Introduction
Atherosclerosis, a major cause of sudden cardiac
death, acute myocardial infarction, and unstable angina pectoris
(1), is a chronic arterial disease
characterized by lipid deposition and inflammation in the vessel
wall (2). Macrophage-derived foam
cells play a key role in the development of atherosclerosis. Foam
cell formation is primarily caused by impaired cholesterol efflux
or uncontrolled uptake of oxidized low-density lipoprotein (ox-LDL)
in macrophages (3). There are
several classes of scavenger receptors (SRs) on macrophage
membrane, in which class A (SR-A) and B (SR-B) attract more
attention (4). For instance, SR-A
and the cluster of differentiation 36 (CD36) are predominantly
responsible for the uptake of ox-LDL (5). In contrast, efflux of intracellular
lipid occurs primarily through reverse cholesterol transporters
(RCTs) including ATP-binding cassette transporter A1 (ABCA1),
ABCG1, and SR-B type I (SR-BI) (6,7).
Hence, foam cell formation is mainly mediated by these RCTs and
SRs. Ample evidence has indicated that regulation of these SRs or
RCTs by antioxidants inhibits the lipid accumulation in foam cells,
retarding the progress of atherosclerosis (3,8).
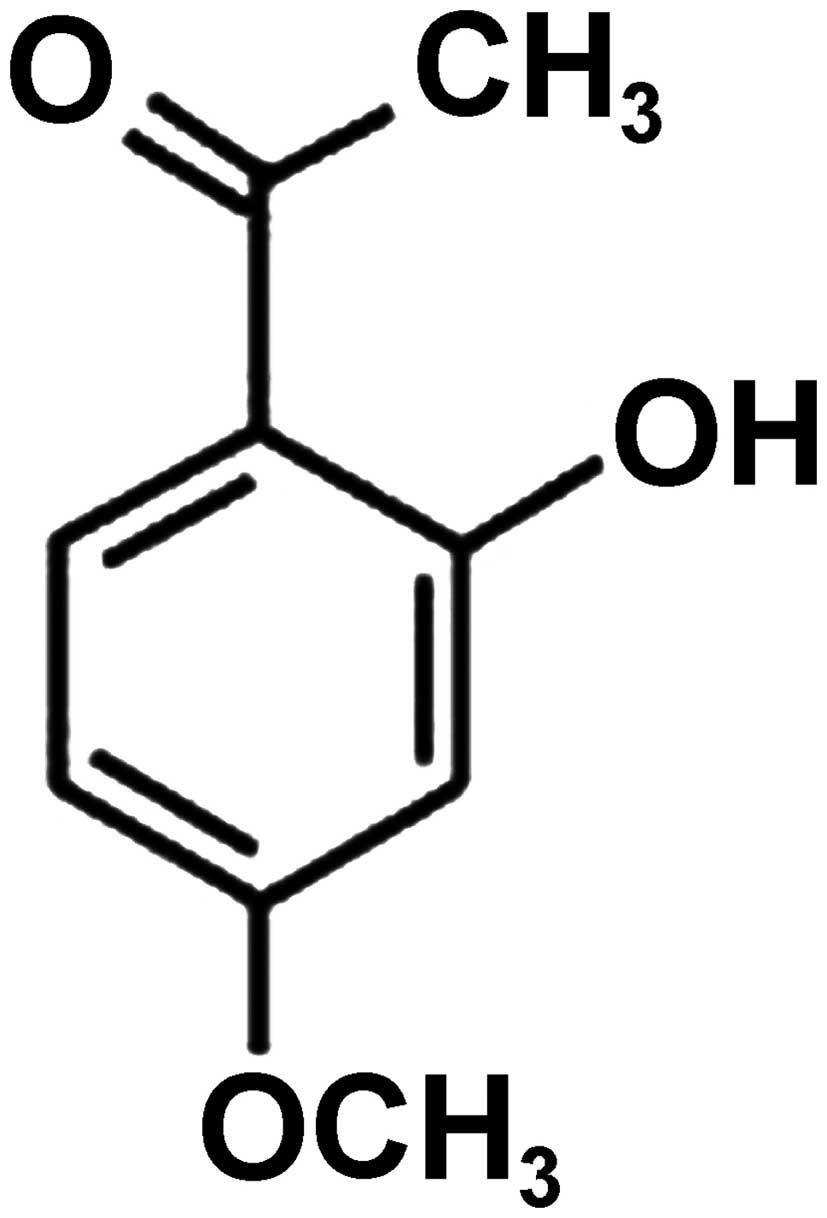
Paeonol (Fig. 1),
an active compound isolated from cortex moutan, possesses
anti-proliferative properties and apoptosis-inducing activity in
different cancer cell types (9–12).
Besides its anticancer effect, paeonol has been reported to display
several beneficial effects in cardiovascular system including
vascular dilation (13), reduction
of platelet aggregation (14) and
improvement of ischemia reperfusion injury in animals (15). Moreover, paeonol is effective in
treatment of atherosclerosis by inhibiting inflammation (16), decreasing thromboxane A2 and free
radical formation (17,18), protecting vascular endothelial
cells (19), and regulating lipid
metabolism (20). Nevertheless,
studies on the effects and molecular mechanism by which paeonol
mediates lipid accumulation in macrophage-derived foam cells are
not well documented. Additionally, various genetic population
studies emphasized the importance of haem oxygenase-1 (HO-1)
expression in the protection against human atherosclerotic lesions
(21). However, whether HO-1 is
involved in the anti-atherogenic effect of paeonol on foam cell
formation remains to be investigated.
In the present study, we investigated the effects of
paeonol on atherosclerosis and the underlying mechanisms in
RAW264.7 cells (mouse macrophage cell line) and apolipoprotein
E-deficient (ApoE−/−) mice. We found that paeonol not
only significantly reduced the formation of foam cells in
vitro, but also inhibited atherosclerotic plaque area in aortas
from ApoE−/− mice. The anti-atherosclerotic effect of
paeonol may be attributed to the upregulation of ABCA1 and
downregulation of CD36 via promotion of HO-1 expression.
Materials and methods
Reagents
Paeonol (purity, 98.0%) and cycloheximide (CHX) were
purchased from Sigma (St. Louis, MO, USA). Goat anti-SR-A antibody
was from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Mouse anti-ABCA1 as well as anti-calpain, anti-calpastatin,
anti-ABCG1, anti-CD36 and anti-SR-BI rabbit antibodies were from
Abcam (Cambridge, MA, USA). Anti-HO-1, anti-c-Jun, anti-c-fos
rabbit antibodies were purchased from Cell Signaling Technology,
Inc. (Beverly, MA, USA). Assay kit for calpain activity was
obtained from BioVision (Lyon, France). Scrambled and HO-1 small
hairpin RNAs (shRNAs) were purchased from Shanghai GenePharma Co.,
Ltd. (Shanghai, China). ox-LDL was supplied by Guangzhou Yiyuan
Biotechnology Co., Ltd. (Guangzhou, China). 3-Dodecanoyl-NBD
cholesterol was obtained from Cayman Chemical Co. (Ann Arbor, MI,
USA).
Cell culture and transfection
RAW264.7 was obtained from the American Type Culture
Collection (ATCC) (Manassas, VA, USA), and maintained in RPMI-1640
medium (Invitrogen Life Technologies, Carlsbad, CA, USA) with 10%
fetal bovine serum in a 37°C incubator with 5% CO2. An
shRNA-containing plasmid that targeted HO-1 was designed and
constructed by Shanghai GeneChem Co., Ltd. (Shanghai, China). The
gene sequence of the shRNA was 5′-GCTGACAGAGGAACACAAAGA-3′. Cell
transfections were performed with the SuperFect fragment (Qiagen,
Valencia, CA, USA) according to the manufacturer’s instructions
using scrambled or HO-1 shRNA in a 50-ml flask. Cells were
incubated for 24 h after transfection and used for the indicated
experiments (8).
Animals
Eight-week-old male ApoE−/− mice and
C57BL/6J mice from Jackson Laboratory (Bar Harbor, ME, USA), which
were a generous gift from Dr Xiong-Zhong Ruan (Key Laboratory of
Lipid and Glucose Metabolism, Chongqing, China), were housed in
barrier facilities on a 12-h light / dark cycle. All experimental
mice were allowed access to food and water ad libitum.
Animal procedures were approved by the Animal Care and Use
Committee of the First Affiliated Hospital of Chongqing Medical
University (Chongqing, China) (Item no. 201303).
Animal experimental protocols
After being fed a high-fat diet (15.8% fat and 1.25%
cholesterol) for 8 weeks, ApoE−/− mice were treated
orally for 8 weeks with paeonol (150 mg/kg/day), simvastatin (5
mg/kg/day), or vehicle (20 ml/kg/day, 0.5% sodium carboxyl methyl
cellulose) by gastric gavages (n=10, each group). The doses of
paeonol and simvastatin used in this study is according to previous
reports (22,23). C57BL/6J mice on a common diet as a
control group (n=10) were treated with the same volume of vehicle
only over the same treatment period. Eight weeks after treatment
with paeonol or simvastatin (total diet-fed period was 16 weeks),
mice were euthanized with CO2, hearts and aortas were
collected for Oil Red O staining and western blotting.
Atherosclerotic lesions at the level of the aortic
valve were detected by Oil Red O staining (24) and atherosclerotic plaque area in
the entire aorta was determined as described before (25).
Cell viability assay with MTT
Macrophages were seeded at density of
7.5×104 cells/well in 96-well plates and the cell
viability was determined by methylthiazolyl tetrazolium (MTT)
assay. After the treatment with or without paeonol for 24 h, the
culture supernatant was removed. The following procedures were
performed as previously described (8).
Assessment of foam cell formation by Oil
Red O staining
After washing 3× with PBS, cells were fixed in 4%
paraformaldehyde for 20 min and then stained with 0.5% Oil Red O
staining for 10 min to visualize cellular lipid accumulation.
Hematoxylin was used as counter staining. The stained cells were
photographed by light microscopy with ×400 magnification. The
density of lipid content was detected by alcohol extraction after
Oil Red O staining. The absorbance at 540 nm was determined with a
microplate reader.
Cholesterol efflux assay
We have previously applied NBD cholesterol to study
the experiments of cholesterol efflux in macrophages (8). Macrophages were incubated with
different concentration of paeonol for 24 h, followed by a further
6-h treatment of the equilibration of NBD cholesterol (1 μg/ml) in
the presence of paeonol. NBD cholesterol-labeled cells were washed
with phosphate-buffered saline and incubated in RPMI-1640 medium
for 6 h. The fluorescence-labeled cholesterol released from the
cells into the medium was measured by use of a multilabel counter
(PerkinElmer, Waltham, MA, USA) with 485 nm excitation and 535 nm
emission.
Western blotting
Tissues or cells were harvested and protein extracts
prepared as previously described (26). They were then subjected to western
blotting using primary antibodies. The proteins were visualized and
quantified using a chemiluminescence method (Pierce Biotechnology,
Inc., Rockford, IL, USA) and Quantity One (Bio-Rad, Hercules, CA,
USA) software program.
Immunoprecipitation
Cell lysates containing equal amounts of protein
(1,000 mg) from macrophages treated with or without paeonol for 24
h were incubated with specific primary antibody overnight at 4°C,
and then with protein A/G-Sepharose for 2 h. Immune complexes were
collected and eluted in lysis buffer. Eluted protein samples were
then boiled in SDS-PAGE loading buffer for subsequent western
blotting.
Quantitative real-time polymerase chain
reaction (RT-qPCR)
Total RNA was isolated by TRIzol reagent (Invitrogen
Life Technologies). cDNA synthesis was performed using MuLV Reverse
Transcriptase (Applied Biosystems, Foster City, CA, USA). Real-time
PCR was performed using a SYBR-Green PCR Master Mix kit [Tiangen
Biotech (Beijing) Co., Ltd., Beijing, China]. Primer sequences were
as follows: SR-A forward, 5′-TGGTCCACCTGGTGCTCC-3′ and reverse,
5′-ACCTCCAGGGAAGCCAATTT-3′; CD36 forward,
5′-CAGTTGGAGACCTGCTTATCC-3′ and reverse,
5′-GCGTCCTGGGTTACATTTTC-3′; β-actin forward,
5′-TTGTCCCTGTATGCCTCTGG-3′ and reverse,
5′-GAGGTCTTTACGGATGTCAACG-3′; ABCA1 forward,
5′-GGTTTGGAGATGGTTATACAATAGTTGT-3′ and reverse,
5′-CCCGGAAACGCAAGTCC-3′; ABCG1 forward, 5′-TTCCCCTGGAGATGAGTGTC-3′
and reverse, 5′-CAGTAGGCCACAGGGAACAT-3′; SR-BI forward,
5′-ACCCTAACCCAAAGGAGCAT-3′ and reverse, 5′-CACAGCAACGGCAGAACTAC-3′.
The reaction of RT-qPCR was performed under the following
conditions: 3 min at 95°C for 1 cycle, 10 sec at 95°C, 30 sec at
60°C for 39 cycles, 95°C for 5 sec.
Measurement of calpain activity
Calpain activity was measured as previously
described (3). Briefly, cellular
lysates (100 mg) were mixed with reaction buffer and fluorogenic
substrate Ac-LLY-AFC. The level of released AFC was measured >1
h at 37°C by fluorometry using 400 nm excitation and 505 nm
emission filters.
Statistical analysis
Data are presented as mean ± SEM. Statistical
analysis was performed using one-way ANOVA followed by Bonferroni
post-hoc test or unpaired Student’s t-test. Continuous variables
were tested for normal distribution using the Kolmogorov-Smirnov
test. Differences were considered statistically significant when
P<0.05. All calculations were carried out with SPSS 15.0
version.
Results
Paeonol inhibits lipid accumulation and
promotes cholesterol efflux in RAW264.7 macrophages
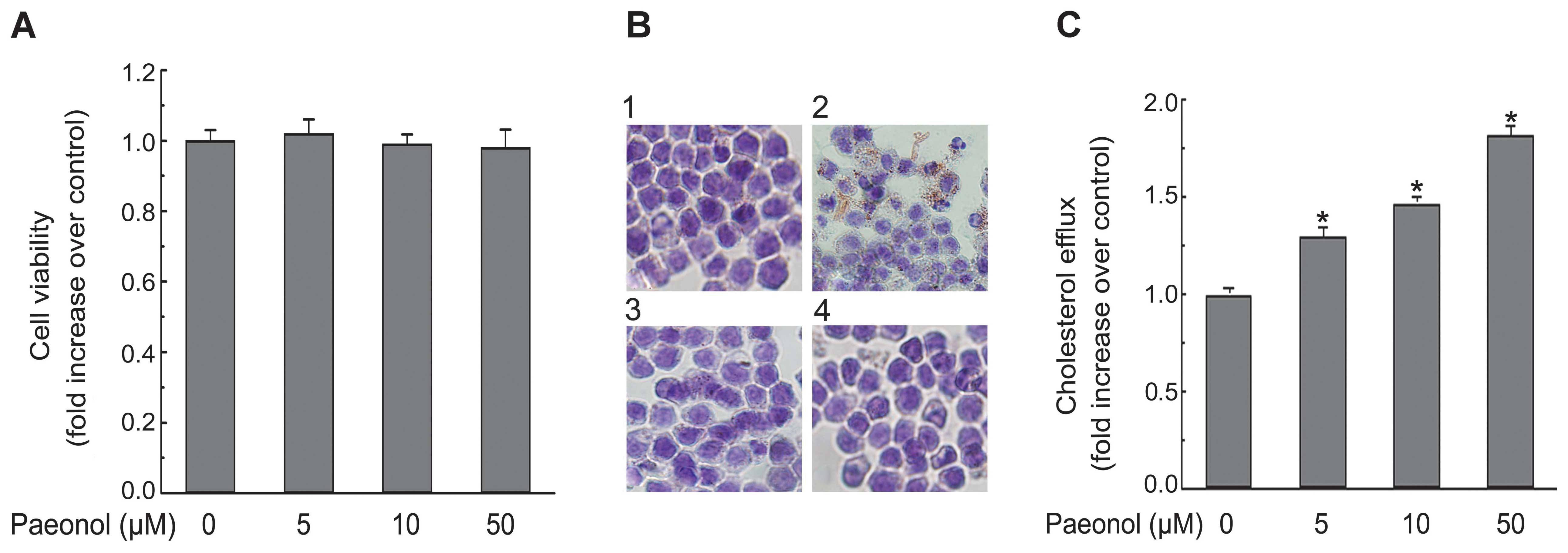
To investigate the toxic effect of paeonol
treatment, cell viability was determined using an MTT assay. As
shown in Fig. 2A, paeonol
treatment for 24 h did not influence cell viability at the tested
concentrations (5, 10, 50 μM). A similar result was reported
previously (27). Therefore, a
concentration range of 5–50 μM was chosen for subsequent
experiments. Lipid accumulation, a hallmark of foam cell formation,
was detected in cells treated with ox-LDL in the presence or
absence of paeonol. Intracellular lipid accumulation in
ox-LDL-treated macrophages was significantly declined when
macrophages were co-incubated with paeonol and ox-LDL as revealed
by Oil Red O staining (Fig. 2B).
The data suggest that paeonol retarded ox-LDL uptake and foam cell
formation in macrophages. Accordingly, paeonol treatment caused a
dose-dependent increase of cholesterol efflux in macrophages as
performed by using NBD-labeled cholesterol (Fig. 2C). This is likely to contribute to
the protective effect by paeonol on macrophage foam cell
formation.
Paeonol decreases the expression of CD36,
but increases the expression of ABCA1 in RAW264.7 macrophages
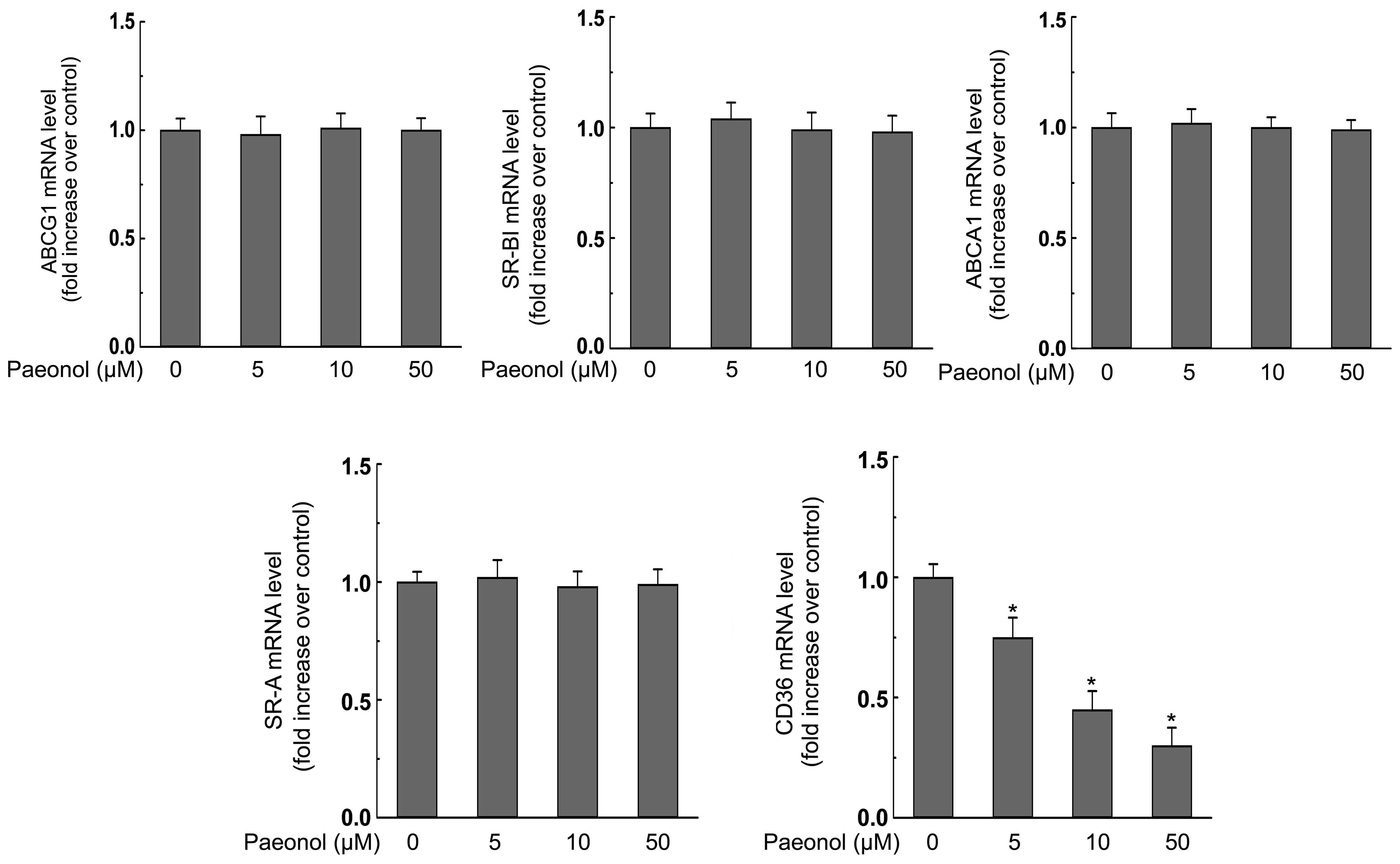
We next explored the effect of paeonol on the
expression of ABCA1, ABCG1, SR-BI, SR-A, CD36, which are reported
to play the most critical roles during the lipid accumulation in
macrophages (3). We found that
paeonol treatment at various concentrations (0, 5, 10, 50 μM) for
24 h dose-dependently decreased both mRNA and protein expression of
CD36 but had no effect on the expression of SR-A (Figs. 3 and 4). Additionally, the mRNA and protein
expression of ABCG1 and SR-BI were not affected by paeonol either
(Figs. 3 and 4). However, ABCA1 protein level in
macrophages was promoted significantly in response to paeonol
treatment (Fig. 3). Importantly,
paeonol did not affect the mRNA expression of ABCA1 (Fig. 4).
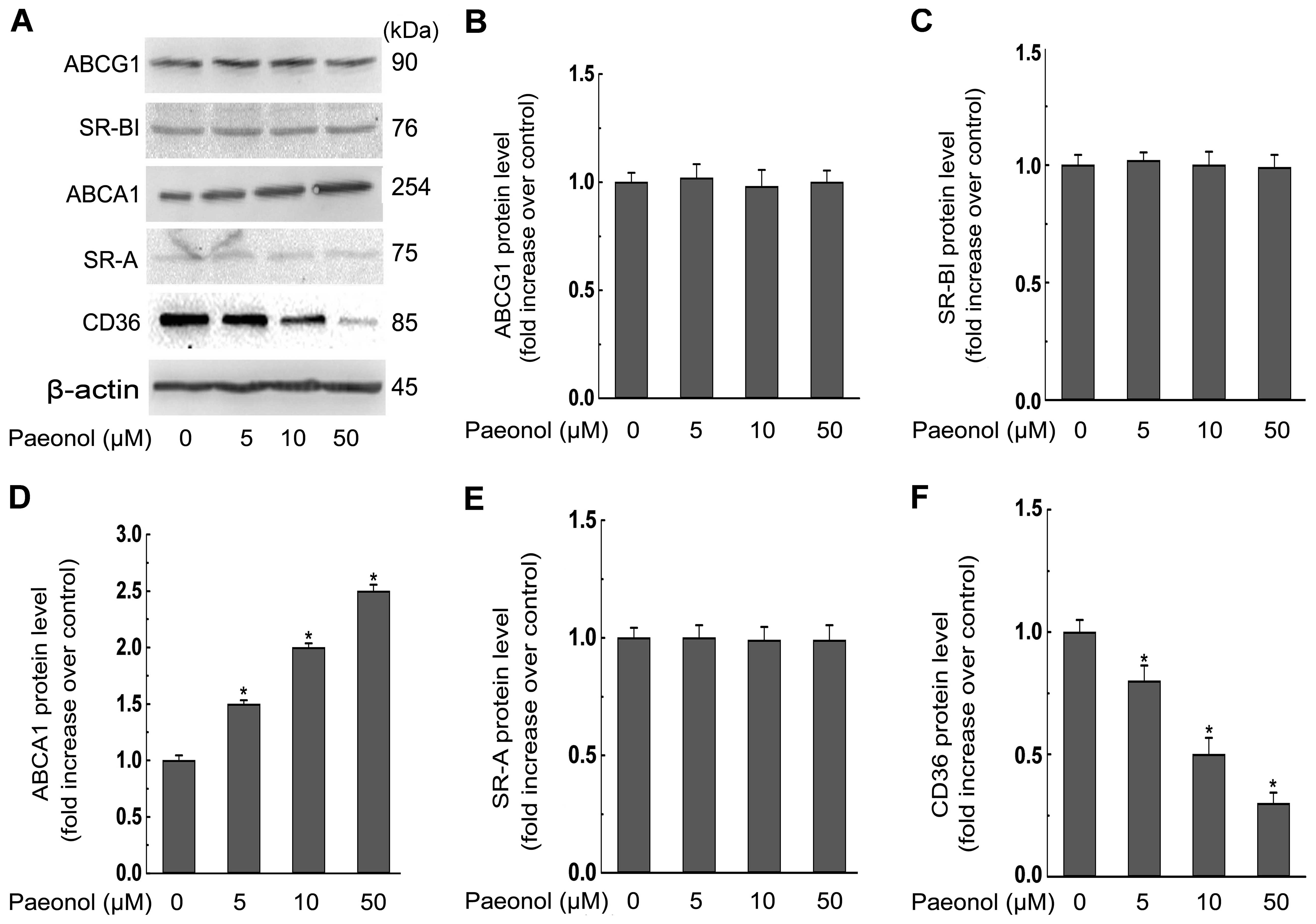 | Figure 3Paeonol decreases protein expression
of the cluster of differentiation 36 (CD36), but increases protein
expression of the ATP-binding cassette transporter A1 (ABCA1) in
RAW264.7 macrophages. (A) Macrophages were treated with indicated
concentrations (0, 5, 10, 50 μM) of paeonol for 24 h and the
protein level of scavenger receptor class A (SR-A), CD36, SR-B type
I (SR-BI), ABCA1, ABCG1, or β-actin was determined by western
blotting. (B–F) The relative protein levels of CD36, SR-A, SR-BI,
ABCA1, ABCG1 are presented as mean ± SEM of optical density from
three separated experiments. *P<0.05 compared with
control. |
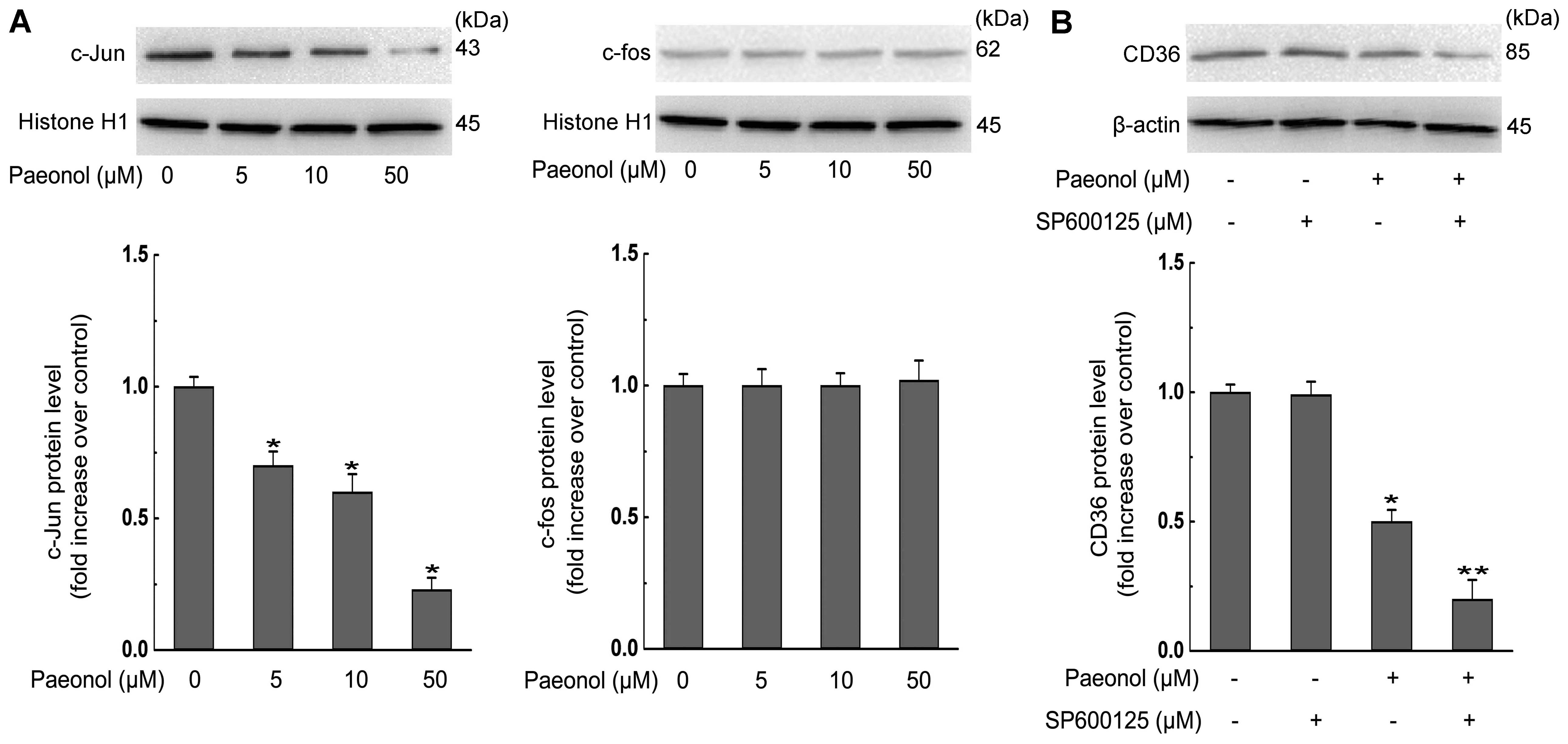
c-Jun-AP-1 pathway is involved in the
inhibition effect of paeonol on CD36 expression in RAW264.7
macrophages
c-Jun and c-fos [the subunits of activator protein-1
(AP-1)] have been reported to participate in CD36 and SR-A
expression in macrophages (3,8).
Therefore, we investigated the role of AP-1 in paeonol-invoked
inhibition of CD36 expression. As shown in Fig. 5A, treatment of macrophages with
paeonol caused a dose-dependent decrease in c-Jun nuclear level
without affecting c-fos nuclear level. A c-Jun N-terminal kinase
(JNK) inhibitor SP600125, which is a potent, cell-permeable
selective and reversible inhibitor of JNK1, 2, and 3, augmented
paeonol-induced decrease in CD36 expression (Fig. 5B). Collectively, these results show
that the suppression of CD36 expression and subsequent alleviation
of foam cell formation by paeonol are partly
c-Jun-AP-1-dependent.
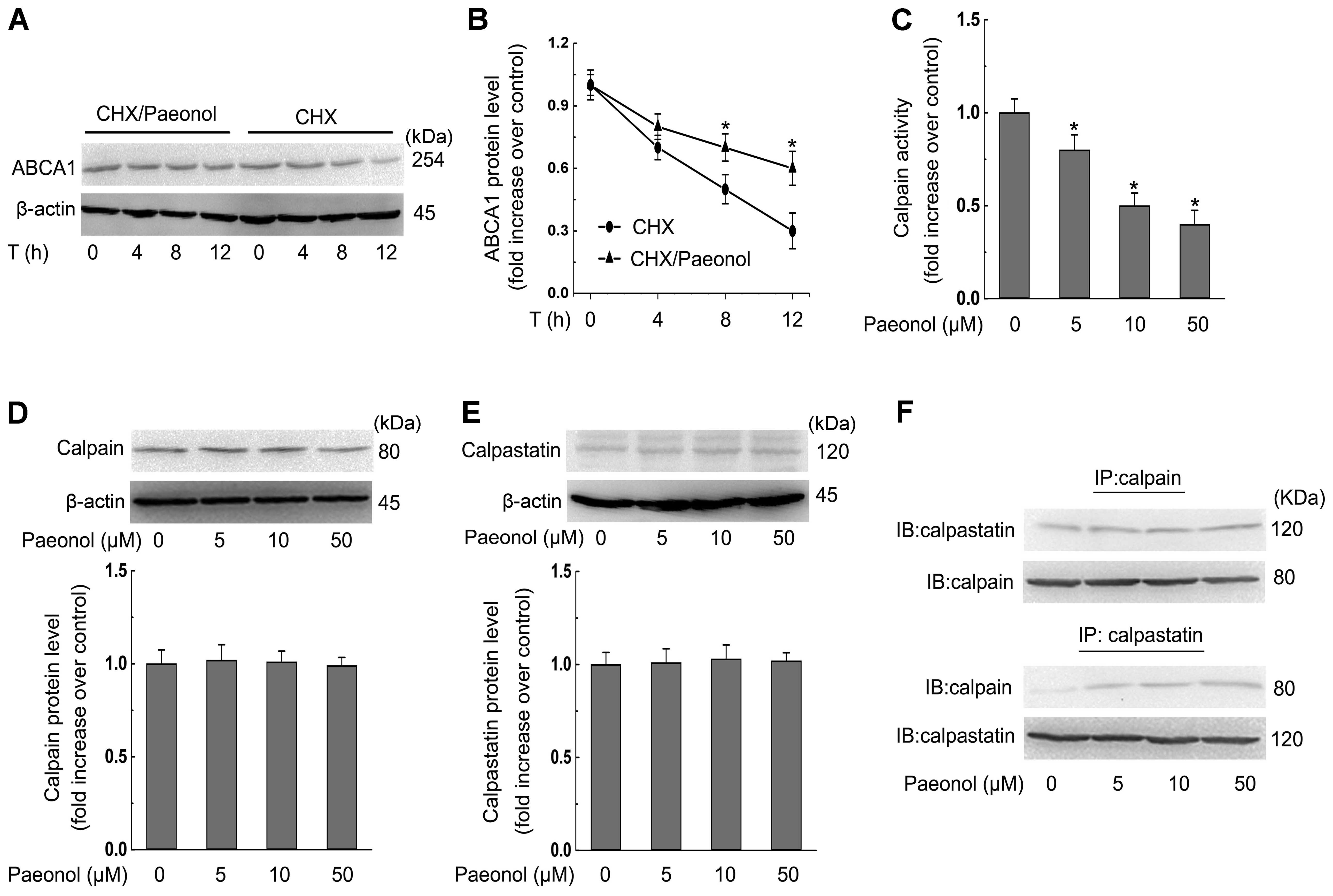
Decreased calpain activity is required
for paeonol-invoked stabilization of ABCA1 protein in RAW264.7
macrophages
We further examined the molecular mechanisms
involved in the effect of paeonol on the protein expression of
ABCA1 by examining the protein stability of ABCA1 with paeonol
treatment. In the presence of CHX (an inhibitor of de novo
protein synthesis), the degradation rate of ABCA1 protein was
dose-dependently inhibited by paeonol treatment (Fig. 6A and B). To define the possible
mechanisms underlying the effect of paeonol on protein stability of
ABCA1, we evaluated the activity of calpain, a protease involved in
ABCA1 proteolysis, after the treatment of paeonol for 24 h in
macrophages. We found that paeonol inhibited calpain activity in a
dose-dependent manner (Fig. 6C).
However, the expression of calpain and calpastatin (the endogenous
inhibitor for calpain) were not altered by paeonol (Fig. 6D and E). Obviously, the declined
calpain activity resulted from an increase in the protein
interaction between calpain and calpastatin (Fig. 6F).
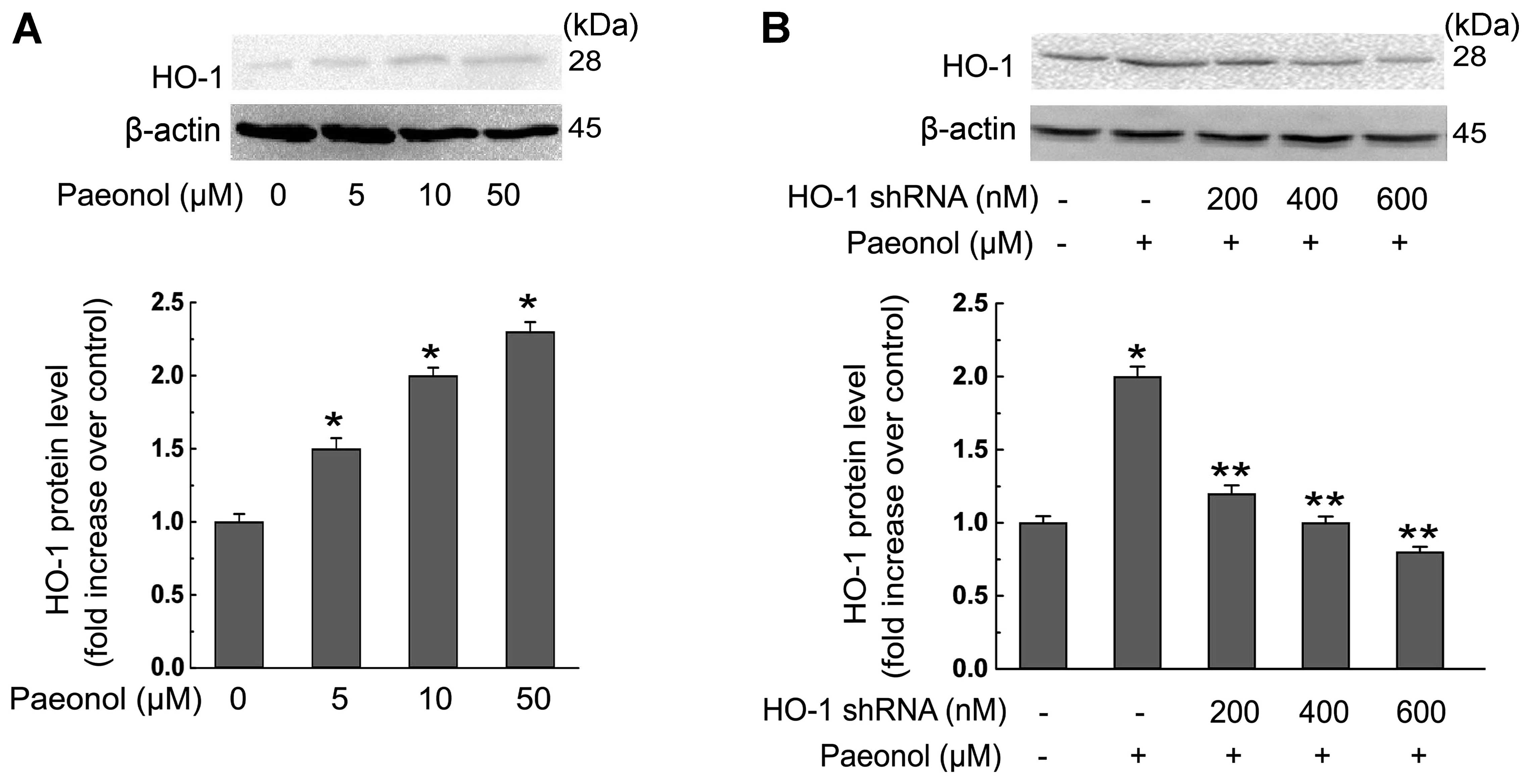
The suppression effect of paeonol on foam
cell formation is regulated by HO-1
We explored the role of HO-1 in paeonol-induced
inhibition of foam cell formation. The protein level of HO-1 in
macrophages was dose-dependently increased in response to paeonol
as shown in Fig. 7A. Moreover,
transfection of the HO-1 shRNA at the concentration of 600 nM
reversed the induction effect of paeonol on HO-1 protein expression
in macrophages (Fig. 7B), whereas
transfection with corresponding scrambled shRNA failed to do so.
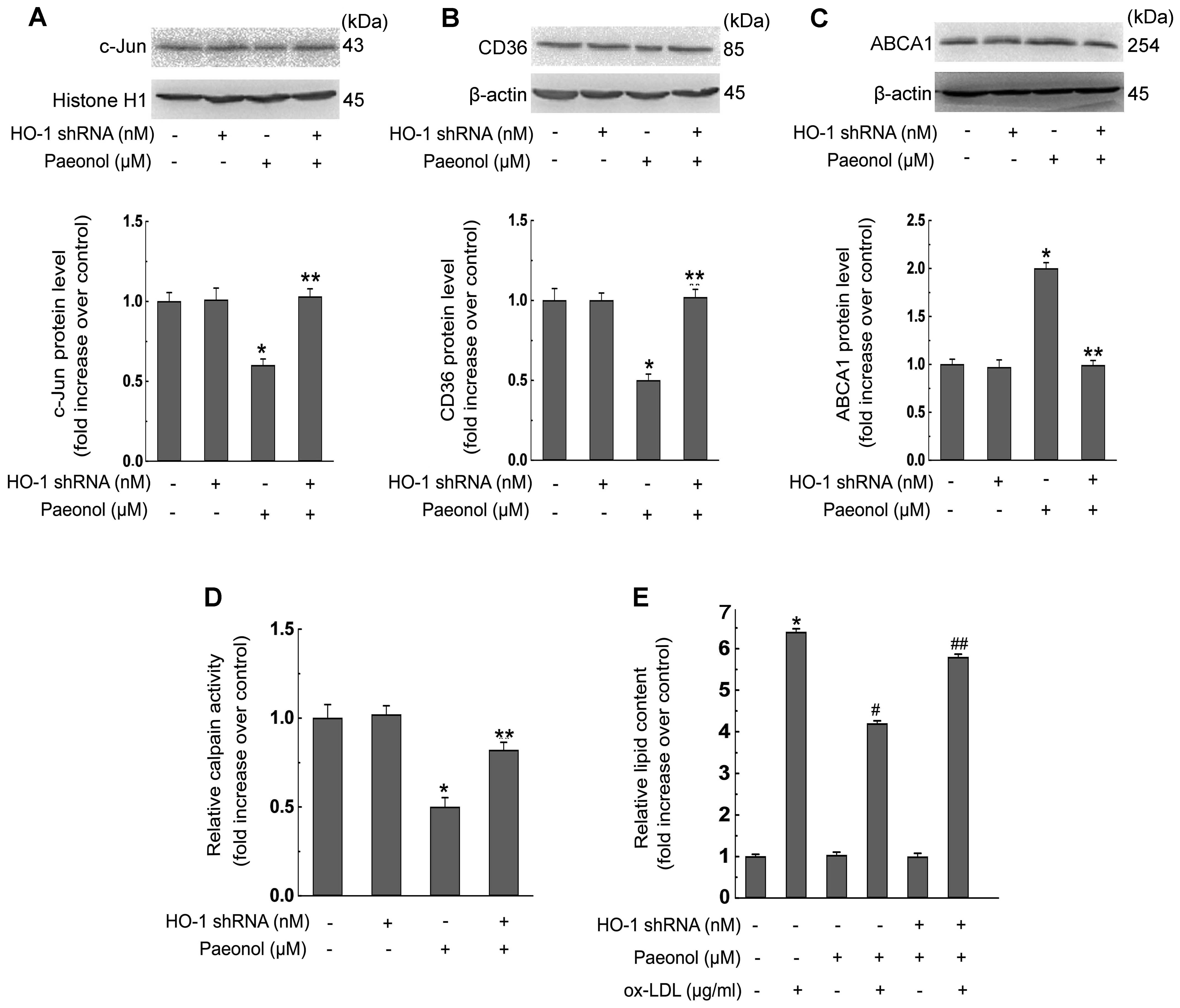
Additionally, HO-1 shRNA transfection decreased paeonol-invoked
effects on the inhibition of c-Jun (Fig. 8A), and CD36 protein expression
(Fig. 8B), promotion of ABCA1
protein expression (Fig. 8C),
attenuation of calpain activity (Fig.
8D), and lipid accumulation (Fig.
8E) in macrophages, indicating the crucial role of HO-1 in
paeonol-mediated protection in foam cells.
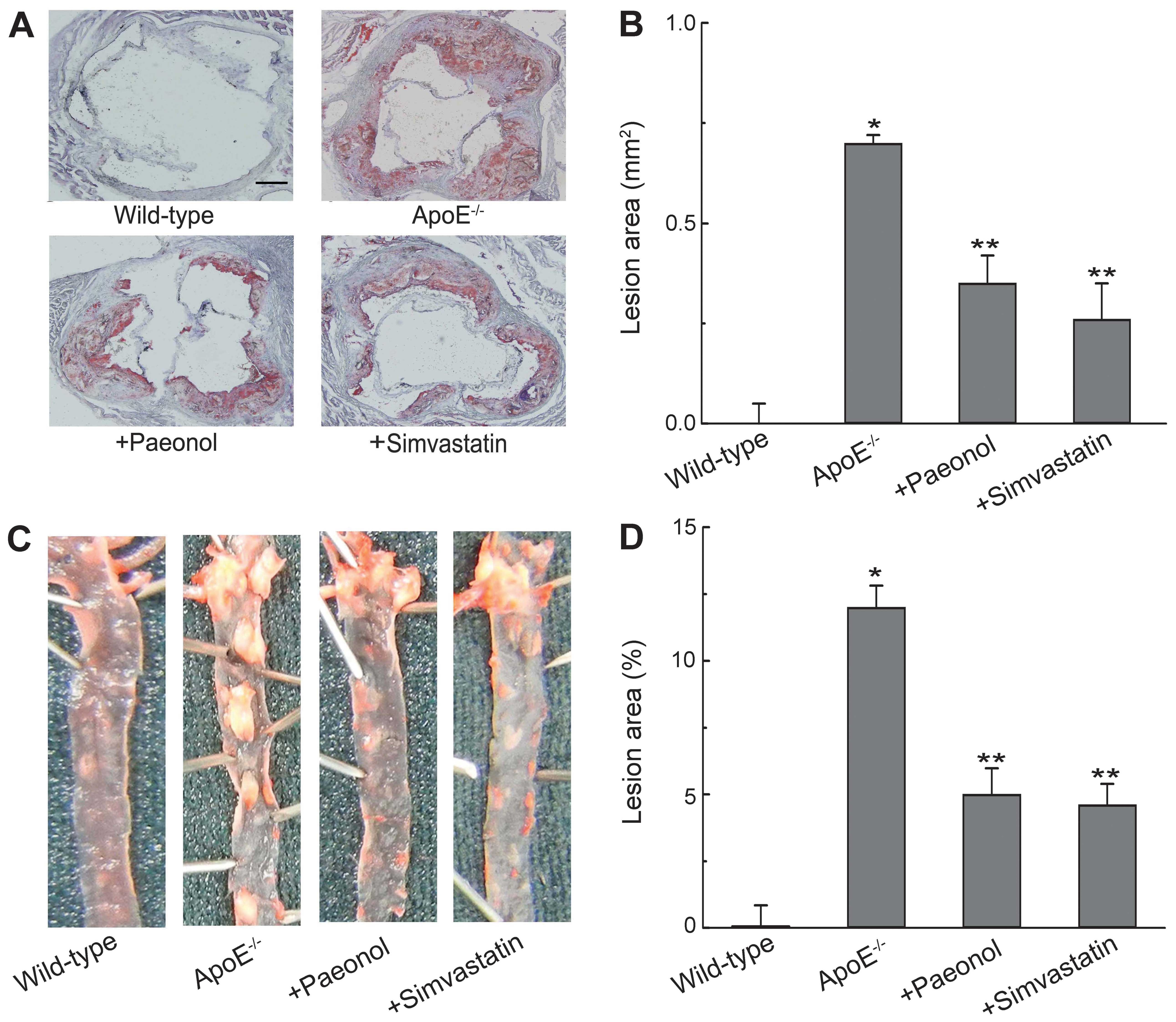
Paeonol decreases atherosclerotic lesion
formation in ApoE−/− mice
We further determined the anti-atherogenic function
of paeonol in vivo. Morphological observations as shown in
Fig. 9A and B, the lesion area in
the aortic root was increased significantly in the
ApoE−/− mice treated with vehicle compared with the
control group. Treatment of 16-week-old ApoE−/− mice
with paeonol (150 mg/kg) for 8 weeks significantly reduced
atherosclerotic lesion formation compared to ApoE−/−
mice treated with vehicle. However, no significant differences were
found in lesion areas between paeonol-treated group and
simvastatin-treated group. Similar results were obtained in
atherosclerotic plaque formation as revealed by Oil Red O staining
of whole aortas (Fig. 9C and D).
Moreover, paeonol treatment increased ABCA1 protein expression in
aortas, while decreased CD36 protein in aortas (Fig. 10). These results are consistent
with the findings of in vitro experiments.
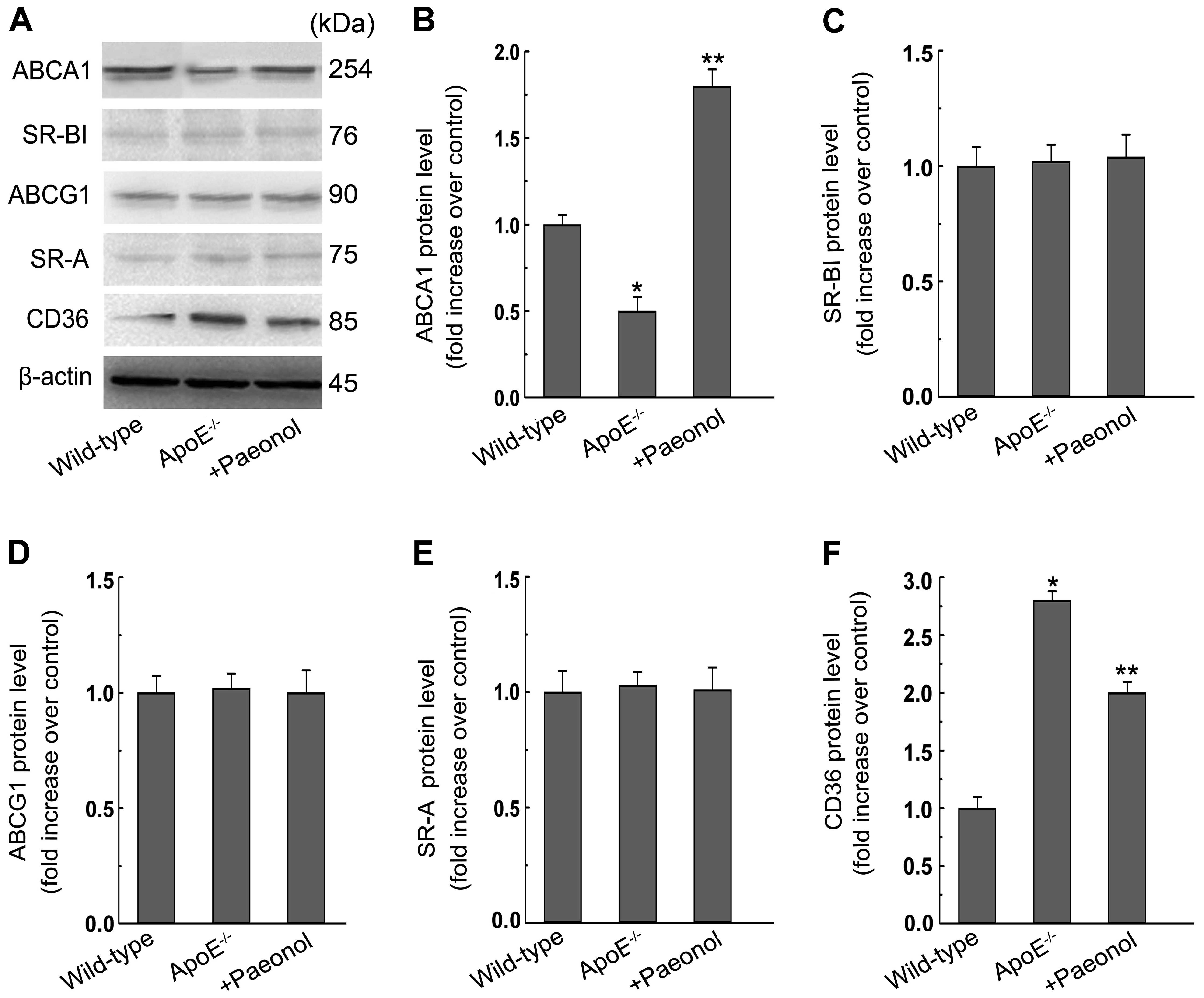 | Figure 10Paeonol promotes ATP-binding cassette
transporter A1 (ABCA1) protein expression, but decreases the
protein expression of the cluster of differentiation 36 (CD36) in
aortas of apolipoprotein E-deficient (ApoE−/−) mice. (A)
After the treatment of wild-type or ApoE−/− mice as
described in the Materials and methods section, aortas were
collected and subjected to western blotting to detect the protein
expression of ABCA1, ABCG1, SR-B type I (SR-BI), scavenger receptor
class A (SR-A), CD36. One representative blot is shown. (B–F) The
relative protein levels of CD36, SR-A, SR-BI, ABCA1, ABCG1 are
presented as mean ± SEM of optical density from three separated
experiments. *P<0.05 compared with wild-type,
**P<0.05 compared with ApoE−/−. Wild-type,
C57BL/6J mice; paeonol, paeonol at 150 mg/kg/day. |
Discussion
It is well established that paeonol confers
protection against atherosclerosis (16–20).
Nevertheless, the effect and molecular mechanism by which paeonol
mediates lipid accumulation in macrophage-derived foam cells
remained to be resolved. We provide new insights into the molecular
mechanisms involved in the anti-atherogenic property of paeonol in
the macrophage-derived foam cell formation and in
ApoE−/− mouse aortas. In RAW264.7 macrophages, paeonol
treatment decreased ox-LDL-induced lipid accumulation by promoting
ABCA1-dependent cholesterol efflux and attenuating CD36-dependent
ox-LDL uptake. The increased ABCA1-regulated cholesterol efflux
resulted from increased protein stability of ABCA1. Our results
strongly indicate that paeonol has a beneficial function in
maintenance of lipid level during the transformation of foam cells
in atherosclerosis and we delineated the underlying molecular
mechanism.
SR-dependent ox-LDL internalization and RCT-mediated
cholesterol efflux play a key role in the mediation of
intracellular lipid level of foam cells (28). We may be the first to demonstrate
that attenuation of intracellular lipid accumulation by paeonol in
macrophage foam cells is possibly via decreased ox-LDL uptake and
increased cholesterol efflux. In addition, paeonol significantly
decreased both the protein and mRNA expression of CD36 without
affecting SR-A expression. SR-A and CD36 are two major types of SRs
involved in the uptake of ox-LDL in macrophages (29). Genetic inactivation of CD36 reduced
ox-LDL uptake in vitro and atherosclerotic lesions in mice
(30). Extensive research also
indicates that CD36 expression can be decreased by anti-atherogenic
antioxidants, which reveals its key role in the pathogenesis of
atherogenesis (31,32). In the present study, we demonstrate
that the suppressive effect of paeonol on ox-LDL uptaking was via
downregulation of CD36 expression, which could be caused by
decreased nuclear translocation of c-Jun (a subunit of AP-1).
Indeed, pharmacological inhibition of AP-1 pathway augmented the
effect of paeonol on CD36 expression. Our findings show that
inactivation of the c-Jun-AP-1 pathway is required for the
athero-protective action of paeonol in macrophages. In view of the
function of CD36, the attenuation of CD36 expression by paeonol may
contribute to the reduction of ox-LDL internalization and
subsequent inhibition of foam cell formation.
In addition to its inhibitory effect on CD36
expression, paeonol promoted cholesterol efflux by increasing ABCA1
protein expression without altering ABCA1 mRNA level. The increase
of ABCA1 protein expression induced by paeonol likely results from
increased stability of ABCA1 protein. ABCA1 is the most important
RCT responsible for cholesterol efflux from macrophage foam cells
(28). Defects in ABCA1-mediated
reverse cholesterol transport result in increased intracellular
cholesterol accumulation, and ABCA1 plays a critical role in
protecting against atherosclerosis in both humans and animals
(33). Moreover, the expression of
ABCA1 is known to be upregulated by various antioxidants with
anti-atherogenic properties (34,35).
In view of ABCA1 function, the increased expression of ABCA1 by
paeonol observed in this study is also likely to facilitate the
cholesterol efflux and subsequent suppression of foam cell
formation. Importantly, our findings indicate that the induced
expression of ABCA1 by paeonol is accompanied by decreased calpain
activity. We further show that the reduced calpain activity may be
due to the increased protein interaction between calpain and
calpastatin as revealed by immunoprecipitation. The critical role
of calpain in the stabilization of ABCA1 protein is well
established (36,37). Furthermore, wogonin increases the
protein stability of ABCA1 via PP2B-mediated dephosphorylation
(38). Whether PP2B-mediated
dephosphorylation is involved in paeonol-invoked increase in ABCA1
protein stability remains to be further examined.
Ample studies reported that HO-1 is correlated with
the suppression of atherosclerotic damage (39,40).
Indeed, HO-1 overexpression by pharmacological inducers or viral
gene transfer inhibits atherogenesis in hypercholesterolemic animal
models (39,41). On the other hand, our previous
study shows that gene knockdown of HO-1 reversed kaempferol-invoked
protection against foam cell formation (8). However, whether HO-1 is involved in
the athero-protective action of paeonol remains unknown. In this
study, we found that paeonol profoundly elicited HO-1 protein
expression in macrophages. Additionally, our findings indicated
that gene knockdown of HO-1 prevented paeonol effects on the
inhibition of c-Jun/AP-1 nuclear translocation, increase of ABCA1
expression, decrease of CD36 expression, attenuation of calpain
activity, and lipid accumulation in macrophages. Collectively,
these data strongly suggest the essential role of HO-1 in the
paeonol-mediated suppression of foam cell formation. Moreover, the
metabolites of HO-1, biliverdin and carbon monoxide, are known to
provide protection in cardiovascular diseases (42). Nevertheless, whether these
metabolites are involved in the anti-atherogenic effect of paeonol
on the formation of foam cells remains to be examined in the
future.
Our findings of paeonol-invoked inhibition of foam
cell formation are not limited to the cell culture system. Our
in vivo experiments indicated that the positive drug
simvastatin decreases atherosclerotic lesion formation, which is
consistent with a previous study (22). This result suggests that the
atherosclerotic model was established successfully. Moreover,
paeonol retarded atherosclerotic progression in ApoE−/−
mice. In fact, studies have reported that paeonol displayed several
beneficial effects on atherosclerosis such as inhibition of
monocyte adhesion to vascular endothelial cells (43), inhibiting inflammatory response
(16) and downregulating platelet
aggregation (15). Here, we
further found increased protein levels of ABCA1 and decreased
protein expression of CD36 in paeonol-treated apoE−/−
mice. Hence, these therapeutic effects seem to be widespread, but
the target cells for paeonol cannot be identified under this
hyperlipidemic situation. Accordingly, paeonol may have beneficial
effect in various organs via dissimilar mechanisms.
In conclusion, our data show that HO-1 plays a
crucial role in the anti-atherogenic effect of paeonol on the
formation of foam cells, which reduces intracellular lipid
accumulation in foam cells via downregulating CD36 and
post-transcriptionally upregulating ABCA1 expression. We provide
new insights for better understanding the molecular mechanisms
involved in paeonol-invoked inhibition of foam cells formation in
atherosclerosis. Paeonol may exert anti-atherosclerotic effect by
reducing serum lipid. Whether paeonol could inhibit serum lipid and
the involved mechanisms in apoE−/− mice need to be
investigated in the future.
Acknowledgements
We thank Professor Chao Yu from Chongqing Medical
University (Chongqing, China) for providing experimental platform
during the course of this study. This investigation was supported
by a grant from Public Health Bureau of Chongqing, China, grant no.
2009-1-6.
References
|
1
|
Sung HJ, Kim J, Kim Y, Jang SW and Ko J:
N-acetyl cysteine suppresses the foam cell formation that is
induced by oxidized low density lipoprotein via regulation of gene
expression. Mol Biol Rep. 39:3001–3007. 2012. View Article : Google Scholar
|
|
2
|
Cai Y, Li JD and Yan C: Vinpocetine
attenuates lipid accumulation and atherosclerosis formation.
Biochem Biophys Res Commun. 434:439–443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsai JY, Su KH, Shyue SK, et al: EGb761
ameliorates the formation of foam cells by regulating the
expression of SR-A and ABCA1: role of haem oxygenase-1. Cardiovasc
Res. 88:415–423. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cai Y, Wan Z, Sun T, et al:
Diarylquinoline compounds induce autophagy-associated cell death by
inhibiting the Akt pathway and increasing reactive oxygen species
in human nasopharyngeal carcinoma cells. Oncol Rep. 29:983–992.
2013.PubMed/NCBI
|
|
5
|
Lin CY, Lee TS, Chen CC, et al:
Endothelin-1 exacerbates lipid accumulation by increasing the
protein degradation of the ATP-binding cassette transporter G1 in
macrophages. J Cell Physiol. 226:2198–2205. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Saeed O, Otsuka F, Polavarapu R, et al:
Pharmacological suppression of hepcidin increases macrophage
cholesterol efflux and reduces foam cell formation and
atherosclerosis. Arterioscler Thromb Vasc Biol. 32:299–307. 2012.
View Article : Google Scholar :
|
|
7
|
Zhou F, Pan Y, Huang Z, et al: Visfatin
induces cholesterol accumulation in macrophages through
up-regulation of scavenger receptor-A and CD36. Cell Stress
Chaperones. 18:643–652. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li XY, Kong LX, Li J, He HX and Zhou YD:
Kaempferol suppresses lipid accumulation in macrophages through the
downregulation of cluster of differentiation 36 and the
upregulation of scavenger receptor class B type I and ATP-binding
cassette transporters A1 and G1. Int J Mol Med. 31:331–338.
2013.
|
|
9
|
Chen B, Ning M and Yang G: Effect of
paeonol on antioxidant and immune regulatory activity in
hepatocellular carcinoma rats. Molecules. 17:4672–4683. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li M, Tan SY, Zhang J and You HX: Effects
of paeonol on intracellular calcium concentration and expression of
RUNX3 in LoVo human colon cancer cells. Mol Med Rep. 7:1425–1430.
2013.PubMed/NCBI
|
|
11
|
Li N, Fan LL, Sun GP, et al: Paeonol
inhibits tumor growth in gastric cancer in vitro and in vivo. World
J Gastroenterol. 16:4483–4490. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun GP, Wan X, Xu SP, Wang H, Liu SH and
Wang ZG: Antiproliferation and apoptosis induction of paeonol in
human esophageal cancer cell lines. Dis Esophagus. 21:723–729.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li YJ, Bao JX, Xu JW, Murad F and Bian K:
Vascular dilation by paeonol - a mechanism study. Vascul Pharmacol.
53:169–176. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hsieh CL, Cheng CY, Tsai TH, et al:
Paeonol reduced cerebral infarction involving the superoxide anion
and microglia activation in ischemia-reperfusion injured rats. J
Ethnopharmacol. 106:208–215. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nizamutdinova IT, Jin YC, Kim JS, et al:
Paeonol and paeoniflorin, the main active principles of Paeonia
albiflora, protect the heart from myocardial ischemia/reperfusion
injury in rats. Planta Med. 74:14–18. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li H, Dai M and Jia W: Paeonol attenuates
high-fat-diet-induced atherosclerosis in rabbits by
anti-inflammatory activity. Planta Med. 75:7–11. 2009. View Article : Google Scholar
|
|
17
|
Dai M, Zhi X, Peng D and Liu Q: Inhibitory
effect of paeonol on experimental atherosclerosis in quails.
Zhongguo Zhong Yao Za Zhi. 24:488–490. 5121999.(In Chinese).
|
|
18
|
Shi L, Fan PS, Fang JX and Han ZX:
Inhibitory effects of paeonol on experimental atherosclerosis and
platelet aggregation of rabbit. Zhongguo Yao Li Xue Bao. 9:555–558.
1988.(In Chinese). PubMed/NCBI
|
|
19
|
Hansson GK: Inflammation, atherosclerosis,
and coronary artery disease. N Engl J Med. 352:1685–1695. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hirai A, Terano T, Hamazaki T, et al:
Studies on the mechanism of antiaggregatory effect of Moutan
Cortex. Thromb Res. 31:29–40. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Araujo JA, Zhang M and Yin F: Heme
oxygenase-1, oxidation, inflammation, and atherosclerosis. Front
Pharmacol. 3:1192012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Song G, Liu J, Zhao Z, et al: Simvastatin
reduces atherogenesis and promotes the expression of hepatic genes
associated with reverse cholesterol transport in apoE-knockout mice
fed high-fat diet. Lipids Health Dis. 10:82011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hu S, Shen G, Zhao W, Wang F, Jiang X and
Huang D: Paeonol, the main active principles of Paeonia moutan,
ameliorates alcoholic steatohepatitis in mice. J Ethnopharmacol.
128:100–106. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rudolph TK, Rudolph V, Edreira MM, et al:
Nitro-fatty acids reduce atherosclerosis in apolipoprotein
E-deficient mice. Arterioscler Thromb Vasc Biol. 30:938–945. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagy N, Melchior-Becker A and Fischer JW:
Long-term treatment with the AT1-receptor antagonist telmisartan
inhibits biglycan accumulation in murine atherosclerosis. Basic Res
Cardiol. 105:29–38. 2010. View Article : Google Scholar
|
|
26
|
Ayaori M, Sawada S, Yonemura A, et al:
Glucocorticoid receptor regulates ATP-binding cassette
transporter-A1 expression and apolipoprotein-mediated cholesterol
efflux from macrophages. Arterioscler Thromb Vasc Biol. 26:163–168.
2006. View Article : Google Scholar
|
|
27
|
Huang H, Chang EJ, Lee Y, Kim JS, Kang SS
and Kim HH: A genome-wide microarray analysis reveals
anti-inflammatory target genes of paeonol in macrophages. Inflamm
Res. 57:189–198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao JF, Ching LC, Huang YC, et al:
Molecular mechanism of curcumin on the suppression of cholesterol
accumulation in macrophage foam cells and atherosclerosis. Mol Nutr
Food Res. 56:691–701. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Witztum JL: You are right too! J Clin
Invest. 115:2072–2075. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kunjathoor VV, Febbraio M, Podrez EA, et
al: Scavenger receptors class A-I/II and CD36 are the principal
receptors responsible for the uptake of modified low density
lipoprotein leading to lipid loading in macrophages. J Biol Chem.
277:49982–49988. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fuhrman B, Volkova N and Aviram M:
Oxidative stress increases the expression of the CD36 scavenger
receptor and the cellular uptake of oxidized low-density
lipoprotein in macrophages from atherosclerotic mice: protective
role of antioxidants and of paraoxonase. Atherosclerosis.
161:307–316. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Venugopal SK, Devaraj S and Jialal I:
RRR-alpha-tocopherol decreases the expression of the major
scavenger receptor, CD36, in human macrophages via inhibition of
tyrosine kinase (Tyk2). Atherosclerosis. 175:213–220. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tang C and Oram JF: The cell cholesterol
exporter ABCA1 as a protector from cardiovascular disease and
diabetes. Biochim Biophys Acta. 1791:563–572. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chang YC, Lee TS and Chiang AN: Quercetin
enhances ABCA1 expression and cholesterol efflux through a
p38-dependent pathway in macrophages. J Lipid Res. 53:1840–1850.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao J, Xu Y, Yang Y, et al: Identification
of upregulators of human ATP-binding cassette transporter A1 via
high-throughput screening of a synthetic and natural compound
library. J Biomol Screen. 13:648–656. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Arakawa R and Yokoyama S: Helical
apolipoproteins stabilize ATP-binding cassette transporter A1 by
protecting it from thiol protease-mediated degradation. J Biol
Chem. 277:22426–22429. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Martinez LO, Agerholm-Larsen B, Wang N,
Chen W and Tall AR: Phosphorylation of a pest sequence in ABCA1
promotes calpain degradation and is reversed by ApoA-I. J Biol
Chem. 278:37368–37374. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen CY, Shyue SK, Ching LC, et al:
Wogonin promotes cholesterol efflux by increasing protein
phosphatase 2B-dependent dephosphorylation at ATP-binding cassette
transporter-A1 in macrophages. J Nutr Biochem. 22:1015–1021. 2011.
View Article : Google Scholar
|
|
39
|
Juan SH, Lee TS, Tseng KW, et al:
Adenovirus-mediated heme oxygenase-1 gene transfer inhibits the
development of atherosclerosis in apolipoprotein E-deficient mice.
Circulation. 104:1519–1525. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yet SF, Layne MD, Liu X, et al: Absence of
heme oxygenase-1 exacerbates atherosclerotic lesion formation and
vascular remodeling. FASEB J. 17:1759–1761. 2003.PubMed/NCBI
|
|
41
|
Ishikawa K, Sugawara D, Goto J, et al:
Heme oxygenase-1 inhibits atherogenesis in Watanabe heritable
hyperlipidemic rabbits. Circulation. 104:1831–1836. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Idriss NK, Blann AD and Lip GY:
Hemoxygenase-1 in cardiovascular disease. J Am Coll Cardiol.
52:971–978. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang YQ, Dai M, Zhong JC and Yin DK:
Paeonol inhibits oxidized low density lipoprotein-induced monocyte
adhesion to vascular endothelial cells by inhibiting the mitogen
activated protein kinase pathway. Biol Pharm Bull. 35:767–772.
2012. View Article : Google Scholar : PubMed/NCBI
|