Introduction
Skeleton is the most common site of metastasis in
breast cancer, which occurs in over 70% of patients suffering this
disease. Bone metastasis, which seriously deteriorates the
patients’ life quality, is life-threatening and leads to several
complications or skeletal-related events, such as pathologic
fracture, pain, disability and nerve compression (1). Although huge efforts have been made
in breast cancer research, what we can do is still limited with
respect to bone metastasis of breast cancer. Safe and effective
treatment to prevent or treat bone metastasis require sufficient
validated targets. Hence, further insights into the molecular
mechanisms underlying the bone metastasis of breast cancer are
urgently needed (2).
The spreading from primary site to distant bones of
breast cancer cells is very complex involving invasion,
intravasation and extravasation of capillary vascular system, and
locating at as well as flourishing in bones. Especially, various
factors that promote cancer cell migration and invasion play a
vital role in the process. It is widely accepted that tumor cells
could travel to distant organs because of some acquired genetic
alterations that facilitate invasion and metastasis (3).
Recent investigations in the field of cancer cell
metastasis rely heavily on the availability of in vivo
animal models. An ideal animal model should mimic the process of
breast cancer cells developing spontaneously metastasis in distant
organs. However, the animal models of bone metastasis we can build
just represent unique stages of human breast cancer bone
metastasis, for this phenomenon occurs rarely in animals (4,5).
miRNA is a class of endogenous small non-coding RNAs
which could modulate gene expression. In almost all cases, these
RNAs negatively regulate gene expression via RNA-RNA binding to the
3′ untranslated region of target mRNAs in a manner of imperfect
complementary, which then leads to either mRNA degradation or
translational inhibition. Accumulating evidence demonstrates that
miRNAs play crucial roles in tumor initiation and progression,
which is exemplified by the ability to regulate every aspect of
tumor pathological process including proliferation, migration,
invasion, differentiation and escape from senescence and apoptosis
(6–9). For example, aberrant expression of
miR-106b promotes hepatocellular carcinoma cell migration in
vitro and metastasis in vivo by stimulating
epithelial-mesenchymal transition (10). miR-145, on the other hand, inhibits
the migration and invasion of metastatic melanoma cells (11).
Several miRNAs have been found to be involved into
breast cancer metastasis. For instance, miR-720 suppresses invasion
and migration in breast cancer cells by targeting TWIST1 (12). miR-29b regulates migration of human
breast cancer cells by modulating PTEN expression (13). In the present study, we established
a bone metastasis mouse model by injecting MDA-MB-231 cells into
the left ventricle and then compared the differentially expressed
miRNAs between the parental malignant cells and the metastatic
cells derived from the bones. The distinct expression pattern of
the miRNAs provides important insight into the molecular mechanism
of bone metastasis of breast cancer. Especially, we focus on the
biological function of miR-429 through its target genes ZEB1 and
CRKL.
Materials and methods
Cell culture and animals
Human mammary carcinoma cell line MDA-MB-231 was
purchased from the American Type Culture Collection (ATCC,
Manassas, VA, USA) and the cells cultured in the Leibovitz L-15
medium supplemented with 10% fetal bovine serum (FBS), streptomycin
(100 mg/ml) and penicillin (100 U/ml). The same culture medium,
adding 200 μg/ml G418 (Life Technologies, Carlsbad, CA, USA) was
also applied to parental MDA-MB-231 (231-P) cells and the
metastatic subline of MDA-MB-231 (231-B) cells derived from the
bones of our animal models, both of which had stable luciferase
expression. All these cells were maintained in the incubator
without gas exchange with outside air.
Five-week-old female Balb/c nude mice were purchased
from Vital River Laboratories (Beijing, China) and bred in the SPF
Animal Center of Cancer Institute and Hospital, Chinese Academy of
Medical Sciences. All the animal experiments were approved by the
Institutional Review Board of the Chinese Academy of Medical
Sciences Cancer Institute.
Generation of lentivirus expression
vehicles
To facilitate in vivo observation and the
following the primary culture of 231-B cells, we constructed a
pLVX-IRES-Neo vector containing luciferase gene and then produced
the lentivirus by transfecting 6.5 μg pLVX-luciferase-Neo vector
and corresponding amounts of package vectors into 293T cells, which
were seeded in 10-cm flasks before the day of the transfection,
following the standard method of Lipofectamine 2000 (Invitrogen).
Forty-eight hours after the transfection, supernatant of these 293T
cells were harvested, centrifuged (3,000 rpm, 4°C), filtered using
0.45-μm filter flasks, aliquoted and stored at −80°C (14).
For transduction, 10 μl supernatant containing
lentivirus was added into the MDA-MB-231 cells, which were
maintained in 2 ml of L-15 complete culture medium. In addition, 8
μg/ml of polybrene (Sigma-Aldrich) was also present in order to aid
transduction. Twenty-four hours later, the medium were replaced by
fresh L15 complemented with 10% FBS. Simultaneously, 400 μg/ml G418
was used to screen out the uninfected cells.
Establishment of bone metastasis
model
The bone metastasis model of breast cancer was
described by Yin and colleagues (15). In general, amounts of 231-P cells
were harvested using 0.25% trypsin and 0.53 mM EDTA, resuspended in
sterilized PBS, and adjusted to a concentration of
5×106/ml. Then, mice were anesthetized by
intraperitoneal injection of 1% (w/v) pentobarbital sodium (90
mg/kg). After confirming that mice were under proper anesthesia, we
injected 100 μl of the suspended cells into the left ventricle via
the third intercostal space with a 29-gauge needle. A successful
injection was confirmed by the pumping of arterial blood into the
syringe (16). Mice were
sterilized with 70% alcohol, and then were bred in the following
weeks.
In vivo detection of bone metastasis
Mice anesthetized by 1% (w/v) pentobarbital sodium
(90 mg/kg) were intraperitoneally injected with 150 mg/kg of
D-Luciferin (Perkin-Elmer) in DPBS. Bioluminescence images were
acquired between 10 and 15 min after injection using Xenogen
Corporation Optical in vivo imaging (IVIS Lumina). Image
acquisition time at the beginning was 60 sec and then it was
reduced in accordance with signal strength to avoid saturation. All
images were analyzed with Living Image software. Intensity of
bioluminescence was calculated as
photons/sec/cm2/steradian of a region of interest (ROI).
Average background reads were obtained from sites of the same mice
without bioluminescence.
Moreover, radiographic analysis was adopted to
further confirm the bone metastases of 231-P cells. Mice were
anesthetized using 1% (w/v) pentobarbital sodium (90 mg/kg) and
arranged in proper position and exposed to μCT at 60 kV for 800
msec using an Inveon MM Gantry-LG CT.
Primary cell culture of metastatic breast
tumor cells
To isolate tumor cells from the osteolytic lesions,
mice with overt bone metastases were sacrificed, and then the
affected hindlimbs were separated from the body at the joints. Both
ends of the long bones were cut open after skin and muscle was
removed using a scalpel. Mouse bone marrow cells as well as tumor
cells were washed out by PBS using a 1-ml syringe with a 29G
needle. All those cells were collected by centrifuging and washed
with PBS before being cultured in 5-cm plates using L-15 medium
supplemented with 10% FBS.
Mouse bone marrow cells that failed to attach to
plates and thus could be washed off with PBS after the other cells
(almost all of them are tumor cells) adhered to culture plates.
After continuous culture using 400 μg/ml G418 for more than 1 week,
we obtained the subline of bone metastasis named as 231-B cells,
which were verified by measuring the luciferase activity
(Promega).
RNA extraction and expression profiles of
miRNA and mRNA
Cells with about 80% confluence on 6-well plates
were washed twice using ice-cold PBS, and 1 ml TRIzol (Invitrogen)
was added to a well to obtain RNA. Cell lysate in TRIzol was stored
at −80°C and then sent for screening differentially expressed miRNA
and mRNA using Affymetrix GeneChip miRNA 3.0 Arrays and Human mRNA
4×180K (Agilent Technologies). The quantity and quality of
extracted RNA were analyzed by CapitalBio. All data were normalized
and further analyses were carried out by the College of
Bioinformatics Science and Technology, Harbin Medical
University.
Reverse transcription of RNA and
real-time PCR
Total RNA was obtained following the standard method
of TRIzol extraction. cDNA was synthesized using Quant cDNA with
random primers (Tiangen). Moreover, miRNAs were reverse-transcribed
following the instructions with minor alteration (17). In general, one specific reverse
primer targeting individual miRNA was designed to complete the
reverse transcription. Then, one miRNA-specific forward primer and
one universal reverse primer were used in the subsequent real-time
PCR detection. Real-time PCR was completed with SYBR Premix Ex Taq™
II (Takara). The PCR procedure followed the instructions of Takara
in StepOne Plus Real-time PCR system (Applied Biosystems). The
results were analyzed using the 2−ΔΔCt method (18).
Primers used in this study were the following:
miR-429 forward, 5′-ACACTCCAGCTGGGTAATACTGTCTGGTAA-3′ and miR-429
reverse, 5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACGGTTTT-3′,
universal reverse, 5′-TGGTGTCGTGGAGTCG-3′; U6 forward,
CTCGCTTCGGCAGCACA and U6 reverse, AACGCTTCACGAATTTGCGT.
Transfection of microRNA mimics
HiPerFect agent (Qiagen) was used to deliver miRNA
mimics into cells maintained in 6-well plates. According to the
manufacturer’s instruction, cells were grown to ~70% confluence and
20 nM miRNA mimics and 12 μl of transfection agent was mixed
thoroughly in 100 μl of Opti-MEM. After 10 min at room temperature,
the mixture was added into cells. The subsequent experiments were
performed 48 h after the transfection.
Cell invasion assays
The upper chambers of Transwells with 8-μm membrane
pores (Corning) were pre-coated with 60 μl Matrigel matrix gel (BD
Biosciences) at least 1 h before seeding of the tested cells. A
total of 3×104 231-B cells in 100 μl of L15 medium
without FBS were added into the upper chambers and 600 μl of L15
medium with 10% FBS was placed to lower chambers as
chemoattractant. Twelve hours later, the upper chambers were
removed from lower chambers and then wiped using cotton swabs. The
invaded cells were fixed using methanol at room temperature for 15
min, visualized and quantified using crystal violet. Three fields
of each chamber were photographed (x10 magnification) and the
results were from duplicate chambers and are presented as mean ±
SD. This experiment was independently repeated at least twice.
Immunoblot assay
Cells reaching 80% confluence on 6-well plates were
washed twice using ice-cold PBS and 80–100 μl cell lysis buffer (50
mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 1% Triton-100) with protease
inhibitor cocktail (Complete Mini EDTA-free; Roche) added into
plates. On ice, cells were carefully collected with scrapers and
were subjected to lysis for 30 min. Proper amount of Laemmli buffer
was added into 20–30 μg cell lysate and boiled at 95°C for 5 min.
Then the denaturalized protein was resolved on a 10% gel. After
being transferred to PVDF membrane, the protein blot was detected
using corresponding antibodies. The immunoblot images were acquired
and analyzed using ImageQuant LAS 4000 System. The primary
antibodies used in this experiment were ZEB1 and β-actin.
Statistical analyses
GraphPad 6 was used to evaluate the statistical
significance of data. Unless otherwise indicated, t-test was
adopted to calculate the statistical significance. A P-value
<0.05 was considered significant.
Results
Breast cancer cells invade bones of
Balb/c nude mice
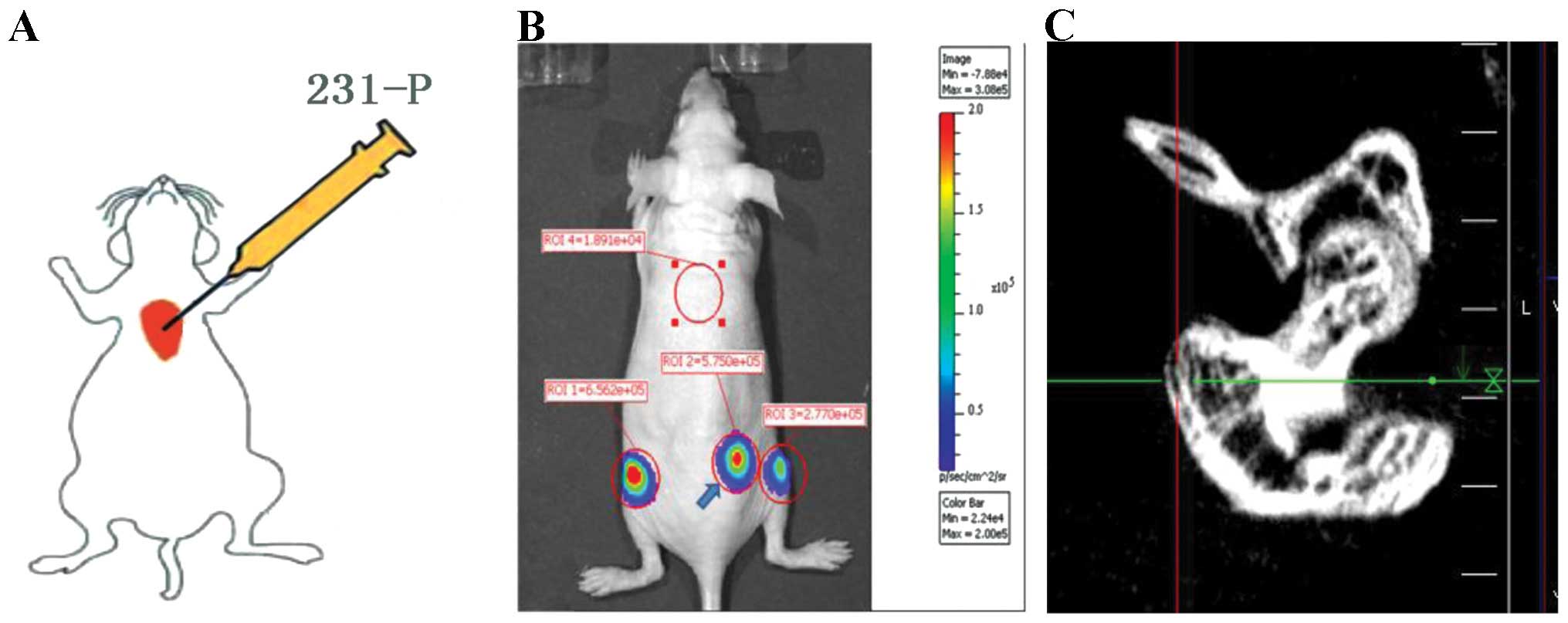
In the present study, we have established a mouse
model of bone metastasis. The human mammary carcinoma MDA-MB-231
cells, which constitutively expressed luciferase, were inoculated
into the left cardiac ventricle of immunodeficient mice (Fig. 1A). Eight to twelve weeks later, 5
mice eventually developed bone metastasis among the 15 mice under
experiment. The bone lesions mainly occur on the long bones of the
hind limbs and the ribs, which were observed by Xenogen Corporation
Optical in vivo imaging and μCT (Fig. 1B and C).
Primary culture of bone metastatic breast
cancer cells
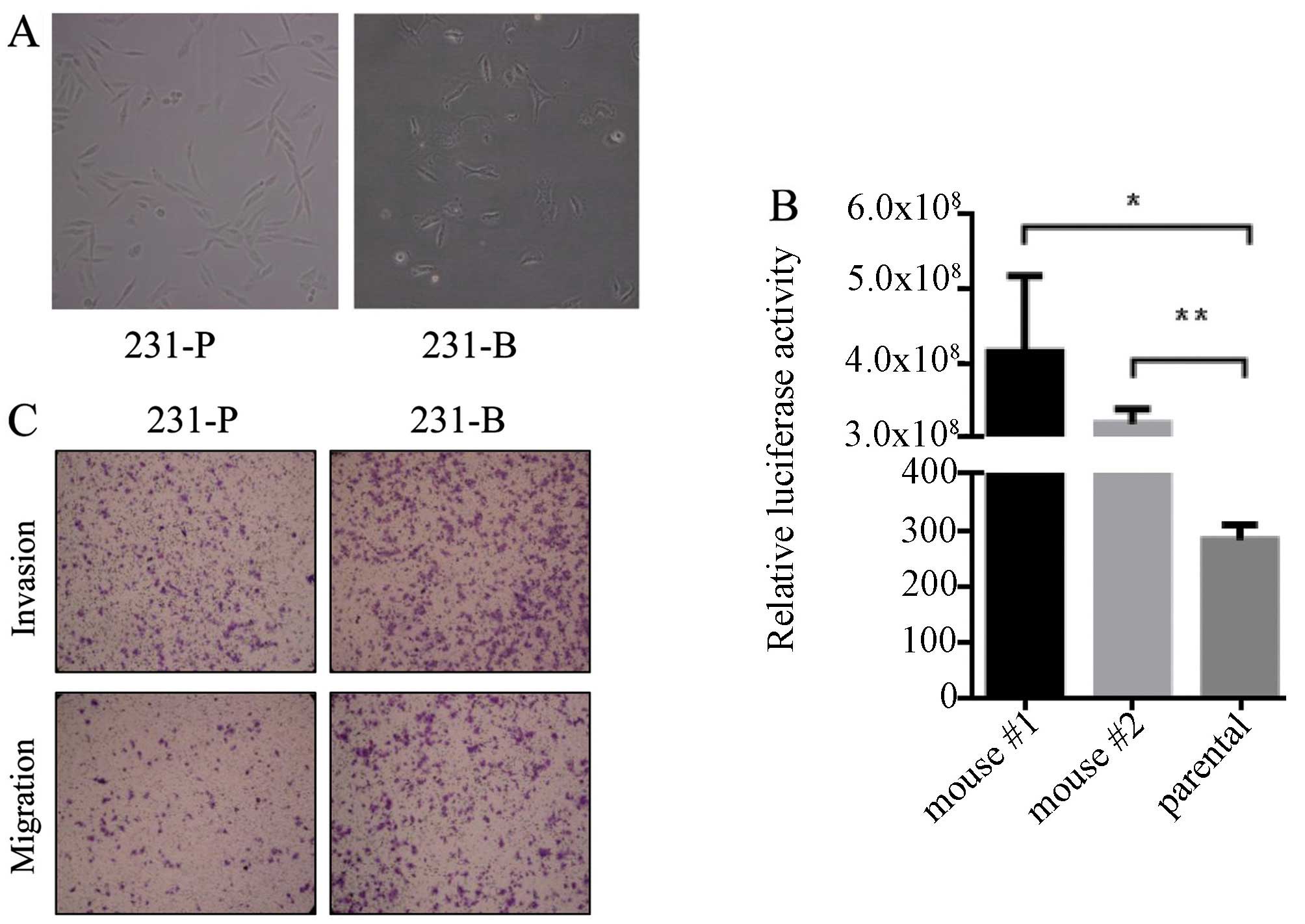
After verifying the presence of bone metastases, we
collected the metastatic tumor cells from the affected bones. G418
were added into the culture medium to wipe out non-malignant cells.
After about 10 days, we obtained 231-B cells (Fig. 2A) of which morphology was different
from 231-P cells. Then, we tested the luciferase activity of 231-B
cells, obtaining much higher luciferase activity compared with the
negative control cells (Fig. 2B).
Moreover, in vitro invasion assay showed that 231-B cells
had stronger invading capability than 231-P cells (Fig. 2C). Thus, we obtained bone
metastatic breast tumor cells from the established model.
Differentially expressed miRNAs of 231-P
and 231-B cells
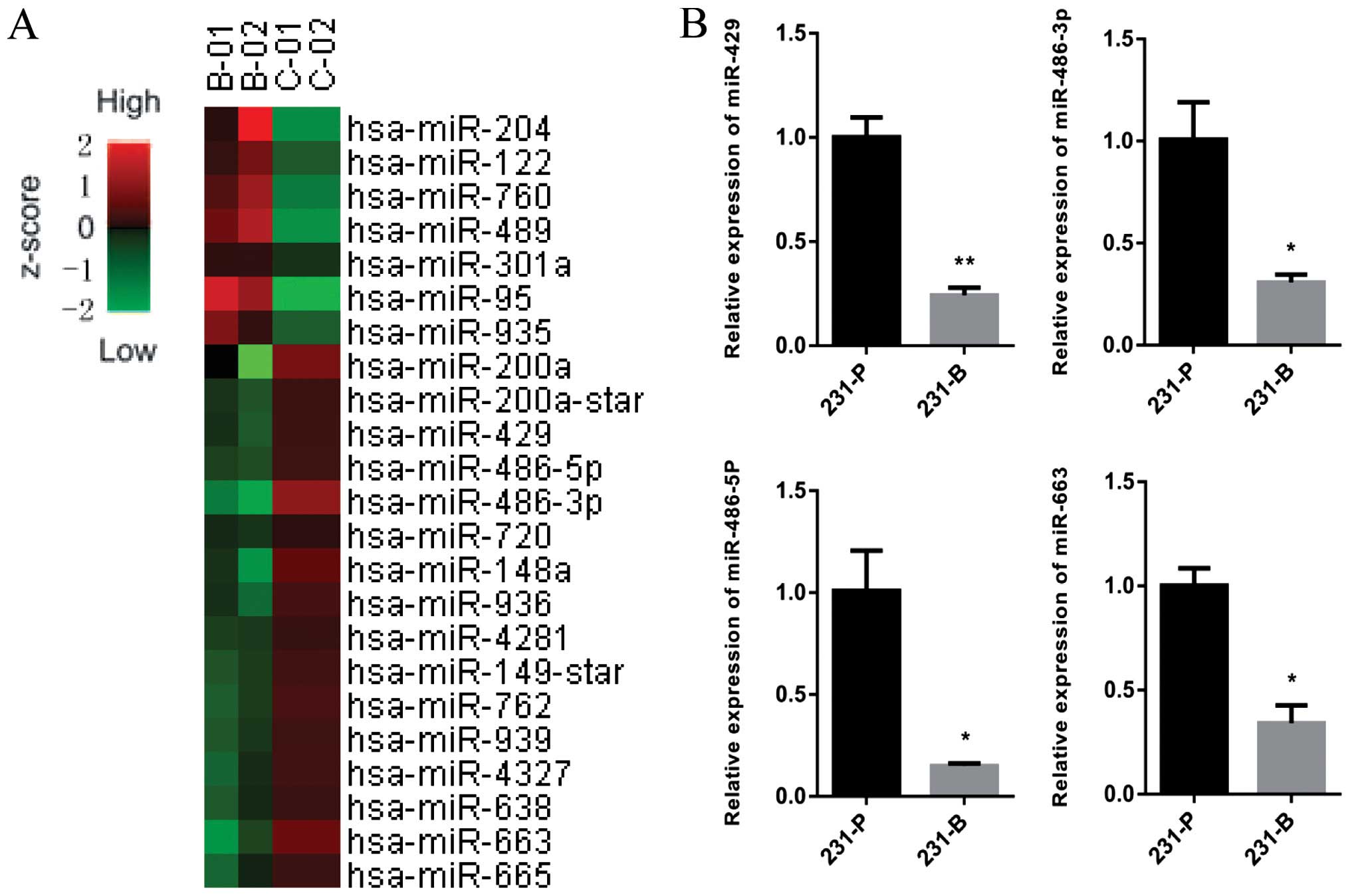
RNA samples from 231-P and 231-B cells were prepared
and analyzed using Affymetrix GeneChip miRNA 3.0 Arrays. Among the
detected miRNAs that amounted to 3,949, 118 miRNAs manifested
differential expression between 231-P and 231-B cells, of which 16
upregulated whereas 102 downregulated >2-fold (P<0.05). Some
of the differentially expressed miRNA are shown in Fig. 3A. Subsequent GO enrichment analysis
showed that the functions of these miRNAs focused on ‘regulation of
cell death’ (P=1.49xe−6), ‘signaling’
(P=3.25xe−6), ‘regulation of localization’ (P=6.31
xe−6) and ‘regulation of cell migration’
(P=0.00028).
In addition, miR-429, miR-663, miR-486-5p and
miR-486-3p were verified in RNA samples derived from 231-P and
231-B cells by real-time PCR assay. Results showed that these
miRNAs indeed had distinct expression patterns, indicating that
they probably play important roles in bone metastasis of breast
cancer cells (Fig. 3B).
miR-429 suppresses invasion of 231-B
cells in vitro
Previous reports showed that miR-429 could inhibit
migration and metastasis of several types of cancers.
Overexpression of miR-429 inhibits invasion and promotes apoptosis
in esophageal carcinoma cells by targeting Bcl-2 and SP1 (19). miR-429 inhibits cells invasion by
targeting Onecut2 in colorectal carcinoma (20). Our results were in line with the
above investigations.
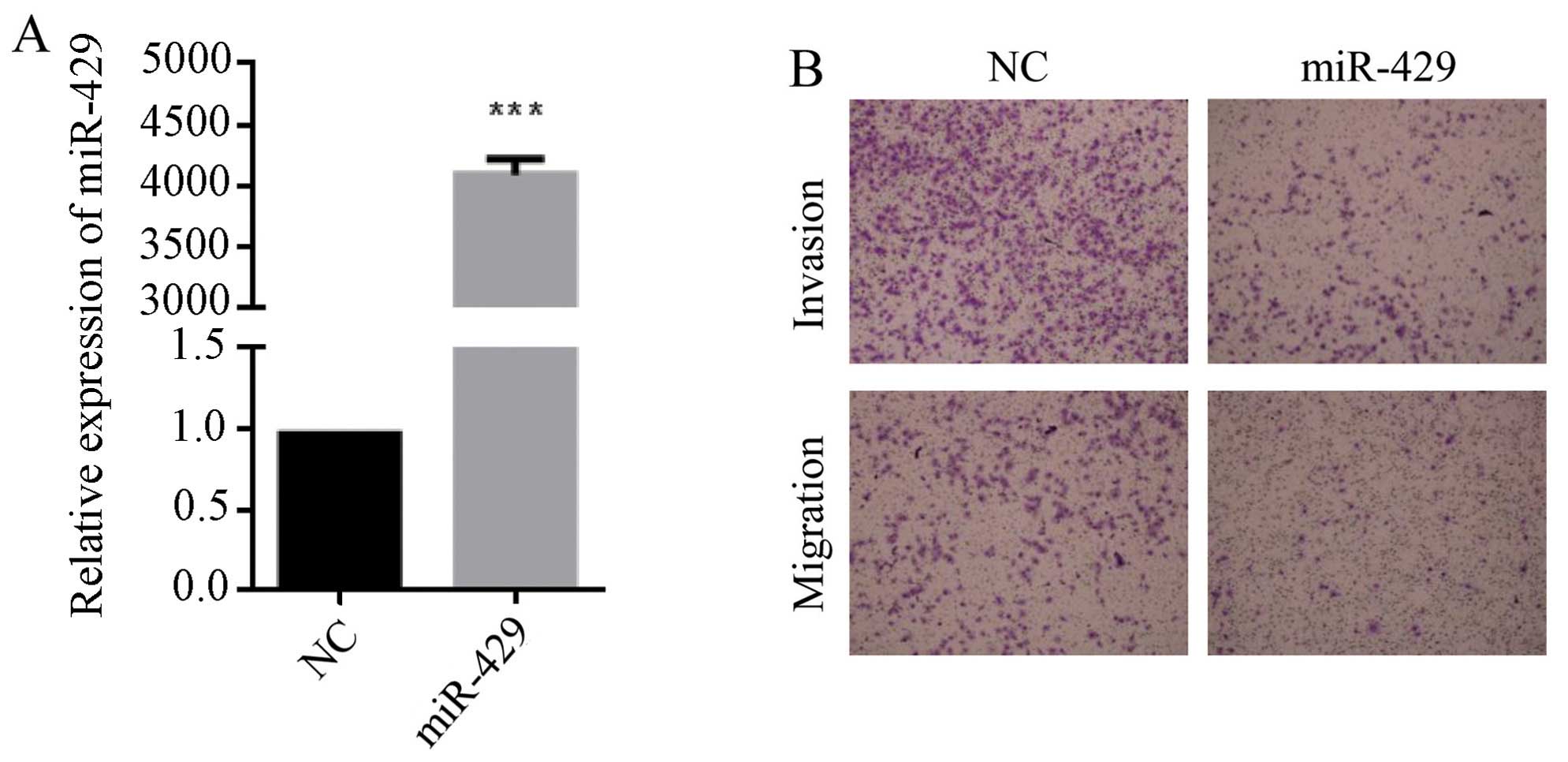
We noted that miR-429 expression was significantly
lower in 231-B cells than that of 231-P cells (Fig. 3B). We transfected miR-429 mimics
into 231-B cells, which was verified by real-time PCR assay
(Fig. 4A), and found that
overexpression of miR-429 could dramatically reduce the migration
and invasiveness of 231-B cells (Fig.
4B). Taken together, miR-429 probably regulated bone metastasis
of breast cancer cells in a negative manner.
miR-429 mediates inhibitory function by
targeting multiple genes
To gain insight into the roles of miR-429 inhibiting
bone metastasis of MDA-MB-231 cells, several computational
algorithms (including TargetScan, DIANA-microT, microRNA.org,
miRDB, RNA22-HSA and PITA) were used to predict the candidate
targets of miR-429, ZEB1, ZEB2, WIPF1, MARCKS, TRIM33, WASF3, CRKL,
WAPAL and NTRK2 were predicted as candidate targets of miR-429
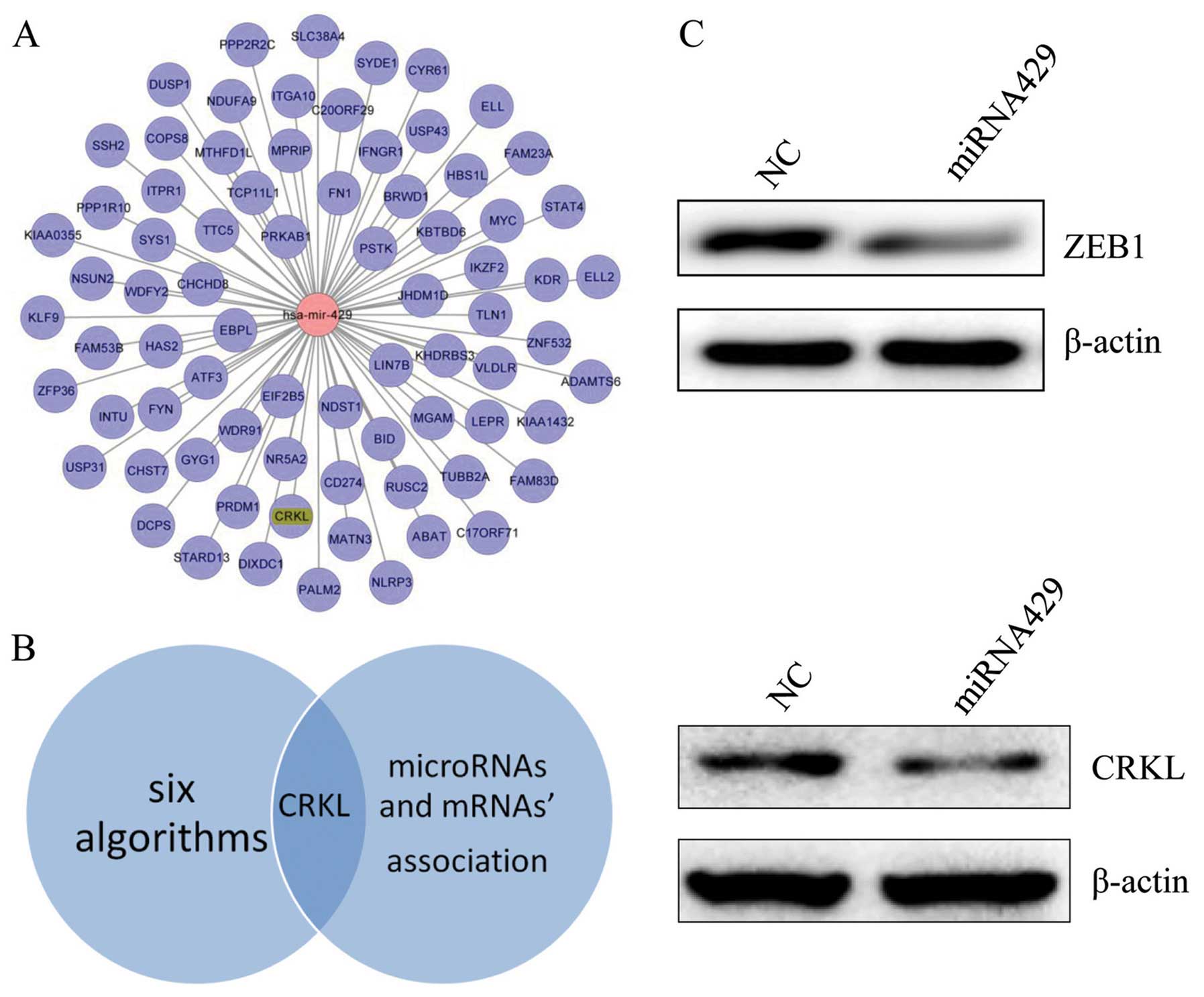
(Table I). Fig. 5A shows the possible regulation
network between miRNAs and mRNAs. Then, we combined the in
silico analysis of miR-429 targets and global transcriptional
profile (Fig. 5B). Among all these
results, CRKL was the target gene on both sides, indicating this
gene was probably a fundamental node in controlling bone metastasis
of breast cancer. Moreover, immunoblot assay showed that miR-429
reduced ZEB1 and CRKL significantly, which is a classical EMT
inducer in breast cancer (Fig.
5C). Collectively, miR-429 negatively modulated several key
invasion and metastasis inducers to suppress motility of breast
cancer cells.
 | Table IThe candidate targets of miR-429,
which were verified to play a vital role in the development of
certain cancer, were predicted in six algorithms. |
Table I
The candidate targets of miR-429,
which were verified to play a vital role in the development of
certain cancer, were predicted in six algorithms.
| TargetScan | DIANA-microT | microRNA.org | miRDB | RNA22-HSA | PITA |
|---|
| ZEB1 | + | + | + | + | + | + |
| ZEB2 | + | + | + | + | + | |
| WIPF1 | + | + | + | + | | + |
| MARCKS | + | + | + | + | | + |
| TRIM33 | + | + | + | + | | + |
| WASF3 | + | + | + | + | | + |
| CRKL | + | + | + | + | | + |
| WAPAL | + | + | + | + | | + |
| NTRK2 | + | | + | + | + | + |
Discussion
Breast cancer morbidity ranks the first accounting
for 29% in all the women suffering cancer, and the mortality ranks
the second accounting for 14% among the women cancer victims in
western countries (21,22). Although great progress has been
made toward preventing and treating breast cancer with surgical
operation, chemotherapy, and radiation therapy, metastasis of
breast cancer still remains one grave clinical challenge due to the
limits of early detection and effective treatment. Breast cancer is
likely to form distant metastases in lung, bone and liver (23). A myriad of factors influence
malignant mammary cells to colonize in bones (24,25).
In this investigation, we set up an in vivo model to study
how microRNAs regulate bone metastases of breast cancer cells.
We established a bone metastasis model using
MDA-MB-231 cells, and further miRNA array analysis revealed the
altered expression of more than 100 miRNAs between 231-P and 231-B
cells. Among these miRNAs, several have already been found to get
involved in invasion and metastasis of tumors. miR-663 could
inhibit proliferation and invasion of glioblastoma in vitro
and in vivo by directly targeting PIK3CD, thus, reducing the
expression of three downstream effectors: CCND1, MMP2 and MMP7
(26). On the other hand, in
castration-resistant prostate cancer cells, miR-663 acted as
OncomiR, which could promote invasion of malignant cells. Moreover,
this miRNA correlated with TNM stage and could be used as an
independent prognostic marker in clinical recurrence (27). Besides, miR-486-5p, expression of
which in 231-B cells was decreased by 70% compared with that of
231-P cells, was recently reported to inhibit metastasis of lung
cancer by reducing the protein level of ARHGAP5 (28). Taken together, most array results
from comparison between 231-P and 231-B cells were in line with
previous literature and these differentially expressed miRNAs are
probably implicated in bone metastasis of breast cancer.
miR-429 is a member of the miR-200 family, four
members of this family have been found to play important roles in
the regulation of EMT in various types of tumors (29). EMT is widely accepted as an
essential mechanism to prompt malignant cells to invade and
colonize distant organs (30). In
the present study, we showed that downregulation of miR-429 could
promote bone metastasis of breast cancer, which is in line with
previously reported results in several types of cancers. Besides in
breast cancer, overexpression of miR-429 suppressed invasion and
metastasis of colorectal carcinoma and ovarian cancer cells,
respectively (20,31).
miRNAs usually function through base-pairing
specific sequences located in 3′-UTRs of targeted genes to hinder
translation of mRNA or enhance degradation of mRNA (32). ZEB1 is the most well studied target
of miR-429, since this transcription factor is an important EMT
inducer (33). Besides ZEB1,
several candidate targets of miR-429 were also identified essential
for invasion and metastasis in cancer models. For example, Onecut2
was proved to be one bona fide target of miR-429 in colon
cancer (20). Combined analysis of
expression array results and in silico target search
revealed CRKL as a common candidate. CRKL protein is a kinase
harboring SH2 and SH3 domains. CRKL transformed fibroblasts and
received signals from BCR-ABL tyrosine kinase (34–36).
CRKL influences integrin mediated adhesion to fibronectin (37). For cellular motility, CRKL enhanced
motion through forming a complex with Cbl and C3G to relay signals
in hematopoietic Ba/F3 cells (38). In addition to promoting invasion of
cells, CRKL also could suppress caspase-8 mediated apoptosis, which
was also essential for distant metastasis (39). Another research group also found
the involvement of CRKL in integrin-triggered cell migration, and
they further identified that CRKL performed by acting as downstream
effector of Src (40). Considering
that CRKL is important in proliferation, migration and evading
apoptosis, we speculate that CRKL is probably essential in bone
metastasis of breast cancer, however this needs further
investigation.
In conclusion, we established an in vivo bone
metastasis model of breast cancer, and we obtained a wealth of
information on how mammary malignant cells invade and finally form
overt metastases in bones. Further research is urgently needed to
clarify the complicated mechanisms underlying this fatal
pathological condition to provide valuable prevention and treatment
targets.
Acknowledgements
We appreciate our colleagues in the laboratory of Dr
Zhihua Liu for helpful discussion, and Professor Burton Yang for
his critical reading of this manuscript.
References
|
1
|
Yang W, Lam P, Kitching R, et al: Breast
cancer metastasis in a human bone NOD/SCID mouse model. Cancer Biol
Ther. 6:1289–1294. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xia TS, Wang GZ, Ding Q, et al: Bone
metastasis in a novel breast cancer mouse model containing human
breast and human bone. Breast Cancer Res Treat. 132:471–486. 2012.
View Article : Google Scholar
|
|
3
|
Fidler IJ: Critical determinants of
metastasis. Semin Cancer Biol. 12:89–96. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li Z, Schem C, Shi YH, Medina D and Zhang
M: Increased COX2 expression enhances tumor-induced osteoclastic
lesions in breast cancer bone metastasis. Clin Exp Metastasis.
25:389–400. 2008. View Article : Google Scholar
|
|
5
|
Rosol TJ, Tannehill-Gregg SH, Corn S,
Schneider A and McCauley LK: Animal models of bone metastasis.
Cancer Treat Res. 118:47–81. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lynam-Lennon N, Maher SG and Reynolds JV:
The roles of microRNA in cancer and apoptosis. Biol Rev Camb Philos
Soc. 84:55–71. 2009. View Article : Google Scholar
|
|
7
|
Qiao J, Lee S, Paul P, et al: miR-335 and
miR-363 regulation of neuroblastoma tumorigenesis and metastasis.
Surgery. 154:226–233. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yi C, Wang Q, Wang L, et al: MiR-663, a
microRNA targeting p21WAF1/CIP1, promotes the
proliferation and tumorigenesis of nasopharyngeal carcinoma.
Oncogene. 31:4421–4433. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pan J, Hu H, Zhou Z, et al:
Tumor-suppressive miR-663 gene induces mitotic catastrophe growth
arrest in human gastric cancer cells. Oncol Rep. 24:105–112.
2010.PubMed/NCBI
|
|
10
|
Yau WL, Lam CS, Ng L, et al:
Over-expression of miR-106b promotes cell migration and metastasis
in hepatocellular carcinoma by activating epithelial-mesenchymal
transition process. PLoS One. 8:e578822013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dynoodt P, Speeckaert R, De Wever O, et
al: miR-145 overexpression suppresses the migration and invasion of
metastatic melanoma cells. Int J Oncol. 42:1443–1451.
2013.PubMed/NCBI
|
|
12
|
Li LZ, Zhang CZ, Liu LL, et al: miR-720
inhibits tumor invasion and migration in breast cancer by targeting
TWIST1. Carcinogenesis. 35:469–478. 2014. View Article : Google Scholar
|
|
13
|
Wang C, Bian Z, Wei D and Zhang JG:
miR-29b regulates migration of human breast cancer cells. Mol Cell
Biochem. 352:197–207. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kutner RH, Zhang XY and Reiser J:
Production, concentration and titration of pseudotyped HIV-1-based
lentiviral vectors. Nat Protoc. 4:495–505. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yin JJ, Selander K, Chirgwin JM, et al:
TGF-beta signaling blockade inhibits PTHrP secretion by breast
cancer cells and bone metastases development. J Clin Invest.
103:197–206. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kang Y, Siegel PM, Shu W, et al: A
multigenic program mediating breast cancer metastasis to bone.
Cancer Cell. 3:537–549. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen C, Ridzon DA, Broomer AJ, et al:
Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic
Acids Res. 33:e1792005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Li M, Zang W, et al: MiR-429
up-regulation induces apoptosis and suppresses invasion by
targeting Bcl-2 and SP-1 in esophageal carcinoma. Cell Oncol
(Dordr). 36:385–394. 2013. View Article : Google Scholar
|
|
20
|
Sun Y, Shen S, Liu X, et al: miR-429
inhibits cells growth and invasion and regulates EMT-related marker
genes by targeting Onecut2 in colorectal carcinoma. Mol Cell
Biochem. 390:19–30. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Smith RA, Brooks D, Cokkinides V, Saslow D
and Brawley OW: Cancer screening in the United States, 2013: a
review of current American Cancer Society guidelines, current
issues in cancer screening, and new guidance on cervical cancer
screening and lung cancer screening. CA Cancer J Clin. 63:88–105.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Weigelt B, Peterse JL and van’t Veer LJ:
Breast cancer metastasis: markers and models. Nat Rev Cancer.
5:591–602. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mundy GR: Metastasis to bone: causes,
consequences and therapeutic opportunities. Nat Rev Cancer.
2:584–593. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Weilbaecher KN, Guise TA and McCauley LK:
Cancer to bone: a fatal attraction. Nat Rev Cancer. 11:411–425.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shi Y, Chen C, Zhang X, et al:
Primate-specific miR-663 functions as a tumor suppressor by
targeting PIK3CD and predicts the prognosis of human glioblastoma.
Clin Cancer Res. 20:1803–1813. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiao L, Deng Z, Xu C, et al: miR-663
induces castration-resistant prostate cancer transformation and
predicts clinical recurrence. J Cell Physiol. 229:834–844. 2014.
View Article : Google Scholar
|
|
28
|
Wang J, Tian X, Han R, et al:
Downregulation of miR-486-5p contributes to tumor progression and
metastasis by targeting protumorigenic ARHGAP5 in lung cancer.
Oncogene. 33:1181–1189. 2014. View Article : Google Scholar :
|
|
29
|
Korpal M and Kang Y: The emerging role of
miR-200 family of microRNAs in epithelial-mesenchymal transition
and cancer metastasis. RNA Biol. 5:115–119. 2008. View Article : Google Scholar
|
|
30
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen J, Wang L, Matyunina LV, Hill CG and
McDonald JF: Overexpression of miR-429 induces
mesenchymal-to-epithelial transition (MET) in metastatic ovarian
cancer cells. Gynecol Oncol. 121:200–205. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pasquinelli AE: MicroRNAs and their
targets: recognition, regulation and an emerging reciprocal
relationship. Nat Rev Genet. 13:271–282. 2012.PubMed/NCBI
|
|
33
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: an alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ten HJ, Kaartinen V, Fioretos T, et al:
Cellular interactions of CRKL, and SH2-SH3 adaptor protein. Cancer
Res. 54:2563–2567. 1994.
|
|
35
|
Heaney C, Kolibaba K, Bhat A, et al:
Direct binding of CRKL to BCR-ABL is not required for BCR-ABL
transformation. Blood. 89:297–306. 1997.PubMed/NCBI
|
|
36
|
Senechal K, Halpern J and Sawyers CL: The
CRKL adaptor protein transforms fibroblasts and functions in
transformation by the BCR-ABL oncogene. J Biol Chem.
271:23255–23261. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Uemura N, Salgia R, Ewaniuk DS, Little MT
and Griffin JD: Involvement of the adapter protein CRKL in
integrin-mediated adhesion. Oncogene. 18:3343–3353. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Uemura N and Griffin JD: The adapter
protein Crkl links Cbl to C3G after integrin ligation and enhances
cell migration. J Biol Chem. 274:37525–37532. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Graf R, Barbero S, Keller N, et al:
Src-inducible association of CrkL with procaspase-8 promotes cell
migration. Cell Adh Migr. 7:362–369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li L, Guris DL, Okura M and Imamoto A:
Translocation of CrkL to focal adhesions mediates integrin-induced
migration downstream of Src family kinases. Mol Cell Biol.
23:2883–2892. 2003. View Article : Google Scholar : PubMed/NCBI
|