Introduction
Breast cancer is one of the most common types of
cancer in women. Women with advanced breast cancer ultimately
develop bone metastasis which increases significantly morbidity and
mortality. Approximately 20–30% of human tumors contain mutated
versions of Ras proteins. Ras is an important signal transducing
protein for growth factor activated pathways. Normal Ras binds GTP
and in the GTP-bound state interacts with numerous effectors
including the Raf proto-oncogene kinase and
phosphatidylinositol-3-kinase (PI3K). The Rho family of GTPases
includes the Rho, Rac and CDC42 genes (1). While the functions of these GTPases
have not been clarified, it appears that the Rho proteins play a
role in regulating cell morphology (2,3).
The p53 gene is known as the guardian of the
genome (4). A major biological
function of p53 is to respond to stress signals and activate the
transcription of downstream target genes involved in important
cellular mechanisms such as cell cycle control, DNA repair and
apoptosis. The p53 has two very distinct roles in cell cycle
control mechanisms. The first is a protective (cytostatic) one in
which p53 arrests cells in the G1 phase of the cell cycle upon
sensing DNA damage. In a second role, p53 initiates apoptosis upon
irreparable damage to the cell.
Serpins are serine protease inhibitors structurally
conserved molecules encompassing nearly all branches of life
(5). Among newly characterized
serpin functions, regulation of cellular proliferation through
apoptosis modulation and proteasome disturbance seem to play a
major role. Accordingly, several serpins were found to be
overexpressed in tumor cells. Indeed, apoptosis dysregulation is
likely to be a cornerstone in both tumorigenesis and autoimmunity,
since uncontrolled cellular viability results in tumor
proliferation, while inefficient disposal of apoptotic debris may
favor the rescue of auto-reactive immune cells.
Caveolin is a specialized lipid raft on the plasma
membrane found in mesenchymal cells (6–9). The
caveolin family consists of three members, caveolin-1 (cav-1),
caveolin-2 and caveolin-3. Cav-1 is widely expressed in various
tissues. Previous studies demonstrated the essential role of
caveolin in a number of human diseases including cancer (6). Previously, both the epithelial and
stromal caveolin have been detected in breast cancer patients to
determine the prognosis (9).
Apoptosis has been defined as a genetic death
program leading to the ordered destruction of cellular components,
while membrane integrity is maintained (10). The balance between pro- and
anti-apoptotic signals guarantee biological homeostasis and its
disturbance is highly related to malignant transformation (10). Two distinct major pathways have
been described that lead to activation of aspartate-specific
cysteine proteases (caspases) that finally mediate apoptosis. The
extrinsic pathway involves binding of extracellular death ligands
to specific cell surface death receptors and formation of the
death-inducing signaling complex. The intrinsic pathway depends on
mitochondria, is initiated by intracellular signals such as DNA
damage and needs activation of the tumor suppressor p53 (11).
NFκB is a family of 5 transcriptional factors
including p50, p52, p65 (Rel-A), Rel-B and c-Rel (12). NFκB is recognized as a key positive
regulator of cancer cell proliferation and survival via its ability
to transcriptionally activate many pro-survival and anti-apoptotic
genes such as XIAP, Bcl-2, Bcl-xL, IkB-a, cIAP1, cIAP-2 and
survivin (12,13).
Bisphosphonates are potent antiresorptive drugs used
to the treatment of metabolic bone disease. It has been shown that
these compounds have direct effects on tumor cells in vitro,
including the induction of apoptosis in human myeloma cell lines
(14,15). Pamidronate is a member of
nitrogen-containing bisphosphonates used in the treatment of bone
metastasis of breast and prostate cancer. It has been reported as a
mechanism of action inhibiting the mevalonate pathway which is a
requirement for the prenylation (activation) of small GTP-binding
proteins such as farnesylpyrophospate synthase and/or
geranylgeranylpyrophosphate synthase thus blocking the generation
of isoprenoid compounds, farnesylpyrophosphate and/or
geranylgeranylpyrophosphate. The aim of this study was to evaluate
antitumor activity of pamidronate in breast cancer cell model with
cells transformed by low doses of α-particles and estrogen in
vitro (16).
Materials and methods
Breast cancer cell lines
The spontaneously immortalized breast epithelial
cell line, MCF-10F cells was grown in DMEM/F-12 (1:1) medium
supplemented with antibiotics 100 U/ml penicillin, 100 μg/ml
streptomycin, 2.5 μg/ml amphotericin B (all from Life Technologies,
Grand Island, NY, USA) and 10 μg/ml and 5% equine serum (Biofluids,
Rockville, MD, USA), 0.5 μg/ml hydrocortisone (Sigma, St. Louis,
MO, USA) and 0.02 μg/ml epidermal growth factor (Collaborative
Research, Bedford, MA). An in vitro experimental breast
cancer model developed by exposure of the immortalized human breast
epithelial cell line was used. MCF-10F was exposed to low doses of
high LET (linear energy transfer) α-particle radiation (150 keV/μm)
and subsequent growth in the presence or absence of 17β-estradiol
at 10−8 M (E or Estrogen) (Sigma-Aldrich). This model
consisted of human breast epithelial cells in different stages of
transformation: i) a control cell line, MCF-10F; ii) a
non-malignant cell line, named Alpha3; iii) a malignant and
tumorigenic, cell line named Alpha5 and iv) Tumor2 derived from
cells originated from a tumor after injection of Alpha5 cells into
the nude mice (16).
MTT assay
The metabolic activity of living cells, as indicator
of viability, was determined by 3-(4, 5-dimethylthiazol-2-yl)-2,
5-diphenyltetrazolium bromide (MTT) assay. MCF-10F cell line
(5×104) was seeded in 24-well microplates and incubated
in culture medium for 24 and 48 h at 37°C and 5% CO2.
After incubation, MCF-10F was treated with a series of
concentrations (10, 30, 50 and 80 μM) of pamidronate
(Sigma-Aldrich, Oakville, ON, Canada). After the treatment of
cells, reduction of MTT was determined following manufacturer’s
instructions. The treatment groups were compared with the control
group and the results were expressed as percentage of viable
cells.
Cell invasion and migration assays
Invasiveness and migration was carried out as
previously described (16) using
modified Boyden’s chambers (Corning, New York, NY, USA) constructed
with multiwell cell culture plates and cell culture inserts. The
upper chambers of Transwells with 8-μm membrane pores were
pre-coated with 60 μl Matrigel matrix gel (BD Biosciences) at least
1 h before seeding the cells studied. A total of 3×104
cells in 100 μl of medium without fetal bovine serum (FBS) was
added into the upper chambers and 600 μl of medium with 10% FBS was
placed to lower chambers as chemoattractant. Twelve hours later,
the upper chambers were removed from lower chambers and then wiped
using cotton swabs. The invaded and migrated cells were fixed using
methanol at room temperature for 15 min, visualized and quantified
using crystal violet. Three fields of each chamber were
photographed (magnification, ×40) and the results are from
duplicate chambers and are presented as mean ± SEM. The experiment
was independently repeated at least twice.
Western blot analysis
Cells were lysated with 1 ml lysis buffer (pH 7.2)
(Tris Base (50 mM), EDTA (1 mM), NaCl (100 mM), PMSF (1 mM),
ortovanadate (1 mM), Triton X-100 (0.1%) and centrifuged (10,000
rpm × 10 min). The supernatant with cellular proteins were
dissolved in SDS-PAGE sample solution (60 mM) Tris, pH 6.5, 10%
(w/v) glycerol, 5% (w/v) β-mercaptoethanol, 20% (w/v) SDS, and
0.025% (w/v) bromophenol blue and denatured by boiling (2×5 min),
sonication (2×5 min in a sonication bath), and vortex mixing (2×30
seg). The total amount of protein was 40 μg in each lane with
standard protein markers (Bio-Rad Laboratories, Hercules, CA, USA).
After fractionation by SDS-PAGE on gels (7×14 cm), proteins were
electro-blotted onto PVDF membrane (Amersham Biosciences, Amersham,
UK) using a blotting apparatus (Bio-Rad Laboratories). Prestained
SDS-PAGE (Standards) blots were blocked for 2 h in 10% defatted dry
milk-TBS-0.1% Tween and then incubated for 2 h at room temperature
with corresponding primary antibodies (1:200) Rho-A (26C4) sc-418
and β-actin (C4) sc-47778 followed by incubation with secondary
peroxidase-conjugated mouse IgG (1:5000) (Cell Signaling, CA, USA)
in 5% defatted dry milk-TBS-0.1% Tween. All steps were performed at
room temperature, and blots were rinsed between incubation steps
with TBS-0.1% Tween. Cell blots were probed with mouse anti-actin
antibody as control. Immunoreactive bands were visualized by using
the ECLTM Western Blotting Detection Reagent detection
method (Amersham, Dübendorf, Switzerland) and exposure of the
membrane to X-ray film. Protein determination was performed using
the Bicinchoninic Acid Method (Bio-Rad Laboratories) and BSA as the
standards. Experiments were performed three times.
RNA extraction and cDNA synthesis
Total RNA from control and treated cells were
isolated using TRIzol reagent (Invitrogen Corp., Carlsbad, CA, USA)
according to the manufacturer’s recommendations. Total RNA (2 μg)
was reverse-transcribed to cDNA using High capacity cDNA Reverse
Transcription kit (Applied Biosystems, Carlsbad, CA, USA) and 10
units of RNase inhibitor (Applied Biosystems).
RT-qPCR
An aliquot of cDNA (2 μl) was used in 20 μl qPCR
reaction containing SYBR Green PCR Master Mix (Agilent, La Jolla,
CA, USA) and 5 μM of each primer for the target genes Rho-A,
c-Ha-ras, p53, Serpin-1, Caveolin-1, Bcl-xL and NFκB or
reference gene (β-actin). Table I
shows the primers for the gene selected to develop cDNA probes. The
reaction was performed in a CFX 96 Touch Real-Time PCR Detection
Systems (Bio-Rad Laboratories) with the following conditions: 95°C
for 10 min and 40 cycles of a 2-step program of 95°C for 10 sec and
61°C for 45 sec when fluorescence-reading occurs. After
amplification, PCR product was monitored through dissociation curve
analysis (measurement of fluorescence during an increasing heating
of 2°C/min from 61 to 95°C). At this step, undesirable DNA
contamination (if present) could be detected since primers were
designed to encompass an intron. Reactions were performed in
triplicate and the threshold cycle (Ct) was obtained using Bio-Rad
CFX Manager 2.1 software and the averaged gene expression was
normalized using the reference housekeeping gene β-actin and
relative expression level was calculated. Relative expression was
normalized to the average in normal breast cells.
 | Table IPrimers for genes selected to develop
cDNA probes. |
Table I
Primers for genes selected to develop
cDNA probes.
| Gene name | Product length
(bp)a | Primer
sequenceb |
|---|
| Rho-A | 140 | F:
CCATCATCCTGGTTGGGAAT
R: CATGTACCCAAAAGCGCCA |
| c-Ha-ras | 112 | F:
CCAGTACAGGGAGCAGAT
R: GAGCCTGCCGAGATTCCACA |
| p53 | 128 | F:
CCTCAGCATCTTATCCGAGTGG
R: TGGATGGTGGTACAGTCAGAGC |
| Serpin-1 | 101 | F:
GAGACAGGCAGCTCGGATTC
R: GGCCTCCCAAAGTGCATTAC |
|
Caveolin-1 | 79 | F:
AACGATGACGTGGTCAAGATTG
R: TCCAAATGCCGTCAAAACTGT |
| Bcl-xL | 211 | F:
CTGAATCGGAGATGGAGACC
R: TGGGATGTCAGGTCACTGAA |
| NFκB
(RelA) | 114 | F:
ATCTGCCGAGTGAACCGAAACT
R: CCAGCCTGGTCCCGTGAAA |
| β-actin | 569 | F:
ACTACCTCATGAAGATCCTC
R: TAGAAGCATTTGCGGTGGACGATGG |
Flow cytometry
Phosphatidylserine (PS) is located inside the cell
membrane in normal cells and is transferred to the surface during
the early stage of cell apoptosis. Annexin V, a
Ca2+-dependent phospholipid binding protein, has a
strong binding affinity for PS. Thus, an Annexin V-FITC/7-AAD
staining kit was used to assess pamidronate-induced cell apoptosis.
MCF-10F, Alpha5 and Tumor2 cell lines were cultured to 70%
confluence, then pamidronate with indicated concentrations was
added. After 48 h, cells were harvested and washed twice with
pre-cold PBS and then resuspended in 1X binding buffer at a
concentration of 1×106 cells/ml, 100 μl of such solution
(1×105 cells) was mixed with 10 μl of Annexin V-FITC and
20 μl of 7-amino actinomycin D (7-AAD) (Beckman Coulter, Fullerton,
CA, USA) according to the manufacturer’s instructions. The mixed
solution was incubated at room temperature (25°C) away from light
for 15 min. Then 400 μl of 1X dilution buffer was added to each
tube. Analysis was performed by Beckman Coulter FC500 Flow
Cytometry System with CXP software (Beckman Coulter) within 1
h.
Statistical analysis
Numerical data were expressed as the average ±
standard error of the mean (SEM). Comparison between treated groups
and controls was carried out by ANOVA and Dunnet’s test. A
p<0.05 and p<0.01 were considered to be significant. Lethal
dose at 50% (LD50) was calculated by a non-linear
regression curve using GraphPad Prism 5.0 for Windows (GraphPad
Software, San Diego, CA, USA).
Results
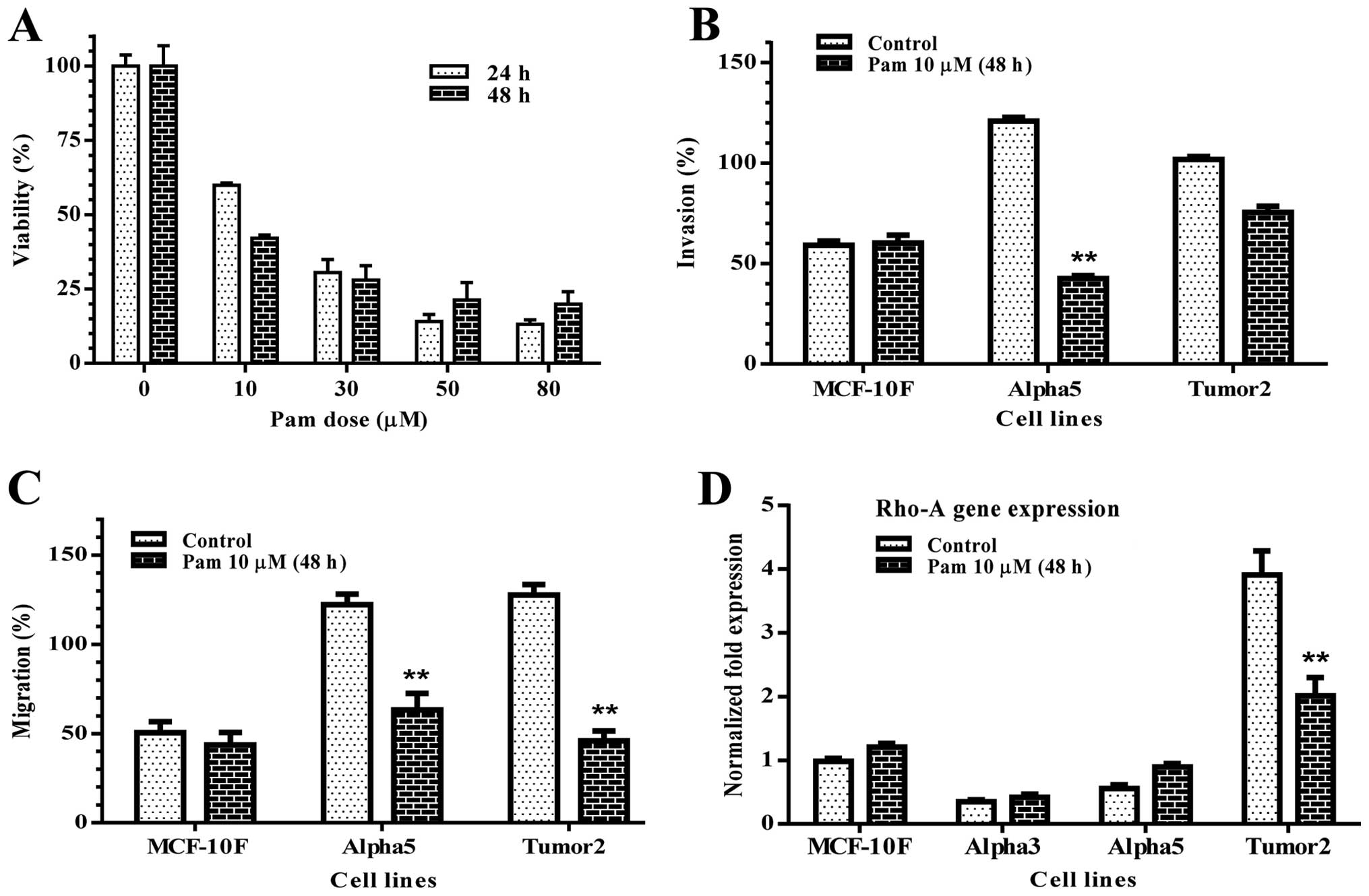
The effect of pamidronate in cell viability in
vitro was analyzed using the MTT assay. Treatment with
increasing concentrations of pamidronate ranging from 0 to 80 μM
resulted in a concentration-dependent decrease in cell number after
24 and 48 h in MCF-10F cell line. Results in Fig. 1A showed that the mean
LD50 was 10 μM. Then, all the experiments were carried
out with 10 μM pamidronate. Migration and invasion assays were
performed to analyze the effect of pamidronate on MCF-10F, Alpha5
and Tumor2 cell lines where basic fibroblast growth factor (b-FGF)
induced the migration of such cells. Results indicated that
pamidronate significantly inhibited (p<0.01) the b-FGF-induced
migration and invasion in both Alpha5 and Tumor2 cell lines
(Fig. 1B and C) compared to the
control MCF-10F. These results suggest that pamidronate influences
cell migration and invasion.
Rho-A is a small GTPase protein known to regulate
the actin cytoskeleton in the formation of stress fibers and it is
generally distributed in the nuclei of cancer cells. Rho-A gene and
protein expression were studied by RT-qPCR and western blot
analysis. Results of the experiments indicated that pamidronate
significantly decreased Rho-A gene expression of the malignant and
tumorigenic cell line Tumor2 (p<0.01) in comparison with its
counterpart (Fig. 1D). In
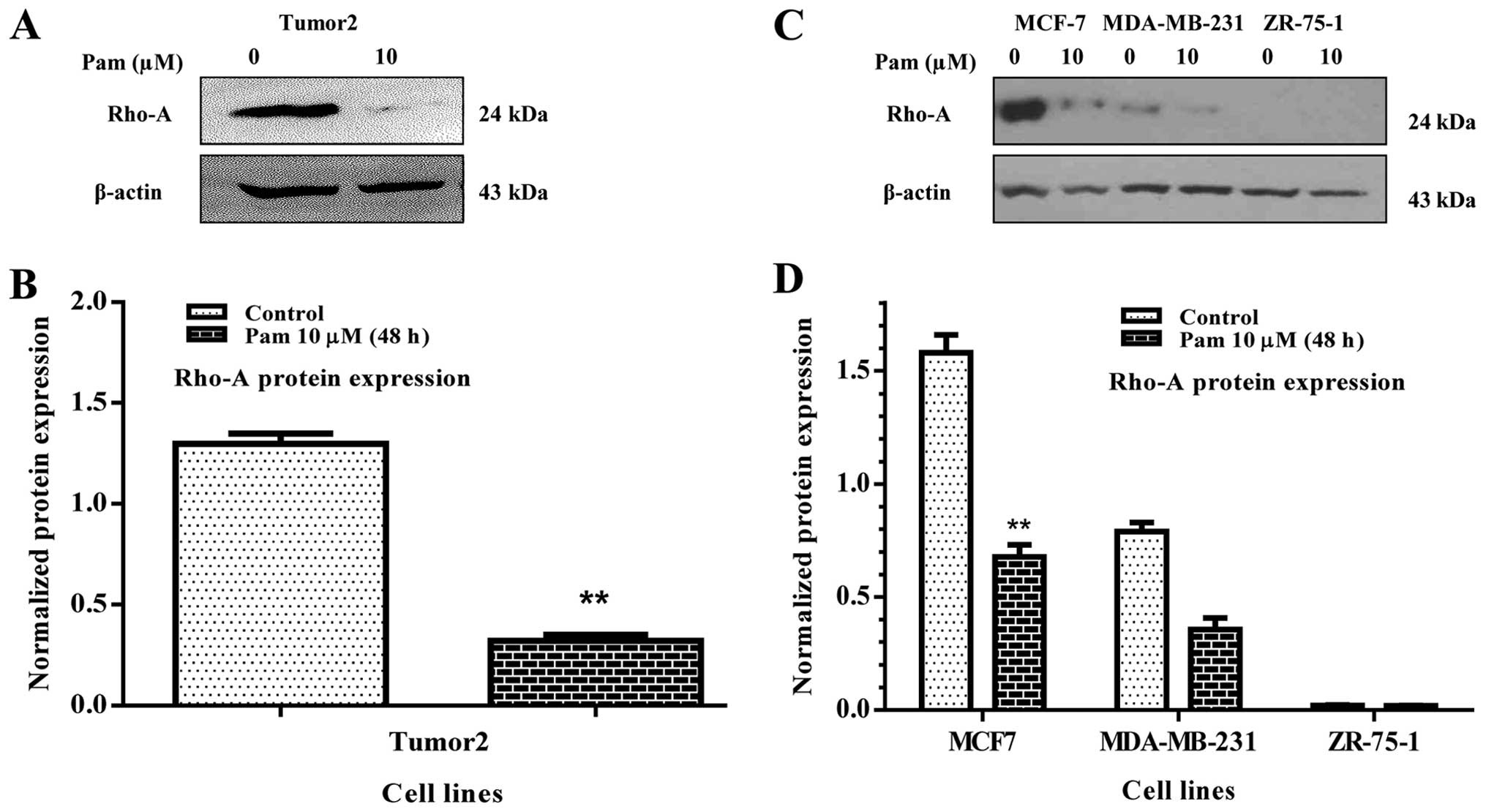
addition, Rho-A protein expression was evaluated in Tumor2 cell
line and other malignant cell lines such as MCF7, MDA-MB-231 and
ZR-75-1. It was found that pamidronate significantly decreased
Rho-A protein expression in Tumor2 (Fig. 2A and B) and MCF7 and MDA-MB-231
(p<0.01) in comparison to their counterparts. There was no Rho-A
protein expression in ZR-75-1 (Fig. 2C
and D).
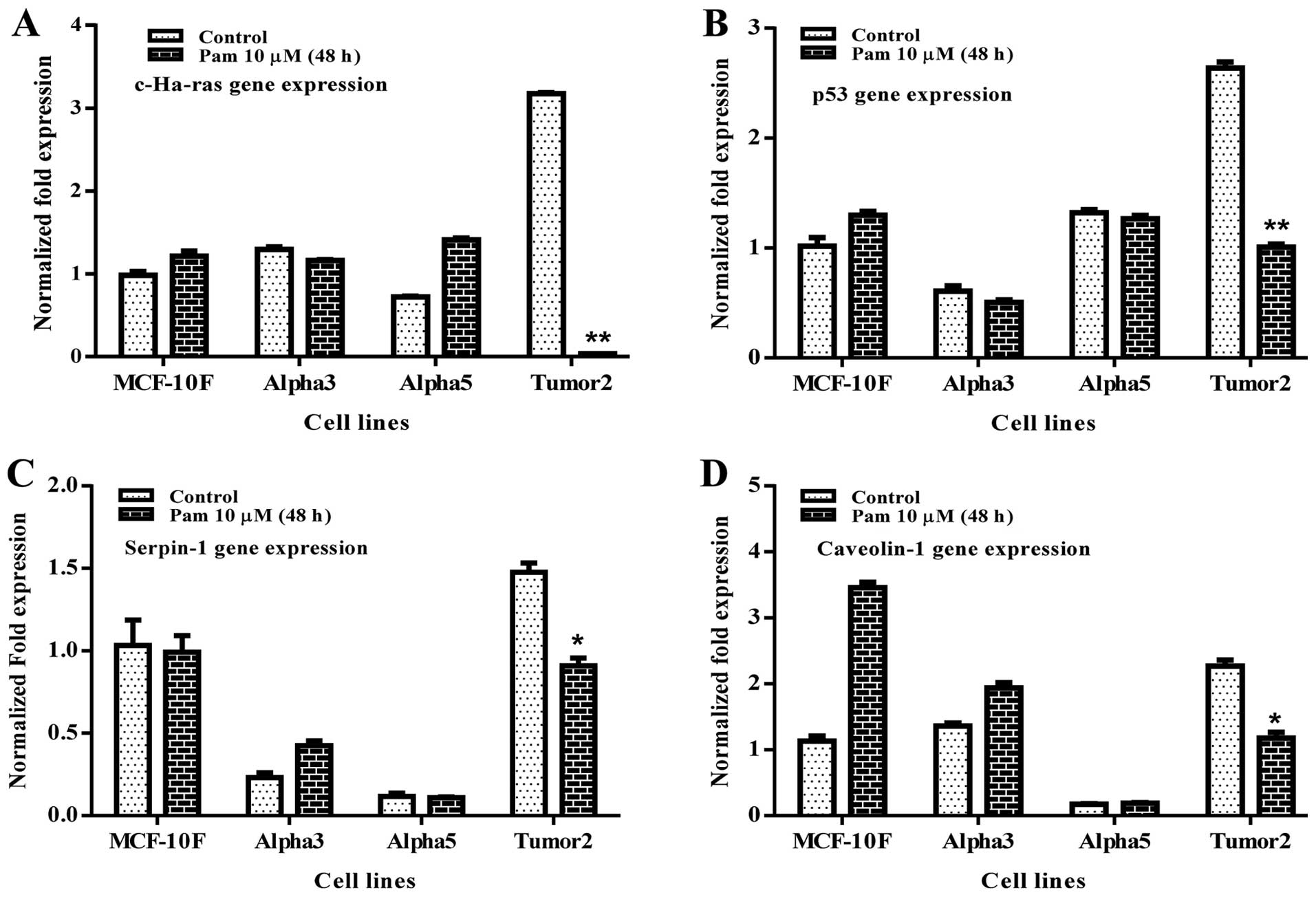
Analysis by real-time PCR indicated that pamidronate
significantly decreased gene expression of c-Ha-ras
(p<0.01), p53 (p<0.01), Serpin-1 (p<0.05)
and Caveolin-1 (p<0.05) in Tumor2 cell line in comparison
to its counterparts. However, Alpha3 and Alpha5 cell lines did not
show any significant difference with their counterparts (Fig. 3).
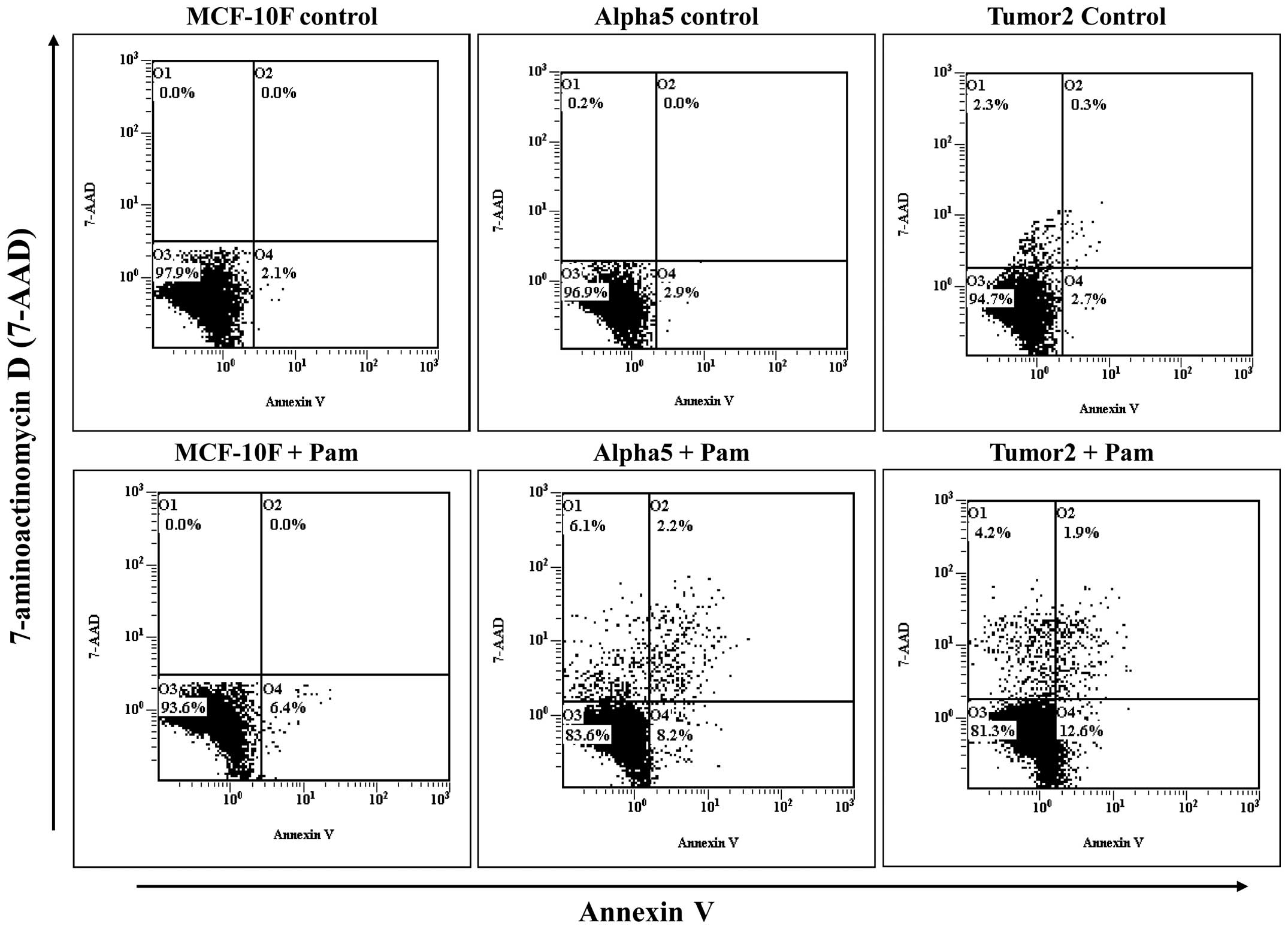
The apoptotic effects of pamidronate on MCF-10F,
Alpha5 and Tumor2 cell lines analyzed by flow cytometry showed 6.4%
of apoptotic cells in the control MCF-10F. On the other hand,
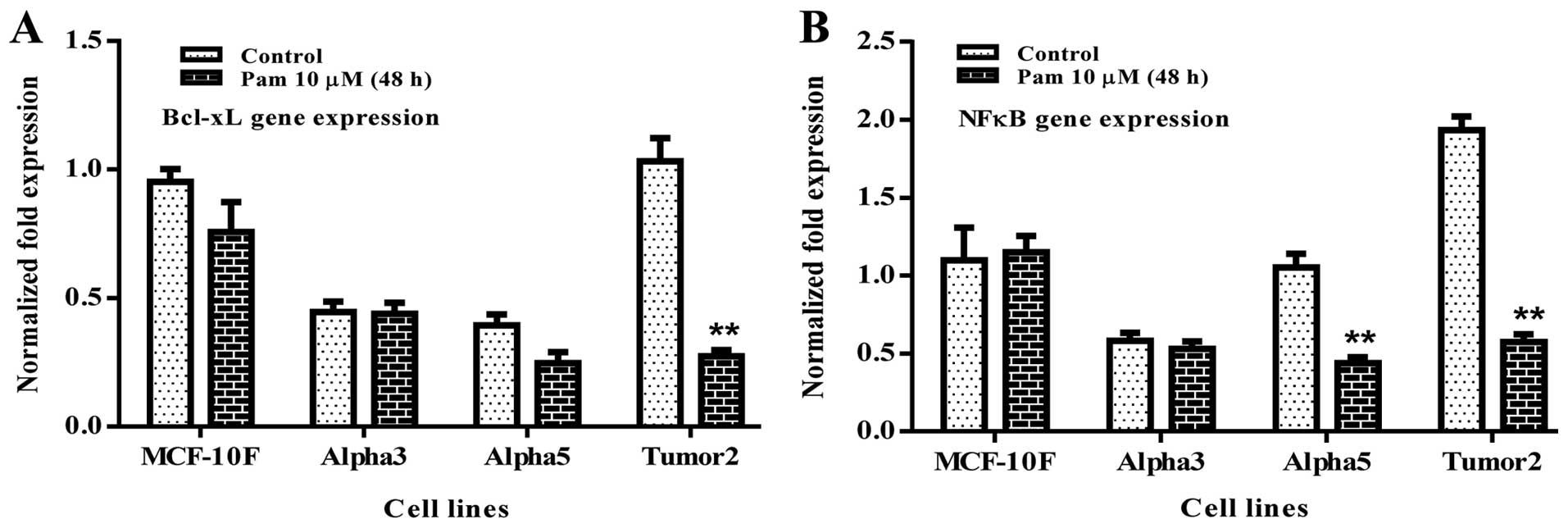
Alpha5 and Tumor2 showed 10.4 and 14.5%, respectively (Fig. 4). Results indicated that
Bcl-xL (p<0.01) and NFκB (p<0.01) were
significantly reduced in Tumor2 in comparison to its counterparts
(Fig. 5A and B).
Discussion
In the present study, the in vitro effects of
pamidronate in breast cancer cell lines were evaluated by several
parameters. Pamidronate showed direct antitumor and apoptotic
activities in breast cancer cell lines, which is in agreement with
results from previous studies (17), highlighting the role of signal
transduction pathways controlled by the Rho family of small GTPases
(18). Treatment of pamidronate
decreased Rho-A gene and protein expression in Tumor2, MCF7,
MDA-MB-231 in comparison to their counterparts. The inhibition of
Rho proteins might provide a possibility to reduce metastasis
through interference with this pathway according to invasion and
migration assays where pamidronate reduced their percentage in
Alpha5 and Tumor2 cell lines. Recent studies have indicated that
pamidronate induced apoptotic effects in myeloma cells in
vitro (19–27). The involvement of the inhibitory
effect of pamidronate on isoprenoid biosynthesis in induction of
apoptosis could be tested by using farnesol and geranylgeraniol to
circumvent the blockade of geraniol synthesis. Geranylgeraniol was
more potent in abolishing pamidronate-induced apoptosis than
farnesol (19). These authors
demonstrated that geranylgeraniol reduced apoptosis by
approximately 75%, suggesting geranylgeranylated proteins such as
Rho proteins were main target of the pamidronate effect.
Pamidronate decreased c-Ha-ras gene
expression in Tumor2 cell line in comparison to its counterpart.
Authors have reported that c-Ha-ras expression in myeloma cells was
valuable in predicting the therapeutic effects of pamidronate.
Others demonstrated that resistance to pamidronate may result from
low levels of GTPase-activating proteins, such as N-ras and H-ras
in tumor cells (20). Pamidronate
has been shown to be a highly effective inhibitor of human cell
proliferation by inactivating the Ras/ERK pathway since it induced
apoptosis by inhibiting farnesylproteins containing Ras protein
(20,21). The prenylation of monomeric
G-proteins such as the members of the Ras superfamily Rho proteins
was reduced by bisphos-phonate treatment (22,23).
It seems that effects of H-ras on cell motility appeared to be
through activation of a MAP kinase cascade, presumably via the Ras
effector Raf (24). Pamidronate
and zoledronic acid induced apoptosis and growth inhibition in
epidermoid cancer cells that occur together with depression of ras
signaling of ERK and Akt survival pathways (25).
The present results indicated that p53 gene
expression decreased by pamidronate in malignant and tumorigenic
cell line Tumor2 in comparison to its counterpart. Other authors
have indicated that pamidronate inhibited cell growth and induced
apoptosis in human melanoma cells in vitro (22). Susceptibility to pamidronate did
not correlate to CD95 ligand sensitivity or p53 mutational status.
Other studies (21) showed that
p-JNK diminished while p53 was unaltered following pamidronate
treatment, ruling out activation of apoptosis via these two
pathways.
Regarding the genes related to metastasis,
Serpin-1 and Caveolin-1 were evaluated where
pamidronate decreased their gene expression in Tumor2 cell line in
comparison to their counterparts. The mitogenic activity of the
C-terminal section of serpin A1 is localized within the last 26
amino acids (25). The activity of
the peptide was sensitive to PKC inhibitors and T24 phosphorylation
and resulted in increased activity in MCF7 cells. Serpin A1 plays a
key role in the neutralization of neutrophil elastase. Tumor growth
and metastasis of breast cancer cells could be enhanced at sites of
high proteolytic activity, in which there is an excess of serine
proteases over serpin A1 (24).
In vitro studies have shown that both stromal and epithelial
cav-1 play a protective role against mammary hyperplasia and
tumorigenesis in breast cancer (8,9).
Genes related to apoptosis such as Bcl-xL
decreased its gene expression of the malignant and tumorigenic
Tumor2 cell line by the effect of pamidronate. Another key positive
regulator of cancer cell proliferation and survival is NFκB
(12,13) which it has the ability to
transcriptionally activate many pro-survival and anti-apoptotic
genes such as Bcl-xL. It was observed that NFκB gene
expression was decreased in Tumor2 cell line in comparison to its
counterpart. It can be concluded that pamidronate may exert
antitumor activity in breast cancer cells transformed by low doses
of α-particles and estrogen in vitro by downregulating
Bcl-xL and NFκB gene expression.
Acknowledgements
The secretarial assistance of Leodán Crispin is
greatly appreciated. This study was supported by Grant support
FONDECYT #1120006 (G.M.C) and MINEDUC-UTA (G.M.C).
References
|
1
|
Hall A: The cellular functions of small
GTP-binding proteins. Science. 249:635–640. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ridley AJ and Hall A: The small
GTP-binding protein rho regulates the assembly of focal adhesions
and actin stress fibers in response to growth factors. Cell.
70:389–399. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ridley AJ, Paterson HF, Johnston CL,
Diekmann D and Hall A: The small GTP-binding protein rac regulates
growth factor-induced membrane ruffling. Cell. 70:401–410. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lane DP: Cancer p53, guardian of the
genome. Nature. 358:15–16. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gatto M, Iaccarino L, Ghirardello A, Bassi
N, Pontisso P, Punzi L, Shoenfeld Y and Doria A: Serpins, immunity
and autoimmunity: Old molecules, new functions. Clin Rev Allergy
Immunol. 45:267–280. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hnasko R and Lisanti MP: The biology of
caveolae: Lessons from caveolin knockout mice and implications for
human disease. Mol Interv. 3:445–464. 2003. View Article : Google Scholar
|
|
7
|
Witkiewicz AK, Dasgupta A, Sammons S, Er
O, Potoczek MB, Guiles F, Sotgia F, Brody JR, Mitchell EP and
Lisanti MP: Loss of stromal caveolin-1 expression predicts poor
clinical outcome in triple negative and basal-like breast cancers.
Cancer Biol Ther. 10:135–143. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sotgia F, Schubert W, Pestell RG and
Lisanti MP: Genetic ablation of caveolin-1 in mammary epithelial
cells increases milk production and hyper-activates STAT5a
signaling. Cancer Biol Ther. 5:292–297. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Williams TM, Sotgia F, Lee H, Hassan G, Di
Vizio D, Bonuccelli G, Capozza F, Mercier I, Rui H, Pestell RG, et
al: Stromal and epithelial caveolin-1 both confer a protective
effect against mammary hyperplasia and tumorigenesis: Caveolin-1
antagonizes cyclin D1 function in mammary epithelial cells. Am J
Pathol. 169:1784–1801. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Reed JC: Apoptosis mechanisms:
Implications for cancer drug discovery. Oncology (Williston Park).
18(Suppl 10): 11–20. 2004.
|
|
11
|
Riedl SJ and Shi Y: Molecular mechanisms
of caspase regulation during apoptosis. Nat Rev Mol Cell Biol.
5:897–907. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sethi G, Ahn KS and Aggarwal BB: Targeting
nuclear factor-kappa B activation pathway by thymoquinone: Role in
suppression of antiapoptotic gene products and enhancement of
apoptosis. Mol Cancer Res. 6:1059–1070. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dolcet X, Llobet D, Pallares J and
Matias-Guiu X: NF-κB in development and progression of human
cancer. Virchows Arch. 446:475–482. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shipman R, Schraml P, Moch H, Colombi M,
Sauter G, Mihatsch M and Ludwig C: p53 protein accumulation and p53
gene alterations (RFLP, VNTR and p53 gene mutations) in
non-invasive versus invasive human transitional bladder cancer. Int
J Oncol. 10:801–806. 1997.PubMed/NCBI
|
|
15
|
Donehower LA, Godley LA, Aldaz CM, Pyle R,
Shi YP, Pinkel D, Gray J, Bradley A, Medina D and Varmus HE: The
role of p53 loss in genomic instability and tumor progression in a
murine mammary cancer model. Prog Clin Biol Res. 395:1–11.
1996.PubMed/NCBI
|
|
16
|
Calaf GM and Hei TK: Establishment of a
radiation- and estrogen-induced breast cancer model.
Carcinogenesis. 21:769–776. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wada A, Fukui K, Sawai Y, Imanaka K, Kiso
S, Tamura S, Shimomura I and Hayashi N: Pamidronate induced
anti-proliferative, apoptotic, and anti-migratory effects in
hepatocellular carcinoma. J Hepatol. 44:142–150. 2006. View Article : Google Scholar
|
|
18
|
Clemons MJ, Dranitsaris G, Ooi WS,
Yogendran G, Sukovic T, Wong BY, Verma S, Pritchard KI, Trudeau M
and Cole DE: Phase II trial evaluating the palliative benefit of
second-line zoledronic acid in breast cancer patients with either a
skeletal-related event or progressive bone metastases despite
first-line bisphosphonate therapy. J Clin Oncol. 24:4895–4900.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Riebeling C, Forsea AM, Raisova M, Orfanos
CE and Geilen CC: The bisphosphonate pamidronate induces apoptosis
in human melanoma cells in vitro. Br J Cancer. 87:366–371. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang PL, Lun M, Siegelmann-Danieli N,
Blasick TM and Brown RE: Pamidronate resistance and associated low
ras levels in breast cancer cells: A role for combinatorial
therapy. Ann Clin Lab Sci. 34:263–270. 2004.PubMed/NCBI
|
|
21
|
Zhang PL, Quiery AT Jr, Blasick TM and
Brown RE: Morpho-proteomic expression of H-ras (p21ras) correlates
with serum monoclonal immunoglobulin reduction in multiple myeloma
patients following pamidronate treatment. Ann Clin Lab Sci.
37:34–38. 2007.PubMed/NCBI
|
|
22
|
Nguyen DH, Catling AD, Webb DJ, Sankovic
M, Walker LA, Somlyo AV, Weber MJ and Gonias SL: Myosin light chain
kinase functions downstream of Ras/ERK to promote migration of
urokinase-type plasminogen activator-stimulated cells in an
integrin-selective manner. J Cell Biol. 146:149–164. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oxford G and Theodorescu D: Ras
superfamily monomeric G proteins in carcinoma cell motility. Cancer
Lett. 189:117–128. 2003. View Article : Google Scholar
|
|
24
|
Caraglia M, D’Alessandro AM, Marra M,
Giuberti G, Vitale G, Viscomi C, Colao A, Prete SD, Tagliaferri P,
Tassone P, et al: The farnesyl transferase inhibitor R115777
(Zarnestra) synergistically enhances growth inhibition and
apoptosis induced on epidermoid cancer cells by Zoledronic acid
(Zometa) and Pamidronate. Oncogene. 23:6900–6913. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Congote LF and Temmel N: The C-terminal
26-residue peptide of serpin A1 stimulates proliferation of breast
and liver cancer cells: Role of protein kinase C and CD47. FEBS
Lett. 576:343–347. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aparicio A, Gardner A, Tu Y, Savage A,
Berenson J and Lichtenstein A: In vitro cytoreductive effects on
multiple myeloma cells induced by bisphosphonates. Leukemia.
12:220–229. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Senaratne SG, Pirianov G, Mansi JL, Arnett
TR and Colston KW: Bisphosphonates induce apoptosis in human breast
cancer cell lines. Br J Cancer. 82:1459–1468. 2000. View Article : Google Scholar : PubMed/NCBI
|