Introduction
Immunotoxins, consisting of a plant or bacterial
toxin coupled to an antibody or antibody fragment, selectively kill
tumor cells by specifically recognizing molecular abnormalities on
the malignant cell surface. A well-studied toxin is a truncated
version of exotoxin A from Pseudomonas aeruginosa (PE38),
where the natural cell-binding domain (1a) has been deleted
(1). It delivers its cytotoxic
effect in the cytosol by ribosylating elongation factor 2 (EF-2),
preventing proper ribosomal function, which leads to cell death
(1). Immunotoxins, including
immunoglobulin-based targeting domains in various formats, such as
Fv- and scFv of IgG, interacting with different cell surface
receptors, have been found efficacious in pre-clinical as well as
clinical studies (2). PE38 has
also been coupled to targeting domains of non-immunoglobulin origin
including designed ankyrin repeat proteins (3) and affibody molecules (4,5) and
such fusion toxins has been found efficacious in pre-clinical
models of cancer. PE38-based toxins are immunogenic, preventing
multiple repeated injections, thus limiting the suitability in
cancer treatment regimens. Recently, deimmuniztion efforts to
remove B- and T-cell epitopes have been undertaken (6–10),
which has successfully identified less immunogenic variants such as
PE38X8 (7). Such toxins may
potentially be more suited for clinical use.
The human epidermal growth factor receptor 2 (HER2)
is a tyrosine kinase receptor that is often overexpressed in many
types of cancer, including breast carcinoma (11). Overexpression of HER2 has been
found to correlate with a poor patient prognosis and early relapse
post-surgery (12). Since the
expression levels of HER2 on normal tissues are relatively low,
this receptor has been used to differentially target HER2-positive
tumors. Current treatment regimens for advanced HER2-positive
breast cancer may therefore include the HER2-specific antibody
trastuzumab, marketed as Herceptin (13). Recently, an antibody-drug conjugate
where trastuzumab has been functionalized with the toxin emtansine
(T-DM1) has also been approved for clinical treatment of patients
with advanced HER2-positive breast cancer (14). Thus far, immunotoxins targeting
HER2-positive tumors has not reached clinical use; however, based
on the success of trastuzumab and T-DM1, it is likely that such
immunotoxins could be developed if short-comings related to, for
example immunogenicity can be overcome.
Affibody molecules are a class of folded,
non-immunoglobulin based affinity ligands based on the triple
helical B-domain of staphylococcal protein A (15). They are only 58 amino acids long
and devoid of cysteins in their framework. Affibody molecules
interacting specifically and with high affinity (in the low
nanomolar to picomolar range) with several tumor cell markers
including IGF-1R, EGFR, HER2 and HER3 have been described (16–19).
In particular, the affibody molecule ZHER2:2891 have
been found to specifically interact with the HER2 receptor with an
equilibrium dissociation constant (KD) of 60 pM
(20). ZHER2:2891 is a
derivative of ZHER2:342 (18) that has increased melting point,
stability, and overall hydrophilicity. The suitability of
ZHER2:2891 for use in man has been documented by a
clinical study where HER2-positive tumors were visualized by
radionuclide molecular imaging (21). When used for radionuclide molecular
imaging, the small size of the affibody molecules enable very rapid
clearance of unbound tracers. This provides a low background for
the affibody-based imaging. A short residence time in circulation
for affibody-based fusion toxins has similarly been shown, but in
that case necessitated multiple injections (5). Thus, an extension of in vivo
half-life of affibody-based fusion toxins would provide a clinical
benefit. Streptococcal Protein G includes three motifs with high
affinity for albumin from several species (22). One of them, the GA148-GA3 domain,
consists of 46 amino acids and has a cysteine-free three-helix
structure. This albumin binding domain (ABD) has been engineered to
yield variants such as ABD035 with femtomolar affinity
for human serum albumin as well as high affinity for serum albumin
from rat, mouse and cynomolgus monkey (23). Inclusion of an albumin binding
domain in a fusion protein causes a strong non-covalent interaction
with serum albumin when injected into the blood (24,25).
This has been shown to increase the serum half-life, both by
increasing the molecular weight of the complex and by allowing
serum albumin-mediated interaction with the neonatal Fc receptor
(FcRn), leading to rescue from lysosomal degradation (26,27).
To further extend the potential of a deimmunized and
truncated version of Pseudomonas exotoxin A (PE38X8) for
targeted strategies, a novel tripartite fusion toxin, including
PE38X8 fused to the HER2-binding affibody molecule
ZHER2:2891 and the half-life extension albumin binding
domain ABD035 was evaluated. The interactions with HER2
and serum albumins from different species were analyzed and the
cytotoxic potential was determined.
Materials and methods
General
All chemicals were from Merck (Darmstadt, Germany)
or Sigma-Aldrich (St. Louis, MO, USA) unless otherwise noted. DNA
modifying and restriction enzymes were from New England Biolabs
(Ipswich, MA, USA).
Gene construction
An expression vector encoding
His6-ZHER2:342-PE38 (4) was a kind gift from Jacek Capala. The
gene enoding ZHER2:2891-ABD-PE38X8 with the N-terminal
amino acid sequence HEHEHE and (S4G)3 linkers
connecting the three domains in the expression vector pET-26b(+)
(Merck) was obtained from GenScript USA Inc. (Piscataway, NJ, USA).
The construct was fitted with NdeI and BamHI
restriction sites surrounding ZHER2:2891 and two
NcoI restriction sites surrounding the ABD domain. The
albumin binding domain (ABD) used was ABD035, a version
engineered for high affinity to human serum albumin (23). PE38X8 was a deimmunized version of
PE38 with the following amino acid alterations: R313A, Q332S,
R432G, R467A, R490A, R513A, E548S, K590S (7). The expression vector for
ZTaq-ABD-PE38X8 was created by PCR amplification of
ZTaq (28) followed by
replacement with ZHER2:2891 in pET-26b(+) encoding
ZHER2:2891-ABD-PE38X8, using the NdeI and
BamHI restriction sites. Expression vectors for
ZHER2:2891-PE38X8 and ZTaq-PE38X8 were
created by digestion of ZHER2:2891-ABD-PE38X8 and
ZTaq-ABD-PE38X8, respectively, with NcoI followed
by religation of the vectors. All constructs were verified by
DNA-sequencing.
Protein expression and purification
Escherichia coli [Rosetta (DE3) pLysS]
(Merck) was used for expression of the fusion toxins essentially
according to the manufacturer’s protocol. Cells harboring the
expression plasmids were grown at 37°C until OD600
reached 1.5 after which protein expression was induced by addition
of Isopropyl β-D-1-thiogalactopyranoside (IPTG) to a final
concentration of 1 mM. Protein expression was carried out for 2.5 h
after which the cells were harvested by centrifugation and
resuspended in 20 ml IMAC-loading buffer (300 mM NaCl, 50 mM
Na-phosphate, pH 7.0) supplemented with Complete EDTA-free protease
inhibitor cocktail (Roche Diagnostic, Basel, Switzerland). The
cells were broken by sonication and the fusion toxins were purified
from the supernatants by immobilized metal-ion affinity
chromatography (IMAC) on a Ni-Sepharose 6 Fast Flow resin (GE
Healthcare, Uppsala, Sweden) under native conditions with Imidazole
elution according to the manufacturer’s protocol. Eluted material
was pooled and the buffer was changed to 20 mM Bis-Tris (pH 6.5)
using PD-10 columns (GE Healthcare). The fusion toxins were further
purified by anion exchange chromatography on 1 ml HiTrap Q HP
columns (GE Healthcare) using 20 mM Bis-Tris (pH 6.5) as running
buffer. Bound material was eluted by a NaCl-gradient from 0 to 0.6
M. Eluted material was pooled and further purified by gel
filtration on a Superdex 75 column (GE Healthcare) with
phosphate-buffered saline (PBS) as running buffer. The molecular
masses of the fusion toxins, previously alkylated with
2-iodoacetamide, were determined by liquid chromatography
electrospray ionization mass spectrometry (Agilent Technologies,
Santa Clara, CA, USA). Purified, free targeting domain
(ZHER2:342), was a kind gift from Lisa Sandersjöö and
John Löfblom (29).
Biosensor analysis
A Biacore 3000 instrument (GE Healthcare) was used
for biosensor analysis. The extracellular domain of HER2
(HER2ECD) (Sino Biological, Beijing, China) was
immobilized on a CM5-chip by amine coupling in 50 mM sodium acetate
buffer (pH 4.5). On a second CM5-chip, HSA, (Novozymes, Bagsvaerd,
Denmark), MSA (Sigma-Aldrich), and BSA (Merck) were immobilized in
the same way. The final immobilization levels of
HER2ECD, HSA, MSA and BSA were 202, 869, 584 and 779 RU,
respectively. Reference flow cells were created on both chips by
activation and deactivation. HBS-EP [10 mM HEPES, 150 mM NaCl, 3 mM
EDTA, 0.05% Tween-20, (pH 7.4)] was used as running buffer and for
dilution of the analytes. All experiments were performed at 25°C
with a flow-rate 50 μl/min. On- and off-rates were determined by
BIAevaluation 4.1 software using a 1:1 Langmuir binding model.
Cell lines
The SKOV-3 and SKBR-3 cell lines were obtained from
American Type Culture Collection (ATCC) through LGC Standards
(Wesel, Germany) and were grown in McCoy’s 5A supplemented with 10%
fetal bovine serum in a humidified incubator at 37°C in 5%
CO2 atmosphere. The A549 cell line was also obtained
from ATCC and was grown in Dulbecco’s modified Eagle’s medium
supplemented with 10% fetal bovine serum at the same conditions.
HER2 expression levels were determined by incubating
2×106 cells with Trastuzumab (Roche) (5 μg/ml) as
primary antibody for 45 min followed by Alexa Fluor 488 conjugated
goat anti-human antibody (Life Technologies, Carlsbad, CA, USA) (5
μg/ml) as secondary antibody for 45 min. The cells were
subsequently analyzed on a Gallios flow cytometer (Beckman Coulter,
Brea, CA, USA). 10,000 events were recorded and analyzed by Kaluza
software (Beckman Coulter).
Cytotoxicity measurements
Approximately 5,000 cells/well were seeded in a
96-well microtiter plate and were allowed to attach overnight.
Subsequently, the medium was replaced with fresh medium containing
fusion toxins and other proteins followed by incubation for 72 h
unless noted otherwise. Cell viability was determined using a cell
counting kit-8 (CCK-8) (Sigma-Aldrich) according to manufacturer’s
protocol with determination of A450 in each well. The
obtained absorbance values were analyzed by GraphPad Prism
(GraphPad Software, La Jolla, CA, USA).
Results
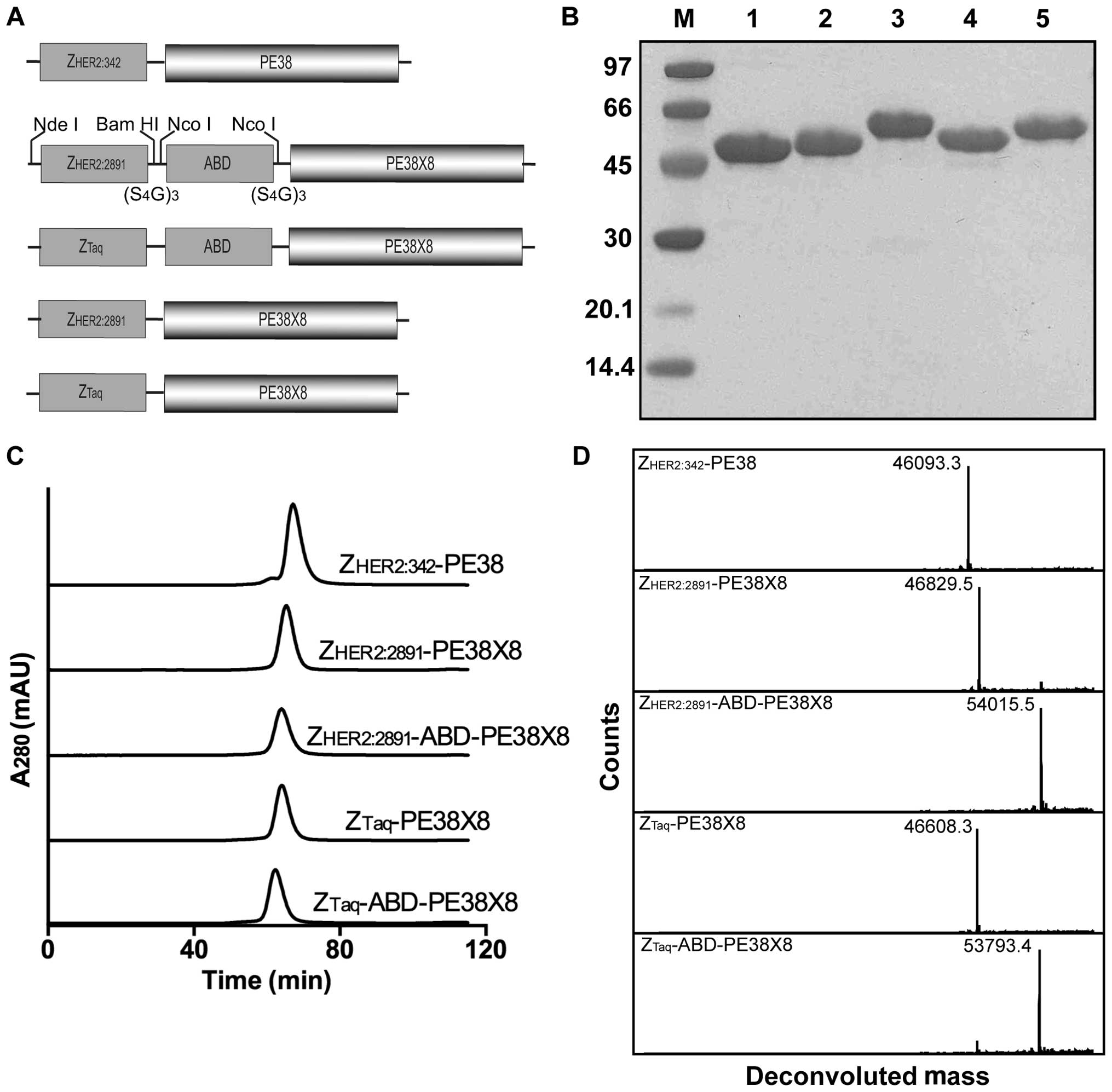
Construction of fusion toxins
Four fusion toxins were investigated in this study,
consisting of a deimmunized variant of PE38 coupled to an affibody
molecule binding to human epidermal growth factor receptor-2
(ZHER2:2891) or to a control affibody molecule not
expected to interact with any human protein (ZTaq) and
with or without the half-life extension albumin binding domain
(ABD) (Fig. 1A). A tag with the
amino acid sequence HEHEHE was added to the N-terminus to allow
purification by immobilized metal-ion affinity chromatography
(IMAC) (30). The characteristics
of the fusion toxins were compared to ZHER2:342-PE38, a
fusion toxin consisting of the wild-type PE38 domain coupled to
ZHER2:342, a predecessor to ZHER2:2891, with
an N-terminal His6-tag.
Production and initial characterization
of the fusion toxins
The proteins were expressed in Escherichia
coli followed by purification using three sequential
chromatographic steps: IMAC, anion exchange chromatography and gel
filtration. The chromatograms recorded during the gel filtration
are displayed in Fig. 1C and show
single, essentially symmetrical peaks, indicating the absence of
multimer formation. Proteins eluted after the final gel filtration
step were analyzed by SDS-PAGE, showing pure proteins with the
expected molecular weights (Fig.
1B). More accurate measurements of the molecular masses were
obtained by mass spectrometry and the results showed the expected
molecular masses with <2 Da error (Fig. 1D and Table I).
 | Table ICharacterization of fusion
toxins. |
Table I
Characterization of fusion
toxins.
| Theoretical MW
(Da) | Detected MW
(Da) | Measured
KD (nM)a |
|---|
|
ZHER2:342-PE38 | 45,978.1 | 45,979.2 | 2b |
|
ZHER2:2891-PE38X8 | 46,714.6 | 46,715.4 | 5 |
|
ZHER2:2891-ABD-PE38X8 | 53,899.6 | 53,901.4 | 5 |
|
ZTaq-PE38X8 | 46,493.3 | 46,494.2 | NBc |
|
ZTaq-ABD-PE38X8 | 53,678.3 | 53,679.3 | NB |
Biosensor analysis of the interaction
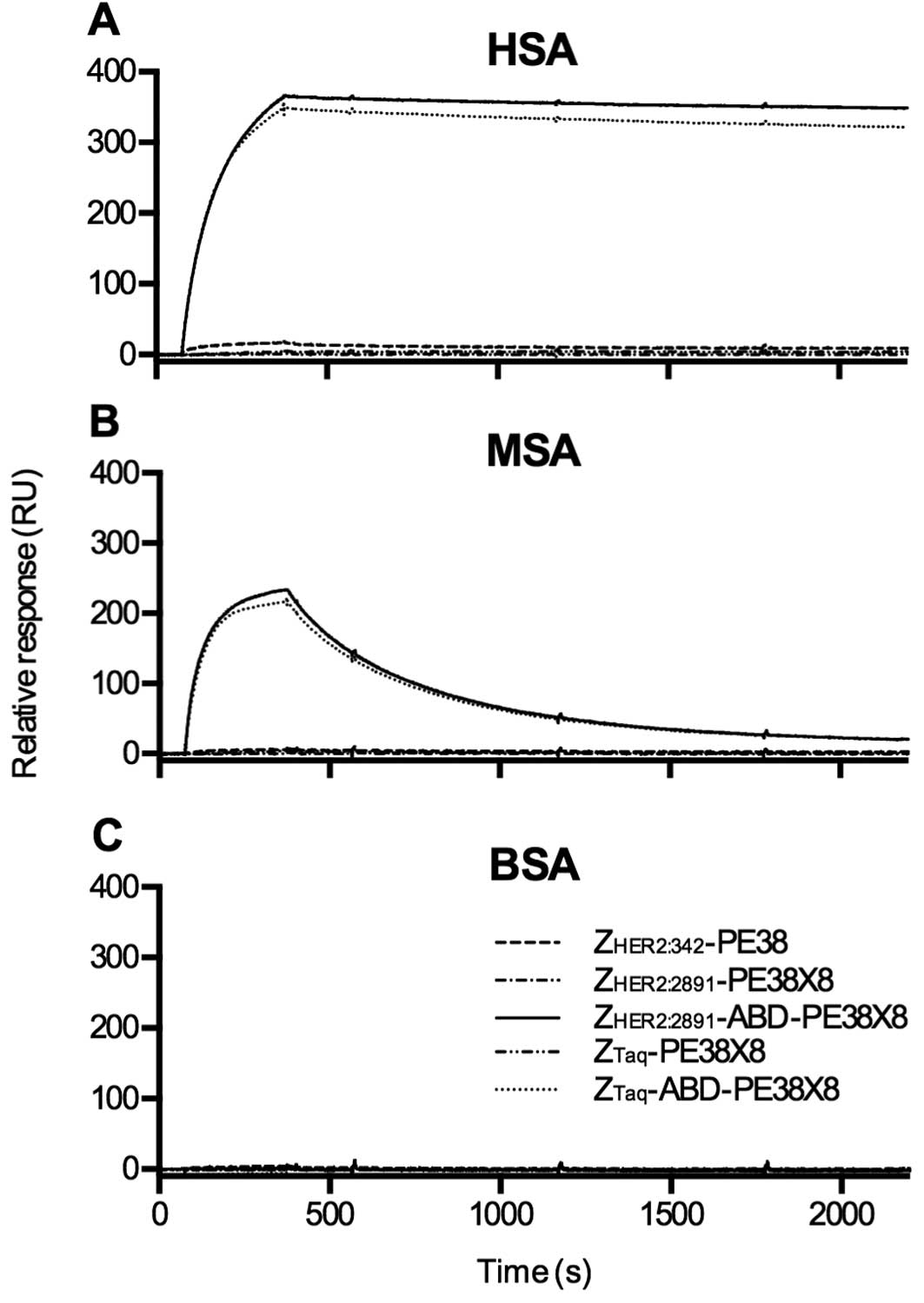
between the fusion toxins and serum albumins
The inclusion of an ABD in the fusion toxins
potentially allowed interaction with serum albumin. To investigate
the interaction with serum albumin from different species, the
fusion toxins were injected over flow cells with immobilized HSA,
MSA or BSA. The two toxins including the ABD interacted with HSA
and to a lesser extent MSA (Fig. 2A
and B). They did not interact with BSA (Fig. 2C). As expected, the three fusion
toxins lacking the ABD did not interact with any of the serum
albumins.
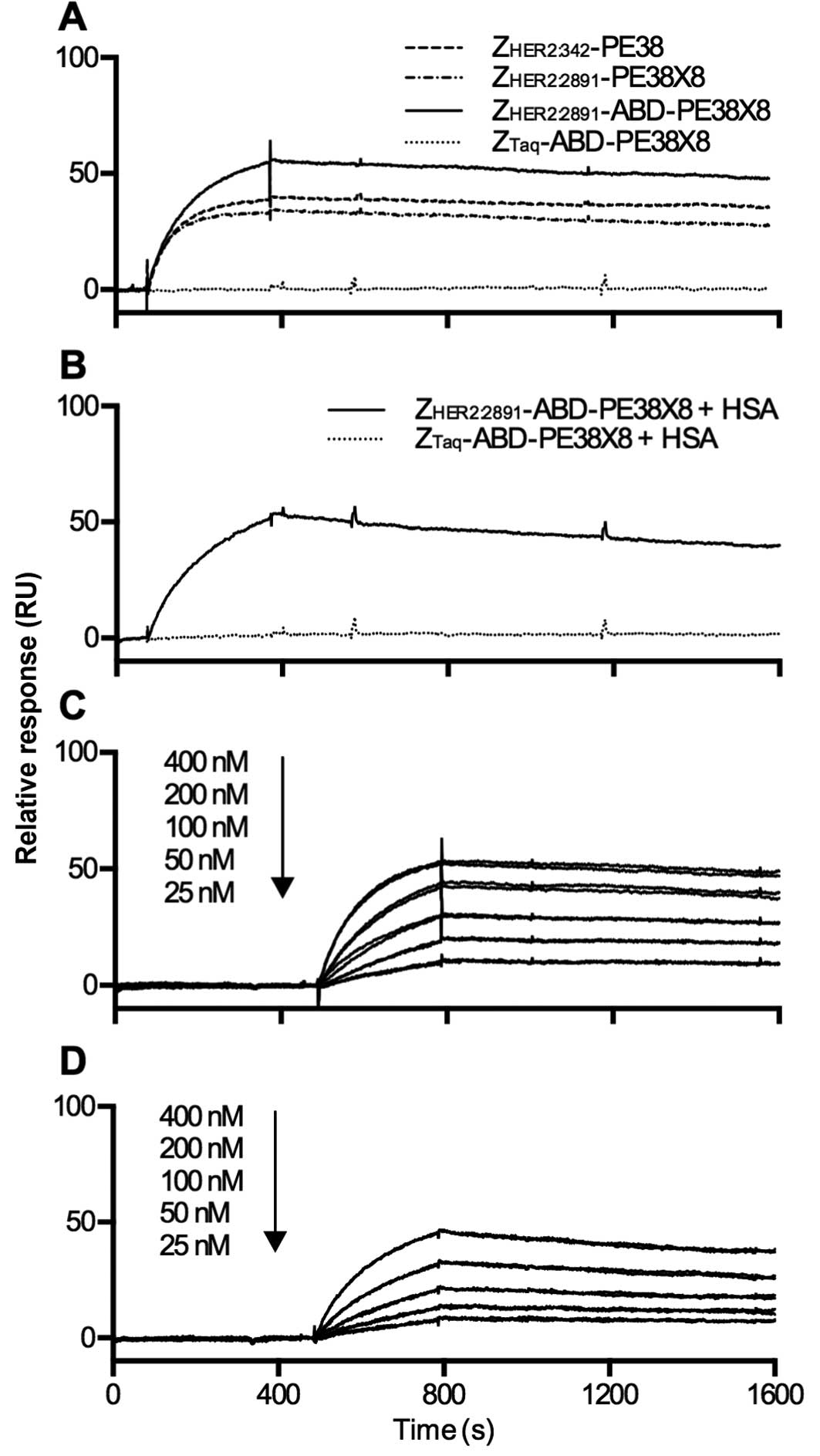
Biosensor analysis of the interaction
between the fusion toxins and HER2
Fig. 3A displays an
overlay of sequential injections of the three
ZHER2-containing and one ZTaq-containing
(control) fusion toxins over a flow cell with immobilized
HER2ECD. As expected, the three fusion toxins which
contain ZHER2:342 or ZHER2:2891 interact with
HER2 and the control does not. To investigate a possible
interference in the fusion toxin/HER2 interaction, when the fusion
toxin is in complex with serum albumin,
ZHER2:2891-ABD-PE38X8 was also injected over the
HER2-flow cell after pre-incubation with an excess of HSA (Fig. 3B). ZHER2:2891-ABD-PE38X8
was found to interact with HER2 while in a complex with HSA. As a
control, ZTaq-ABD-PE38X8 was also pre-incubated with HSA
and injected over the flow cell with immoblilized HER2, which
showed no interaction. The affinity between
ZHER2:2891-ABD-PE38X8 and HER2 was determined in the
absence and presence of HSA by injecting serial dilutions of the
fusion toxins (Fig. 3C and D). The
resulting equilibrium dissociation constants were 5 and 12 nM in
the absence or presence of HSA. In addition, the affinity between
ZHER2:2891-PE38X8 and HER2 was determined similarly and
the equilibrium dissociation constant was found to be 5 nM.
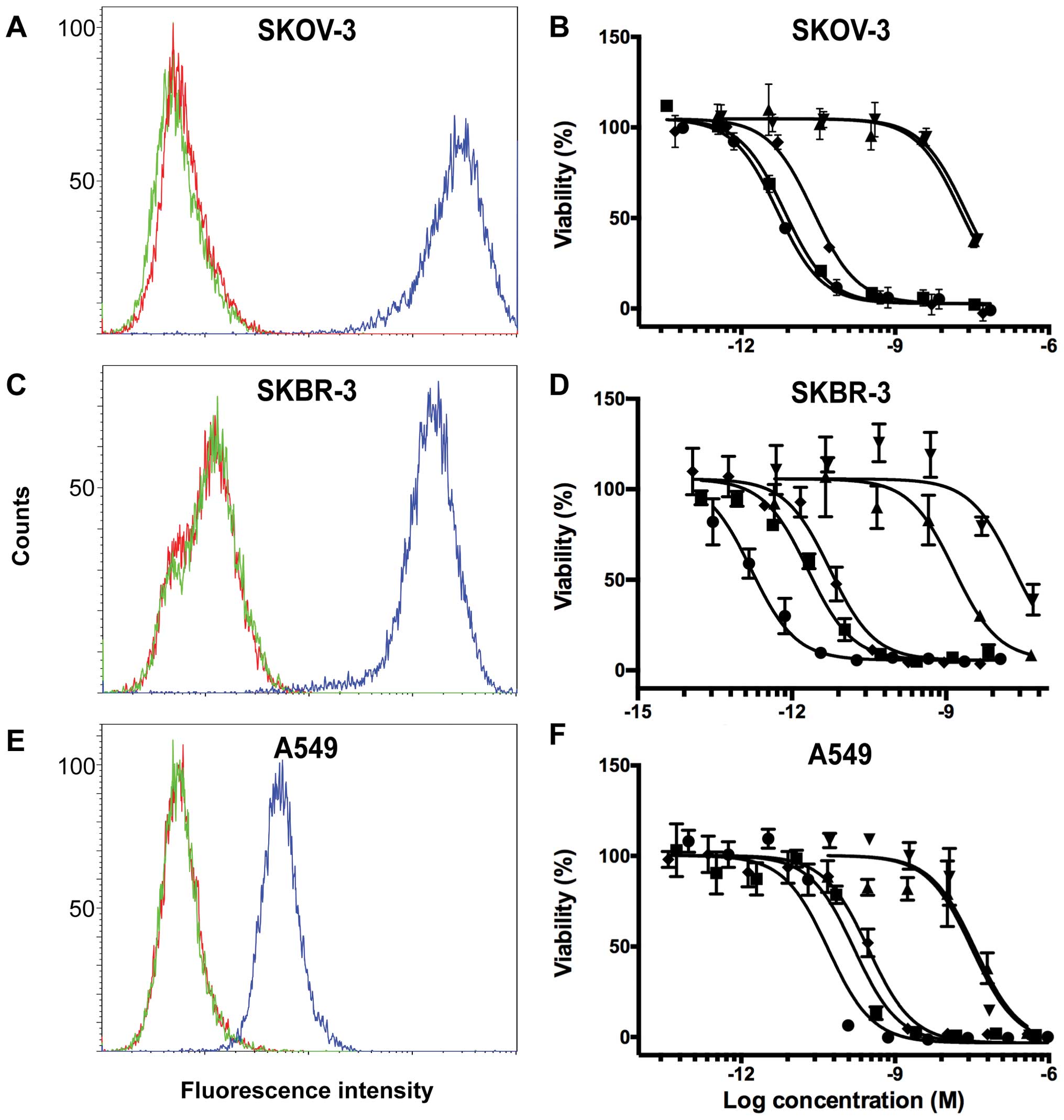
Determination of the cytotoxicity of the
fusion toxins on cells expressing different levels of HER2
The relative expression levels of HER2 on SKOV-3,
SKBR-3 and A549 cells were determined by flow cytometry. SKOV-3 and
SKBR-3 have relatively high expression levels of HER2 in contrast
to A549, which has a moderate expression level (Fig. 4A, C and E). To determine the
cytotoxicity of the fusion toxins, the cell lines were incubated
with serial dilutions of the fusion toxins for 72 h, after which
cell viability was measured and plotted as a function of the
logarithm of the toxin concentration (Fig. 4B, D and F). IC50 values
were determined from the viability plots and are displayed in
Table II. On SKOV-3 cells, the
two fusion toxins including ZHER2:2891 and the control
including ZHER2:342, had IC50 values ranging
between 5 and 25 pM which were 4,000–1,000 times lower than the
IC50 values of the control fusion toxins including
ZTaq. The IC50 values of
His6-ZHER2:342-PE38 and
ZHER2:2891-PE38X8 were similar and 5-fold lower than the
IC50 value of ZHER2:2891-ABD-PE38X8,
indicating that inclusion of ABD lowers the cytotoxicity somewhat.
SKBR-3 cells have a similar density of HER2 on the cell surface and
similar IC50 values for the fusion toxins were measured
on this cell line. Here, a slightly lower IC50 value was
measured for His6-ZHER2:342-PE38 compared to
ZHER2:2891- PE38X8, indicating that PE38X8 is slightly
less cytotoxic compared to PE38. Since a similar decrease in
cytotoxicity was not found on SKOV-3 cells, the result suggests
that the difference is dependent on the cell line. The A549 cell
line has a lower HER2 density than SKOV-3 and SKBR-3 and
consequently the IC50 values for the fusion toxins
including ZHER2:2891 or ZHER2:342 was higher,
ranging from 50 to 300 pM. The IC50 values for the
control fusion toxins including ZTaq, were similar for
A549, SKOV-3 and SKBR-3, except for ZTaq-ABD-PE38X8 on
SKBR-3 cells, which was lower. In combination, these results
indicate that the fusion toxins interacting with HER2 are more
cytotoxic to cells with a higher surface density of HER2 than cells
with a lower, further proving that the fusion toxins are HER2
specific.
| Table IICytotoxicity of fusion toxins. |
Table II
Cytotoxicity of fusion toxins.
| | IC50
(pM) |
|---|
| |
|
|---|
| Cell line | HER2 expression
level |
ZHER2:342-PE38 |
ZHER2:2891-PE38X8 |
ZHER2:2891-ABD-PE38X8 |
ZTaq-PE38X8 |
ZTaq-ABD-PE38X8 |
|---|
| SKBR-3 | High | 0.16
(0.12–0.23)a | 2.1 (1.5–2.9) | 5.6 (4.0–7.8) | 22,000
(14,000–32,000) | 1,400
(900–2,100) |
| SKOV-3 | High | 5.1 (4.1–6.4) | 6.9 (5.5–8.8) | 25 (20–32) | 24,000
(19,000–30,000) | 19,000
(15,000–24,000) |
| A549 | Moderate | 50 (35–71) | 160 (110–230) | 300 (210–430) | 32,000
(22,000–45,000) | 37,000
(26,000–52,000) |
Analysis of toxin specificity
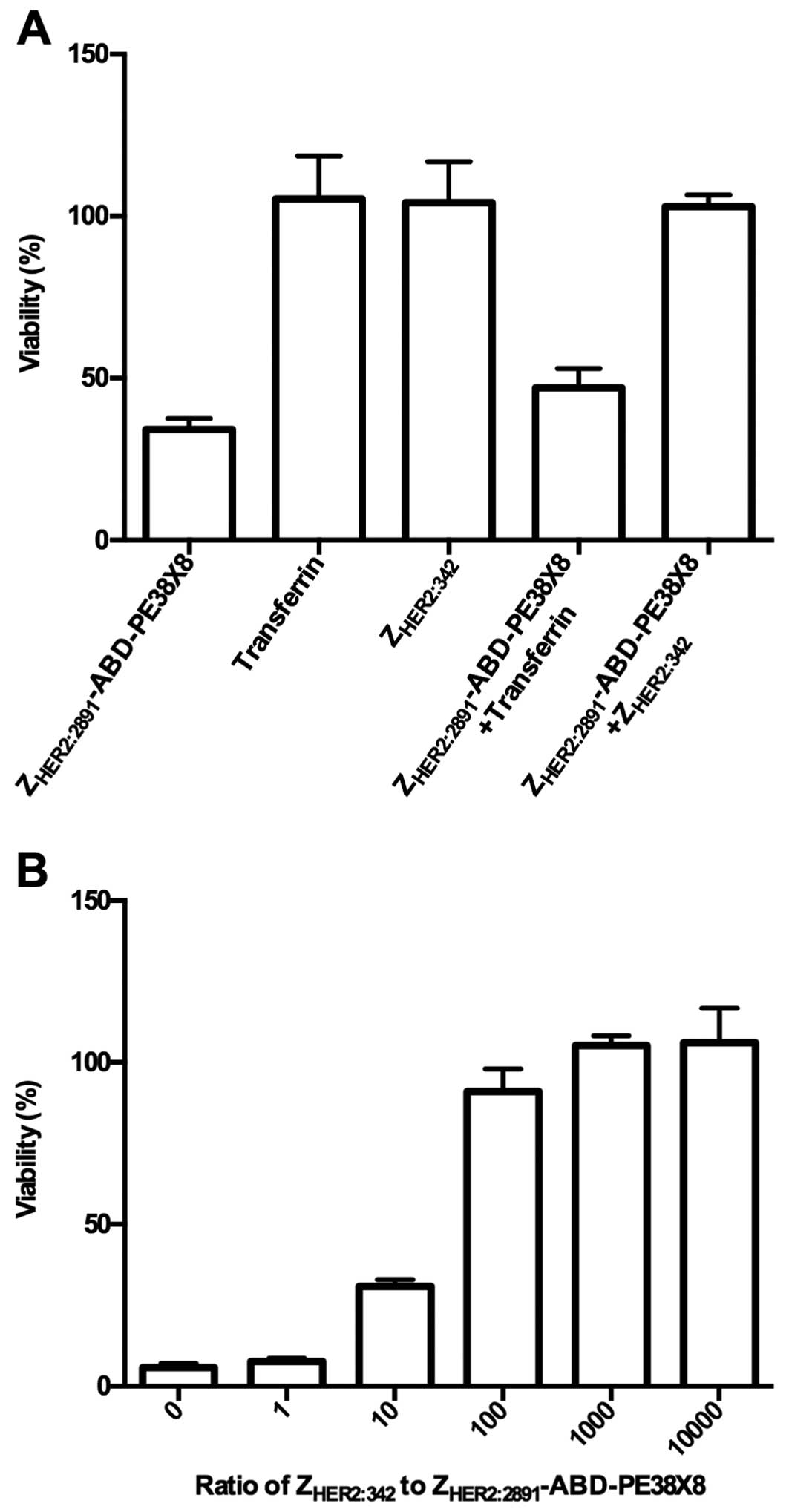
To further investigate the specificity of the fusion
toxins, SKOV-3 cells were incubated with
ZHER2:2891-ABD-PE38X8 after the cells had been
pre-incubated with an excess of free targeting domain
(ZHER2:342) which was expected to block available HER2
receptors on the cells, or with an excess of control protein
(transferrin), which was not expected to interact with the HER2
receptor. Fig. 5A shows that
ZHER2:342 and transferrin does not affect cell viability
by themselves. However, pre-incubation of the cells with
ZHER2:342 rescues the cells from
ZHER2:2891-ABD-PE38X8 cytotoxicity. Pre-incubation with
the control protein transferrin does not rescue cells similarly,
showing that the cytotoxic potential of
ZHER2:2891-ABD-PE38X8 is HER2-dependent. The cytotoxic
potential of ZHER2:2891-ABD-PE38X8 in combination with
the free targeting domain ZHER2:342 was further
investigated by pre-incubating SKOV-3 cells with increasing
concentrations of ZHER2:342 followed by addition of
ZHER2:2891-ABD-PE38X8. The results showed that cell
viability increases with increasing concentration of
ZHER2:342 (Fig. 5B),
further corroborating the finding that the cytotoxicity of
ZHER2:2891-ABD-PE38X8 is dependent on the level of free
HER2 on the cell surface.
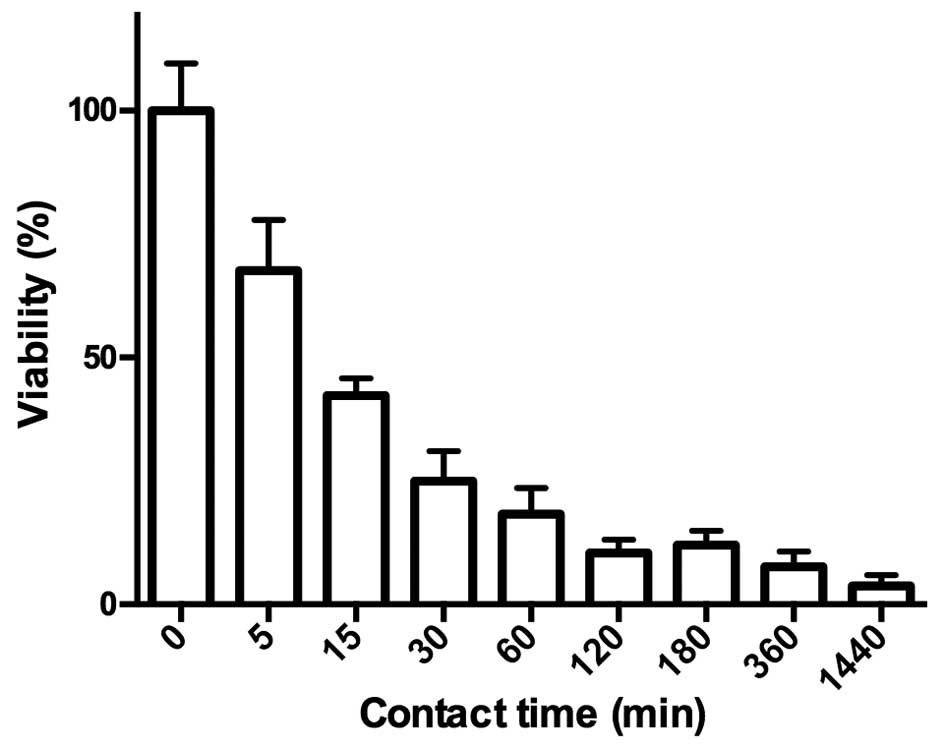
Analysis of the effect of contact time on
cytotoxicity
To investigate the influence of contact time on
cytotoxicity, SKOV-3 cells were incubated for different amounts of
time with a concentration of ZHER2:2891-ABD-PE38X8
expected to reduce cell viability close to 0 after incubation for
72 h, followed by measurement of cell viability. The results are
plotted in Fig. 6 and show that
cell viability is reduced with increasing exposure time. The
longest exposure time of 1,440 min (24 h) reduces cell viability
close to 0. A 50% reduction of cell viability is found after
exposure for 10 min.
Discussion
The use of immunotoxins for treatment of cancer has
been hampered partly by the immunogenicity of the toxin part, which
may lead to formation of neutralizing antibodies after only few
injections (31). Deimmunized
toxins such as PE38X8, where mouse B-cell epitopes have been
removed, may be more suited for immunotoxin construction since
formation of neutralizing antibodies is significantly lower while
the potency of the toxin is intact (7). However, for use in humans, further
engineered versions, where for example also human B-cell epitopes
have been removed, could prove even more useful, despite their
lower potency (8). Deimmunized
PE38-variants, such as PE38X8, have not yet been evaluated in
combination with non-immunglobulin based targeting domains and one
of the goals of this study was to compare PE38X8 with PE38 using a
HER2-interacting affibody molecule (ZHER2:324 or the
derivative ZHER2:2891) as targeting domain. The results
showed that ZHER2:2891-PE38X8 had a comparable toxicity
to ZHER2:342-PE38 on SKOV-3 cells but was slightly less
toxic for SKBR-3 and A549 cells. In a previous study, PE38X8 was
found to be slightly more toxic than PE38 to Raji cells (7). Thus, the difference in toxicity of
PE38X8 and PE38 appears to be cell line-dependent. Inclusion of an
albumin binding domain (ABD) for half-life extension also slightly
lowered the cytotoxicity on all three cell lines. The tendency of
lower toxicity of ZHER2:2891-ABD-PE38X8 and
ZHER2:2891-PE38X8 is possibly a consequence of their
slightly lower affinity for HER2 (5 nM) in comparison with
ZHER2:342-PE38 (2 nM). The lower affinity is in turn a
consequence of the lower affinity between HER2 and
ZHER2:2891 (60 nM) in comparison with HER2 and
ZHER2:342 (22 pM) (18,20).
The affinity of ZHER2:2891-ABD-PE38X8 was
200-fold weaker compared to the affinity between
ZHER:2891 and HER2 (20). A similar reduction in affinity has
previously been reported for ZHER2:342-PE38 (4). Despite this reduction, the difference
in cytotoxic potency of ZHER2:2891-ABD-PE38X8 and the
control ZTaq-ABD-PE38X8 on SKOV-3 and SKBR3 cells, which
express high levels of HER2, was 1,000-fold which suggests a rather
wide therapeutic window.
The half-life of ZHER2:342-PE38 was
previously found to be only 9 min (5), which is less than the contact time
needed to kill 50% of cells in vitro (4). Inclusion of an ABD in the current
construct is likely to increase the serum half-life significantly,
since a strong interaction with both human and mouse serum albumin
was detected (Fig. 2). A longer
serum half-life may lead to a higher in vivo efficacy,
although a longer serum half-life also gives the immune system a
longer time to interact with the fusion toxin to form neutralizing
antibodies. An important issue is that interaction with the
comparatively large serum albumin does not impart HER2-binding and
as a consequence cytotoxicity. Interestingly,
ZHER2:2891-ABD-PE38X8 appeared to be able to interact
with HER2 while in a complex with HSA (Fig. 3B). This suggests that
ZHER2:2891-ABD-PE38X8 should be cytotoxic also when in
complex with serum albumin.
Off-target toxicity has been an obstacle when
developing immunotoxins and fusion toxins. A particular problem
with early constructs including Pseudomonas exotoxin A was
damage to the vasculature, which set the limiting dose (32). Fortunately, with the development of
PE38, damage to the vasculature has been reduced. Also of
importance is that the construct does not unspecifically accumulate
in healthy organs. In a study by Zielinski et al (5), where ZHER2:342-PE38 was
used to treat xenografted tumors in mice, some unspecific
accumulation of the fusion toxin in the liver was observed. A
combination of free ZHER2:342 with an N-terminally
placed His6-tag, as in ZHER2:342-PE38, has
previously been found to lead to unspecific uptake in the liver
(30). A solution was to modify
the His6-tag to the amino acid sequence HEHEHE, which
was found to reduce liver accumulation significantly. In the
currently investigated fusion toxin,
ZHER2:2891-ABD-PE38X8, a HEHEHE-tag was placed in the
N-terminus instead of the His6-tag present in
ZHER2:342-PE38. Even though half of the histidines were
substituted with glutamic acid, ZHER2:2891-ABD-PE38X8
could still be efficiently purified by IMAC. It is possible that
this substitution may lead to lower unspecific uptake in the liver
in future in vivo studies.
In conclusion, a tripartite fusion toxin based on a
deimmunized version of PE38 was successfully constructed. All three
parts were found to function properly. Future experiments will
reveal if this fusion toxin also have the ability to kill
HER2-expressing malignant cells in vivo.
Acknowledgements
This study was financially supported by grants from
Swedish Cancer Society (Cancerfonden).
Abbreviations:
|
ABD
|
albumin binding domain
|
|
BSA
|
bovine serum albumin
|
|
EF-2
|
elongation factor 2
|
|
Fv
|
variable fragment
|
|
HER2
|
human epidermal growth factor receptor
2
|
|
HSA
|
human serum albumin
|
|
IMAC
|
immobilized metal-ion affinity
chromatography
|
|
KD
|
equilibrium dissociation constant
|
|
MSA
|
mouse serum albumin
|
|
PBS
|
phosphate-buffered saline
|
|
scFv
|
single-chain variable fragment
|
|
PE38
|
truncated exotoxin A from
Pseudomonas aeruginosa
|
References
|
1
|
Kreitman RJ: Recombinant immunotoxins
containing truncated bacterial toxins for the treatment of
hematologic malignancies. BioDrugs. 23:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Alewine C, Hassan R and Pastan I: Advances
in anticancer immunotoxin therapy. Oncologist. 20:176–185. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Martin-Killias P, Stefan N, Rothschild S,
Plückthun A and Zangemeister-Wittke U and Zangemeister-Wittke U: A
novel fusion toxin derived from an EpCAM-specific designed ankyrin
repeat protein has potent antitumor activity. Clin Cancer Res.
17:100–110. 2011. View Article : Google Scholar
|
|
4
|
Zielinski R, Lyakhov I, Jacobs A, Chertov
O, Kramer-Marek G, Francella N, Stephen A, Fisher R, Blumenthal R
and Capala J: Affitoxin - a novel recombinant, HER2-specific,
anticancer agent for targeted therapy of HER2-positive tumors. J
Immunother. 32:817–825. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zielinski R, Lyakhov I, Hassan M, Kuban M,
Shafer-Weaver K, Gandjbakhche A and Capala J: HER2-affitoxin: A
potent therapeutic agent for the treatment of HER2-overexpressing
tumors. Clin Cancer Res. 17:5071–5081. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mazor R, Vassall AN, Eberle JA, Beers R,
Weldon JE, Venzon DJ, Tsang KY, Benhar I and Pastan I:
Identification and elimination of an immunodominant T-cell epitope
in recombinant immunotoxins based on Pseudomonas exotoxin A. Proc
Natl Acad Sci USA. 109:E3597–E3603. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Onda M, Beers R, Xiang L, Nagata S, Wang
Q-C and Pastan I: An immunotoxin with greatly reduced
immunogenicity by identification and removal of B cell epitopes.
Proc Natl Acad Sci USA. 105:11311–11316. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu W, Onda M, Lee B, Kreitman RJ, Hassan
R, Xiang L and Pastan I: Recombinant immunotoxin engineered for low
immunogenicity and antigenicity by identifying and silencing human
B-cell epitopes. Proc Natl Acad Sci USA. 109:11782–11787. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Onda M, Beers R, Xiang L, Lee B, Weldon
JE, Kreitman RJ and Pastan I: Recombinant immunotoxin against
B-cell malignancies with no immunogenicity in mice by removal of
B-cell epitopes. Proc Natl Acad Sci USA. 108:5742–5747. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
King C, Garza EN, Mazor R, Linehan JL,
Pastan I, Pepper M and Baker D: Removing T-cell epitopes with
computational protein design. Proc Natl Acad Sci USA.
111:8577–8582. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Carlsson J, Nordgren H, Sjöström J, Wester
K, Villman K, Bengtsson NO, Ostenstad B, Lundqvist H and Blomqvist
C: HER2 expression in breast cancer primary tumours and
corresponding metastases. Original data and literature review. Br J
Cancer. 90:2344–2348. 2004.PubMed/NCBI
|
|
12
|
Ménard S, Casalini P, Campiglio M, Pupa S,
Agresti R and Tagliabue E: HER2 overexpression in various tumor
types, focussing on its relationship to the development of invasive
breast cancer. Ann Oncol. 12(Suppl 1): S15–S19. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ahmed S, Sami A and Xiang J: HER2-directed
therapy: Current treatment options for HER2-positive breast cancer.
Breast Cancer. 22:101–116. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Amiri-Kordestani L, Blumenthal GM, Xu QC,
Zhang L, Tang SW, Ha L, Weinberg WC, Chi B, Candau-Chacon R, Hughes
P, et al: FDA approval: Ado-trastuzumab emtansine for the treatment
of patients with HER2-positive metastatic breast cancer. Clin
Cancer Res. 20:4436–4441. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Löfblom J, Feldwisch J, Tolmachev V,
Carlsson J, Ståhl S and Frejd FY: Affibody molecules: Engineered
proteins for therapeutic, diagnostic and biotechnological
applications. FEBS Lett. 584:2670–2680. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Lundberg E, Vernet E, Larsson B,
Höidén-Guthenberg I and Gräslund T: Selection of affibody molecules
to the ligand-binding site of the insulin-like growth factor-1
receptor. Biotechnol Appl Biochem. 55:99–109. 2010.PubMed/NCBI
|
|
17
|
Friedman M, Nordberg E, Höidén-Guthenberg
I, Brismar H, Adams GP, Nilsson FY, Carlsson J and Ståhl S: Phage
display selection of Affibody molecules with specific binding to
the extracellular domain of the epidermal growth factor receptor.
Protein Eng Des Sel. 20:189–199. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Orlova A, Magnusson M, Eriksson TLJ,
Nilsson M, Larsson B, Höidén-Guthenberg I, Widström C, Carlsson J,
Tolmachev V, Ståhl S, et al: Tumor imaging using a picomolar
affinity HER2 binding affibody molecule. Cancer Res. 66:4339–4348.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kronqvist N, Malm M, Göstring L,
Gunneriusson E, Nilsson M, Höidén Guthenberg I, Gedda L, Frejd FY,
Ståhl S and Löfblom J: Combining phage and staphylococcal surface
display for generation of ErbB3-specific Affibody molecules.
Protein Eng Des Sel. 24:385–396. 2011. View Article : Google Scholar
|
|
20
|
Feldwisch J, Tolmachev V, Lendel C, Herne
N, Sjöberg A, Larsson B, Rosik D, Lindqvist E, Fant G,
Höidén-Guthenberg I, et al: Design of an optimized scaffold for
affibody molecules. J Mol Biol. 398:232–247. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sörensen J, Sandberg D, Sandström M,
Wennborg A, Feldwisch J, Tolmachev V, Åström G, Lubberink M,
Garske-Román U, Carlsson J, et al: First-in-human molecular imaging
of HER2 expression in breast cancer metastases using the
111In-ABY-025 affibody molecule. J Nucl Med. 55:730–735. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Makrides SC, Nygren PA, Andrews B, Ford
PJ, Evans KS, Hayman EG, Adari H, Uhlén M and Toth CA: Extended in
vivo half-life of human soluble complement receptor type 1 fused to
a serum albumin-binding receptor. J Pharmacol Exp Ther.
277:534–542. 1996.PubMed/NCBI
|
|
23
|
Jonsson A, Dogan J, Herne N, Abrahmsén L
and Nygren P-A: Engineering of a femtomolar affinity binding
protein to human serum albumin. Protein Eng Des Sel. 21:515–527.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Orlova A, Jonsson A, Rosik D, Lundqvist H,
Lindborg M, Abrahmsen L, Ekblad C, Frejd FY and Tolmachev V:
Site-specific radiometal labeling and improved biodistribution
using ABY-027, a novel HER2-targeting affibody
molecule-albumin-binding domain fusion protein. J Nucl Med.
54:961–968. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tolmachev V, Orlova A, Pehrson R, Galli J,
Baastrup B, Andersson K, Sandström M, Rosik D, Carlsson J,
Lundqvist H, et al: Radionuclide therapy of HER2-positive
microxenografts using a 177Lu-labeled HER2-specific Affibody
molecule. Cancer Res. 67:2773–2782. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hopp J, Hornig N, Zettlitz KA, Schwarz A,
Fuss N, Müller D and Kontermann RE: The effects of affinity and
valency of an albumin-binding domain (ABD) on the half-life of a
single-chain diabody-ABD fusion protein. Protein Eng Des Sel.
23:827–834. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Andersen JT, Pehrson R, Tolmachev V, Daba
MB, Abrahmsén L and Ekblad C: Extending half-life by indirect
targeting of the neonatal Fc receptor (FcRn) using a minimal
albumin binding domain. J Biol Chem. 286:5234–5241. 2011.
View Article : Google Scholar :
|
|
28
|
Gunneriusson E, Nord K, Uhlén M and Nygren
P: Affinity maturation of a Taq DNA polymerase specific affibody by
helix shuffling. Protein Eng. 12:873–878. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sandersjöö L, Jonsson A and Löfblom J: A
new prodrug form of Affibody molecules (pro-Affibody) is
selectively activated by cancer-associated proteases. Cell Mol Life
Sci. 72:1405–1415. 2015. View Article : Google Scholar
|
|
30
|
Tolmachev V, Hofström C, Malmberg J,
Ahlgren S, Hosseinimehr SJ, Sandström M, Abrahmsén L, Orlova A and
Gräslund T: HEHEHE-tagged affibody molecule may be purified by
IMAC, is conveniently labeled with
[99mTc(CO)3]+, and shows improved
biodistribution with reduced hepatic radioactivity accumulation.
Bioconjug Chem. 21:2013–2022. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hassan R, Bullock S, Premkumar A, Kreitman
RJ, Kindler H, Willingham MC and Pastan I: Phase I study of SS1P, a
recombinant anti-mesothelin immunotoxin given as a bolus I.V.
infusion to patients with mesothelin-expressing mesothelioma,
ovarian, and pancreatic cancers. Clin Cancer Res. 13:5144–5149.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kuan CT, Pai LH and Pastan I: Immunotoxins
containing Pseudomonas exotoxin that target LeY damage human
endothelial cells in an antibody-specific mode: Relevance to
vascular leak syndrome. Clin Cancer Res. 1:1589–1594.
1995.PubMed/NCBI
|