Introduction
The fight or flight response triggered by stress is
elicited by the production of mediators, such as the
catecholamines, norepinephrine (NE) and epinephrine (E), from the
sympathetic nervous system and the adrenal medulla. The
hypothalamic-pituitary-adrenal response includes the release of
corticotrophin-releasing hormone from the hypothalamus and the
secretion of adrenocorticotrophic hormone from the anterior
pituitary. This process results in a downstream release of
glucocorticoids, such as cortisol, from the adrenal cortex. The
tumor is not independent from its microenvironment (1,2); in
fact, it interacts with its adjacent cells and also with
neuroendocrine system. Tumor cells express different
neurotransmitter receptors and react with different
neurotransmitters released by the autonomic nervous system from the
brain, peripheral plexuses, ganglia and adrenal medulla (1,2).
Therefore, it is increasingly clear that malignant tumors are under
the regulatory control of the nervous system, and that these
signals may represent a metastatic drive to tumor cells. This
process is called neoneurogenesis and, as angiogenesis forms new
vessel in the tumors, it occurs when produced neurotrophins
stimulate adjacent nerve cells to develop nerve endings into the
tumor. This innervation could stimulate the development of
metastases, since tumor cells respond to neurotransmitters with
increased migratory activity (e.g., norepinephrine). Actually, many
biological cancer findings are focused on the regulation of
migration of tumor cells that is a prerequisite for cancer invasion
and metastasis. For example, it has been shown that chemokinesis
induces the migration of tumor cells (1,2), and
directs the tumor cells to specific organs (3). Norepinephrine is a neurotransmitter
released in stress reactions. The long-lasting elevation of
catecholamines attributable to chronic stress, is known to be a
risk factor for heart disease as well as for cancer. Previous
studies provided molecular evidence for a functional link between
psychoneurological events, through the increasing release of the
neurotransmitter norepinephrine in vivo, and their
promigratory influence in tumor cells in vitro. In previous
studies, some authors examined the expression of β2-adrenoreceptor
(β2-AR) in breast, colon and prostatic cancer, and raised the
possibility that β2-AR could be involved in metastasis to the
location of these cancers (4–8).
Epithelial-mesenchymal transition (EMT) has a key role in the
progression of cancer to an invasive phenotype. The differential
remodeling of E-cadherin and vimentin has been characterized in
cells that have undergone EMT. The loss of E-cadherin occurs in
most epithelial cancers, and can be correlated to higher mobility
and invasiveness of tumor cells (9,10).
On the other hand, the increased vimentin expression is usually
associated with increased in vitro motility of cancer cell
lines (9,10).
In the present study, we found that treatment with
norepinephrine induces an increased in vitro and in
vivo migration of DU145 human hormone-independent prostate
cancer cell line. Moreover, in norepinephrine treated mice the
increased expression of MMP2 and MMP9 in tumor tissues is
paralleled by the presence of metastasis in lymph nodes proximal to
the tumor. These effects are antagonized by β2AR antagonist
propranolol. Collectively, the data show that norepinephrine
initiates and drives migration providing a rationale for both the
development and localization of metastases and that β-blockers such
as propranolol could be used for chemoprevention of metastasis
development.
Materials and methods
Cells
Human prostate cancer cell line hormone-independent
DU145, was purchased from American Type Culture Collection (ATCC,
Manassas, VA, USA) and grown in Dulbecco’s Modified Eagle’s medium
(DMEM) supplemented with glutamine, essential and non-essential
amino acids, vitamins, antibiotics, and 10% heat-inactivated fetal
bovine serum (Gibco/Invitrogen, Grand Island, NY, USA). The cells
were maintained at 37°C in a humidified atmosphere of 5%
CO2. All experiments were performed with cultures grown
for no longer than 6 weeks after recovery from frozen stocks.
Animals
Thirty-eight-week-old male Foxnnu/nu mice
were purchased by Harlan, San Pietro al Natisone (Italy). Mice were
housed 5 per cage and maintained on a 12-h light:12-h dark cycle
(lights on at 7.00 a.m.) in a temperature-controlled room (22±2°C)
and with food and water ad libitum. The experimental
protocols were in compliance with the European Community Council
directive (86/609/EEC). After 1-week of acclimation to the housing
conditions, mice were injected subcutaneously (s.c.) with a
suspension of DU145 cells (1×106 cells/mouse in the
right hind limb).
After 1 week, mice were randomized for tumor volume
and weight according to Fisher’s exact test. Tumor growth was
measured every 2–3 days with a digital caliper 2BIOL (Besozzo,
Italy) and expressed as volume, according to the formula: V = (a ×
b2)/2 (a = the largest superficial diameter and b = the smallest
superficial diameter). Animals, were sacrificed with cervical
dislocation when they reached the cut-off of 1500 mm3 or
when presenting signs of pain.
Drug administration
When the tumor mass was palpable and measurable,
mice were treated with S-propranolol hydrochloride, a non-selective
β-adrenoreceptor antagonist (Sigma-Aldrich, St. Louis, MO, USA) (2
mg/kg/day), and with 28 μM of norepinephrine tartrate
(Sigma-Aldrich) via intra-peritoneal injection, or either
combination every day until they reached the cut-off.
Wound healing assay or proliferation
assay
DU145 cells were seeded at the density of
40×103 cells per well into a 24-multiwell plate, and
cultured in their respective culture medium supplemented with 1%
FBS. At the time of confluence, the cells were incubated in the
absence or presence of norepinephrine and propranolol and their
combination at same concentration (10 μM) for 24 and 48 h. Then, a
slit was made horizontally with a white tip at the center of each
confluent well, the medium was changed after gentle rinse and cells
were cultured for 24 h with or without norepinephrine and
propranolol. Cell invasion on the slit of the confluent well, was
assessed at time 0, 24 and 48 h in each condition, by light
microscopy.
Invasion test
In vitro cell migration was assayed in
24-well cell culture plates using inserts with 8-μm pore membranes
(Costar, San Diego, CA, USA) according to the method described by
Katayama et al (11). In
brief, membrane was precoated with 20 μl per insert of Matrigel (BD
Pharmingen, Bedford, MA, USA) diluted 1:1 by RPMI-1640. DU145 cell
line (1×105 per ml) were suspended in 500 μl migration
buffer (DMEM/0.5% bovine serum albumin) per well and loaded into
the upper compartment of the chamber. The lower compartment of the
chamber was loaded with 10–400 ng/ml norepinephrine (Sigma, St.
Louis, MO, USA) in 500 μl migration buffer. After incubation at
37°C for 24 h, the cells on the lower surface of membrane were
fixed in 70% ethanol, stained with hematoxylin, and counted under a
microscope (Olympus, Tokyo, Japan). Each value was expressed as a
mean number of five different fields. Three experiments were
performed independently and the results were compared.
MTT
Cell viability was evaluated using 3-(4, 5-dimethyl
(thia zol-2-yl)-2, 5-diphenyltetrazolium bromide) (MTT), the
reagent that measures the metabolic activity of cells. The stock
solution of MTT (5 mg/ml) was prepared in phosphate-buffered saline
(PBS) and stored in the dark at 4°C. A 100-μl aliquot of a dilution
prepared in culture medium (1 mg/ml final) was filtered (0.22 μm)
and added to the cells growing with/without CBGM. After 3 h of
incubation the supernatant was removed. Formazan crystals in viable
cells were dissolved in ethanol (96% v/v) and absorbance was
measured by the Multiskan® FC microplate photometer
(Thermo Scientific, USA) at 540 nm. After the treatment described
above, MTT (Sigma) was added to a final concentration of 0.5 mg/ml
and incubated for 4 h at 37 °C. The culture medium was then
removed, and the remaining blue precipitate was solubilized in DMSO
followed by an absorbance reading at 570 nm in a plate reader using
630 nm as a reference (Spectra Max 340; Molecular Devices,
Sunnyvale, CA, USA). This reading was divided by the adjusted
absorbance reading of untreated cells in control wells to obtain
the percentage of cell survival. All experiments were carried out
in triplicates.
Histology and immunohistochemistry
(IHC)
Sections were deparaffinized in xylene for 10 min
each and then were rehydrated through graded alcohols. To inhibit
endogenous peroxide activity, sections were rinsed in mixture of
100% methanol and 0.3% hydrogen peroxide for 40 min. Then, they
were put in a microwave oven in a jar filled with 10 mmol/l sodium
citrate buffer (pH 6.0) for 10 min. Following this treatment,
sections were allowed to cool at room temperature. Sections were
incubated with normal goat serum (Zymed, San Diego, CA, USA) at RT
for 20 min and then incubated with chicken anti-MMP-2, MMP-9 5–15
μg/ml (R&D Systems, Minneapolis, MN, USA) and pan cytokeratin
antibody 1:50 (Dako, Italy) at room temperature for 1 h. A negative
control was used in all experiments without the primary antibody.
After the period of incubation, sections were washed three times
with PBS. Then, they were incubated with the linking reagent
(biotin-labeled affinity purified antibody; KPL, Gaithersburg, MD,
USA) at room temperature for 1 h. After three washes with PBS, the
sections were incubated with a complex of avidin DH and
biotinylated enzyme (Zymed) at room temperature for 30 min. Then,
sections were washed three times with PBS again and incubated with
a mixture of an equal volume of 0.02% hydrogen peroxide and
diaminobenzidine tetrahydrochloride (Dako, Italy) for 1 min in the
dark. After chromogenic development, sections were washed in water
and counter-stained with hematoxylin. The stained slides were
investigated independently by two pathologists who had no knowledge
of the clinical parameters and outcomes. For microscope analysis of
the slides, they each selected five high-powered fields whereby
each field contained >200 tumor cells, and counted both positive
and negative cancer cells. In total, >1,000 tumor cells were
counted. We calculated the average of 10 readings of positive cells
as a percentage of all the cells to determine staining scores. We
judged the expression of MMP2 and MMP9 and Pan cytokeratin. The
cases were graded as either positive of marker expression (MMP-2,
MMP-9, Pan cytokeratin >30%) or negative cases (<30% positive
tumor cells) for the subsequent statistical analyses.
Immunostaining and confocal
microscopy
DU145 cells were treated with 10 μM norepinephrine
(NE10), 10 μM propranolol (P10) and N10-P10 combination. After 48
h, DU145 cells were fixed in PBS 4% paraformaldehyde then
permeabilized for 5 min with PBS 1% Triton. Immunostaining was
carried out by incubation with anti-E-cadherin and anti-vimentin
antibodies 1:1,000 followed by revelation using Alexa Fluor
633-conjugated anti-rabbit immunoglobulin (Ig)G antibodies and
Alexa Fluor 488-conjugated anti-rabbit IgG antibodies, respectively
(Jackson Immunoresearch Laboratories, West Grove, PA, USA) at a
dilution of 1:1,000 for 1 h. The cells were analyzed by an LSM-410
Zeiss confocal microscope.
Semiquantitative Rt-PCR analysis
Cells (DU145) grown in monolayers were harvested at
early confluence. Total RNA was extracted from cultured cells using
TRIzol reagent. The RNA concentration was measured by Biophotometer
(Eppendorf) at 260 nm. Then 1 μl of the product was subjected to
PCR amplification using Mastercycler Personal (Eppendorf, Italy)
according to the manufacturer’s instructions. Semiquantitative
RT-PCR was performed starting with 1cDNA, followed by specific gene
product amplification with One-Step RT-PCR kit (Invitrogen,
Burlington, ON, Canada). PCR cycling conditions for b2-AR mRNA were
95°C for 5 min; then 32 cycles of 94°C for 1 min, 54°C for 1 min,
72°C for 1 min; and finally extension at 72°C for 7 min. The primer
sequences of human b2-AR were designed as follows: forward
5′-ACGCAGCAAAGGGACGAG-3′ and reverse 5′-CACACCATCAGAATGATCAC-3′.
Experiments were performed three times, for densitometric analysis
of each set of data. PCR products were resolved by electrophoresis
in 1–2% agarose gels and visualized by ethidium bromide staining.
The densities of the b2-AR bands were divided by those of the GAPDH
bands.
Protein extraction and western blot
analysis
Cell lysates were prepared by adding ice-cold lysis
buffer [0.5% Triton X-100, 50 mmol/l Tris (pH 7.2), 140 mmol/l
NaCl, 10 mmol/l EDTA, 50 mmol/lNaF, 1 mmol/l
Na3VO4] containing the protease inhibitor
cocktail Complete Mini (Roche, Mannheim, Germany). Protein samples
were mixed with an equal volume of 2% sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE) sample buffer, boiled
for 5 min, and then separated using 10% SDS-PAGE. After
electrophoresis, proteins were transferred to PVDF membranes by
semi-dry electrophoretic transfer. The membranes were blocked in 5%
dry milk, rinsed and then incubated with primary antibody of p53
(Sigma) overnight at 4°C. The primary antibody was removed,
membranes were washed four times and followed by HRP-conjugated
secondary antibody. Detection was then performed using an enhanced
chemiluminescence kit and exposed to X-ray film. Anti α-actin was
used as loading control.
Statistical analysis
Normally distributed data are presented as mean ±
SEM. Two-way ANOVA and Bonferroni post hoc test, were used
to examine the significance of differences among groups (Graph pad
Prism 5.0).
Results
Norepinephrine increases proliferation of
DU145 cancer cells
We first determined whether norepinephrine increases
the proliferation of human prostate cancer cells DU145 by
performing in vitro assays. Proliferation test was performed
by wound healing assay on cells treated with norepinephrine at 10
μM (NE10), propranolol at 10 μM (P10) and their combination at the
same concentration (NE+P10). Our results indicate that
norepinephrine (Fig. 1B, F and J)
increased the proliferation of DU145 at 48 h compared to controls
(Fig. 1A, E and I), to propranolol
(Fig. 1C, G and K) and the
combination (Fig. 1D, H and L).
These results were also confirmed by MTT assay (data not
shown).
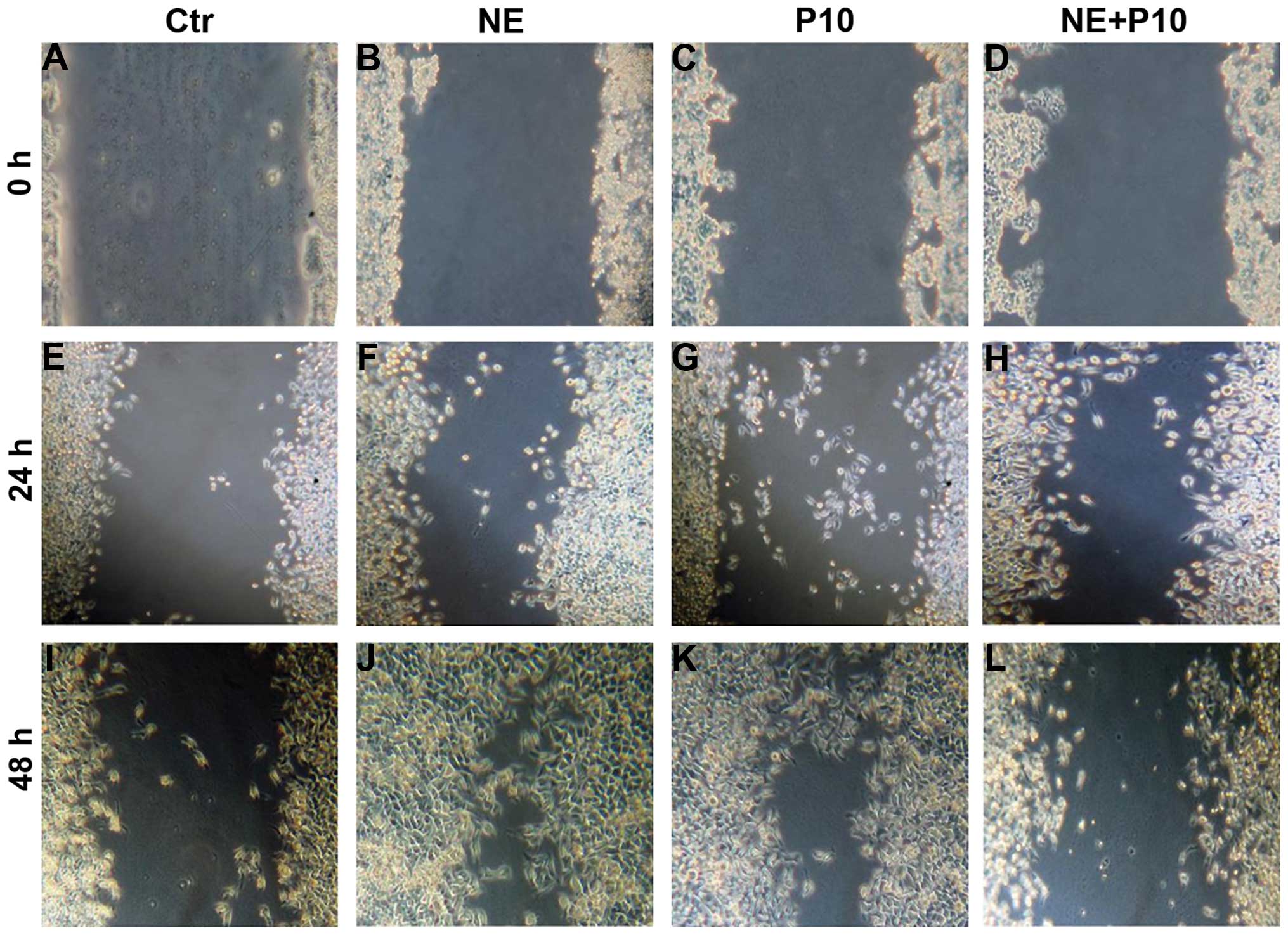 | Figure 1Norepinephrine stimulates
proliferation in DU145 cell lines. DU145 cancer cells were
incubated in medium containing (A, E and I) medium (Ctr), (B, F and
J) 10 μm norepinephrine (NE10), (C, G and K), 10 μm propranolol
(P10), (D, H and L) 10 μm propanolol and 10 μm propranolol
(NE10+P10). Cell migration rates were quantitatively assessed by
counting the number of cells in the denuded area at 0, 24, and 48 h
after wound induction. At 48 h after wound induction, there were
clearly more cells in the denuded area of norepinephrine-treated
cells than untreated cells or propranolol or combination-treated
cells. |
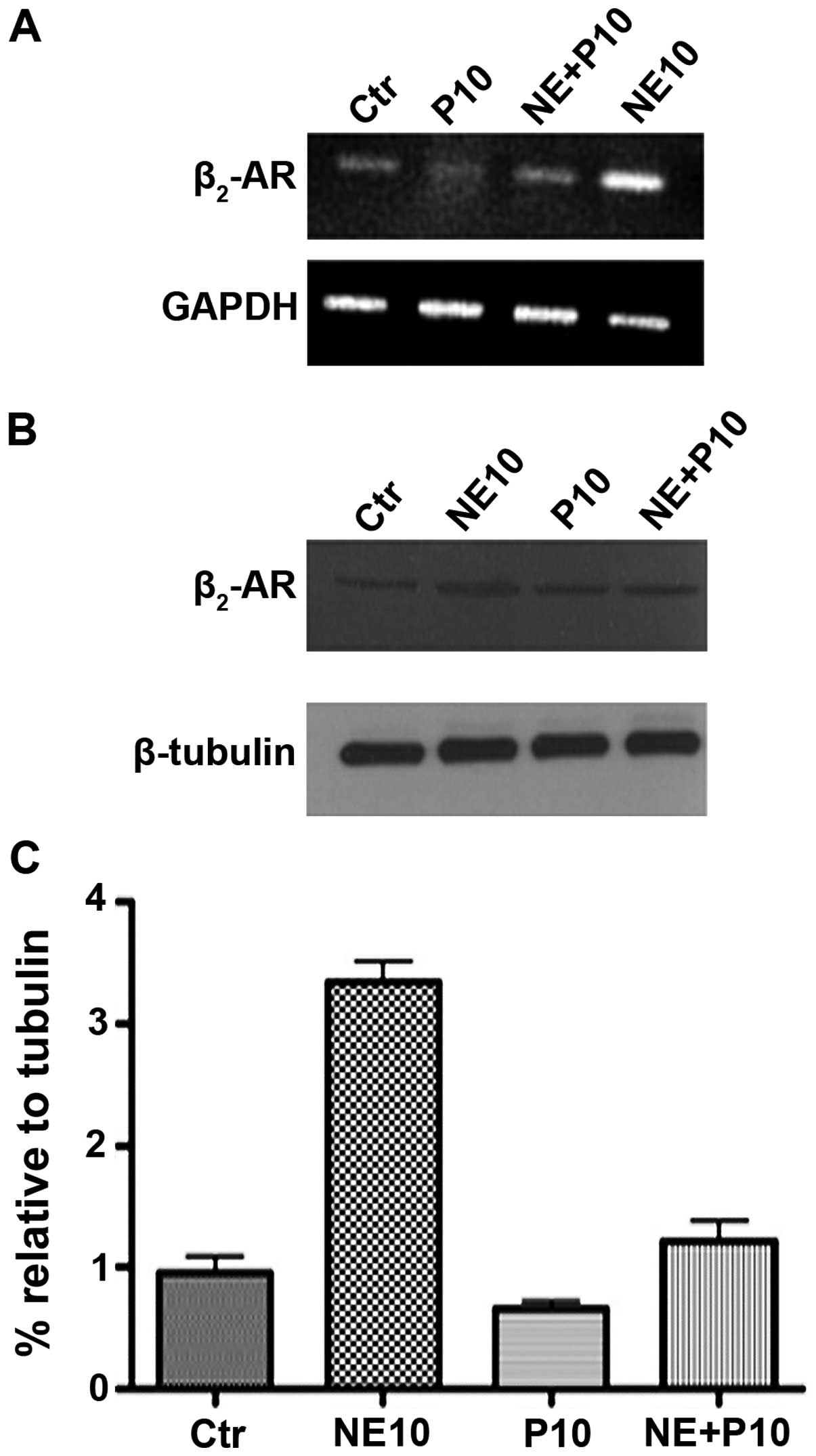
Norepinephrine influences the expression
of β2-adrenoreceptor in DU145 cell line
Since it has been demonstrated that the expression
of β2-adrenoreceptor (β2-AR) in DU145 cell line is overexpressed
when these cells are treated with norepinephrine, we performed
expression studies of β2-AR on RNAs and proteins extracted from
DU145 cells treated with 10 μM norepinephrine (NE10), 10 μM
propranolol (P10) and their combination (NE+P10). RT-PCR analysis
and western blotting results (Fig. 2A
and B) showed that norepinephrine causes approximately 3-fold
increase of β2-AR protein expression paralleled by enhanced
expression of its mRNA. Propanolol almost completely antagonized
this effect in DU145 cells.
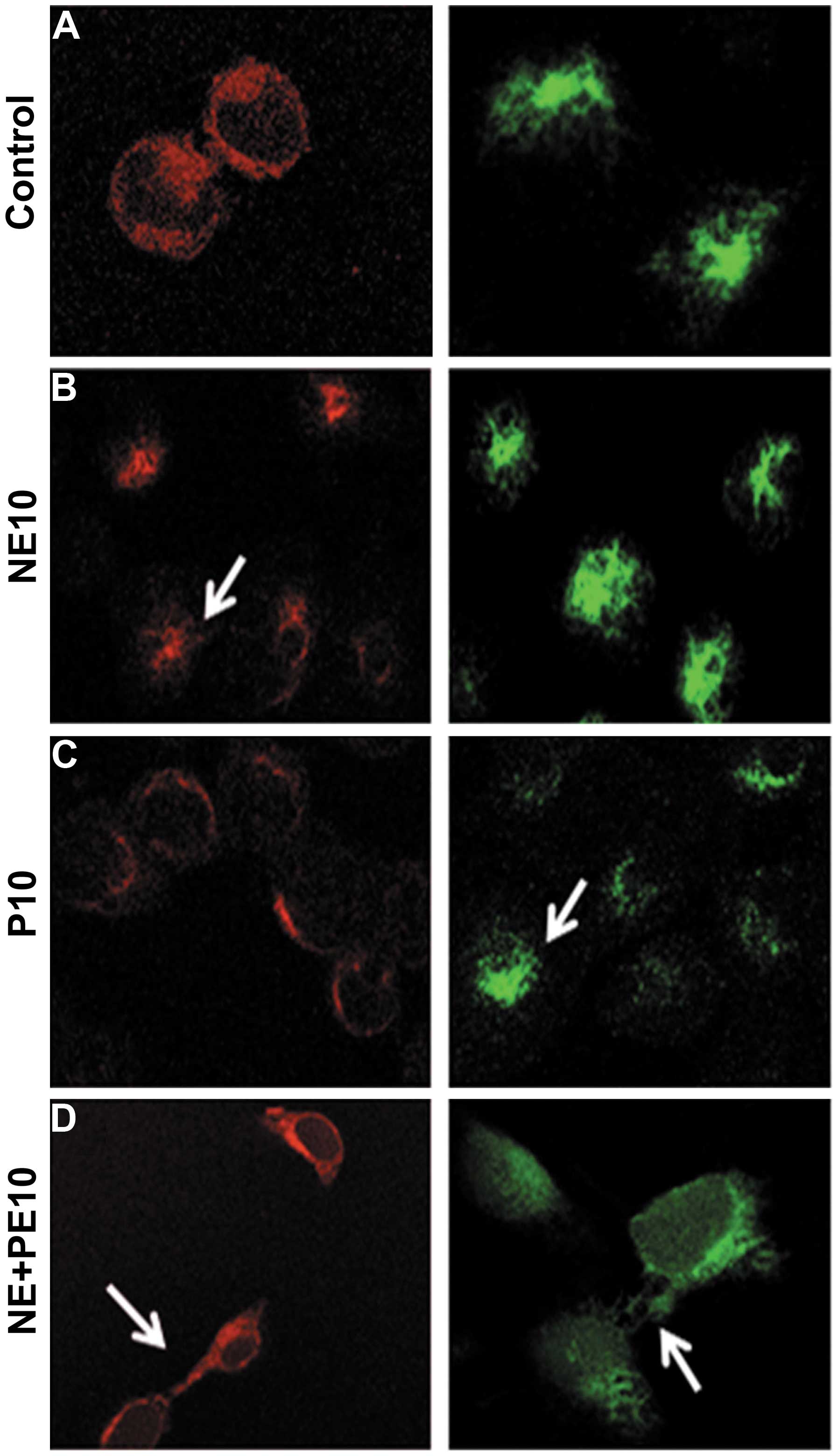
Propranolol antagonizes the effects of
norepinephrine on both E-cadherin and vimentin expression and
intracellular localization
We investigated the mechanisms of the in vivo
acquisition of tumor cell migratory properties induced by
norepinephrine treatment and its possible correlation to EMT
occurrence. Therefore, we evaluated both cell expression and
localization of E-cadherin and vimentin in prostate cancer DU145
cells. Confocal microscopy showed that 10 μM norepinephrine (NE10)
(Fig. 3B) reduced E-cadherin
expression and caused a delocalization of E-cadherin from cell
membrane to the cytoplasm. The addition of 10 μM propranolol (P10)
30 min before NE10 treatment completely antagonized this effect
(Fig. 3D). Moreover, NE10
increased the expression of vimentin and P10 antagonized again this
effect together with polar localization of vimentin on the inner
side of cell membrane (Fig. 3D).
Interestingly, P10 alone had slight effects on E-cadherin
expression and localization if compared to untreated cells while
inducing a decreased expression and delocalization of vimentin
(Fig. 3C). These effects suggest
that P10 antagonized the EMT occurrence induced by NE10 that mimics
an in vivo stress condition.
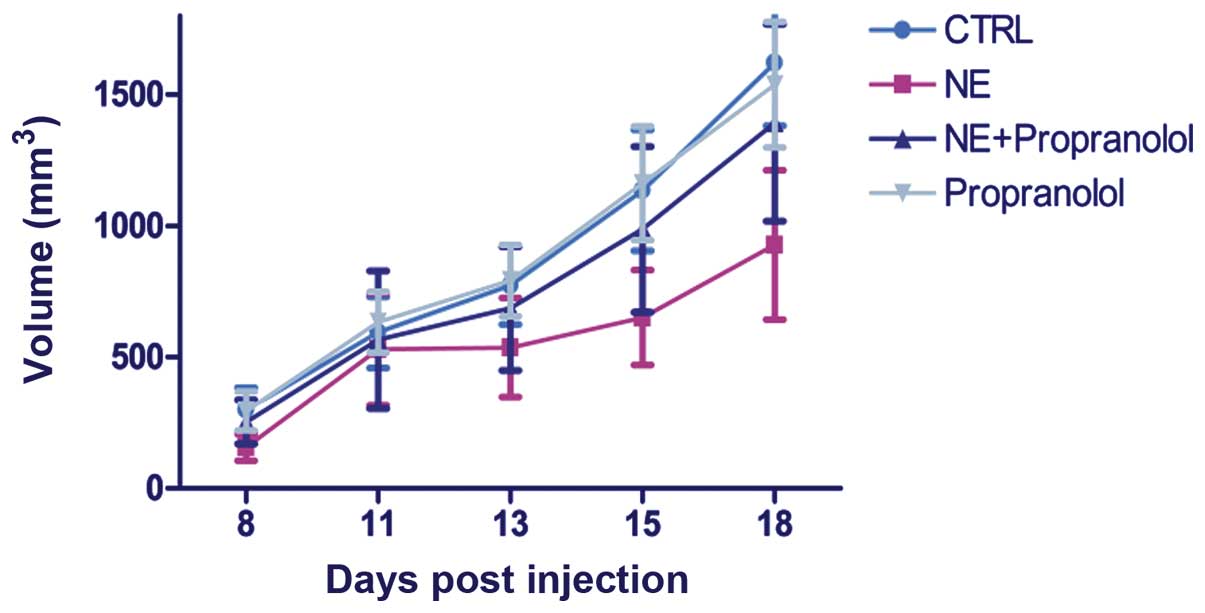
The effects of norepinephrine on tumor
growth and metastasis formation in a xenograft mouse model of
prostate cancer
In order to study the role of norepinephrine on
in vivo tumor growth, we generated a mouse model of prostate
cancer by injection of DU145 cells subcutaneously into the right
flank of mice. When tumors reached ~30–60 mm3, 2 weeks
following cell injection, the mice were randomized into four
groups: a) normal saline (control); b) 28 mM NE; c) 2 mg/kg
propranolol; d) 28 mM NE and 2 mg/kg propanol. Tumor volumes were
monitored once a week by using a digital caliper. Therapy was
continued for 3 weeks. We also monitored the body weight of mice
twice a week until the end of treatment. No difference was observed
between the body weight of the different animal groups, indicating
that treatments of mice with drugs is not associated with toxic
effects. Mice were sacrificed at the end of treatment. Differently
from that demonstrated by in vitro assays, in vivo
experiments showed that norepinephrine is not able to influence the
tumor growth of mice (P>0.05) (Fig.
4). In fact, NE induced an apparent but not significant
decrease of tumor volume and this effect was almost completely
antagonized again by the treatment with propranolol.
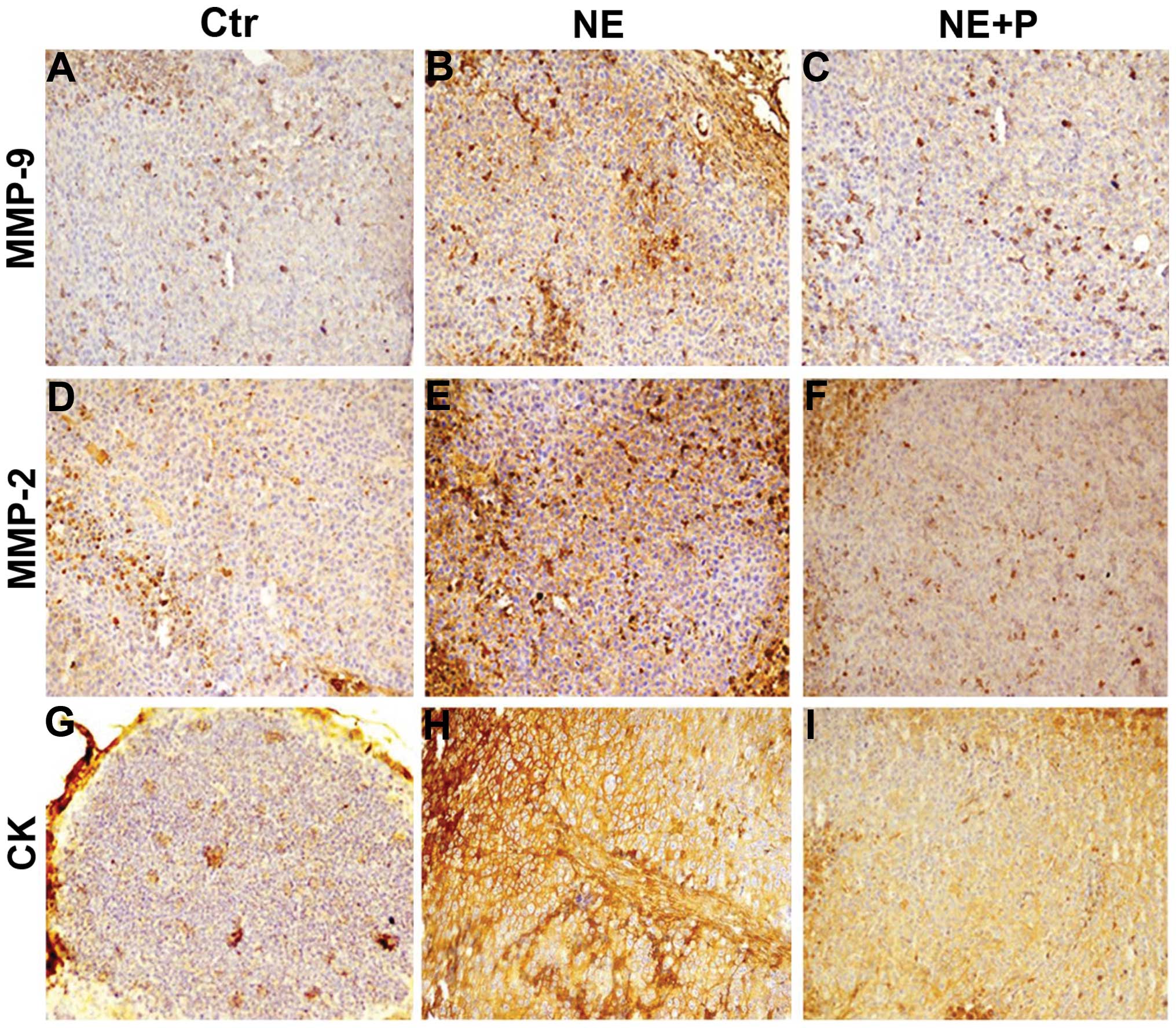
Norepinephrine influences the expression
of cytokeratin (CK), MMP-2 and MMP-9
In order to show whether norepinephrine in
vivo treatment induces the formation of metastasis foci in mice
we performed immunostaining for pan-cytokeratin in inguinal lymph
nodes in order to find epithelial cells that normally are absent.
Our results show a significant increase of the positivity of lymph
nodes for cells expressing CK in all the NE-treated animals
suggesting a strong increase of metastatic cells and this effect
was potently antagonized by propranolol in the combination-treated
animals (Fig. 5G, H and I).
Moreover, overexpression of MMP2 and MMP9 in NE treated tumor
samples (Fig. 5B and E) was
observed compared to those of controls (Fig. 5A and D) and of mice treated with
combination of the two substances (Fig. 5C and F). These data suggest an
in vivo EMT effect induced by NE, with consequent increase
of the metastatic processes, is not paralleled by tumor growth
promotion.
Discussion
Several preclinical and clinical studies have shown
that psychosocial stress has a role in the incidence and
progression of cancer. The molecular basis underlying the
connection between the neuroendocrine system and malignant tumors
is still unclear. In recent years, it has been demonstrated that
the immune system plays a role of mediator between these two
systems and immunosuppression was supposed to enhance tumor
establishment (11,12). Even if this role has been also
demonstrated in our previous work on chronic stress and melanoma
(13–16), in the present investigation, we
dissected the role of NE in a complex living system represented by
athymic mice that lack important parts of the cellular immune
system demonstrating its effects also in an immunocompromised
model. Moreover, it has been demonstrated that norepinephrine
influences the cytotoxicity of natural killer cells (17) that could explain, at least in part,
the effects on metastasis formation as we show in this report. In
general, immunosuppression by NE or other neurotransmitters might
contribute to metastasis formation. Current research has revealed
that several neurotransmitters have direct influence on the
migratory activity and invasiveness of tumor cells (7–9,17)
and does not need the immune system as a mediator. In fact, many
studies have demonstrated in vitro that stress-related
neurotransmitters are the most potent direct stimulators for the
migration of carcinoma cells of various tissue origins. These
neurotransmitters are NE, dopamine and substance P (18–22).
We showed that the in vitro migratory activity of DU145
cells was increased after the treatment with NE. This leads to an
augmented and earlier migration of tumor cells from the primary
tumor into inguinal lymph nodes, and consequently to larger
metastases formation in mice (23–28).
Whether NE leads to organ-specific metastasis to other NE-rich
organs must be shown in further experiments with an enhanced study
design. Tumor cell migration to lymph nodes occurs rarely in an
experimental mouse model of heterotopic cancer because the
xenografted tumor mass is generally well encapsulated. Therefore,
NE treatment induces an increased migratory effect that is in line
with the frequent clinical observation of metastasis development.
Lymph nodes and other organs of the immune system, such as bone
marrow, thymus and spleen (29)
contain noradrenergic and neuropeptidergic nerve fibres and this
could induce cell migration into the regional lymph nodes behaving
as spreaders (28). Further
investigation will be necessary to elucidate the molecular
mechanisms underlying the role of NE or other neurotransmitters in
metastatic migration towards lymph nodes. We used a model of
subcutaneous injection of tumor cells for the investigation of
metastasis development in inguinal lymph node because mice
initially develop solid primary tumors. The pro-migration effect of
NE in vivo was inhibited by the β-blocker propranolol that
antagonized the prometastatic effect of NE. Furthermore, we showed
that propranolol reduced metastases formation in mice compared to
controls. This can be related to the blocking of β2-AR that is
responsible for the effects of norepinephrine endogenously produced
by mice. These results open a new scenario for the use of
β-blockers for chemoprevention of metastasis development in
patients with diagnosed cancer, because the diagnosis of cancer
itself causes stress and especially in prostate cancer patients
that are emotionally involved for the impact of the disease on the
quality of life (30–36). Moreover, we previously showed that
the induction of chronic stress by restraint in mice induced a
significant increase of tumor growth and angiogenesis in mice
bearing murine melanoma B16F10 paralleled by the activation of
β2-AR by neurotransmitters such as E and NE. These effects were
completely abolished in eNOS knockout mice (13). In the present study, we
demonstrated that stress activates the release of several
substances different from catecholamines leading to an increase of
migration of tumor cells from primary tumor to lymph nodes without
affecting tumor growth (6).
Additionally, we demonstrated that norepinephrine increased the
expression of CK, MMP-2 and MMP-9 paralleled by EMT effects
(E-cadherin loss and vimentin augmentation) (9–11,16),
that are indispensable for metastasis formation. The combination of
our results delivers good perspectives for the development of
non-heart active and β2-specific blockers for the inhibition of
metastasis development. In conclusion, our results strongly support
the concept that the development of metastases is not only
genetically determined, but is under the influence of the
organism’s own signal substances. Further investigation will be
necessary in order to dissect the role of neurotransmitters in
specific types of cancer and the use of β-blockers for the
chemopreventive inhibition of metastasis development.
Acknowledgements
The authors would like to specially thank
Massimiliano Spinelli and Alessandra Trocino from Istituto
Nazionale per lo Studio e la Cura dei Tumori ‘Fondazione Giovanni
Pascale’ - IRCCS, Italy for their kind help in providing informatic
assistance. We also thank Michela Falco, Vitale Del Vecchio and
Amalia Luce for technical assistance. This work was supported by
current research programs of Ministry of Health of Istituto
Nazionale per lo Studio e la Cura dei Tumori ‘Fondazione Giovanni
Pascale’, Naples, Italy.
Abbreviations:
|
CK
|
cytokeratin
|
|
E
|
epinephrine
|
|
EMT
|
epithelial-mesenchymal transition
|
|
IHC
|
immunohistochemistry
|
|
Ig
|
immunoglobulin
|
|
NE
|
norepinephrine
|
|
SDS-PAGE
|
sodium dodecyl sulfate polyacrylamide
gel electrophoresis
|
|
β2-AR
|
β2-adrenoreceptor
|
References
|
1
|
Lang K and Bastian P: Neurotransmitter
effects on tumor cells and leukocytes. Prog Exp Tumor Res.
39:99–121. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schuller HM: Neurotransmission and cancer:
Implications for prevention and therapy. Anticancer Drugs.
19:655–671. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liotta LA: An attractive force in
metastasis. Nature. 410:24–25. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Entschladen F, Drell TL IV, Lang K, Joseph
J and Zaenker KS: Tumour-cell migration, invasion, and metastasis:
Navigation by neurotransmitters. Lancet Oncol. 5:254–258. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Entschladen F, Lang K, Drell TL, Joseph J
and Zaenker KS: Neurotransmitters are regulators for the migration
of tumor cells and leukocytes. Cancer Immunol Immunother.
51:467–482. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Palm D, Lang K, Niggemann B, Drell TL IV,
Masur K, Zaenker KS and Entschladen F: The norepinephrine-driven
metastasis development of PC-3 human prostate cancer cells in
BALB/c nude mice is inhibited by beta-blockers. Int J Cancer.
118:2744–2749. 2006. View Article : Google Scholar
|
|
7
|
Drell TL IV, Joseph J, Lang K, Niggemann
B, Zaenker KS and Entschladen F: Effects of neurotransmitters on
the chemokinesis and chemotaxis of MDA-MB-468 human breast
carcinoma cells. Breast Cancer Res Treat. 80:63–70. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Masur K, Niggemann B, Zanker KS and
Entschladen F: Norepinephrine-induced migration of SW 480 colon
carcinoma cells is inhibited by beta-blockers. Cancer Res.
61:2866–2869. 2001.PubMed/NCBI
|
|
9
|
Lang SH, Hyde C, Reid IN, Hitchcock IS,
Hart CA, Bryden AA, Villette JM, Stower MJ and Maitland NJ:
Enhanced expression of vimentin in motile prostate cell lines and
in poorly differentiated and metastatic prostate carcinoma.
Prostate. 52:253–263. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shan T, Cui X, Li W, Lin W, Li Y, Chen X
and Wu T: Novel regulatory program for norepinephrine-induced
epithelial-mesenchymal transition in gastric adenocarcinoma cell
lines. Cancer Sci. 105:847–856. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shakhar G and Ben-Eliyahu S: In vivo
β-adrenergic stimulation suppresses natural killer activity and
compromises resistance to tumor metastasis in rats. J Immunol.
160:3251–3258. 1998.PubMed/NCBI
|
|
12
|
Reiche EM, Nunes SO and Morimoto HK:
Stress, depression, the immune system, and cancer. Lancet Oncol.
5:617–625. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barbieri A, Palma G, Rosati A, Giudice A,
Falco A, Petrillo A, Petrillo M, Bimonte S, Di Benedetto M,
Esposito G, et al: Role of endothelial nitric oxide synthase (eNOS)
in chronic stress-promoted tumour growth. J Cell Mol Med.
16:920–926. 2012. View Article : Google Scholar
|
|
14
|
Moreno-Smith M, Lutgendorf SK and Sood AK:
Impact of stress on cancer metastasis. Future Oncol. 6:1863–1881.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng Z, Liu L, Zhang C, Zheng T, Wang J,
Lin M, Zhao Y, Wang X, Levine AJ and Hu W: Chronic restraint stress
attenuates p53 function and promotes tumorigenesis. Proc Natl Acad
Sci USA. 109:7013–7018. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Moretti S, Massi D, Farini V, Baroni G,
Parri M, Innocenti S, Cecchi R and Chiarugi P: β-adrenoceptors are
upregulated in human melanoma and their activation releases
pro-tumorigenic cytokines and metalloproteases in melanoma cell
lines. Lab Invest. 93:279–290. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lang K, Drell TL, Niggemann B, Zänker KS
and Entschladen F: Neurotransmitters regulate the migration and
cytotoxicity in natural killer cells. Immunol Lett. 90:165–172.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lang K, Drell TL IV, Lindecke A, Niggemann
B, Kaltschmidt C, Zaenker KS and Entschladen F: Induction of a
metastatogenic tumor cell type by neurotransmitters and its
pharmacological inhibition by established drugs. Int J Cancer.
112:231–238. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang D, Ma Q, Shen S and Hu H: Inhibition
of pancreatic cancer cell proliferation by propranolol occurs
through apoptosis induction: The study of beta-adrenoceptor
antagonist’s anticancer effect in pancreatic cancer cell. Pancreas.
38:94–100. 2009. View Article : Google Scholar
|
|
20
|
Paredes A, Gálvez A, Leyton V, Aravena G,
Fiedler JL, Bustamante D and Lara HE: Stress promotes development
of ovarian cysts in rats: The possible role of sympathetic nerve
activation. Endocrine. 8:309–315. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liotta LA and Kohn EC: The
microenvironment of the tumour-host interface. Nature. 411:375–379.
2001. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lutgendorf SK, Sood AK and Antoni MH: Host
factors and cancer progression: Biobehavioral signaling pathways
and interventions. J Clin Oncol. 28:4094–4099. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vilardi BM, Bravo-Calderón DM, Bernabé DG,
Oliveira SH and Oliveira DT: VEGF-C expression in oral cancer by
neurotrans-mitter-induced activation of beta-adrenergic receptors.
Tumour Biol. 34:139–143. 2013. View Article : Google Scholar
|
|
24
|
Flint MS, Baum A, Episcopo B, Knickelbein
KZ, Liegey Dougall AJ, Chambers WH and Jenkins FJ: Chronic exposure
to stress hormones promotes transformation and tumorigenicity of
3T3 mouse fibroblasts. Stress. 16:114–121. 2013. View Article : Google Scholar :
|
|
25
|
Newton PK, Mason J, Bethel K, Bazhenova L,
Nieva J, Norton L and Kuhn P: Spreaders and sponges define
metastasis in lung cancer: a Markov chain Monte Carlo mathematical
model. Cancer Res. 73:2760–2769. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hara MR, Kovacs JJ, Whalen EJ, Rajagopal
S, Strachan RT, Grant W, Towers AJ, Williams B, Lam CM, Xiao K, et
al: A stress response pathway regulates DNA damage through
β2-adrenoreceptors and β-arrestin-1. Nature. 477:349–353. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Entschladen F, Palm D, Lang K, Drell TL IV
and Zaenker KS: Neoneurogenesis: Tumors may initiate their own
innervation by the release of neurotrophic factors in analogy to
lymphangiogenesis and neoangiogenesis. Med Hypotheses. 67:33–35.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Felten DL: Direct innervation of lymphoid
organs: Substrate for neurotransmitter signaling of cells of the
immune system. Neuropsychobiology. 28:110–112. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Muller WA: Regulate globally, act locally:
adrenergic nerves promote leukocyte recruitment. Immunity.
37:189–191. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sokołowska P and Nowak JZ: Constitutive
activity of beta-adrenergic receptors in C6 glioma cells. Pharmacol
Rep. 57:659–663. 2005.
|
|
31
|
Sloan EK, Priceman SJ, Cox BF, Yu S,
Pimentel MA, Tangkanangnukul V, Arevalo JM, Morizono K, Karanikolas
BD, Wu L, et al: The sympathetic nervous system induces a
metastatic switch in primary breast cancer. Cancer Res.
70:7042–7052. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Armaiz-Pena GN, Cole SW, Lutgendorf SK and
Sood AK: Neuroendocrine influences on cancer progression. Brain
Behav Immun. 30(Suppl): S19–S25. 2013. View Article : Google Scholar
|
|
33
|
Cole SW and Sood AK: Molecular pathways:
Beta-adrenergic signaling in cancer. Clin Cancer Res. 18:1201–1206.
2012. View Article : Google Scholar :
|
|
34
|
Hassan S, Karpova Y, Baiz D, Yancey D,
Pullikuth A, Flores A, Register T, Cline JM, D’Agostino R Jr,
Danial N, et al: Behavioral stress accelerates prostate cancer
development in mice. J Clin Invest. 123:874–886. 2013.PubMed/NCBI
|
|
35
|
Yang EV and Eubank TD: The impact of
adrenergic signaling in skin cancer progression: Possible
repurposing of β-blockers for treatment of skin cancer. Cancer
Biomark. 13:155–160. 2013.
|
|
36
|
Sood AK and Lutgendorf SK: Stress
influences on anoikis (Review). Cancer Prev Res (Phila). 4:481–485.
2011. View Article : Google Scholar
|