Introduction
Cancer immunotherapy, especially when combined with
other therapeutic modalities such as chemotherapy, is an attractive
approach to cancer treatment. Synergistic effects of combinations
of immunotherapy and chemotherapy have been demonstrated in a
number of pre-clinical and clinical studies (1,2).
Dendritic cells (DCs) are key players in the immune
response as they are able to capture antigens with their
pattern-recognition receptors, to process and present them to naïve
T-cells, inducing their activation (3), and thus building an essential bridge
between innate and adaptive responses. The possibility of their
generation in vitro enabled their use for immunotherapy of
cancer (4), and a number of
clinical trials have been performed in the last decade (5,6).
Typically, an autologous dendritic cell-based vaccine represents
in vitro cultured dendritic cells pulsed with tumor antigens
that can be in the form of tumor cells with subsequent DC
maturation. For DC pulsing, tumor cells can be inactivated by their
lysis (ultrasonic treatment, repeated freeze-thaw), lethal
irradiation or other methods before mixing them with DC. Selection
of the optimal inactivation method can be crucial for DC vaccine
optimization, together with selection of proper maturation-inducing
agents.
Therefore, a significant effort has also been
invested in increasing the immunogenicity of dying cancer cells
used for vaccine production. Until now several chemotherapeutic
agents [anthracyclines (7),
oxaliplatin, platinum complexes (8), bortezomib (9)] and physical modalities [UV-C,
irradiation (10), HHP] have been
identified as inducers of immunogenic cell death (ICD). ICD is
characterized by the cell-surface expression and release of damage
associated molecular patterns (DAMPs). DAMPs found to be crucial
for ICD include surface exposed chaperone protein calreticulin
(CRT) and heat shock proteins 70 (HSP70) and 90 (HSP90), actively
secreted ATP and passively released high-mobility group box 1
protein (HMGB1). These signals can activate innate immunity and,
importantly, interact with phagocytosis-related receptors,
purinergic receptors and pattern-recognition receptors expressed by
DCs and thereby stimulate presentation of tumor antigens to T
cells.
High hydrostatic pressure (HHP) has been
demonstrated as a convenient tool for tumor cell inactivation
preserving their immunogenic capacity (11,12).
Recently, induction of ICD by HHP has been shown in several human
tumor cell lines. HHP-treated cells were able to induce
monocyte-derived DC maturation, and DC co-cultured with HHP-treated
tumor cells were able to induce T cell activation in vitro.
These encouraging results suggest that HHP can be an important tool
for tumor cell inactivation before their use for DC pulsing or as
cellular vaccines (13).
Chemotherapeutic drugs affect rapidly growing cells
and, as a consequence, cause collateral damage to cells of the
immune system. In this regard, they are considered
immunosuppressive. However, there is increasing evidence that some
cancer chemotherapies may actually aid the immunotherapy by
activating the immune system rather than suppressing it (14,15).
Chemotherapeutic drugs such as cyclophosphamide, doxorubicin,
paclitaxel or docetaxel (16) were
reported to possess immunomodulatory activities and appeared to be
suitable for chemoimmunotherapy (17,18).
Docetaxel is a widely used chemotherapeutic drug and
represents a first-line chemotherapy for metastatic
castration-resistant prostate cancer (19,20).
The autologous dendritic cell-based vaccines are intensively
studied as an immunotherapy for prostate cancer, and the first
cellular immunotherapy based on activated peripheral blood
mononuclear cells, Sipuleucel T, has been FDA-approved (21). Collectively, combination
chemoimmunotherapy based on docetaxel combined with the DC
treatment represents an attractive modality for advanced prostate
cancer therapy.
In the present study, we investigated, using murine
tumor models, the immunogenicity of the HHP-inactivated tumor cells
in vivo and, furthermore, the possibility to use HHP-treated
tumor cells for preparation of DC-based vaccines. We have
demonstrated the therapeutic capacity of the HHP cells-pulsed DC
vaccines in combination with docetaxel treatments to inhibit growth
of the TRAMP-C2 and TC-1 murine tumors. We have focused on the
immunotherapy of poorly immunogenic TRAMP-C2 tumors, an animal
model of prostate cancer treatment. For comparison, the study was
completed with experiments using immunogenic TC-1 tumors
representing a murine model for human papilloma virus 16-associated
tumors, previously shown to be sensitive to the experimental DC
treatments in various settings (22–24).
Materials and methods
Mice
C57BL/6 male mice, 6–8 weeks old, were obtained from
AnLab Ltd., Prague, Czech Republic. Experimental protocols were
approved by the Institutional Animal Care Committee of the
Institute of Molecular Genetics, Prague.
Tumor cell lines
The TC-1 tumor cell line (obtained from the ATCC
collection) was developed by co-transfection of murine C57BL/6 lung
cells with HPV16 E6/E7 genes and activated (G12V) Ha-ras plasmid
DNA (25). TRAMP-C2 tumor cells
(obtained from the ATCC collection), MHC class I-deficient, were
established from a heterogeneous 32-week tumor of the transgenic
adenocarcinoma mouse prostate (TRAMP) model (26). TC-1 cells were maintained in
RPMI-1640 medium (Sigma-Aldrich GmbH, Steinheim, Germany)
supplemented with 10% FCS (PAN Biotech GmbH, Aidenbach, Germany), 2
mM L-glutamine and antibiotics; TRAMP-C2 cells were maintained in
D-MEM medium (Sigma-Aldrich) supplemented with 5% FCS, Nu-Serum IV
(5%; BD Biosciences, Bedford, MA, USA), 0.005 mg/ml human insulin
(Sigma-Aldrich), dehydroisoandrosterone (DHEA, 10 nM;
Sigma-Aldrich) and antibiotics. Both cell lines were cultured at
37ºC in a humidified atmosphere with 5% CO2 cells. In
the in vivo experiments, 5×104 TC-1 cells and
1×106 TRAMP-C2 cells were administered for the
challenge. In our hands, 5×104 TC-1 cells represent 5
TID50 doses and 1×106 TRAMP-C2 cells
represent 3 TID50 doses.
High hydrostatic pressure and irradiation
cell treatments
Tumor cells were treated by HHP (100, 150, 175 and
200 MPa) in the custom-made device (Resato International BV, Roden,
the Netherlands) that is located in the GMP manufacturing facility,
Sotio a.s. (Prague, Czech Republic). This device allows reliable
treatment of the tumor cells by defined levels of HHP for specified
periods of time (10 min in the case of 200 MPa) (13). Inactivation of tumor cells by
irradiation (150 Gy) was performed as previously described
(22).
Dendritic cell preparation
Dendritic cells (DC) were prepared from bone marrow
precursors as described by Indrová et al (24) and Lutz et al (27) with slight modifications (28). Briefly, the bone marrow cells were
cultured for 7 days in the complete RPMI-1640 medium supplemented
with 2×10−5 M mercaptoethanol (Calbiochem, La Jolla, CA,
USA), 10 ng/ml GM-CSF and IL-4 (R&D Systems, Minneapolis, MN,
USA). On day 5, the DC were pulsed with HHP-treated or irradiated
(IR-treated) tumor cells by 48-h incubation in the ratio of 2:1
(DC/tumor cells, 106 DC/ml). DC pulsed with the tumor
cells were treated for 24 h with unmethylated CpG containing
phosphorothioate-modified oligodeoxynucleotide CpG 1826
(5′-TCCATGACGTTCCTGACGTT-3′) (29)
at a final concentration of 5 μg/ml (Generi Biotech, Hradec
Králové, Czech Republic), were sulfur-modified in their backbone
(phosphorothioate) and synthesized under endotoxin-free conditions.
On day 7, non-adherent cells were harvested. These cells,
designated as DC, contained ~60–70% CD11c+ cells. For
mouse immunization experiments, DC were washed twice with PBS and
injected subcutaneously (s.c.) in PBS, 300 μl/2×106
cells/mouse.
Immunization/challenge experiments with
tumor cells
Mice were twice immunized with 5×106
irradiated tumor cells in a three-week interval (s.c., irradiation
dose was 150 Gy, HHP dose was 200 MPa) (13,30,31).
For in vivo studies, 10 days after the second immunization,
mice were challenged s.c. with corresponding tumor cells (TC-1,
5×104; TRAMP-C2, 1×106 cells/mouse). Mice
were observed twice weekly, and the numbers of tumor-bearing mice
and the size of the tumors were recorded. Two perpendicular
diameters of the tumors were measured with a caliper and the tumor
size was expressed as the tumor area (cm2). For in
vitro analyses of the immune response, three mice were
sacrificed. Single-cell suspensions from the spleens were prepared
by homogenization through a cell strainer (70 μm; BD Biosciences,
San Jose, CA, USA). Erythrocytes were osmotically lysed using
ammonium chloride-potassium lysis buffer, the cell suspension was
washed three times in the RPMI-1640 medium and used for further
analysis by FACS, chromium release assay, ELISA (IFNγ) and ELISPOT
(IFNγ).
Immunization/challenge experiments with
dendritic cells
Mice were twice immunized with 2×106
cells of DC-based vaccine in a two-week interval. For in
vivo studies, 10 days after the second immunization, mice were
challenged s.c. with corresponding tumor cells (TC-1,
5×104; TRAMP-C2, 1×106 cells/mouse). Mice
were observed twice weekly, and the numbers of tumor-bearing mice
and the size of the tumors were recorded. Two perpendicular
diameters of the tumors were measured with a caliper and the tumor
size was expressed as the tumor area (cm2). For in
vitro analyses of the immune response, three mice were
sacrificed. Single-cell suspensions from the spleens were prepared
as mentioned above and used for further analysis by FACS, chromium
release assay, ELISA (IFNγ) and ELISPOT (IFNγ).
Therapeutic experiments
The therapeutic schemes were designed for combined
chemoimmunotherapy treatment of early growing tumors. TC-1
(5×104 cells) or TRAMP-C2 (1×106 cells) tumor
cells were s.c. transplanted on day 0. Docetaxel, 30 mg/kg
(Actavis, North Bruncwik, NJ, USA) was repeatedly administered on
days 7, 21 and 35 intraperitoneally (i.p.). Dendritic cells were
administered on days 14, 28 and 42 in the vicinity of the tumor
cell challenge site or peritumorally when the growing tumors
appeared. Mice were observed twice a week and the size of the
tumors was recorded. Two perpendicular diameters of the tumors were
measured with a caliper and the tumor size was expressed as the
tumor area (cm2).
Flow cytometry
Cell surface expression of CRT, HSP90, MHC class I,
CD54 and CD80 on the tumor cells was analyzed by flow cytometry.
Tumor cells were collected from the cell culture 24 h after the HHP
or IR treatment [106 cells/ml/well, 12-well plate (Nunc,
Roskilde, Denmark)]. Cells (5×105/sample) were washed
and labeled with primary antibodies for 25 min at 4ºC, followed by
wash steps and alternatively labeled by incubation with Alexa 647-
or DyLight 649-conjugated secondary antibody for 30 min at 4ºC.
Apoptotic cells were determined by Annexin V apoptosis detection
kit (eBiosciences) according to the manufacturer's instructions.
Samples were kept in the dark and 10 min before the analysis,
Hoechst 33258 was added at a final concentration of 2 μg/ml.
Expression of cell surface molecules on the DC or spleen cells was
analyzed by flow cytometry. Cell suspensions were washed and
preincubated with anti-CD16/CD32 antibody to minimize non-specific
binding for 15 min at 4ºC following washing step and incubation
with labeled primary antibody for 30 min at 4ºC. Relevant isotype
controls of irrelevant specificity were used. FACS buffer (PBS, 1%
FBS, 0.1% NaN3) was used for all washing steps and
analysis. The following antibodies were used for FACS analyses: BD:
anti-MHC class I (PE anti-H-2Db clone KH95 and PE
anti-H-2Kb clone AF6-88.5), FITC anti-I-Ab
(Aβb) (AF6-120.1), PE anti-CD54 (3E2), PE
anti-CD80 (16-10A1), PE anti-CD86 (GL1), PE anti-CD274 (MIH5);
BioLegend, Inc. (San Diego, CA, USA): BV421 or APC anti-CD11c
(HL3), APC-CD45 (30-F11), FITC anti-CD8α (LY-2), BV711 anti-CD4
(RM4-5), PE anti-CD44 (IM7), PE-Cy7 CD62L (MEL-14); R&D Systems
(Basel, Switzerland): anti-HSP70 (242707); Abcam (Cambridge, UK):
anti-CRT (ab2907); Enzo Life Sciences, Inc. (Farmingdale, NY, USA):
anti-HSP90 (AC88). Secondary antibodies anti-mouse conjugated to
DyLight 649 (Jackson ImmunoResearch Laboratories, West Grove, PA,
USA) or anti-rabbit conjugated to Alexa 647 (Cell Signaling
Technology, Danvers, MA, USA) were also used. FACS analysis was
performed using an LSR II flow cytometer (BD Biosciences) and
analyzed by FlowJo 7.6.5 software.
Confocal microscopy
HHP-treated TC-1 and TRAMP-C2 cells were collected
and washed twice with PBS. The cells were then incubated for 30 min
with primary anti-CRT antibody (FMC 75; Enzo Life Sciences) diluted
in PBS, followed by washing and incubation with the Alexa Fluor 488
goat anti-mouse secondary antibody (Molecular Probes). Cells were
washed twice with PBS and fixed in 4% paraformaldehyde for 20 min
and mounted on slides. Cells were examined under a DMI 6000
inverted Leica TCS AOBS SP5 tandem scanning confocal microscope
with an AR (488 nm) laser and an ×63 oil immersion objective.
ELISA
For HMGB1 release, supernatants from the tumor cell
culture were collected 24 h after HHP treatment (106
cell/ml/well, 12-well plate (Nunc) and analyzed by an ELISA kit
(IBL International GmbH, Hamburg, Germany) according to the
manufacturer's instructions. For IL-1β, IL-6, IFNγ and IL-12
production, supernatants from the DC culture were collected 24 h
after the addition of CpG 1826 and analyzed by ELISA kits (BD
Biosciences) according to the manufacturer's instructions. For IFNγ
production, supernatants from the spleen single-cell suspension
were collected after 48-h incubation [2×106
cell/ml/well, 12-well plate (Nunc)] and analyzed by an ELISA kit
(BD Biosciences) according to the manufacturer's instructions.
ELISPOT
To determine the amount of IFNγ-secreting cells, an
ELISPOT kit for detection of murine IFNγ (BD Biosciences, San
Diego, CA, USA) was used. Spleen cells were cultured for 48 h and
then placed into the wells of ELISPOT plates (concentration
5×105, 1×105 and 5×104 cells/well)
for 24 h. The plates were then processed according to the
manufacturer's instructions (BD Biosciences). Colored spots were
counted with CTL Analyzer LLC (CTL, Cleveland, OH, USA) and
analyzed using ImmunoSpot Image Analyzer software.
Chromium release microcytotoxicity
assay
The cytolytic activity of effector cells was tested
in 18-h 51Cr release assay, as previously described
(32,33). Briefly, spleen cells from control
and immunized mice that served as effector cells were treated with
ammonium chloride-potassium lysing buffer (1 min) to deplete
erythrocytes. The mixtures of effector cells with
51Cr-labeled tumor targets were incubated in selected
target/effector cell ratios (1:25, 1:50, 1:100 and 1:200) in
triplicate in 96-well round bottom microtiter plates (Nunc). The
percentage of specific 51Cr release was expressed
according to the formula: [cpm experimental release - cpm control
release/cpm maximum release/cpm control release] × 100.
Statistical analyses
For statistical analyses of in vitro
experiments, Student's t-test was used. For evaluation of in
vivo experiments, analysis of variance (ANOVA) from the NCSS,
Number Cruncher Statistical System (NCSS, LLC, Kaysville, UT, USA)
statistical package was utilized. Standard deviations are indicated
in the figures.
Results
HHP, but not IR, induces ICD markers on
both TC-1 and TRAMP-C2 tumor cells
First, we determined the ability of HHP to induce
ICD in murine TC-1 or TRAMP-C2 cells, and then we compared the
effect of HHP to the effect of irradiation (150 Gy) that has been
standardly used for treatment of cells during preparation of DC
vaccines in our previous studies (22). Fig.
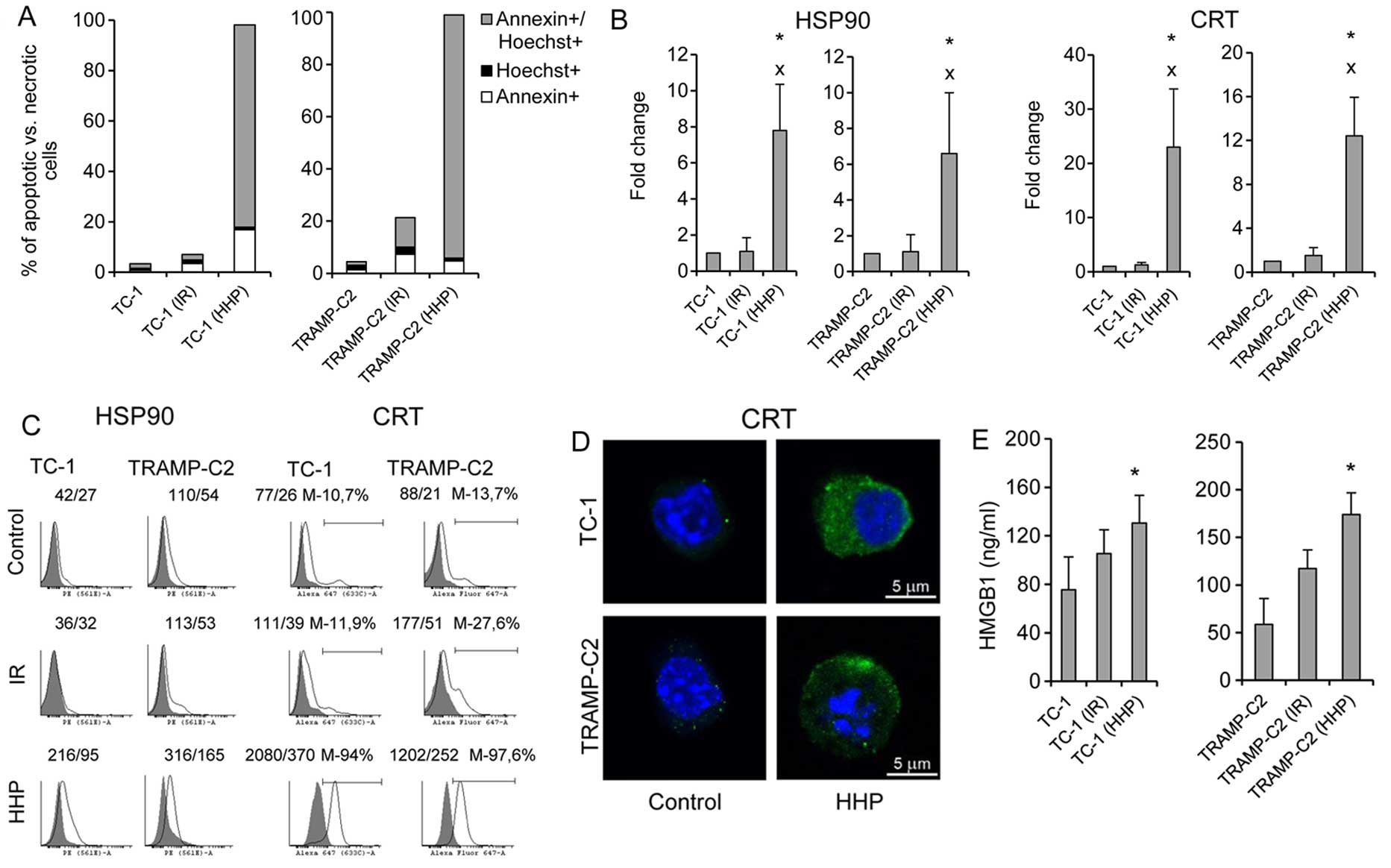
1A shows that the percentage of late apoptotic/dead tumor cells
(Annexin V+/Hoechst+) after the treatment
with 200 MPa HHP was >80% within 24 h. The presence of ICD
markers HSP90 and CRT on the cell surface of the tested cells was
also significantly increased (Fig. 2B
and C). Fluorescence microscopy images of HHP-treated tumor
cells stained for CRT confirmed increased expression of CRT after
HHP treatment (Fig. 1D). Release
of HMGB1, late-stage ICD marker, in the supernatant was further
analyzed. Fig. 1E demonstrates a
significant increase of HMGB1 in the tumor cell supernatants after
HHP treatment. Induction of ICD by 200 MPa HHP was similar both in
TC-1 and TRAMP-C2 tumor cells. No significant upregulation of ICD
markers was detected after irradiation with 150 Gy. The treatment
with HHP of 200 MPa was selected as it was the most effective in
inducing apoptosis and expression of ICD markers and simultaneously
arresting cell proliferation, as determined by colony-forming assay
in experiments, in which the effects of different doses of HHP
(100, 150, 175, 200 and 250 MPa) were compared (data not
shown).
Prophylactic immunization with
HHP-treated tumor cells induces higher immune responses in mice
when compared with IR-treated tumor cells both in TC-1 and TRAMP-C2
tumor models
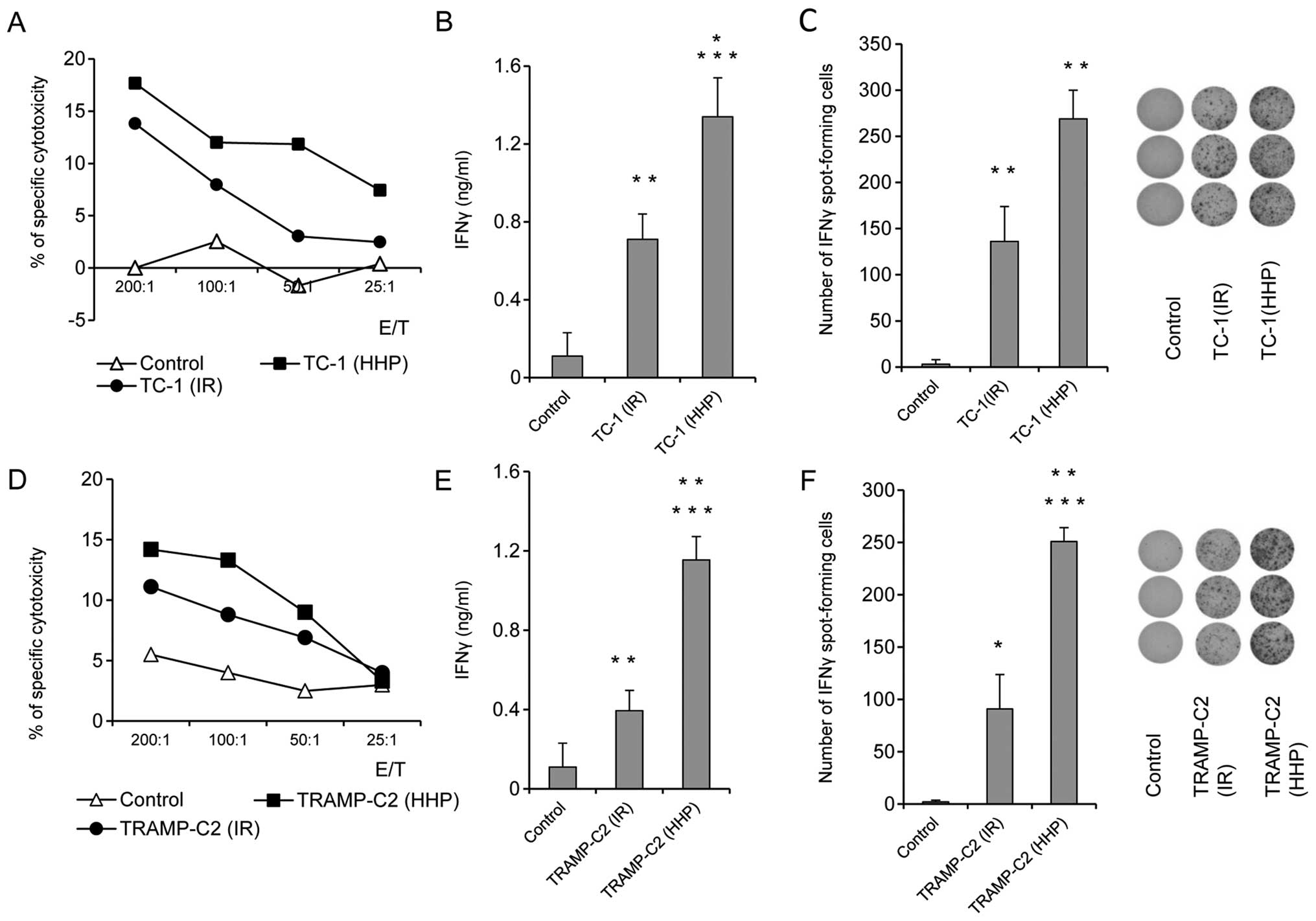
To study the ability of HHP and IR-treated tumor
cells to induce immune response and their antitumor potency, mice
were immunized twice at a three-week interval with 5×106
HHP- or IR-treated TC-1 or TRAMP-C2 tumor cells. Ten days after the
second immunization, mice were challenged with relevant tumor cells
in doses of 5×104 TC-1 or 106 TRAMP-C2. Three
mice from each group were left without challenge and used for
parallel in vitro analyses. In vitro analyses of the
spleen effector cells prepared ten days after the second
immunization with HHP-treated TC-1 or TRAMP-C2 tumor cells showed
an increased cytotoxic effect of spleen effector cells on the
corresponding targets. In the group of mice immunized with
IR-treated tumor cells, a similar but slightly lower effect was
observed (Fig. 2A and D). Despite
the fact that the analysis of the spleen effector cells after
immunization with HHP-treated tumor cells showed only moderately
augmented cytotoxic effect when compared to immunization with
IR-treated tumor cells, analysis of IFNγ production revealed
significant differences. Compared to the IR-treated tumor cells,
mice immunized with HHP-treated tumor cells displayed significantly
increased IFNγ production by spleen cells measured by the ELISA
assay (Fig. 2B and E) and
significantly increased number of IFNγ-producing cells detected by
the ELISPOT assay (Fig. 2C and F).
These results were similar in both tumor models, immunogenic TC-1
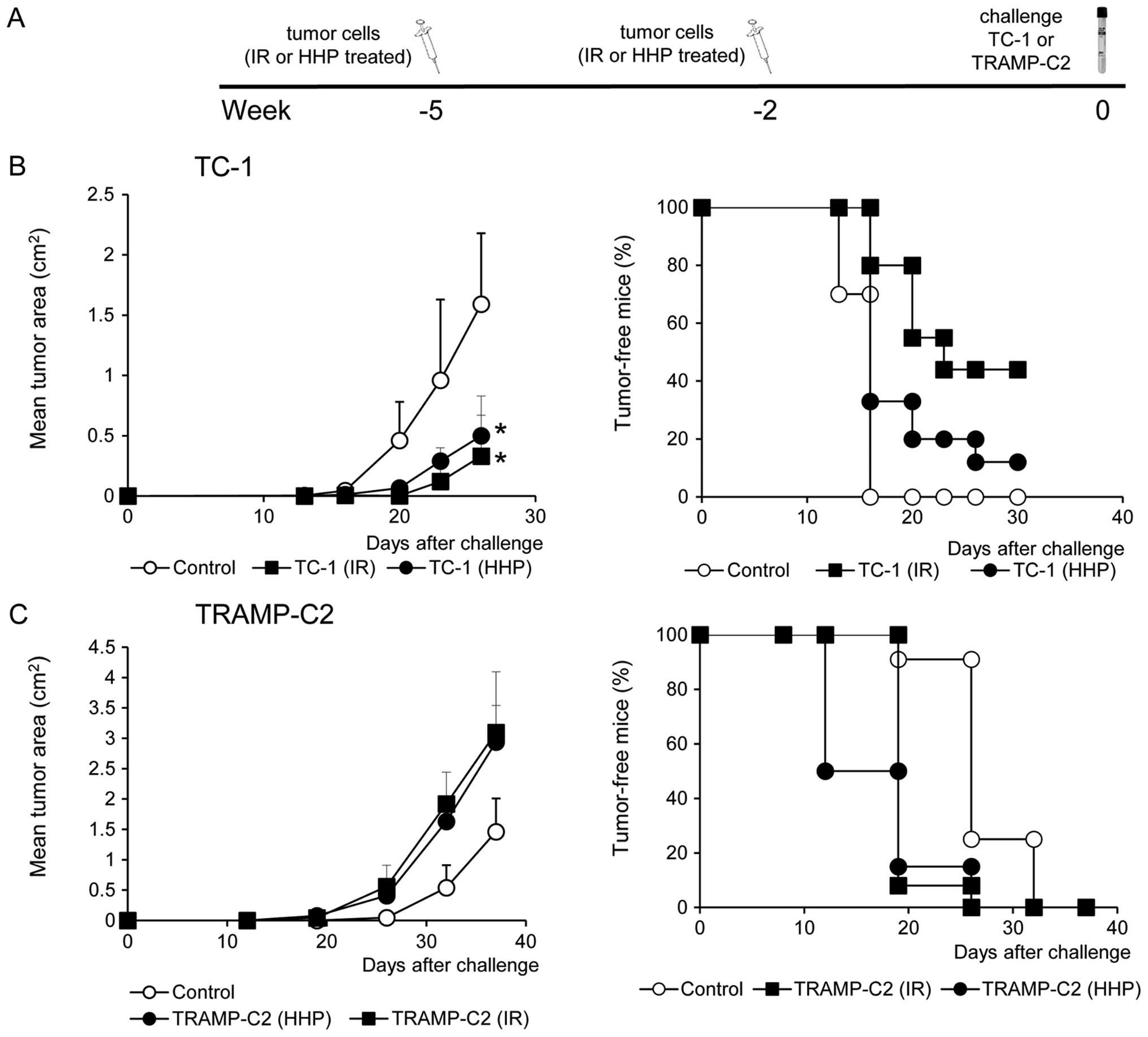
and weakly immunogenic TRAMP-C2. However, after the challenge of
immunized mice with the corresponding tumor cells, significant
inhibition (P<0.05) of tumor growth was recorded only in the
groups of mice immunized with the HHP or IR-treated TC-1 tumor
cells and challenged with corresponding TC-1 cells (Fig. 3B). In contrast, mice immunized with
HHP and IR-treated TRAMP-C2 cells did not exhibit any inhibition of
tumor growth after the challenge with TRAMP-C2 cells (Fig. 3C).
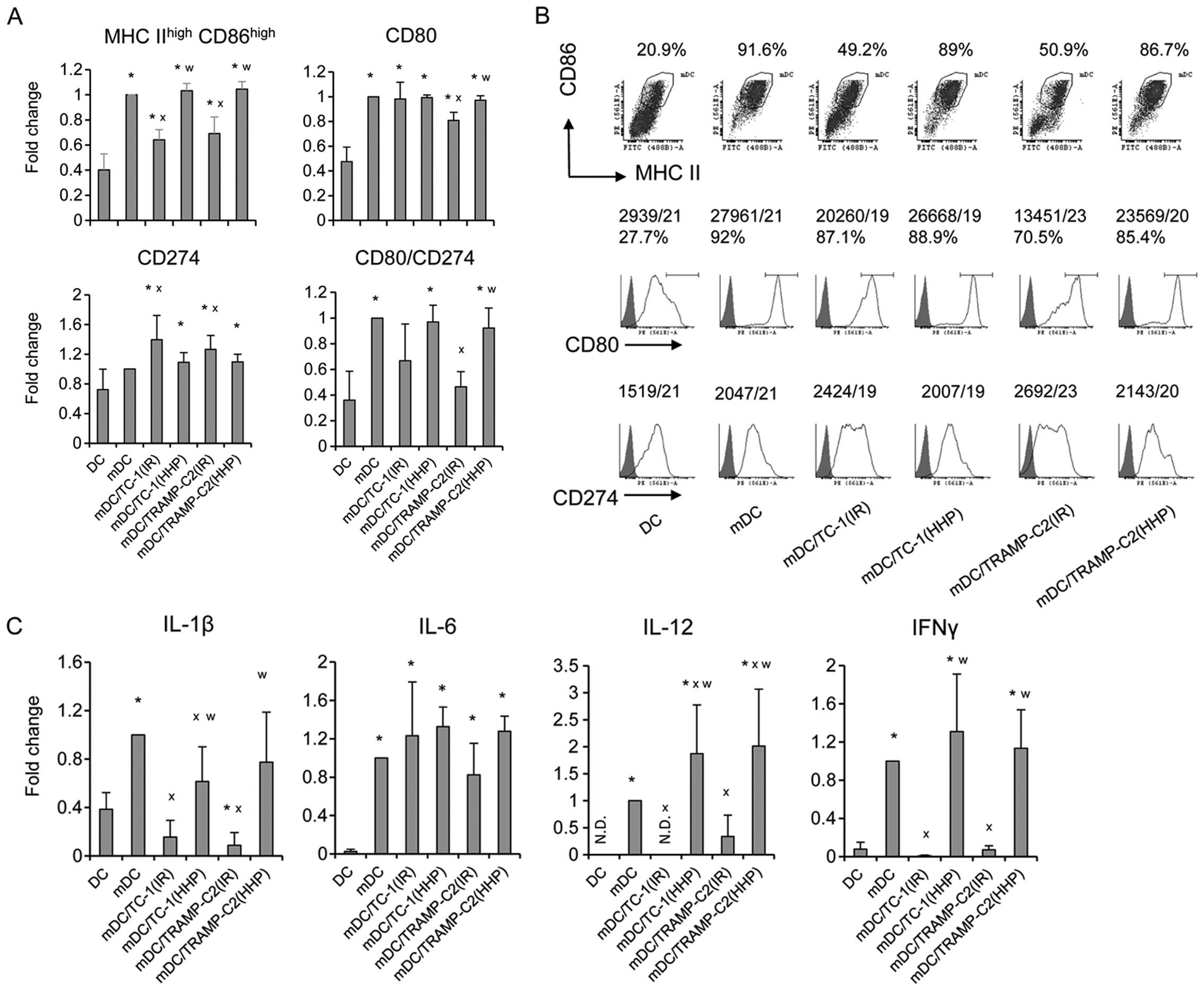
Pulsing with HHP-treated TC-1 or TRAMP-C2
tumor cells increased expression of maturation markers on DC and
stimulated production of cytokines characteristic for matured
DC
Next, the HHP-treated TC-1 and TRAMP-C2 cells were
used for DC pulsing, and the phenotypes of matured DC vaccines,
unpulsed or pulsed with the IR-treated tumor cells or HHP-treated
tumor cells, were compared (Fig.
4). We did not see any significant differences between unpulsed
cells and HHP-treated tumor cells-pulsed DC. In both cases, CpG
ODN1826-mediated maturation increased the proportion of matured MHC
class IIhigh/CD86high dendritic cells and
increased CD80 and CD274 cell surface expression (Fig. 4A and B). The ratio between the CD80
and CD274 cell surface expressions (demonstrated by MFIvalues) was
higher on matured cells compared to the immature controls (Fig. 4A). Both unpulsed cells and
HHP-treated tumor cells-pulsed matured DC produced IL-12, as well
as IL-1β, IL-6 and IFNγ (Fig. 4C).
HHP-treated tumor cell-pulsed matured DC produced significantly
higher amounts of IL-12 and IFNγ as compared to the unpulsed cells.
No significant differences were observed between TRAMP-C2 and TC-1
cell co-culture. On the other hand, pulsing with the IR-treated
tumor cells resulted in reduction of the proportion of matured MHC
class IIhigh/CD86high dendritic cells in the
DC populations, decreased the ratio between the CD80 and CD274 cell
surface expression, and also significantly inhibited IL-12, IFNγ
and IL-1β production, as compared to both unpulsed cells and
HHP-treated tumor cell-pulsed matured DC (Fig. 4). These results indicate that DC
co-culture with IR-treated, but not HHP-treated tumor cells, can
impair DC maturation.
Prophylactic immunization with DC-based
vaccine pulsed with HHP-treated TC-1 or TRAMP-C2 tumor cells
induces strong immune response, but inhibits growth of TC-1 tumors
only
In the next series of experiments, HHP-treated tumor
cell-pulsed matured DC were investigated in vivo.
HHP-treated tumor cell-pulsed matured DC were selected for further
in vivo experiments as pulsing of DC with IR-treated tumor
cells negatively affected DC maturation in terms of expression of
costimulatory molecules and production of selected cytokines. Mice
were immunized twice in a 2-week interval with 2×106
HHP-treated tumor cell-pulsed matured DC. Ten days after the second
immunization, mice were challenged with relevant tumor cells, in
doses of 5×104 TC-1 or 106 TRAMP-C2 tumor
cells. Three mice from each group were left without challenge and
used for parallel in vitro analyses. Both HHP-treated
TRAMP-C2 and TC-1 cells pulsed DC vaccines induced strong immune
responses, as determined by spleen cell analysis performed ten days
after the second immunization (Fig.
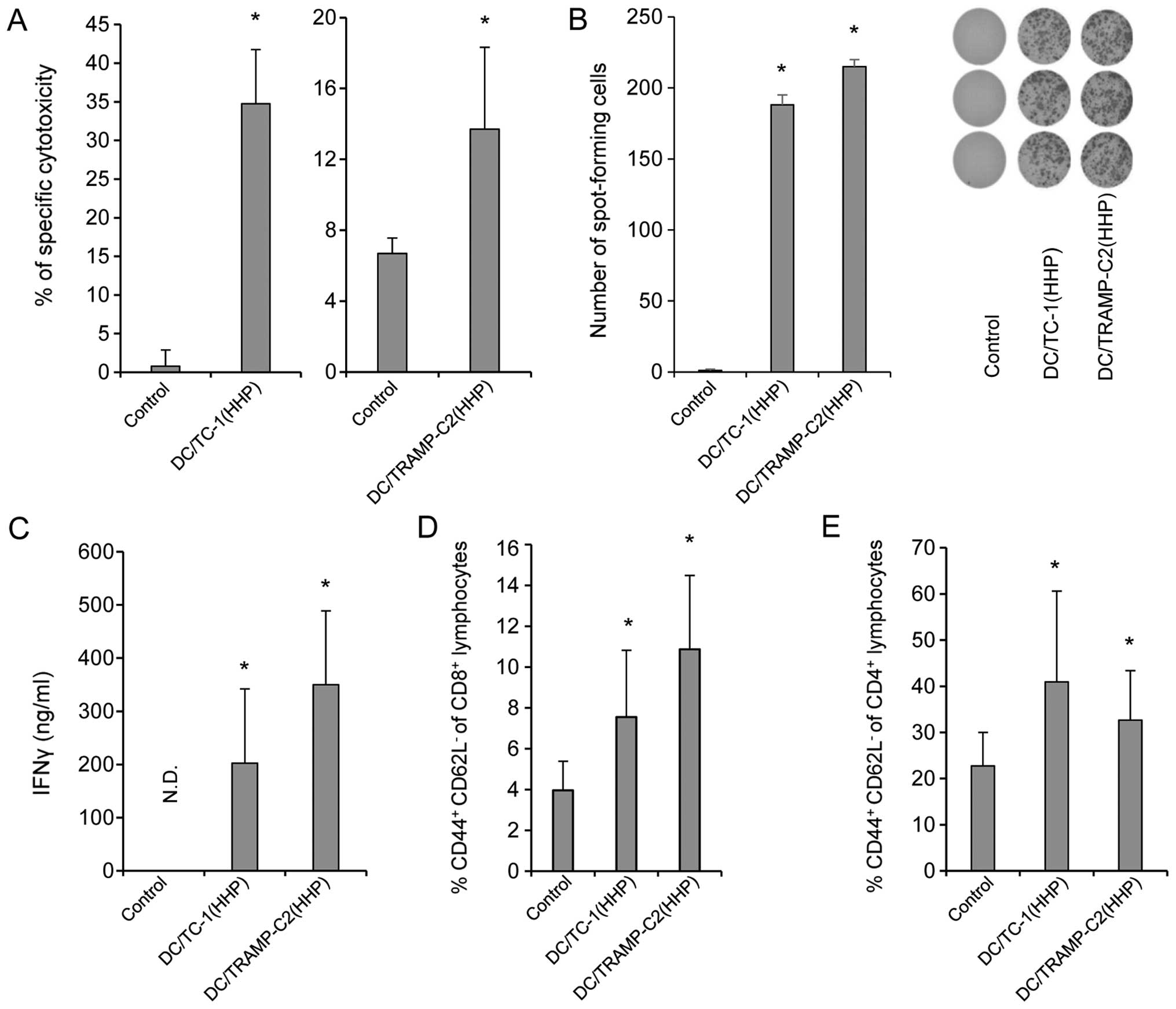
5). Immunization with HHP-treated TC-1 or TRAMP-C2 pulsed DC
vaccines showed a significantly increased cytotoxic effect of
spleen effector cells on the corresponding targets (Fig. 5A). As compared with control mice,
mice immunized with both HHP-treated TRAMP-C2 and TC-1 cells pulsed
DC vaccines displayed significantly increased numbers of
IFNγ-producing cells detected by ELISPOT assay (Fig. 5B) and significantly increased IFNγ
production by spleen cells measured by ELISA assay (Fig. 5C). A significant increase was also
found in the percentage of CD4+ and CD8+
CD44+ CD62L− T lymphocytes (Fig. 5D and E). These results were similar
in both tumor models, immunogenic TC-1 and weakly immunogenic
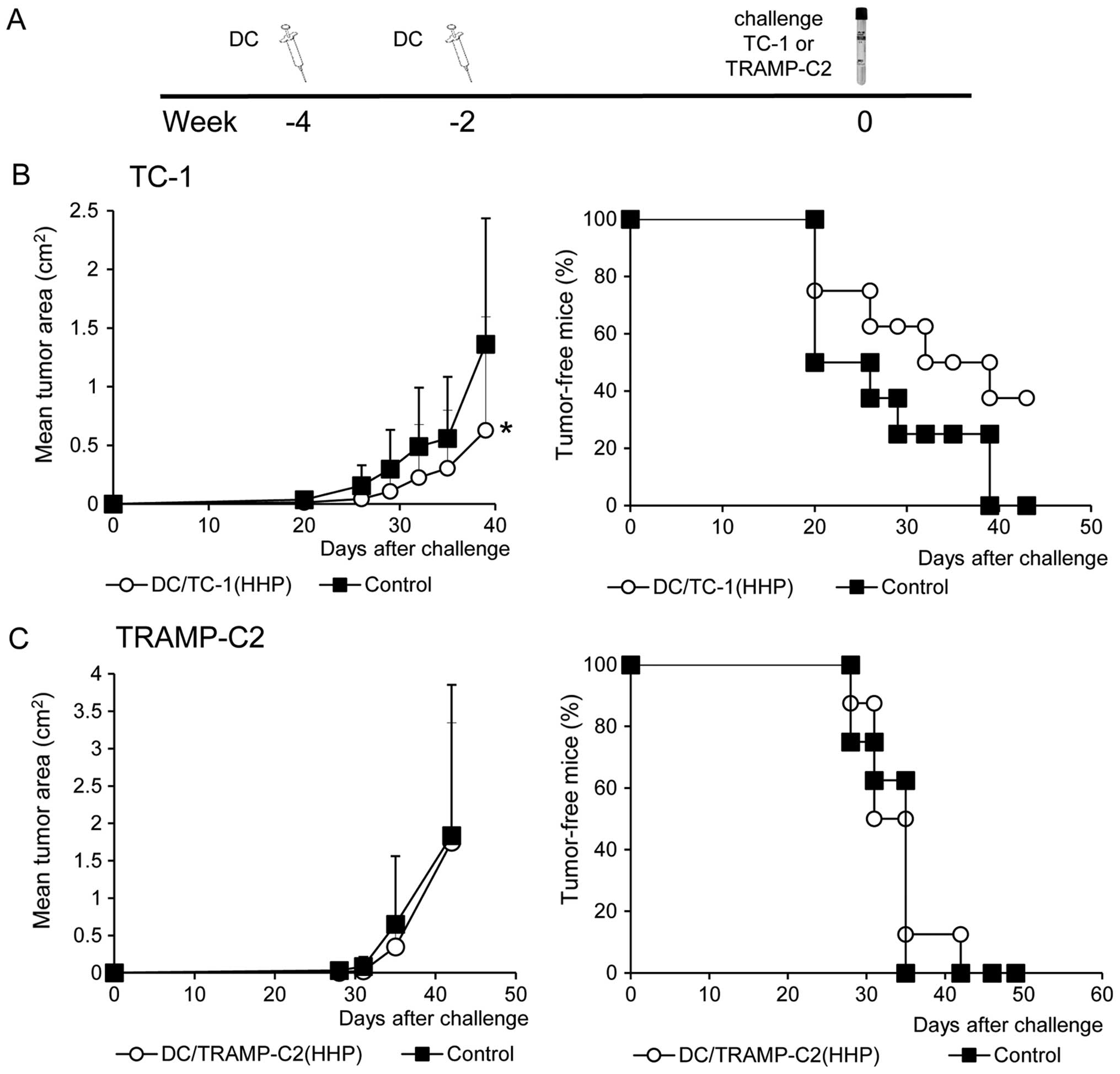
TRAMP-C2. Contrary to the results in vitro, in vivo
analysis showed significant inhibition (P<0.05) of the tumor
growth only in the group of mice immunized with the HHP-treated
TC-1 tumor cell-pulsed matured DC and challenged with corresponding
TC-1 cells (Fig. 6B). Mice
immunized with the HHP-treated TRAMP-C2 tumor cell-pulsed matured
DC did not exhibit any inhibition of tumor growth after the
challenge with TRAMP-C2 cells (Fig.
6C). In both experiments, the percentage of tumor-free mice are
shown in the right panel.
Combined chemoimmunotherapy of TC-1 and
TRAMP-C2 tumors with docetaxel and DC-based vaccine significantly
inhibits growth of subcutaneous tumors
The therapeutic efficacy of HHP-treated tumor
cell-pulsed matured DC was then tested in the therapeutic setting
when a combination of chemotherapy and immunotherapy with DC-based
vaccine was employed. TC-1 (5×104 cells) or TRAMP-C2
(106 cells) tumor cells were s.c. transplanted on day 0
and treated with three doses of docetaxel chemotherapy in a 2-week
interval. Dendritic cells were administered at regular intervals
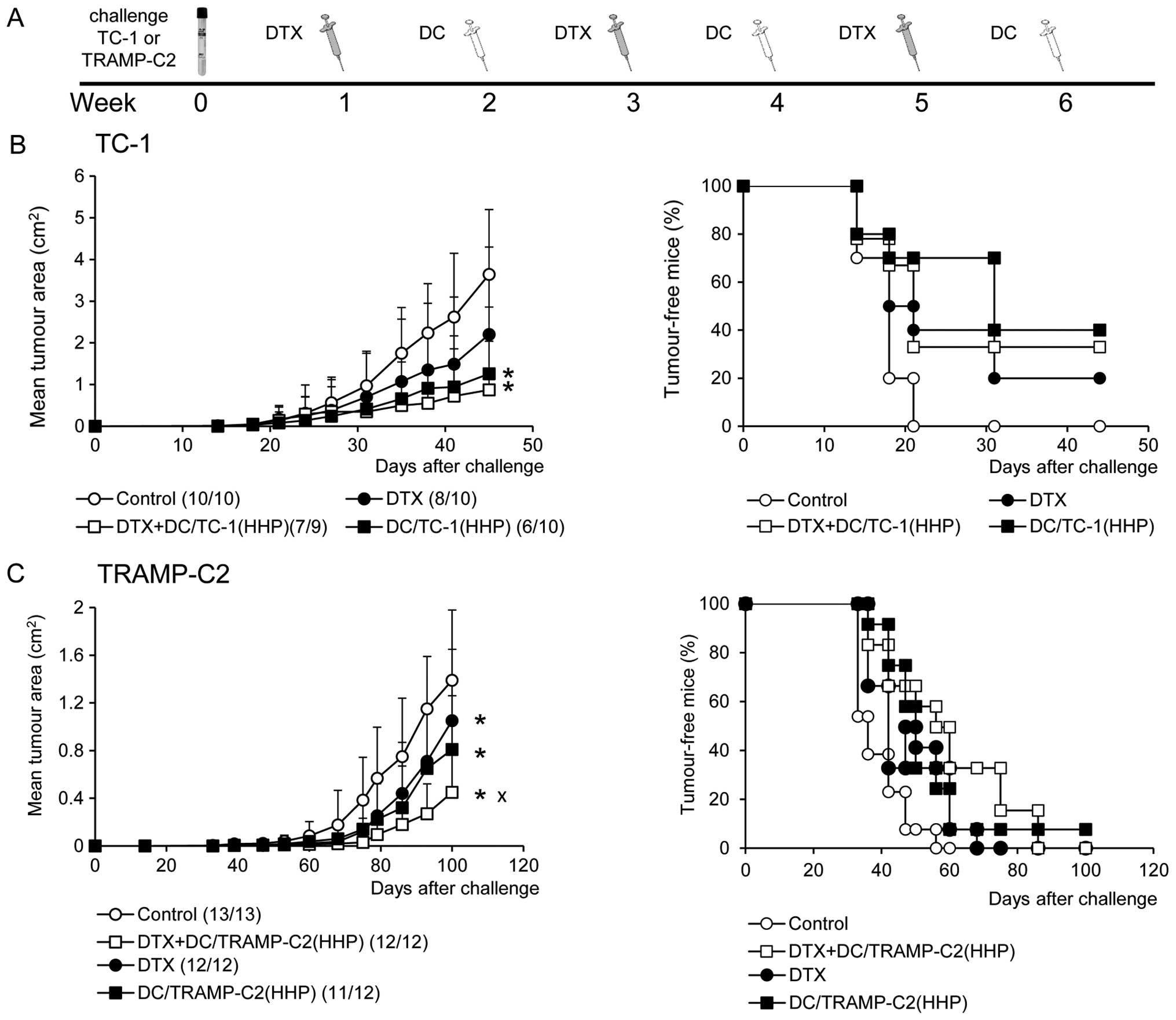
between the docetaxel chemotherapy. As shown in Fig. 7B, the growth of immunogenic TC-1
tumors was significantly inhibited by the treatment with DC alone
or with the combination of docetaxel and DC vaccine [DC/TC-1(HHP)]
(P<0.05 vs. control). Docetaxel alone delayed the growth of
tumors, but no significant difference was evident. A representative
experiment of two independent ones is given in Fig. 7B. When the incidences of tumors in
mice from two performed experiments were merged [Control 19/19,
docetaxel 18/20, docetaxel + DC/TC-1(HHP) 14/19, DC/TC-1-HHP 6/10],
the only significant difference was found between the control group
and the group of combined chemoimmunotherapy (χ2;
docetaxel +DC/TC-1(HHP) vs. Control P<0.01). These results
indicate the beneficial effect of the combination of chemotherapy
with immunotherapy. The same therapeutic setting was also used for
the treatment of poorly immunogenic TRAMP-C2 tumors. As shown in
Fig. 7C, monotherapies with
docetaxel alone or DC/TRAMP-C2(HHP) vaccine alone significantly
inhibited growth of TRAMP-C2 tumors. However, when these
monotherapies were combined, the therapeutic effect was even
stronger. Significant inhibition of tumor growth was found between
docetaxel alone or DC/TRAMP-C2(HHP) alone groups and the group
treated with a combination of docetaxel and DC/TRAMP-C2(HHP)
vaccine (P<0.05). The tumor-inhibitory effect was noted as
reduction of the size of growing tumors; there was no difference
between the incidences of tumors when two independent experiments
were merged [Control 25/26, docetaxel 22/22, docetaxel +
DC/TRAMP-C2(HHP) 22/22, DC/TRAMP-C2(HHP) 21/22].
Discussion
HHP has been previously shown to induce endoplasmic
reticulum stress and consequently ICD in both murine and human cell
lineages (11,13,34).
This suggests that HHP, along with other modalities, such as
irradiation, photodynamic therapy using hypericin, hyperthermia or
treatments with selected chemotherapeutic and cytotoxic agents, can
be used for preparation of tumor cells capable of inducing
effective antitumor immunity (35). HHP could also be used for tumor
cell inactivation before their use as cellular vaccines or as
antigen donors in DC-based vaccines.
In the first part of the study, our aim was to
demonstrate the capability of HHP-treated tumor cells to induce
immune responses in mice, in comparison with irradiated tumor
cells. Lethal irradiation represents a standard procedure used for
tumor cell inactivation before their usage for immunization or for
DC pulsing, and HHP treatment can serve as an attractive
alternative for this procedure. Before performing the in
vivo experiments, we optimized the HHP treatments of TC-1 and
TRAMP-C2 cell lines used for the studies and we demonstrated, in
comparative experiments, that HHP induced higher levels of CRT and
HSP90 expression on tumor cells, as well as HMGB1 production, as
compared to irradiation. The immunogenicity of irradiated and
HHP-treated cells was further monitored in vivo and we noted
higher IFNγ production by spleen cells upon immunization with the
HHP-treated compared to irradiated cells. However, we did not
observe significantly higher cytotoxicity of the spleen cells from
the animals immunized with HHP-treated cells and this finding was
in agreement with the results of immunization-challenge
experiments. We did not see any significant differences between
vaccination with HHP- or IR-treated tumor cells; both vaccinations
inhibited TC-1 tumor growth, as expected and previously observed
for the animals immunized with IR-treated cells (30,31)
while the TRAMP-C2 tumor growth was not blocked by both of the
vaccination protocols. It has been previously shown that TRAMP-C2
tumor cells are not immunogenic, unless their immunogenicity was
increased by IFNγ treatment, inducing MHC class I cell surface
expression (36). Thus, it seems
that HHP treatment, which induces ICD but not MHC class I and
co-stimulatory molecule cell surface expression, does not induce
protective immunity effective against TRAMP-C2 cells. It is of note
that in the case of TC-1 tumors, which are apparently more
sensitive to immune responses, effective immunity was induced by
vaccination with both irradiated and HHP-treated TC-1 cells.
Furthermore, in order to assess the suitability of
HHP as a tool for tumor cell preparation in the DC-based vaccine
preparation protocols, we prepared a DC-based vaccine by co-culture
of immature DC with HHP-treated TC-1 or TRAMP-C2 tumor cells and
subsequent DC maturation with CpG ODN 1826. CpG ODN 1826, an
agonist of Toll-like receptor 9, is a potent maturation agent for
murine DC (37), and the
capability of DC pulsed by co-cultivation with irradiated tumor
cells and matured by CpG ODN 1826 to inhibit the TC-1 tumor growth
has also been demonstrated in our laboratory (38). We have compared the phenotype of
matured DC vaccines, unpulsed or pulsed with the IR- or HHP-treated
tumor cells. The results suggest that DC co-culture with
irradiated, but not HHP-treated tumor cells, interferes with their
subsequent CpG ODN-driven maturation, since the matured DC culture
of the cells pulsed with IR-treated cells displayed lower
proportion of matured DC (defined as MHC class
IIhigh/CD86high), and lower ratio between the
expression of positive costimulatory molecule CD80 (B7.1) vs.
negative costimulatory molecule CD274 (B7-H1). This ratio can be
considered as an important marker suggesting the DC capability to
transmit positive signaling to T cells (39). Selected cytokine expression levels,
including that of IL-12, were lower in DC pulsed with IR-treated
tumor cells, as compared to the unpulsed controls. Notably, this
was not observed when the HHP-treated tumor cells DC were compared
to the unpulsed controls. These results suggest that immature DC
co-culture with HHP-treated cells represents a convenient protocol
for the DC-based vaccine preparation and corroborates previous
findings of Fucikova et al (13).
The next step was therefore to perform in
vivo experiments and to evaluate the immunogenicity of the
matured DC pulsed with the HHP-treated tumor cells. As expected, DC
vaccines induced much higher IFNγ production by spleen cells as
compared to immunization with tumor cells. This, together with
further parameters investigated in the spleens (chromium release
assay, effector memory CD4 and CD8 cell proportion), suggested that
DC vaccines induced strong immunity against TC-1 or TRAMP-C2
tumors, respectively. However, as determined in the
immunization-challenge experiments, DC vaccination in a
prophylactic setting induced protection against TC-1, but not
TRAMP-C2 tumor growth. This was in agreement with IR- and
HHP-treated tumor cell immunization, confirming different
immunogenicity or sensitivity of the TC-1 and TRAMP-C2 tumors to
the immune response induced by prophylactic immunization.
Next, we tested the vaccine efficacy in a
therapeutic setting in combination with docetaxel chemotherapy,
which is clinically relevant especially for prostate cancer
treatment (19,20). DC-based vaccines are in general
intended to be used rather for tumor immunotherapy in a multimodal
setting than for immunization. In our experiments, unlike in
prophylactic use, the DC treatments of both immunogenic TC-1 and
poorly immunogenic/treatable TRAMP-C2 tumors resulted in
significant inhibition of the tumor growth, albeit the effect on
the TRAMP-C2 appeared to be weaker as compared to the TC-1 tumors.
The difference was observed for the therapeutic protocol using
docetaxel and DC combination. This treatment led to the highest
therapeutic effect, as compared to the chemotherapy or
immunotherapy only treatments, in the case of the TRAMP-C2 prostate
cancer model. In this model, both chemo- and immunotherapy, when
used as monotherapies, displayed only moderate antitumor effects,
and additive/synergistic effects were observed when these
treatments were used in combination. On the contrary, synergistic
effects of the combination therapy were not seen for the TC-1
therapy. We can speculate that TC-1 tumors are much more vulnerable
to immunotherapy, as compared to the TRAMP-C2 tumors, and that it
may be difficult to boost it. Moreover, DTX treatment can increase
the TRAMP-C2 tumor cell sensitivity to the immune responses.
In conclusion, in the present study we demonstrated
that HHP-treatment induced ICD in the cells of TRAMP-C2 and TC-1
murine tumor cell lines. Furthermore, our results show that
DC-based vaccines pulsed with HHP-treated cells is an effective
instrument for immunotherapy, mainly when combined with
chemotherapy, as has been demonstrated in the prostate cancer
TRAMP-C2 model, which is poorly immunogenic and difficult to
treat.
Acknowledgements
The present study was supported by research grant
provided by SOTIO a.s., and in part by MEYS (LM2011032), the
Academy of Sciences of the Czech Republic (RVO 68378050), the
project ‘BIOCEV-Biotechnology and Biomedicine Centre of the Academy
of Sciences and Charles University’ (CZ.1.05/1.1.00/02.0109) from
the European Regional Development Fund. The authors are grateful to
Mrs. Renáta Turečková for skillful technical assistance and to Dr
Šárka Takáčová for editorial help. Conflict of interest: J.
Bartůňková and R. Špišek are employees and shareholders of SOTIO
a.s.
References
|
1
|
Ramakrishnan R, Antonia S and Gabrilovich
DI: Combined modality immunotherapy and chemotherapy: A new
perspective. Cancer Immunol Immunother. 57:1523–1529. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nowak AK, Lake RA and Robinson BW:
Combined chemoimmunotherapy of solid tumours: Improving vaccines?
Adv Drug Deliv Rev. 58:975–990. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Banchereau J and Steinman RM: Dendritic
cells and the control of immunity. Nature. 392:245–252. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Palucka K and Banchereau J: Cancer
immunotherapy via dendritic cells. Nat Rev Cancer. 12:265–277.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Galluzzi L, Senovilla L, Vacchelli E,
Eggermont A, Fridman WH, Galon J, Sautès-Fridman C, Tartour E,
Zitvogel L and Kroemer G: Trial watch: Dendritic cell-based
interventions for cancer therapy. Oncoimmunology. 1:1111–1134.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Podrazil M, Horvath R, Becht E, Rozkova D,
Bilkova P, Sochorova K, Hromadkova H, Kayserova J, Vavrova K,
Lastovicka J, et al: Phase I/II clinical trial of dendritic-cell
based immunotherapy (DCVAC/PCa) combined with chemotherapy in
patients with metastatic, castration-resistant prostate cancer.
Oncotarget. 6:18192–18205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fucikova J, Kralikova P, Fialova A,
Brtnicky T, Rob L, Bartunkova J and Spísek R: Human tumor cells
killed by anthracyclines induce a tumor-specific immune response.
Cancer Res. 71:4821–4833. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hato SV, Khong A, de Vries IJ and
Lesterhuis WJ: Molecular pathways: The immunogenic effects of
platinum-based chemo-therapeutics. Clin Cancer Res. 20:2831–2837.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Spisek R, Charalambous A, Mazumder A,
Vesole DH, Jagannath S and Dhodapkar MV: Bortezomib enhances
dendritic cell (DC)-mediated induction of immunity to human myeloma
via exposure of cell surface heat shock protein 90 on dying tumor
cells: Therapeutic implications. Blood. 109:4839–4845. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Galluzzi L, Kepp O and Kroemer G:
Immunogenic cell death in radiation therapy. Oncoimmunology.
2:e265362013. View Article : Google Scholar
|
|
11
|
Weiss EM, Frey B, Rödel F, Herrmann M,
Schlücker E, Voll RE, Fietkau R and Gaipl US: Ex vivo- and in
vivo-induced dead tumor cells as modulators of antitumor responses.
Ann NY Acad Sci. 1209:109–117. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Frey B, Rubner Y, Kulzer L, Werthmöller N,
Weiss EM, Fietkau R and Gaipl US: Antitumor immune responses
induced by ionizing irradiation and further immune stimulation.
Cancer Immunol Immunother. 63:29–36. 2014. View Article : Google Scholar
|
|
13
|
Fucikova J, Moserova I, Truxova I,
Hermanova I, Vancurova I, Partlova S, Fialova A, Sojka L, Cartron
PF, Houska M, et al: High hydrostatic pressure induces immunogenic
cell death in human tumor cells. Int J Cancer. 135:1165–1177. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Emens LA: Chemoimmunotherapy. Cancer J.
16:295–303. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ménard C, Martin F, Apetoh L, Bouyer F and
Ghiringhelli F: Cancer chemotherapy: Not only a direct cytotoxic
effect, but also an adjuvant for antitumor immunity. Cancer Immunol
Immunother. 57:1579–1587. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
van Dodewaard-de Jong JM, Verheul HM,
Bloemendal HJ, de Klerk JM, Carducci MA and van den Eertwegh AJ:
New treatment options for patients with metastatic prostate cancer:
What is the optimal sequence? Clin Genitourin Cancer. 13:271–279.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Machiels JP, Reilly RT, Emens LA, Ercolini
AM, Lei RY, Weintraub D, Okoye FI and Jaffee EM: Cyclophosphamide,
doxorubicin, and paclitaxel enhance the antitumor immune response
of granulocyte/macrophage-colony stimulating factor-secreting
whole-cell vaccines in HER-2/neu tolerized mice. Cancer Res.
61:3689–3697. 2001.PubMed/NCBI
|
|
18
|
Malvicini M, Rizzo M, Alaniz L, Piñero F,
García M, Atorrasagasti C, Aquino JB, Rozados V, Scharovsky OG,
Matar P, et al: A novel synergistic combination of cyclophosphamide
and gene transfer of interleukin-12 eradicates colorectal carcinoma
in mice. Clin Cancer Res. 15:7256–7265. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tannock IF, de Wit R, Berry WR, Horti J,
Pluzanska A, Chi KN, Oudard S, Théodore C, James ND, Turesson I, et
al; TAX 327 Investigators. Docetaxel plus prednisone or
mitoxantrone plus prednisone for advanced prostate cancer. N Engl J
Med. 351:1502–1512. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Petrylak DP, Tangen CM, Hussain MH, Lara
PN Jr, Jones JA, Taplin ME, Burch PA, Berry D, Moinpour C, Kohli M,
et al: Docetaxel and estramustine compared with mitoxantrone and
prednisone for advanced refractory prostate cancer. N Engl J Med.
351:1513–1520. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kantoff PW, Higano CS, Shore ND, Berger
ER, Small EJ, Penson DF, Redfern CH, Ferrari AC, Dreicer R, Sims
RB, et al; IMPACT Study Investigators. Sipuleucel-T immunotherapy
for castration-resistant prostate cancer. N Engl J Med.
363:411–422. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Reinis M, Indrová M, Mendoza L, Mikysková
R, Bieblová J, Bubeník J and Símová J: HPV16-associated tumours:
Therapy of surgical minimal residual disease with dendritic
cell-based vaccines. Int J Oncol. 25:1165–1170. 2004.PubMed/NCBI
|
|
23
|
Reinis M, Stepanek I, Simova J, Bieblova
J, Pribylova H, Indrova M and Bubenik J: Induction of protective
immunity against MHC class I-deficient, HPV16-associated tumours
with peptide and dendritic cell-based vaccines. Int J Oncol.
36:545–551. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Indrová M, Reinis M, Bubeník J, Jandlová
T, Bieblová J, Vonka V and Velek J: Immunogenicity of dendritic
cell-based HPV16 E6/E7 peptide vaccines: CTL activation and
protective effects. Folia Biol (Praha). 50:184–193. 2004.
|
|
25
|
Lin KY, Guarnieri FG, Staveley-O'Carroll
KF, Levitsky HI, August JT, Pardoll DM and Wu TC: Treatment of
established tumors with a novel vaccine that enhances major
histocompatibility class II presentation of tumor antigen. Cancer
Res. 56:21–26. 1996.PubMed/NCBI
|
|
26
|
Foster BA, Gingrich JR, Kwon ED, Madias C
and Greenberg NM: Characterization of prostatic epithelial cell
lines derived from transgenic adenocarcinoma of the mouse prostate
(TRAMP) model. Cancer Res. 57:3325–3330. 1997.PubMed/NCBI
|
|
27
|
Lutz MB, Kukutsch N, Ogilvie AL, Rössner
S, Koch F, Romani N and Schuler G: An advanced culture method for
generating large quantities of highly pure dendritic cells from
mouse bone marrow. J Immunol Methods. 223:77–92. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stepanek I, Indrova M, Bieblova J,
Fucikova J, Spisek R, Bubenik J and Reinis M: Effects of
5-azacytidine and trichostatin A on dendritic cell maturation. J
Biol Regul Homeost Agents. 25:517–529. 2011.
|
|
29
|
Yi AK and Krieg AM: CpG DNA rescue from
anti-IgM-induced WEHI-231 B lymphoma apoptosis via modulation of I
kappa B alpha and I kappa B beta and sustained activation of
nuclear factor-kappa B/c-Rel. J Immunol. 160:1240–1245.
1998.PubMed/NCBI
|
|
30
|
Reinis M, Símová J, Indrová M, Bieblová J,
Pribylová H, Moravcová S, Jandlová T and Bubeník J: Immunization
with MHC class I-negative but not -positive HPV16-associated tumour
cells inhibits growth of MHC class I-negative tumours. Int J Oncol.
30:1011–1017. 2007.PubMed/NCBI
|
|
31
|
Indrová M, Símová J, Bieblová J, Bubeník J
and Reinis M: NK1.1+ cells are important for the
development of protective immunity against MHC I-deficient,
HPV16-associated tumours. Oncol Rep. 25:281–288. 2011.
|
|
32
|
Bubenik J, Zeuthen J, Indrova M,
Bubenikova D and Simova J: Kinetics and function of
peritoneal-exudate cells during local IL-2 gene-therapy of cancer.
Int J Oncol. 4:13–16. 1994.PubMed/NCBI
|
|
33
|
Indrová M, Mikysková R, Jandlová T, Vonka
V, Bubeník J and Bieblová J: Adjuvant cytokine treatment of minimal
residual disease after surgical therapy in mice carrying
HPV16-associated tumours: Cytolytic activity of spleen cells from
tumour regressors. Folia Biol (Praha). 49:217–222. 2003.
|
|
34
|
Frey B, Janko C, Ebel N, Meister S,
Schlücker E, Meyer-Pittroff R, Fietkau R, Herrmann M and Gaipl US:
Cells under pressure - treatment of eukaryotic cells with high
hydrostatic pressure, from physiologic aspects to pressure induced
cell death. Curr Med Chem. 15:2329–2336. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Adkins I, Fucikova J, Garg AD, Agostinis P
and Špíšek R: Physical modalities inducing immunogenic tumor cell
death for cancer immunotherapy. Oncoimmunology. 3:e9684342014.
View Article : Google Scholar
|
|
36
|
Martini M, Testi MG, Pasetto M, Picchio
MC, Innamorati G, Mazzocco M, Ugel S, Cingarlini S, Bronte V,
Zanovello P, et al: IFN-gamma-mediated upmodulation of MHC class I
expression activates tumor-specific immune response in a mouse
model of prostate cancer. Vaccine. 28:3548–3557. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Okamoto M and Sato M: Toll-like receptor
signaling in anti-cancer immunity. J Med Invest. 50:9–24.
2003.PubMed/NCBI
|
|
38
|
Reinis M, Símová J, Indrová M, Bieblová J
and Bubeník J: CpG oligodeoxynucleotides are effective in therapy
of minimal residual tumour disease after chemotherapy or surgery in
a murine model of MHC class I-deficient, HPV16-associated tumours.
Int J Oncol. 30:1247–1251. 2007.PubMed/NCBI
|
|
39
|
Spranger S, Javorovic M, Bürdek M, Wilde
S, Mosetter B, Tippmer S, Bigalke I, Geiger C, Schendel DJ and
Frankenberger B: Generation of Th1-polarizing dendritic cells using
the TLR7/8 agonist CL075. J Immunol. 185:738–747. 2010. View Article : Google Scholar : PubMed/NCBI
|