Introduction
Ewing's sarcoma (ES) is the second most common bone
malignancy after osteosarcoma (1).
ES is characterized by a rapid proliferation of small round tumor
cells, which causes rapid and extensive progressive bone
destruction in children, adolescents and young adults (2). With advances in adjuvant therapy, the
survival rate of patients with a localized form of the disease has
reached ~70% (3). However,
patients with a metastatic, recurrent form of the disease have a
poor prognosis (4).
MicroRNAs (miRs) are evolutionarily conserved,
small, non-coding RNA molecules of ~22 nucleotides in length which
can play critical roles in cell proliferation, apoptosis, and
developmental timing by negatively regulating the stability or
translational efficiency of their target mRNAs (5). Global profiling and functional
studies have demonstrated that multiple miRNAs contribute to cancer
development and biology (6,7).
MicroRNA-138 is downregulated in different cancers, including
prostate, ovarian, and lung cancer (8,9). A
recent study has identified that miR-138 suppresses cancer cell
proliferation or motility by targeting BCR-ABL/GATA-1 in chronic
myeloid leukemia (10), cyclin D1
in nasopharyngeal carcinoma (11)
and SOX4 and HIF-1α in ovarian cancer (12). It has been reported that the
expression level of miR-138 in several types of cancers is reduced
compared to normal tissues, and that miR-138 can prevent the
cellular proliferation or motility via the down-regulation of
oncogenes. However, the role of miR-138 in the proliferation and
invasiveness of ES remains unclear.
Focal adhesion kinase (FAK) is an important receptor
at the proximal position in the regulation of growth factor- and
integrin-stimulated cell motility. Specifically, it plays a key
role in focal contact formation as well as turnover, and FAK may
also function in promoting cell invasion and may regulate cell
adhesion, migration, survival, proliferation, and differentiation
in a variety of cells (13,14).
Some previous studies have strongly implicated FAK in the
development and progression of breast and other cancers (15,16).
Although several miRNAs have been found to target
FAK, including miR-7 (17),
miR-488 (18), and miR-151
(19), the correlation of FAK
expression and miRNAs in ES cells is totally unknown.
In the present study, we analyzed the genome-wide
expression of both miRNAs and mRNAs in five human ES cell lines and
human mesenchymal stem cells (hMSCs). The results suggest that the
expression of miR-138 was repressed, whereas that of FAK was
increased, in all five ES cell lines. Based on the inverse
correlation between miR-138 and FAK expression, we hypothesized
that the effect of FAK in ES cells might be mediated, at least in
part, by miR-138. The purpose of our study was to assess whether
the expression of FAK is regulated by miR-138, and whether the
pathway plays a role in tumorigenesis in ES cells.
Materials and methods
Ethics statement
All animal experiments were performed under the
guidelines for animal experiments as stipulated by the Oita
University Graduate School of Medical Science. All animal
procedures carried out in this study were reviewed, approved, and
supervised by the Oita University Graduate School of Medical
Science, Japan. All animals were kept and the experiments were
performed in accordance with the European Community guidelines for
the use of experimental animals (86/609/EEC).
Cell lines
The human ES cell lines, SKES1, RDES, SKNMC and SCCH
were obtained from JCRB Cell Bank (Tokyo, Japan), and WE68 was
kindly provided by Dr Frans van Valen (Westfalische-Wilhelms
University, Münster, Germany). Human mesenchymal stem cells (hMSCs)
were purchased from Takara Biotechnology (Otsu, Japan). RDES and
SKNMC cells were cultured in Dulbecco's modified Eagle's medium
(DMEM) high glucose medium (Invitrogen, NY, USA) with 10% FBS and
1% penicillin and streptomycin. SKES1 cell were cultured in
RPMI-1640 (Invitrogen) supplemented with 10% FBS. SCCH cells were
grown in minimal essential medium (MEM) supplemented with 10% fetal
bovine serum (FBS; Invitrogen) and 0.1 mmol/l nonessential amino
acids (NEAA). hMSCs were cultured mesenchymal stem cell basal
medium, chemically defined (MSCBM-CD) with MSCGM-CD SingleQuats
(Takara Bio). The cells were maintained at 37ºC incubator supplied
with 5% CO2 and passaged every 2–3 days.
RNA isolation
mRNAs were prepared from the triplicate cell
cultures using RNeasy (Qiagen, Valencia, CA, USA) kit according to
the manufacturer's instructions. The RNA quality was ensured,
before labeling, using RNA 6000 Nano kit and Bioanalyzer 2100
(Agilent, Santa Clara, CA, USA). miRNAs were prepared from
triplicate cell cultures using the miRNeasy Mini kit (Qiagen)
according to the manufacturer's instructions.
Genome-wide miRNA expression
microarray
GeneChip miRNA 3.0 array (Affymetrix, Santa Clara,
CA) was used for miRNA expression profiling in five ES cell lines
and hMSCs. The data were extracted from the images,
quantile-normalized, summarized (median polish), and
log2-transformed with miRNA QC software (Affymetrix). GeneSpring GX
11.0 (Agilent) was used to analyze the array results. The gene list
was filtered with a fold-change cutoff of 2 resulting in the output
of a list of genes that have significant differential expression at
2-fold or more. Pathway analysis was performed using KEGG pathway
database (http://www.genome.jp/kegg/pathway.html).
Analysis of mRNA expression by cDNA
arrays
GeneChip Genome HG U133 Plus 2.0 Array (Affymetrix)
was used for mRNA expression profiling in 5 ES cell lines and hMSC.
Biotinylated cRNA was synthesized from total RNA using the 3' IVT
Express kit (Affymetrix) according to the manufacturer's protocols.
The entire experiment was performed twice. The microarray numerical
values were analyzed using the GeneSpring GX 11.0 software,
according to the RAM16 Algorithm: quantile normalization, filter by
flags (detected), filter by expression on the normalized data
(20.0–100.0th percentile). The gene list was filtered with a
fold-change cutoff of 2, resulting in output of list with genes
that have significant differential expression at 2-fold or
more.
Prediction of binding site and mature
miRNA transfection
Among the predicted target genes of miR-138 in the
TargetScan (http://www.targetscan.org/), DIANA (http://diana.cslab.ece.ntua.gr/microT/),
and PicTar (http://pictar.mdc-berlin.de/) databases, we found that
FAK was one of the top candidates. One day prior to the
transfection, cells were seeded onto 6-well plates
(5×104 cells/well) and incubated with the complete
medium without antibiotics (2 ml/well). The transfection of
miR-138-1-3p mimic (5′-GCUACUUCACAACACCAGG GCC-3′), miR-138-1-3p
mutant (5′-CAGAUGAGCACAACAC CAGGGCC-3′) and negative control (NC)
mRNAs (Invitrogen) was performed using Lipofectamine 2000 reagent
(Invitrogen) in antibiotics-free OptiMEM (Invitrogen) according to
the manufacturer's instructions. After 48-h incubation following
the transfection, the cells were harvested and processed for
further analysis.
RNA extraction, cDNA synthesis, and
quantitative real-time PCR
Total RNA was extracted from prepared treated
samples with the TRizol reagent (Invitrogen) and cDNA was
synthesized according to the manufacturer's protocol (Roche).
Quantitative real-time PCR (qRT-PCR) was performed using a Light
Cycler 480 Probe Master system (Roche), and PCR-specific
amplification was conducted in the LightCycler® Nano
(Roche). The relative expression of genes (FAK and β-actin) was
calculated with the 2−ΔΔCt method. The primers used are
listed here (qRT-PCR; FAK-forward 5′-CCCAGAAAGAAGGTGAACG-3′,
FAK-reverse 5′-GGTCGAGGGCATGGTGTA-3′; GAPDH-forward
5′-CCTCTATGCCAACACAGTGC-3′, GAPDH-reverse
5′-GTACTCCTGCTTGCTGATCC-3′.
Knockdown of FAK expression using
siRNA
siRNA oligonucleotides targeting FAK mRNA was
purchased from Ambion (Tokyo, Japan) and MISSION siRNA Universal
Negative Control was purchased from Sigma-Aldrich (Osaka, Japan).
The siRNAs were transfected into SKES1 and RDES cells using
Lipofectamine 2000 reagent according to the manufacturer's
instructions. The cell lines were harvested 48 h after the
transfection, then subjected to various analyses. The experiment
was repeated three times.
Cell proliferation assay
The cells were plated in 6-well plates
(5×104 cells per well), and were transfected with or
without hsa-miR-138-1-3p mimic, negative control miRNA, FAK-siRNA
or negative control-siRNA. Then the cells were incubated in
antibiotic-free OptiMEM. After 48 h of the cultivation, the cells
were counted using TC10 Automated Cell Counter (Bio-Rad).
Western blot analysis
Whole cell lysates were prepared from the cancer
cell lines and cellular protein (15 μg) was resolved on a precast
10% Tris-HCl Criterion 10-well gel (Bio-Rad) at 200 V (300 mAmp)
for 30 min. Antibodies rabbit anti-human FAK (#3285), RhoA (#2117),
paxillin (#2542), phosphorylation (p)-paxillin (#2541), Src
(#2108), p-Src (#5473), Rac (#4651), p-Rac (#2461), PARP (#9542),
cleaved PARP (#5625) were obtained from Cell Signaling Technology.
Rabbit anti-F-actin (ab130935), anti-p-RhoA (ab41435) and
anti-human β-actin (ab16039) Abcam Technology (Cambridge, UK).
Immunocomplexes were visualized with horseradish
peroxidase-conjugated anti-rabbit immunoglobulin G antibodies (GE
Healthcare, Tokyo, Japan), the blots were developed using ECL Plus
system (GE Healthcare) with a ChemiDoc camera (ImageQuant LAS 4000
mini; GE Healthcare). The quantification of western blot signals
was performed by the densitometry with ImageQuant TL software (GE
Healthcare). All primary antibodies were used at a 1:1,000
dilution. Peroxidase-conjugated anti-Rabbit IgG secondary
antibodies (GE Healthcare) were used at a 1:5,000 dilution. Three
independent experiments were performed for each analysis and the
gels have been run under the same experimental conditions.
Cell motility and migration assays
For the motility assays, stable monoclonal cell
lines (SKES1/miR-negative control, SKES1/miR-138 and
SKES1/siRNA-negative control, SKES1/FAK-siRNA) and 2×105
cells transfected from stable cell lines were resuspended in the
migration medium (medium without FBS), and were plated in the top
chamber without the Matrigel-coated membrane (24-well insert; 8 mm
pore size; BD Biosciences). The lower compartment was filled with
600 ml of a medium containing 30% FBS as a chemoattractant.
Following an incubation for 16 h at 37ºC and 5% CO2 in a
humidified incubator, the cells on the lower surface of the filter
were fixed in 4% formaldehyde for 20 min, then stained with Giemsa
stain for 10 min at room temperature and examined by light
microscopy.
Cell cycle analysis
SKES1 cells were seeded at a density of
5×105 cells/well in a 6-well plate and transfected with
miR-138 and FAK-siRNA. After transfection for 48 h, samples were
trypsinized, resuspended in PBS (Life Technologies), and fixed with
cold 70% ethanol overnight. For cell cycle analysis, cells were
stained with propidium iodide using Cycletest Plus DNA Reagent kit
(BD Biosciences) following the manufacturer's protocol, and the
cell cycle distribution was analyzed by FACS Verse flow cytometer
(BD Biosciences). The percentages of cells in G0/G1, S, and G2/M
phases were counted and compared. The experiments were carried out
in triplicate.
Apoptosis assay
The changes in the expression of apoptotic proteins
were analyzed by western blot analysis using antibodies against
PAR/poly(ADP-ribose) polymerase (PARP) (#9542) and cleaved PARP
(#9541) (Cell Signaling Technology, Tokyo, Japan) as an index of
apoptosis. The FITC Annexin V Apoptosis Detection kit (BD
Pharmingen, San Jose, CA, USA) was used to detect apoptotic cells
by flow cytometry. Annexin V binding buffer was used to resuspend
cells, and the cell suspensions were stained with FITC-Annexin V
and PI staining solution for 15 min at room temperature. The
samples were analyzed by FACSVerse using FACSuite analysis software
(BD Bioscience). As a positive control for apoptosis, SKES1 cells
treated with doxorubicin at 40 mg/ml for 20 h were used.
In vivo experimental and spontaneous
metastasis models
The experimental metastasis model was established by
injection of SKES1 cells (1×106) transfected with
miR-138 suspended in 100 μl of normal saline into the tail veins of
nude mice. Four groups were established: i) untreated (n=7); ii)
transfection of negative control-miRs (n=7); iii) transfection of
miR-138 (n=7) and iv) transfection of FAK-siRNA (n=7). All mice
were fed in standard condition with weight monitoring and
sacrificed 6 weeks after the cell inoculation. Tumor volumes were
measured using a micro-CT apparatus (R_mCT) which allows us to
obtain high-resolution CT images in small living animals. The tumor
volume of the lung nodule was estimated using the formula (π × long
axis × short axis × short axis)/6.
Statistical analysis
Statistical analysis was carried out and all
analyses were conducted using SPSS® 18.0 software (SPSS
Japan Inc., Tokyo, Japan). Two-tailed Student's t-test was used for
analysis of continuous variables. P<0.01 was considered to be
statistically significant. We determined the difference in >3
groups using a nonrepeated measures analysis of variance (ANOVA)
and Scheffe test. Results were expressed as the mean ± standard
deviation, and P<0.01 was considered as statistically
significant.
Results
Downregulation of miR-138 expression in
ES cell lines
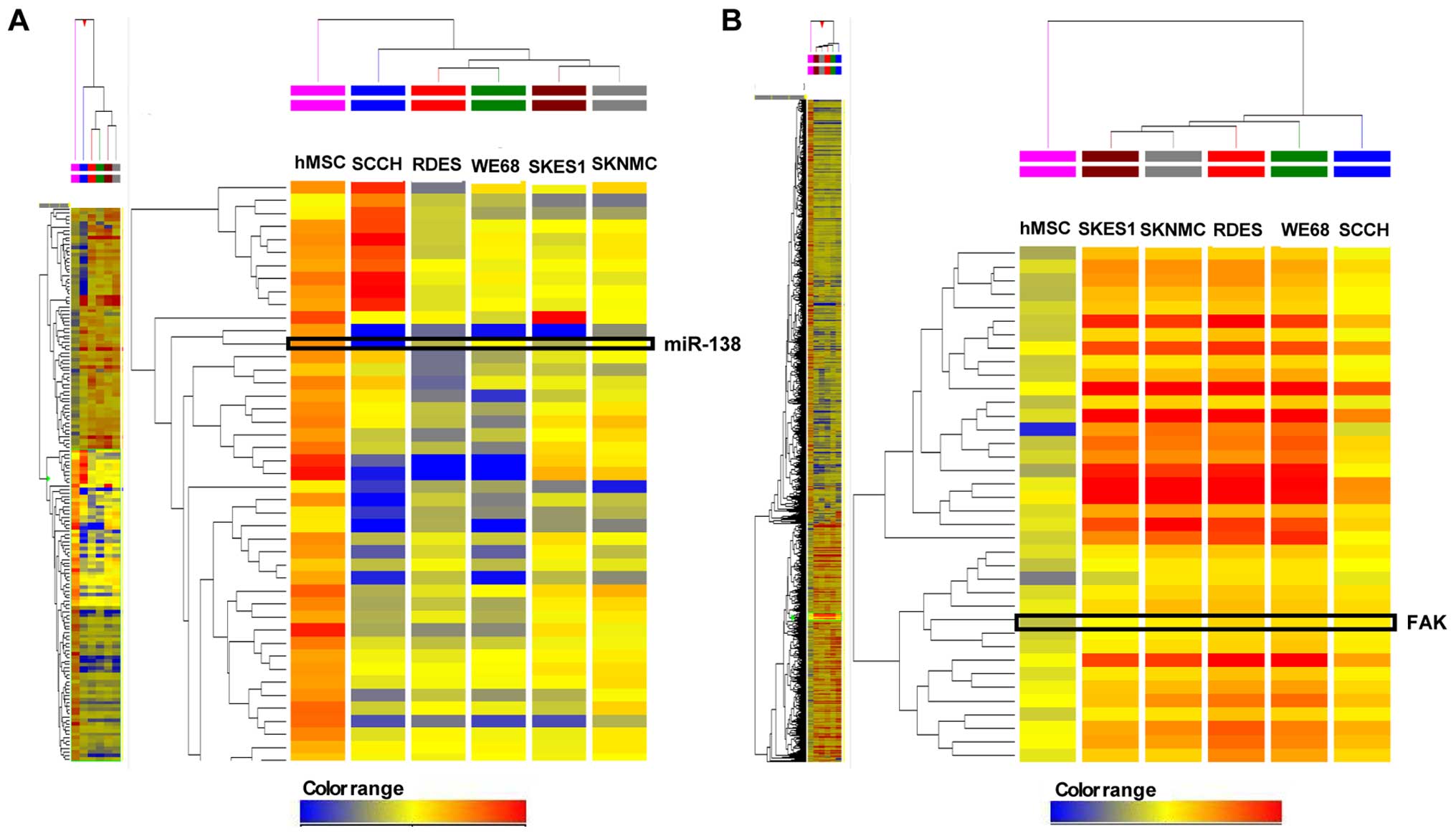
Genome-wide miRNA expression profiling was carried
out using five ES cell lines, to identify miRNAs expressed in ES
cells specifically. The array analysis showed that the expression
of 1,054 miRNAs in ES cells was changed (fold-change >2.0)
compared to the hMSCs (Fig. 1A).
Among these 1,054 miRNAs, 228 were significantly upregulated,
whereas 705 were significantly downregulated, in all tested ES
cells compared to the hMSCs. The remaining 121 miRNAs were up- or
downregulated among the five ES cell lines. In the ES cell lines, a
decrease in the expression of miR-138 by 4.03–179.82-fold was
observed compared to the hMSCs. The rank of fold-change of miR-138
were 66 out of 705 for SCCH, 97 out of 705 for WE68, 217 out of 705
for SKES1, 399 out of 705 for RDES and 531 out of 705 for
SKNMC.
Upregulation of FAK expression in ES cell
lines
The cDNA array analysis demonstrated that the
expression of 3,043 mRNAs was significantly changed between five ES
cell lines and hMSCs (Fig. 1B). We
found that 1,062 genes were significantly upregulated, whereas
1,884 genes were significantly downregulated, and the remaining 97
genes were up- or downregulated in five ES cell lines compared to
hMSCs. The expression of FAK was increased by 2.45–5.07-fold in the
five ES cell lines.
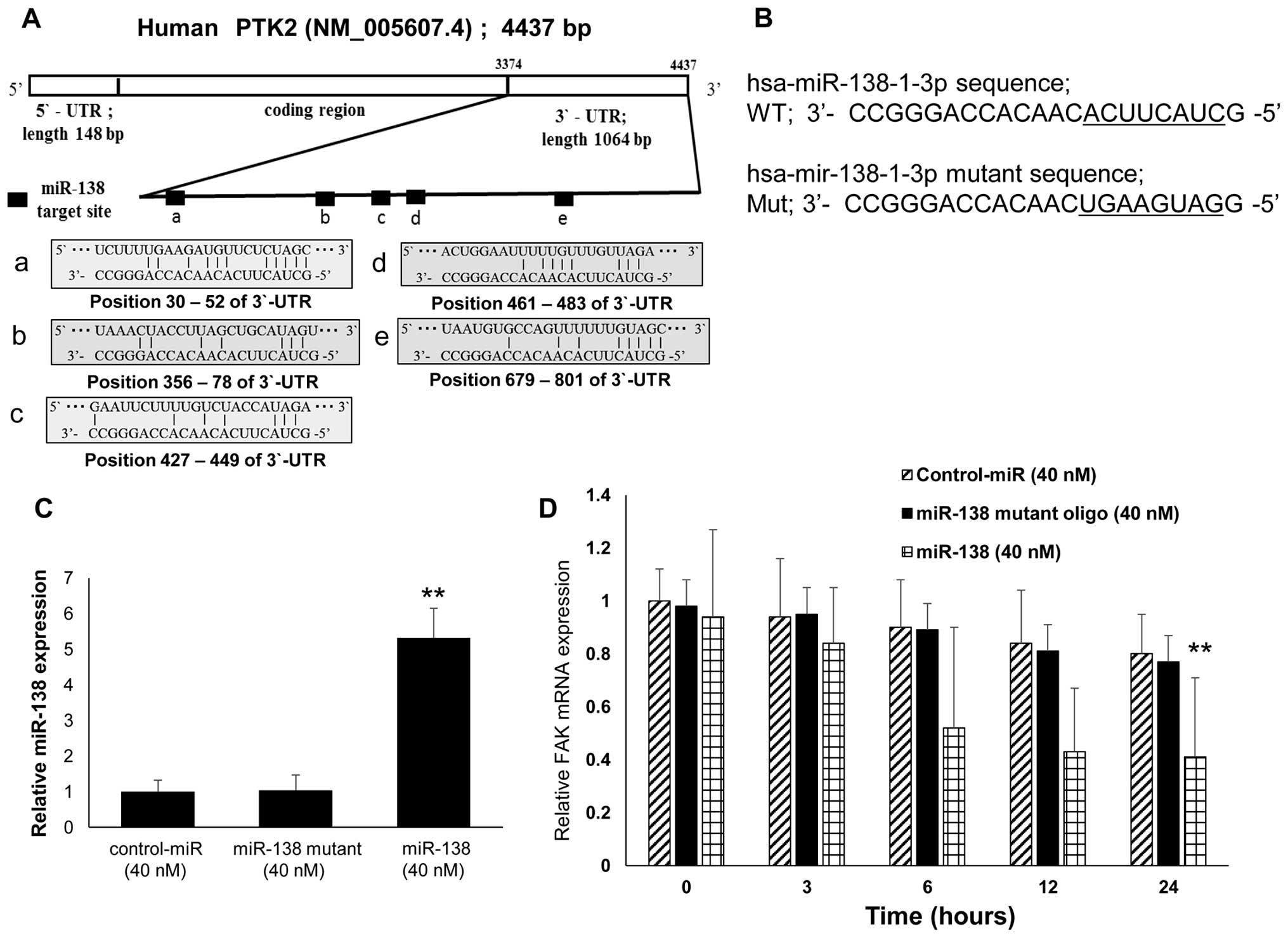
miR-138 directly targets FAK mRNA
The region complementary to the miR-138 seed region
was found in the 3′-UTR of human FAK (Fig. 2A). To test whether miR-138
expression affected endogenous FAK expression, we transfected the
miR-138-1-3p and miR-138-1-3p mutant oligonucleotides, (Fig. 2B) as well as the negative
control-miR, into SKES1 cells. We observed an increased miR-138
expression by 5.32-fold compared with control-miR (Fig. 2C) and decreased FAK expression at
the mRNA level following transfection with the miR-138-1-3p
oligonucleotide (Fig. 2D).
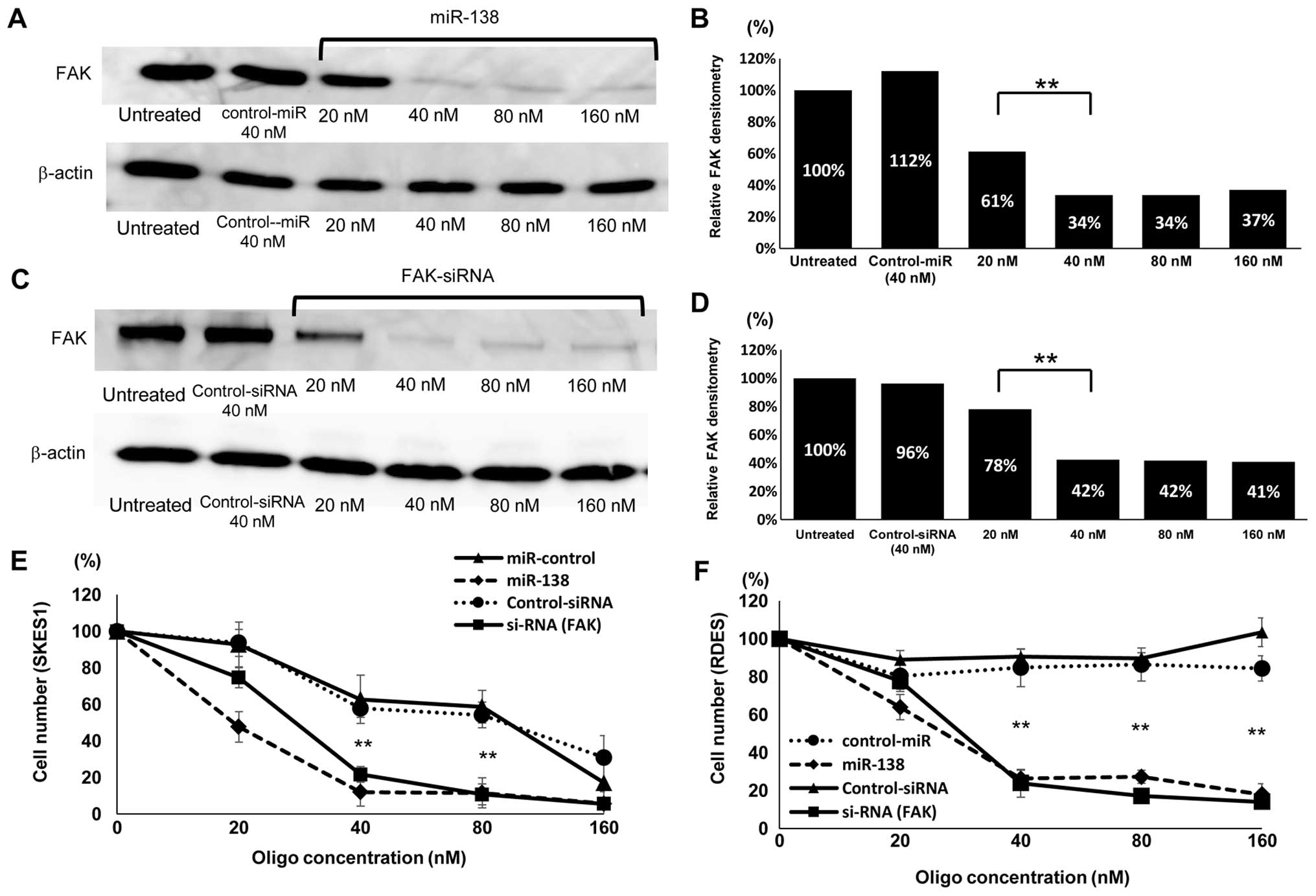
Inhibition of FAK expression by miR-138
and FAK siRNA
To examine the correlation between miR-138 and FAK
in ES cells, miR-138 was transfected into SKES1 cells. A western
blot analysis showed that the expression level of FAK strongly
decreased in miR-138 transfected cells compared to the untreated or
negative control oligo-transfected cells (Fig. 3A). The FAK protein expression level
in the miR-138 (40 nM) transfected cells was reduced to 34% of that
in the control cells (P<0.01) (Fig.
3B). To further confirm the effects of FAK on the growth of ES
cells, a transfection with siRNA-targeting FAK was carried out.
Although the FAK protein expression level in the cells transfected
with the negative control siRNA was not significantly affected,
that in the cells transfected with FAK siRNA was significantly
reduced, as determined by western blot analysis (Fig. 3C). Compared to the control cells
(100%), FAK siRNA (40 nM) transfected cells exhibited a
significantly lower FAK expression level by 42% (P<0.01)
(Fig. 3D).
Suppression of ES cell growth by
transfection of miR-138 and FAK siRNA
FAK is known to play important roles in the
regulation of cell proliferation. As the transfection of miR-138
resulted in the reduction of FAK expression, we then examined the
effects of miR-138 on the proliferation of ES cells. The SKES1 cell
growth was inhibited by the transfection of miR-138 and FAK-siRNA
compared to the negative control-miRNA and negative control-siRNA
transfected cells, respectively, 48 h after the transfection, as
determined by cell counts (Fig.
3E). RDES cells transfected with miR-138 and FAK siRNA
(Fig. 3F) showed a significant
inhibition of cell proliferation compared to the negative
control-miR and siRNA-transfected cells. At the high concentration
of 80 and 160 nM, the challenge of control miR and siRNA also
exhibited growth inhibition in SKES1, but not in RDES cells. The
results suggested that the susceptibility to toxicity of the
oligonucleotides might differ among cell lines.
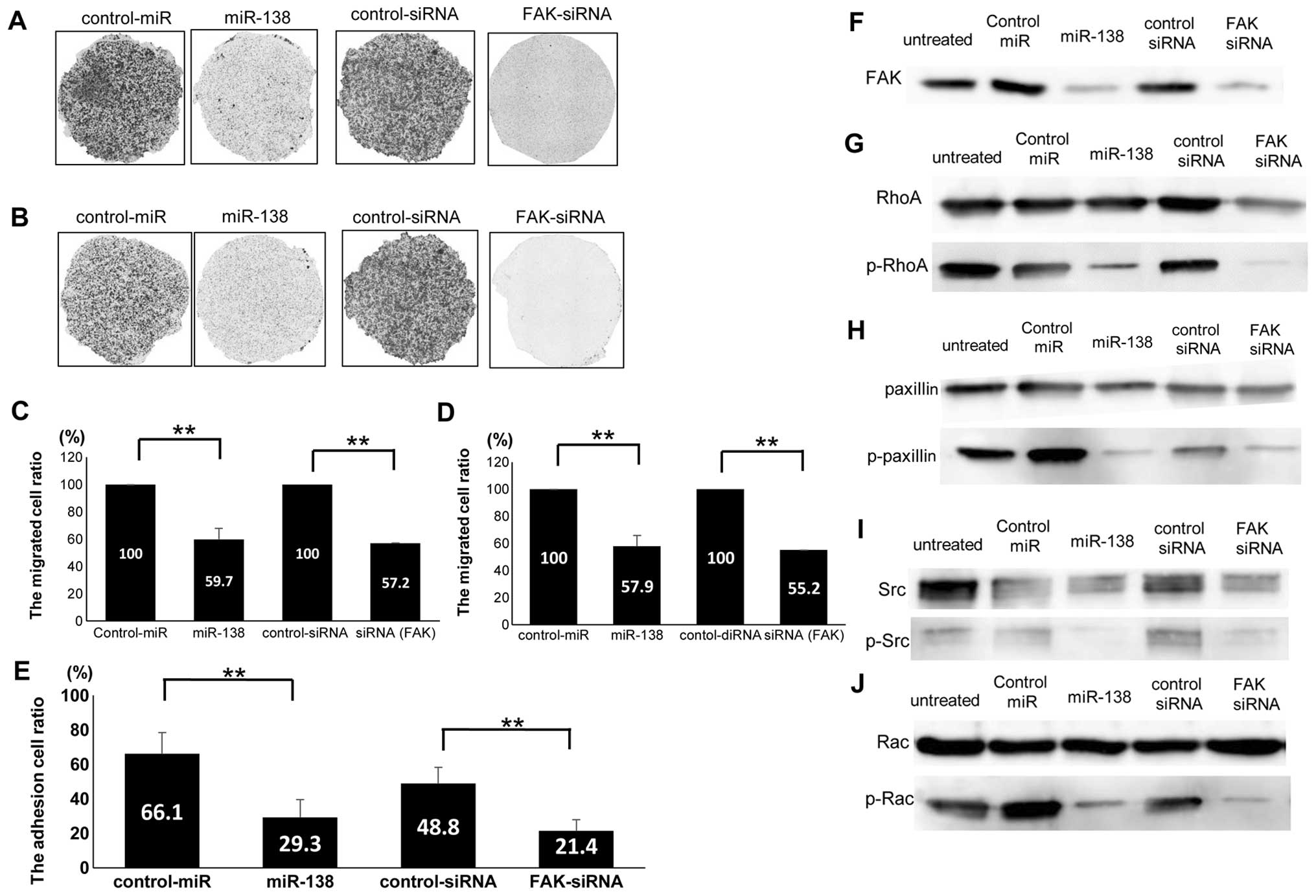
miR-138 attenuates the in vitro motility,
migration and adhesion of ES cells through the targeting of
FAK
We investigated the effects of miR-138
overexpression and target gene restoration on the motility and
migration of SKES1 cells. In a Transwell migration assay, SKES1
cells stably transfected with miR-138 and a FAK-siRNA precursor
presented impaired motility compared to those transfected with
negative control-miR and control-siRNA (Fig. 4A). Transwell cells that went
through the membrane without the Matrigel in miR-138 (59.7±8.2%)
and siRNAs (57.2±7.1%) were significantly fewer than those of the
negative control (Fig. 4C). In a
Matrigel invasion assay, miR-138 overexpression and FAK-siRNA
attenuated the invasiveness of SKES1 compared to the control-miRs
(Fig. 4B). Transwell cells that
went through the membrane with the Matrigel coated in miR-138
(57.9±7.9%) and siRNAs (55.2±6.4%) were significantly fewer than
those of the negative control (Fig.
4D). To evaluate the cell adhesion ability, we counted the
cells that did not float in vitro. Compared to the negative
control miR (66.1±12.2%) or the control siRNA group (48.8±9.4%),
the ratio of adhesion cells was lower in the miR138 (29.3±10.3%)
and the FAK-siRNA group (24.1±6.3%), respectively (Fig. 4E). These results suggest that
miR-138 and FAK-siRNA remarkably attenuated the in vitro
migration and invasion of ES cells through the targeting of FAK.
The expression level of FAK was markedly decreased in miR-138- (40
nM) and FAK-siRNA- (40 nM) transfected cells, than in the untreated
or negative control oligo-transfected cells (Fig. 4F). The protein expression level of
RhoA (Fig. 4G), paxillin (Fig. 4H), F-actin (data not shown), Src
(Fig. 4I) and Rac (Fig. 4J) did not significantly decrease in
miR-138- (40 nM) and FAK-siRNA- (40 nM) transfected cells, compared
to the untreated or negative control oligo-transfected cells. The
phosphorylation level of RhoA, paxillin, Src and Rac significantly
decreased in miR-138 (40 nM) and FAK-siRNA- (40 nM) transfected
cells, compared to the untreated or negative control
oligo-transfected cells.
Downregulation of FAK through miR-138-
and siRNA-induced anoikis
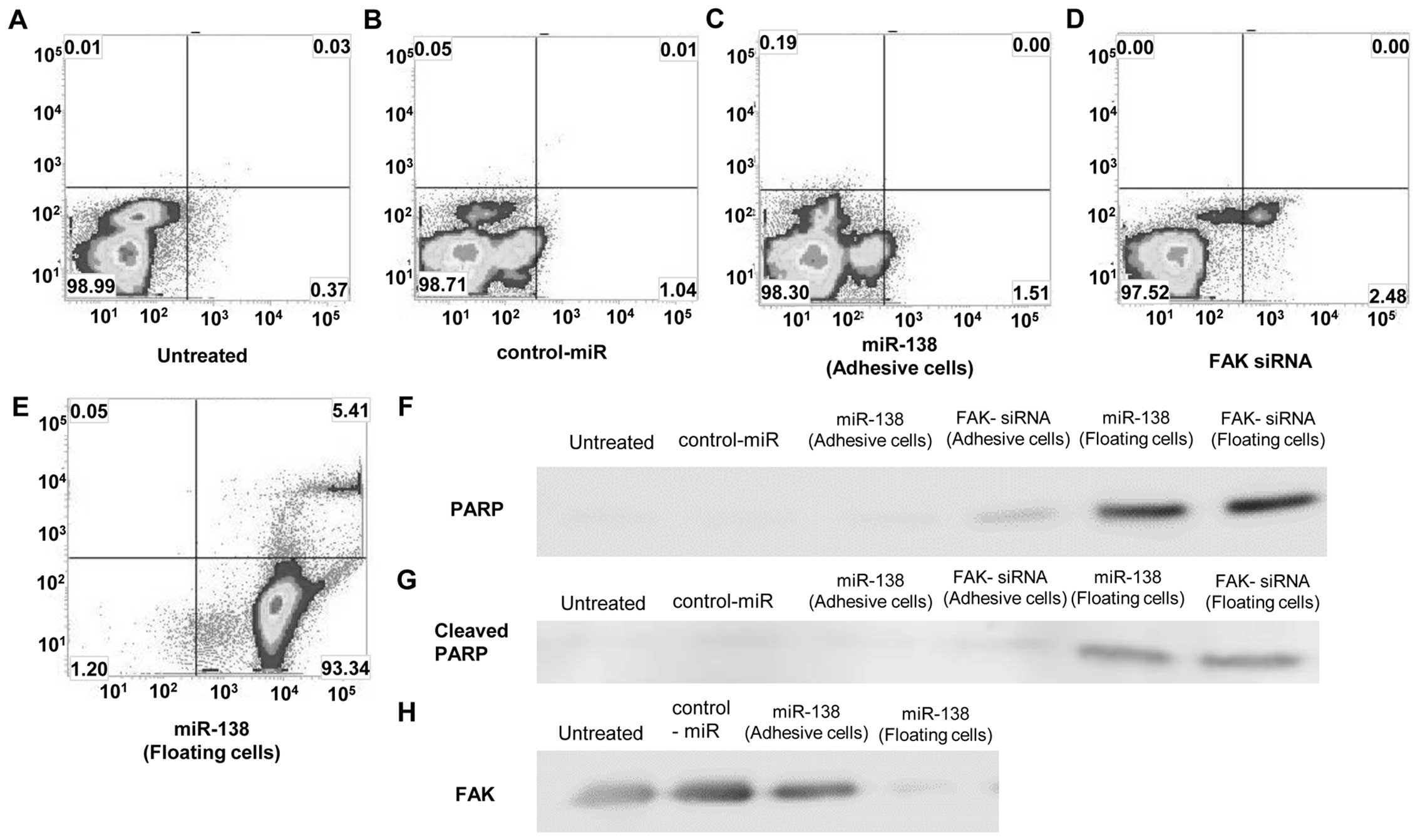
In a fluorescence activated cell sorting (FACS)
analysis of Annexin V-FITC/PI double staining, there was no
significant difference between untreated, 40 nM control-miR, 40 nM
miR-138 and 40 nM FAK-siRNA (Fig.
5A–D). We counted the number of floating cells following
miR-138 treatment, to check the status of these excluded cells.
Almost all of the cells that were floating following miR-138
treatment were sorted in the apoptosis fraction (Fig. 5E), rather than in the other
adhesive cell groups. The effect of programmed cell death or
apoptosis on SKES1 cells was not induced by miR-138 as anoikis
in vitro. The cellular expression of PARP (Fig. 5F) and that of its cleaved product
(Fig. 5G) was assayed by
immunoblotting in the SKES1 cell lines and their transfections. The
presence of the cleaved PARP product, a marker for caspase-mediated
apoptosis, was not expressed in the miR-138 or siRNA against the
FAK transfectants, nor in the untreated cells and negative control
transfectants, in marked contrast to the cells that were floating
following miR-138 and FAK-siRNA transfection (Fig. 5H).
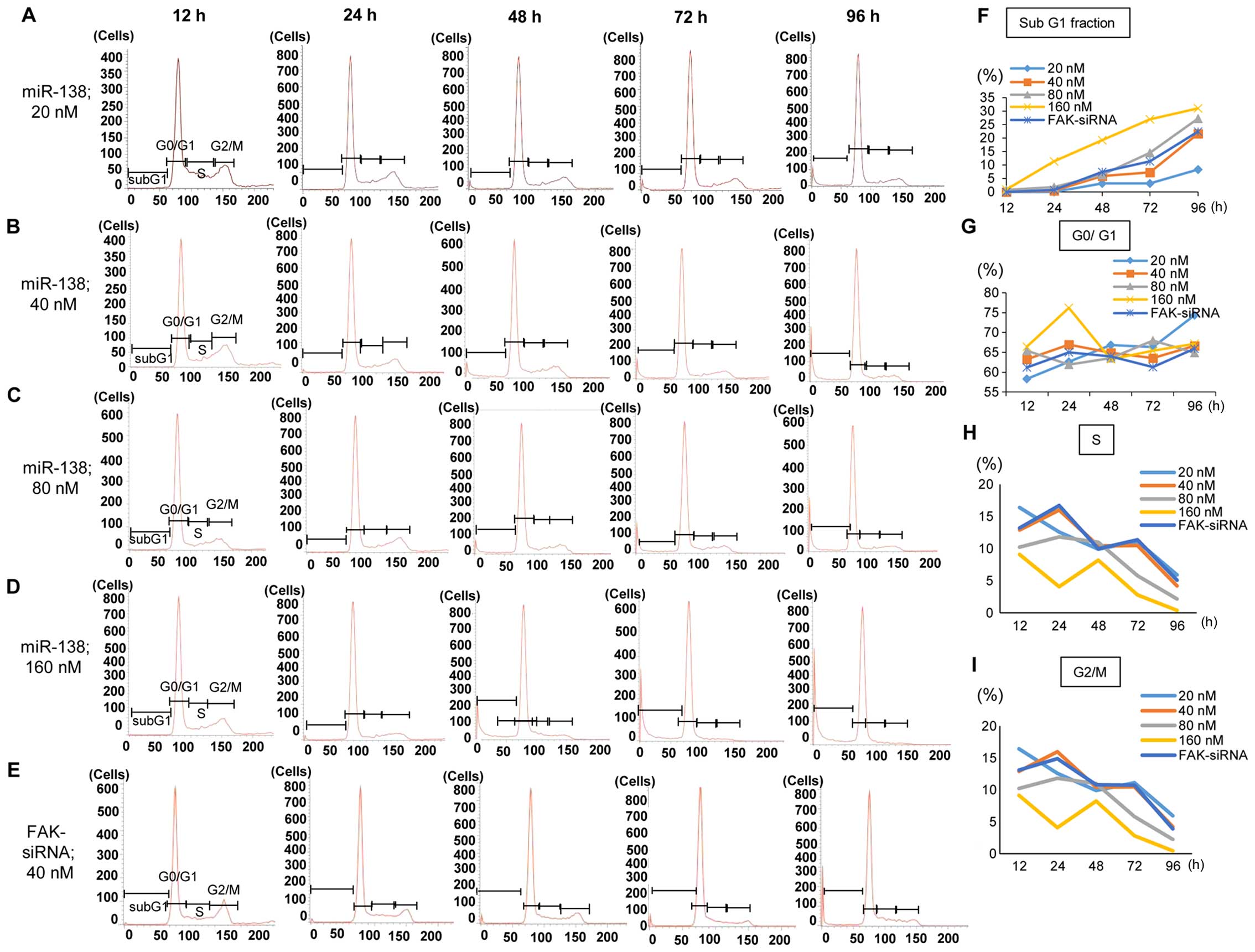
miR-138 effects of cell cycle and
apoptosis are dose- and time-dependent
To determine the cause of the repression of cell
proliferation following the transfection of SKES1 cells with
miR-138 at 20 nM (Fig. 6A), 40 nM
(Fig. 6B), 80 nM (C), 160 nM
(Fig. 6D), and with FAK-siRNA
(Fig. 6E), we carried out a FACS
analysis to measure the cell cycle distribution. In the 20-nM
condition, the S and G2/M phases were reduced, and the sub-G1
fraction was increased after 96 h. Furthermore, we investigated the
proportion of the sub-G1 fraction in PI in all groups, because the
miR-138 transfectants were increased in the cells that were
floating in the culture dish, and we only used adhesive cells in
the cell count analysis. miR-138 might be a repressor of cell
growth via the inhibition of cell adhesion, as the effect of
miR-138 in SKES1 cells is the induction of anoikis in a dose- and
time-dependent manner, resulting from the loss of the adhesive
function in floating cells, and the repression of cell cycle
progression in viable cells that were barely kept alive by
adhesion. The proportion of cells in the sub-G0/G1 fraction was
increased in a time- and miR-138 dose-dependent manner (Fig. 6G), whereas that of cells in the S
phase (Fig. 6H) and the G2/M
(Fig. 6I) phase were decreased,
also depending on the miR-138 dose and reaction time. The
proportion of cells in the G0/G1 fraction showed no significant
trend to be influenced by the ratio of cells in the sub G0/G1, S
and G2/M fractions (Fig. 6G).
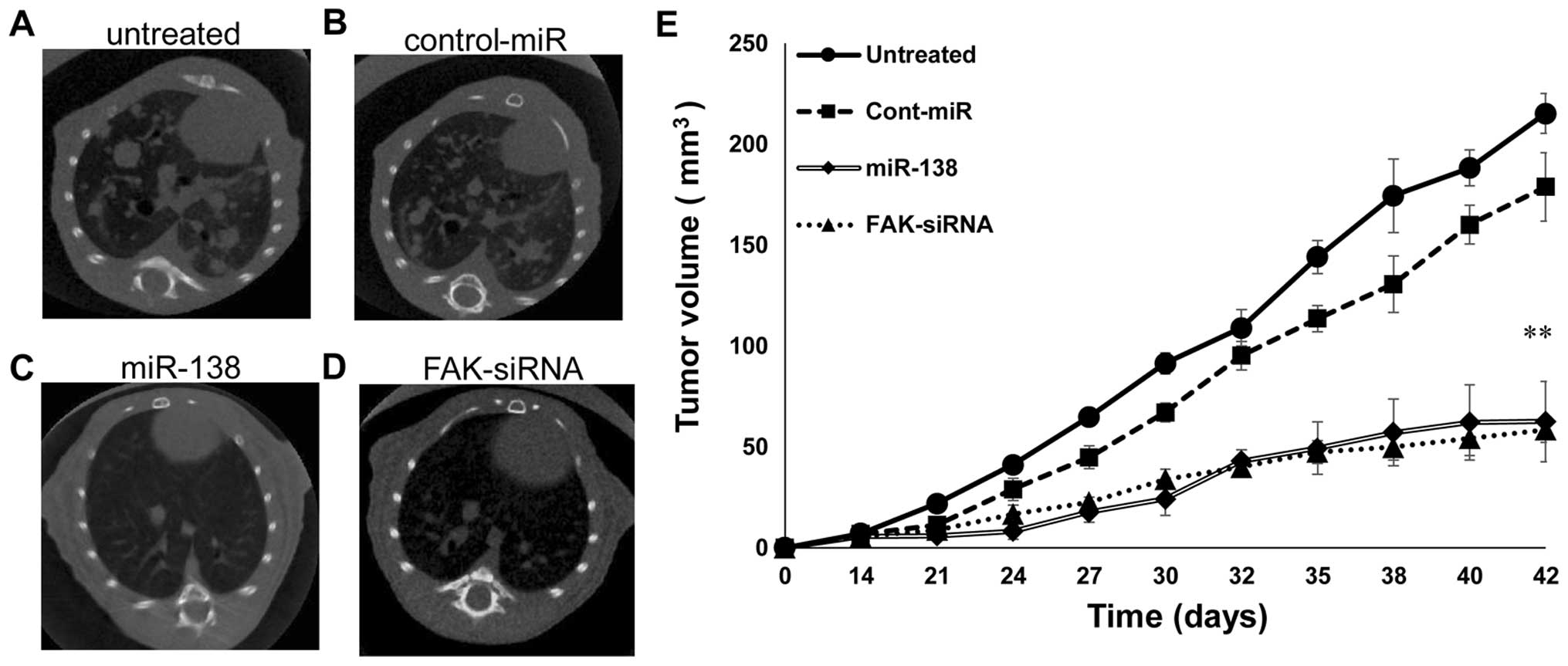
miR-138 inhibits tumor growth in a nude
mouse xenograft model
Next, we investigated the efficacy of miR-138
against tumor growth in vivo. Stable transfection of miR-138
(Fig. 7C) and of a FAK-siRNA
(Fig. 7D) precursor into SKES1
cells resulted in decreased growth of xenograft tumors from the
tail vein of nude mice, compared to those stably transfected with
untreated cells (Fig. 7A) or
control-miR (Fig. 7B). In the
experimental metastasis studies, mice treated with SKES1 cells
stably transfected with miR-138 (62.5±19.9 mm3) and
FAK-siRNA (58.6±6.23 mm3) displayed a statistically
lower (P<0.01) lung metastatic volume than the untreated
(215.3±9.79 mm3) and control-miR (178.9±17
mm3) group (Fig.
7E).
Discussion
miRNAs regulate gene expression
post-transcriptionally by binding to the 3′-UTR of their target
mRNAs. Binding leads to the degradation of the target mRNAs and to
reduced translation of the target proteins. miRNA activity also
affects the expression of genes that are downstream of the direct
targets, and can lead to changes in global protein expression
profiles. Therefore, miRNAs potentially play a critical role in the
progression of human cancer. Aberrant expression of miRNAs has been
shown to play important roles in the dysregulation of cell
functions, including differentiation, proliferation, apoptosis, and
metastasis (20,21).
In the present study, our miRNA array results show
that the expression of miR-138 was downregulated in all of the five
ES cell lines. Several studies have shown that miR-138 is
downregulated, and it is closely related to the abnormal potential
of tumors (11,21–23).
The significant suppression of miR-138 expression in all of our
five ES cell lines suggests that miR-138 might play a tumor
suppressor role and might be related to some malignant potential in
ES. However, neither the functional role nor the targets of miR-138
in ES have been previously defined. The results suggest that the
expression of miR-138 was coordinately upregulated in ES cell
lines, and prompted us to perform genome-wide mRNA profiling in
these lines, using a cDNA array to detect the possible targets of
miR-138 in ES cells.
The data from the cDNA array shows that FAK mRNA
expression was increased in all of the five ES cell lines. FAK
activity is regulated by cellular adhesion and downstream of growth
factors and G-protein-linked receptor activation (24). It localizes to focal contact sites
and receives input from both integrin and growth factor receptors.
FAK is overexpressed in many malignant tumors, including lung
cancer, and its expression level is linked to the poor prognosis of
the corresponding cancers (25).
Our data are consistent with these tumors in that upregulation of
FAK may contribute to malignancy.
Although miR-138 might influence the expression of
many genes, we focused on FAK as the target of miR-138 in ES cells.
It has been reported that miR-138 targets several genes, including
BCR-ABL/GATA-1, CCND1, SOX4 and HIF-1α (10–12).
Our cDNA array analysis demonstrated that FAK was the only
miR-138-target gene whose expression was uniformly upregulated in
all five ES cell lines, whereas the expression of other candidate
genes varied among the ES cells. Using several algorithms, the
analysis further suggested that FAK is the putative target of
miR-138. Thus, we analyzed the possibility that miR-138 might
contribute to anticancer activities by targeting FAK in ES
cells.
Due to the reported importance of FAK in cancer
(26), we further characterized
its role in relation to miR-138. We examined the functions of
miR-138 on the regulation of its predicted target genes and the
changes in the biological characteristics of our ES cell lines. The
forced elevation of miR-138 resulted in the reduction of the
expression of the FAK protein, suggesting that miR-138 might
function as a tumor suppressor gene in ES cells. Interestingly, FAK
protein levels are downregulated by the overexpression of miR-138
are known as tumor suppressor gene. FAK, a protein tyrosine kinase
involved in invasion and metastasis regulation, is a direct target
of miR-138 (27), and FAK has the
ability to promote cell cycle progression (14–16).
Aberrant expression of FAK will lead to abnormal cellular
proliferation. Our results suggest that the same mechanism of
regulation of FAK expression by miR-138 might exist in ES cell
lines.
FAK plays an important role in the regulation of
cellular invasion, migration and metastasis (28). Downregulation of FAK expression by
miR-138 inhibited cell attachment, invasion and migration compared
to transfection negative control-miR in SKES cells. Our motility
analyses showed that miR-138 inhibited the movement of ES cells by
repressing motility. These observations are consistent with
previous reports demonstrating that FAK is necessary for its
motility (29). We can assume that
upregulation of miR-138 regulates the movement of ES cells via
miR-138-mediated FAK.
The percentage of apoptosis was increased following
transfection of 40 nM of miR-138, the tumor growth of which was
repressed in vitro. We found that cells are divided into
adherent cells and floating cells in petri dishes that were
transfected with miR-138. To verify that the repression of FAK via
miR-138 and FAK-siRNA could induce apoptosis, we collected the
floating cells following transfection of miR/siRNA, and analyzed
them to compare with the adhered cells in the culture dishes. We
showed that knocking down through the upregulation of miR-138 and
the transfection of FAK-siRNA induced anoikis in floating cells.
However, apoptosis was not induced in the adhered cells that were
transfected with miR-138/FAK-siRNA, based on the data on Annexin V
and the expression of the cleaved PARP protein.
Duration of 48 h and a concentration of 40 nM
represent the optimal miR-138 transfection conditions for FAK
expression, in which the number of tumor cells is reduced, but it
is monistic. To examine the function of miR-138 and the biology of
FAK in SKES1 in more detail, we tried to change the reaction time
and concentration of miR-138. In miR-138-transfected SKES1 cells,
the cell cycle progression was suppressed in adhesive cells and
apoptosis was simultaneously induced in floating cells. miR-138
exerts its antitumorigenic action in two ways: one is the
repression of the adhesive function, which leads to anoikis, and
the other is the repression of the cell cycle in viable cells that
are barely holding on the ground. Furthermore, these two methods
depend on the duration time and the concentration.
The overexpression of miR-138 in SKES1 cells
resulted in the inhibition of metastatic lung tumor growth in mouse
xeno-graft models. This study shows an important tumor suppressor
role in metastasis through the transfection of miR-138 in SKES1
cells.
In conclusion, the present study suggests for the
first time a correlation between miR-138 and FAK in ES cells. Our
results provide evidence that the expression level of miR-138 in ES
cells was significantly reduced and inversely correlated with the
FAK expression level, and that miR-138 plays important roles in ES
cell proliferation, invasion, and migration by targeting FAK both
in vitro and in vivo. Our data suggest that FAK is
one of the crucial factors that enhance tumor proliferation,
invasion, and migration in ES, as well as in other malignant
tumors. Recent studies have shown that FAK plays critical roles in
host cells in the tumor microenvironment and in the tumor cells
that contribute to metastasis (30,31).
FAK plays pleiotropic roles in cancer, and this research has to
progress to the next step, i.e., until the phenomenon is also
confirmed in human tumor tissue samples. This study is the first
report showing that miR-138 inhibits the proto-oncogenic FAKs,
providing deeper insight into the biology of ES tumorigenesis. Our
results might therefore prove useful to clinical work in the
future.
Acknowledgements
The authors thank Dr Katsuhiro Hanada and Dr Takashi
Kobayashi, for helpful discussion for this study. This study was
supported in part by National Cancer Center Research and
Development Fund (26-A-4), the Grants-in-Aid for Scientific
Research (no. 24592250) and 15K10451 from Japan Society for the
Promotion of Science, and the Grants-in-Aid for Scientific Research
from Japan Agency for Medical Research and Development
(15Ack0106087h0002)..
References
|
1
|
Bleyer A, Barr R, Hayes-Lattin B, Thomas
D, Ellis C and Anderson B: Biology and The distinctive biology of
cancer in adolescents and young adults. Nat Rev Cancer. 8:288–298.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Taylor R, Knowles HJ and Athanasou NA:
Ewing sarcoma cells express RANKL and support osteoclastogenesis. J
Pathol. 225:195–202. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Paulussen M, Craft AW, Lewis I, Hackshaw
A, Douglas C, Dunst J, Schuck A, Winkelmann W, Köhler G, Poremba C,
et al; European Intergroup Cooperative Ewing's Sarcoma Study-92.
Results of the EICESS-92 Study: Two randomized trials of Ewing's
sarcoma treatment - cyclophosphamide compared with ifosfamide in
standard-risk patients and assessment of benefit of etoposide added
to standard treatment in high-risk patients. J Clin Oncol.
26:4385–4393. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ladenstein R, Pötschger U, Le Deley MC,
Whelan J, Paulussen M, Oberlin O, van den Berg H, Dirksen U, Hjorth
L, Michon J, et al: Primary disseminated multifocal Ewing sarcoma:
Results of the Euro-EWING 99 trial. J Clin Oncol. 28:3284–3291.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Meltzer PS: Cancer genomics: Small RNAs
with big impacts. Nature. 435:745–746. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yeh YM, Chuang CM, Chao KC and Wang LH:
MicroRNA-138 suppresses ovarian cancer cell invasion and metastasis
by targeting SOX4 and HIF-1α. Int J Cancer. 133:867–878. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang H, Zhang H, Zhao M, Lv Z, Zhang X,
Qin X, Wang H, Wang S, Su J, Lv X, et al: MiR-138 inhibits tumor
growth through repression of EZH2 in non-small cell lung cancer.
Cell Physiol Biochem. 31:56–65. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu C, Fu H, Gao L, Wang L, Wang W, Li J,
Li Y, Dou L, Gao X, Luo X, et al: BCR-ABL/GATA1/miR-138 mini
circuitry contributes to the leukemogenesis of chronic myeloid
leukemia. Oncogene. 33:44–54. 2014. View Article : Google Scholar
|
|
11
|
Liu X, Lv XB, Wang XP, Sang Y, Xu S, Hu K,
Wu M, Liang Y, Liu P, Tang J, et al: MiR-138 suppressed
nasopharyngeal carcinoma growth and tumorigenesis by targeting the
CCND1 oncogene. Cell Cycle. 11:2495–2506. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kohlhoff J and Barnett B: Parenting
self-efficacy: Links with maternal depression, infant behaviour and
adult attachment. Early Hum Dev. 89:249–256. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schlaepfer DD and Mitra SK: Multiple
connections link FAK to cell motility and invasion. Curr Opin Genet
Dev. 14:92–101. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Siesser PM and Hanks SK: The signaling and
biological implications of FAK overexpression in cancer. Clin
Cancer Res. 12:3233–3237. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
McLean GW, Carragher NO, Avizienyte E,
Evans J, Brunton VG and Frame MC: The role of focal-adhesion kinase
in cancer - a new therapeutic opportunity. Nat Rev Cancer.
5:505–515. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lahlou H, Sanguin-Gendreau V, Zuo D,
Cardiff RD, McLean GW, Frame MC and Muller WJ: Mammary
epithelial-specific disruption of the focal adhesion kinase blocks
mammary tumor progression. Proc Natl Acad Sci USA. 104:20302–20307.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kong X, Li G, Yuan Y, He Y, Wu X, Zhang W,
Wu Z, Chen T, Wu W, Lobie PE, et al: MicroRNA-7 inhibits
epithelial-to-mesenchymal transition and metastasis of breast
cancer cells via targeting FAK expression. PLoS One. 7:e415232012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song J, Kim D and Jin EJ: MicroRNA-488
suppresses cell migration through modulation of the focal adhesion
activity during chondrogenic differentiation of chick limb
mesenchymal cells. Cell Biol Int. 35:179–185. 2011. View Article : Google Scholar
|
|
19
|
Luedde T: MicroRNA-151 and its hosting
gene FAK (focal adhesion kinase) regulate tumor cell migration and
spreading of hepatocellular carcinoma. Hepatology. 52:1164–1166.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lujambio A and Esteller M: How epigenetics
can explain human metastasis: A new role for microRNAs. Cell Cycle.
8:377–382. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Medina PP and Slack FJ: microRNAs and
cancer: An overview. Cell Cycle. 7:2485–2492. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang W, Zhao LJ, Tan YX, Ren H and Qi ZT:
MiR-138 induces cell cycle arrest by targeting cyclin D3 in
hepatocellular carcinoma. Carcinogenesis. 33:1113–1120. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee YC, Tzeng WF, Chiou TJ and Chu ST:
MicroRNA-138 suppresses neutrophil gelatinase-associated lipocalin
expression and inhibits tumorigenicity. PLoS One. 7:e529792012.
View Article : Google Scholar
|
|
24
|
Schlaepfer DD, Hauck CR and Sieg DJ:
Signaling through focal adhesion kinase. Prog Biophys Mol Biol.
71:435–478. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Carelli S, Zadra G, Vaira V, Falleni M,
Bottiglieri L, Nosotti M, Di Giulio AM, Gorio A and Bosari S:
Up-regulation of focal adhesion kinase in non-small cell lung
cancer. Lung Cancer. 53:263–271. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Siejka A, Barabutis N and Schally AV: GHRH
antagonist inhibits focal adhesion kinase (FAK) and decreases
expression of vascular endothelial growth factor (VEGF) in human
lung cancer cells in vitro. Peptides. 37:63–68. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Golubovskaya VM, Sumbler B, Ho B, Yemma M
and Cance WG: MiR-138 and MiR-135 directly target focal adhesion
kinase, inhibit cell invasion, and increase sensitivity to
chemotherapy in cancer cells. Anticancer Agents Med Chem. 14:18–28.
2014. View Article : Google Scholar :
|
|
28
|
Nowicki TS, Zhao H, Darzynkiewicz Z,
Moscatello A, Shin E, Schantz S, Tiwari RK and Geliebter J:
Downregulation of uPAR inhibits migration, invasion, proliferation,
FAK/PI3K/Akt signaling and induces senescence in papillary thyroid
carcinoma cells. Cell Cycle. 10:100–107. 2011. View Article : Google Scholar :
|
|
29
|
Li LH, Zheng MH, Luo Q, Ye Q, Feng B, Lu
AG, Wang ML, Chen XH, Su LP and Liu BY: P21-activated protein
kinase 1 induces colorectal cancer metastasis involving ERK
activation and phosphorylation of FAK at Ser-910. Int J Oncol.
37:951–962. 2010.PubMed/NCBI
|
|
30
|
Jia YL, Shi L, Zhou JN, Fu CJ, Chen L,
Yuan HF, Wang YF, Yan XL, Xu YC, Zeng Q, et al: Epimorphin promotes
human hepatocellular carcinoma invasion and metastasis through
activation of focal adhesion kinase/extracellular signal-regulated
kinase/matrix metalloproteinase-9 axis. Hepatology. 54:1808–1818.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fan H, Zhao X, Sun S, Luo M and Guan JL:
Function of focal adhesion kinase scaffolding to mediate endophilin
A2 phos-phorylation promotes epithelial-mesenchymal transition and
mammary cancer stem cell activities in vivo. J Biol Chem.
288:3322–3333. 2013. View Article : Google Scholar :
|