Introduction
Prostate cancer is one of the most common cancer
types and the second leading cause of cancer-related death in males
in western countries (1). Its
incidence has also kept rising in China during the past few years.
Though surgical and radiological interventions are the most popular
regimens, the lack of long-term clinical treatment options requires
indentifying more effective therapeutic targets to prevent
metastasis and recurrence of prostate cancer. Despite numerous
reported studies, there remain many unaddressed difficulties
regarding to biological mechanisms of prostate cancer initiation
and progression. It is generally believed that prostate
carcinogenesis is a consequence of both genetic and epigenetic
modifications, converting normal prostate glandular epithelium to
pre-neoplastic lesions and finally invasive carcinoma. To develop
more specific diagnostic and prognostic biomarkers, which demands a
better understanding of the molecular pathogenesis of prostate
cancer, is necessary.
Nuclear factor κ-light-chain-enhancer of activated B
cells (NF-κB) family of transcription factors encompasses five
members including RelA, RelB, c-Rel, NF-κB1 (p50 and its precursor
p105), and NF-κB2 (p52 and its precursor p100) in mammalian cells.
NF-κB functions critically in a range of biological processes such
as inflammation, immune response, carcinogenesis, and secondary
lymphoid organogenesis. In most cells, NF-κB subunits are
sequestered in the cytoplasm through binding to an inhibitory IκB
family member and kept inactive. Upon stimulation, the IκB protein
could undergo proteosomal degradation followed by the translocation
of NF-κB dimers into the nucleus to transactivate their target
genes. NF-κB dimers transduce the signaling through two major
pathways: the canonical and the non-canonical. The canonical NF-κB
activity involves the proteasome-mediated IκBα degradation and the
subsequent activation of RelA-p50 heterodimers. In the
non-canonical pathway, NF-κB2/p100 functions as an IκB-like protein
and retains RelB-p100 heterodimers in the cytoplasm. The
stimulatory signaling triggers NF-κB2/p100 phosphorylation and
proteasome-mediated degradation in an NF-κB-inducing kinase (NIK)
and IκB kinase (IKK)-dependent manner, thereby producing active
RelB-p52 heterodimers. The canonical NF-κB signaling pathway plays
a role in immune responses, and inflammation whereas the
non-canonical pathway is rather implicated in the secondary
lymphoid organogenesis and the generation of B lymphocytes
(2–4).
Dysregulated NF-κB activity leads to aberrant cell
proliferation and survival, angiogenesis, metastasis, and other
malignant phenotypes, which contributes to the development of many
human malignancies including prostate cancer (5). Previous studies have addressed
intensively the biological significance of the canonical NF-κB
activity in carcinogenesis of prostate cancer (6). Increasing attention has been paid to
the understanding of the role of the non-canonical NF-κB activity
in the pathogenesis of diverse hematological malignancies and solid
tumors; nevertheless, the significance of the non-canonical NF-κB
activity in regulating the carcinogenesis of prostate cancer is
still largely unclear.
RelB complexes, representing the non-canonical NF-κB
activity, is the most frequently detected NF-κB subunit in the
nucleus of prostate cancer tissue and is correlated directly with
Gleason score, suggesting that RelB might be involved in prostate
cancer initiation or progression (7). Both the canonical and the
non-canonical NF-κB activities are present in the
androgen-independent human prostate cancer cell lines such as PC-3
and DU145, while the non-canonical NF-κB activity is minimally
expressed in the androgen-dependent prostate cancer cell line such
as LnCaP (8,9). Therefore, it is suggested that RelB
is likely implicated in the transition from androgen-dependent into
androgen-independent status. RelB-targeting decreases cancer
incidence and growth rate, and sensitizes prostate cancer cells to
ionizing radiation. RelB exerts a radio-protective role in
aggressive prostate cancer cells, partially due to the induction of
the manganese superoxide dismutase (MnSOD) gene. Targeting
RelB is suggested to be a valuable strategy in overcoming the
radiation-resistance in both prostate and breast cancer (10–12).
Recent studies have suggested that IKKα, which is a crucial
upstream molecule in the NF-κB signaling pathway, is involved in
both canonical and non-canonical NF-κB activation, promotes
malignant phenotypes of prostate cancer. IKKα is thereby considered
to be an important therapeutic target in treating prostate cancer
(13,14).
Here, we examined systemically the functional roles
of RelB in prostate cancer cells and studied its significance as a
therapeutic target. Targeting RelB using short hairpin RNA (shRNA)
approach in DU145 prostate cancer cells had an effect on varied
biological behaviors of cells. Knockdown of RelB slowed down cell
growth due to increased apoptosis. The downregulated expression of
anti-apoptotic gene Bcl-2 played critical roles in regulating
spontaneous and radiation-induced apoptosis in the presence of RelB
knockdown. Importantly, RelB knockdown significantly diminished the
migration and invasion abilities of DU145 prostate cancer cells,
attributed at least partially to the clear reduction of integrin
β-1. RelB knockdown also enhanced the sensitivity to proteasome
inhibitor treatment. Collectively, we provided evidence that RelB
functioned as an oncogene in prostate cancer and shows potential
for targeted therapy in human prostate cancer.
Materials and methods
Tissue culture and reagents
The human prostate cancer cell line DU145 was
cultured in RPMI-1640 media supplemented with 10% fetal bovine
serum (FBS), 2 mM L-glutamine, 100 U/ml penicillin, and 100 μg/ml
streptomycin at 37°C in a humidified atmosphere containing 5%
CO2. Neomycin (cat no. E859-5G) was purchased from
Amresco. MG-132 (cat no. 474790) was purchased from Millipore.
Cell transfection
An shRNA carrying sequence targeting the RelB
gene (275–293: 5′-GCACAGATGAATTGGAGAT-3′) was designed and
chemically synthesized by Invitrogen (Beijing, China). The
shRNA-RelB was subcloned into the pSilencer3.1-H1-neo plasmid (cat
no. 5770, Thermo Scientific™, China), which was linearized by
restriction endonucleases HindIII and BamHI. The
recombinant plasmid pSilencer3.1-siRelB (carrying an shRNA
targeting RelB) and the scrambled control plasmid were then
transduced into cells using Lipofectamine 2000 (cat no. 12566014,
Thermo Scientific™, China) according to the manufacturer's
instructions. To obtain stably transfected cell lines, cells were
selected in the presence of neomycin for two weeks.
Quantitative real-time PCR (qRT-PCR)
Total RNA was extracted using TRIzol reagent
(Tiangen Biotech Co., Ltd., Beijing, China) according to the
manufacturer's instructions. RNA yield and purity were determined
spectrophotometrically at 260–280 nm and the integrity of RNA was
verified by Nanodrop-1000 (Thermo Scientific). Total RNA (2 μg) was
reverse-transcribed with Superscript M-MLV (Promega, China)
according to the manufacturer's instructions. Triplicates were
performed for all reactions with a LightCycler 480 System (Roche,
China). Primers for qRT-PCR were designed using Primer-BLAST
(PubMed) and synthesized from Invitrogen. A housekeeping gene,
β-actin, was always run together with target genes. Data were
analyzed with Pfafflmethod that provides a means for quantification
of a target gene transcript in comparison to a reference gene
(15).
Western blot analysis
RIPA buffer supplemented with proteinase inhibitor
cocktail was used to prepare cell lysates. Cell lysates including
whole-cell extracts and cytoplasmic/nuclear extracts were denatured
by boiling in SDS-PAGE sample buffer, fractionated on SDS-PAGE
gels, and transferred onto nitrocellulose membranes. The membranes
were blocked in 5% skim milk for 1 h at room temperature and then
incubated with the indicated primary antibodies (Abs) overnight
followed by appropriate secondary Abs. Both primary and secondary
Abs were diluted well in advance according to the manufacturer's
instructions. Proteins were detected and scanned with an
Odyssey® infrared imaging system (LI-COR Biosciences,
USA). Band density was normalized to α-tubulin, β-actin or lamin
reference. Abs against Bcl-2 (C-2, sc-7382), NF-κB p65 (C-20,
sc-372), RelB (C-19, sc-226), c-Rel (N, sc-70), NF-B pκ50 (H-119,
sc-7178), NF-κB p52 (K-27, sc-298), and Lamin A/C (H-110, sc-20681)
were purchased from Santa Cruz Biotechnology, Inc. Abs against
Bcl-xL (#2764), Bim (#2819), Mcl-1 ((#4572), Survivin (#2808),
MMP-2 (#4022), MMP-9 (#2270), and integrin β-1 (#9699S) were
obtained from Cell Signaling Technology. β-actin (AT0001) was
purchased from CMCTAG, Inc. α-tubulin (AJ1034a) was purchased from
Abgent. IRDye 680CW (926-32222) and IRDye 800CW secondary Abs
(926-32210) were obtained from LI-COR Biosciences.
Electro-mobility shift assay (EMSA)
Nuclear extract preparation and EMSA were performed
as previously described (16). The
integrity of nuclear extracts was checked using an Oct-1
oligodeoxynucleotide probe. Briefly, nuclear extracts were
incubated with an IRDye 800-labeled oligonucleotide probe. The
binding reactions occurred under specific salt/pH conditions in a
binding buffer. Poly (dI:dC) (Sigma-Aldrich, China) was added to
prevent non-specific binding of proteins to the κB
oligonucleotide probe. After binding, the samples were separated on
5% non-denaturing PAGE gels and bands were detected by scanning
using Odyssey infrared imaging system.
NF-κB activity assay
The activity of individual NF-κB members was
quantified using an ELISA-based NF-κB family transcription factor
assay kit (Active Motif, Carlsbad, CA, USA). In brief, 2 μg of
nuclear extracts were incubated in a 96-well plate with immobilized
NF-κB consensus oligo-nucleotides. Captured complexes were
incubated with NF-κB subunit-specific primary Abs and subsequently
detected with an HRP-conjugated secondary Ab, which provided a
sensitive colorimetric readout at 450 nm that was easily quantified
by spectrophotometry. All samples were tested in triplicates.
Cell growth assay
The xCelligence RTCA instrument (Roche) was applied
to monitor cell growth. In the assay, impedance for indicated times
was continuously monitored by the system, and the value was
indicated as ‘cell index’, which was determined by the number of
cells seeded, the overall size and morphology of the cells, and the
degree to which the cells interact with the sensor surface.
Following running the background blank with 100 μl RPMI-1640 media
supplemented with 10% FBS, cells were seeded in wells and then the
program was run. The cell index was continuously monitored by the
system, and data were analyzed by RTCA software 1.2.
Cell migration and invasion assay
The cell migration and invasion assay was also
performed with the xCelligence RTCA instrument. In this assay, a
CIM-plate assembled with an upper and lower chamber was used.
RPMI-1640 (180 μl) media supplemented with 10% FBS was added to
each well on the lower chamber. Cells were suspended in the
serum-free media and added into the upper chamber. For the cell
invasion assay, wells of the upper chamber were pre-coated with
Matrigel (cat. no. 356234, BD Biosciences, China) for ≥4 h.
Following attachment, cell migration or invasion through Matrigel
towards the lower chamber containing RPMI-1640 media supplemented
with 10% FBS was continuously monitored, and data was collected and
analyzed by RTCA software 1.2.
Cell cycle analysis
The cell cycle analysis and cellular DNA content
measurement were carried out by flow cytometry. Cells were
harvested and fixed with 70% ethanol for 24 h at 4°C. Subsequently,
the single cell suspensions were prepared to stain DNA using
propidium iodide (PI) according to the manufacturer's instructions.
Cell cycle was measured by FACSCalibur™ cytometer (BD Biosciences)
with at least three independent experiments performed.
CFSE assay
Cells (1×106) were suspended in 1 ml
phosphate-buffered saline (PBS) and then stained with 2 μl CFSE
(cat no. C-1311, Molecular Probes). Afterwards, 5 ml pre-cooled
serum-free RPMI-1640 media was added to stop the staining. After
washed by RPMI-1640 media three times, cells were seeded into
dishes and the fluorescence intensity was detected using
FACSCalibur™ cytometer.
Scratch healing assay
The confluent cell layers were scratched using a 200
μl sterile pipette tip of and washed three times with PBS. The
scratched area was then imaged continuously at magnifications of
x10 with the light System Microscope IX71 (Olympus, Japan). The
migratory distance is used to measure the migratory ability of
cells.
Gelatinase zymography
Gelatin zymography assay was performed in an 8%
SDS-PAGE gel in the presence of 0.1% gelatin under non-reducing
conditions. Culture media with sample buffer were loaded for
SDS-PAGE with Tris-glycine SDS buffer. Samples were not boiled
before electrophoresis. Following electrophoresis, the gels were
washed twice in 2.5% Triton X-100 for 30 min at room temperature to
remove SDS. The gels were then incubated at 37°C overnight in
substrate buffer containing 50 mM Tris-HCl and 10 mM
CaCl2 at pH 8.0, and stained with 0.5% Coomassie Blue
R250 in 50% methanol and 10% glacial acetic acid for 30 min, and
then destained. Upon renaturation of the enzyme, the gelatinases
digested the gelatin in the gel to produce clear bands against an
intensely stained background.
Apoptosis assay
Cells were harvested and stained with Annexin V and
PI alone or in combination for 15 min at room temperature according
to the manufacturer's instructions (Invitrogen, China). Cells were
subjected to flow cytometric analysis with a FACSCalibur™ cytometer
by using CellQuestPro software.
Irradiation treatment
Cells were cultured overnight in 100-mm2
plates at a density of 2×106/dish. Irradiation was
performed using Siemens Primus-M linear accelerator at room
temperature. Cells received a dose of 4 or 8 Gy per treatment at an
average dose rate of 2 Gy/min. The distance between the radiation
source and cells was 100 cm. Following the radiation treatment,
cells were incubated in RPMI-1640 media with 10% FBS for 24, 48 and
72 h respectively, and harvested for the following experiments.
Statistical analysis
Data were expressed as mean ± SD of at least three
separate experiments. All statistical analysis was performed by
Graphpad software. Differences between groups were valued by
Student's t-test. A p-value of ≤0.05 was considered significant, a
p-value of ≤0.01 was considered as highly significant, and a
p-value of ≤0.001 was considered as very highly significant.
Results
Introduction of shRNA-RelB into DU145
prostate cancer cells
To verify the functional significance of RelB in
prostate carcinogenesis, a plasmid containing either shRNA-RelB or
shRNA-control was constructed and then transfected into
androgen-independent DU145 prostate cancer cells, respectively.
Cells were cultured in the presence of neomycin (800 ng/μl) until
single colonies appeared. The selected monoclones were further
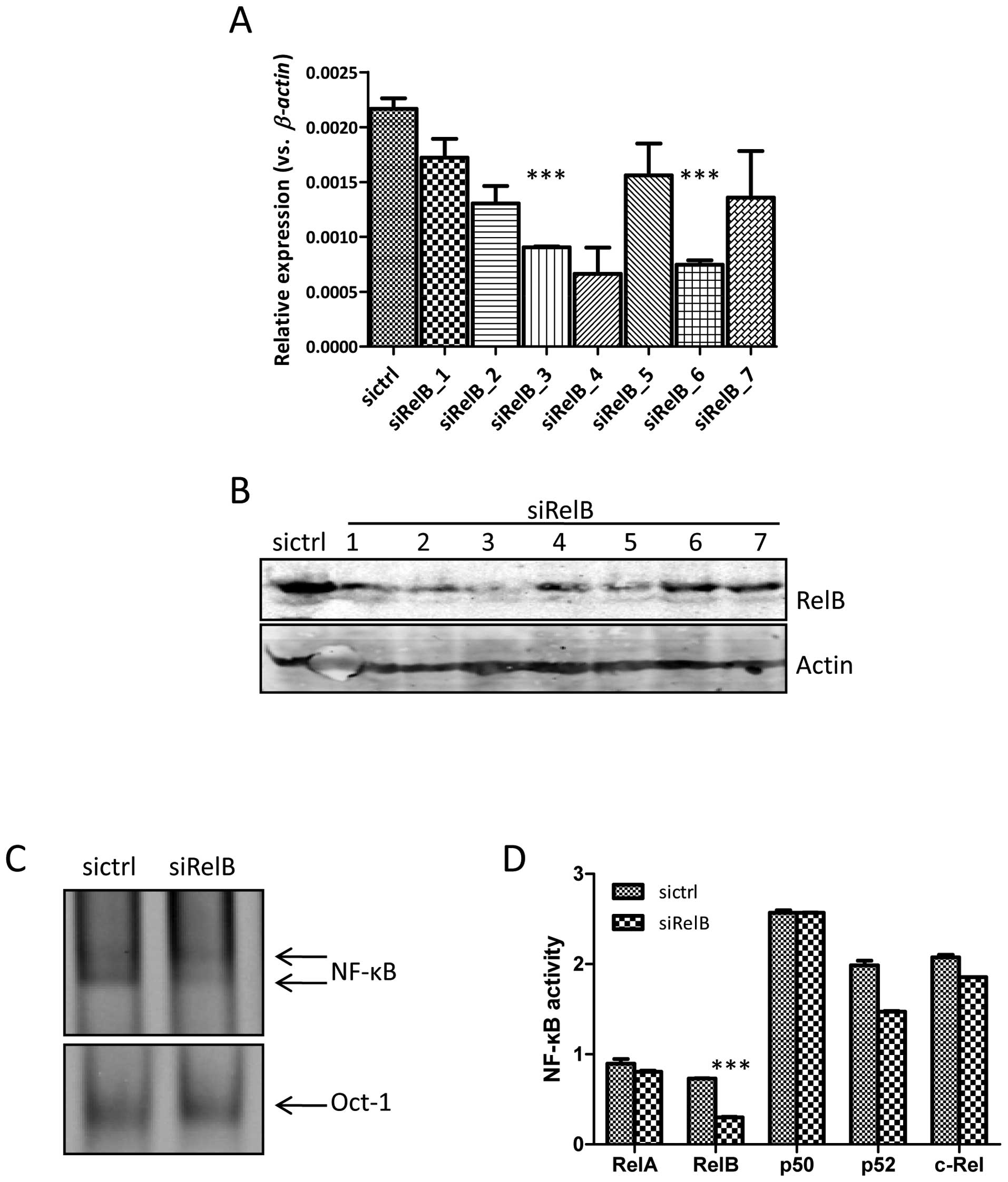
expanded and examined for the RelB expression. As shown in Fig. 1A, the RelB expression at
mRNA level detected by qRT-PCR was generally lower in the
DU145-siRelB group (transfected with the plasmid containing
shRNA-RelB) as compared with that in the DU145-sictrl group
(transfected with the plasmid containing shRNA-control), albeit at
different levels. The RelB expression of clone nos. 3 and 6
in the DU145-siRelB group was significantly downregulated compared
to that of the DU145-sictrl group. In line with the mRNA level, the
RelB expression of clone no. 3 in the DU145-siRelB group at protein
level was also evidently reduced by western blot analysis (Fig. 1B), indicating a successful RNA
interference (RNAi) of the RelB gene. Therefore,
DU145-siRelB clone no. 3 was chosen to carry out the following
study. The κB-DNA-binding activity in nuclear extracts of
DU145-siRelB and DU145-sictrl cells was measured by EMSA. Fig. 1C shows that the upper complex was
similar between the two cell lines, however, the lower complex was
attenuated in the DU145-siRelB cells as compared with that in the
DU145-sictrl cells. To dissect the exact contribution of RelA and
RelB to the κB-DNA binding complexes, an ELISA-based NF-κB activity
assay was performed. The average RelB activity in nuclear extracts
of DU145-siRelB was significantly decreased as compared with that
in the DU145-sictrl cells, while the average RelA, p50, p52, and
c-Rel activity was comparable (Fig.
1D).
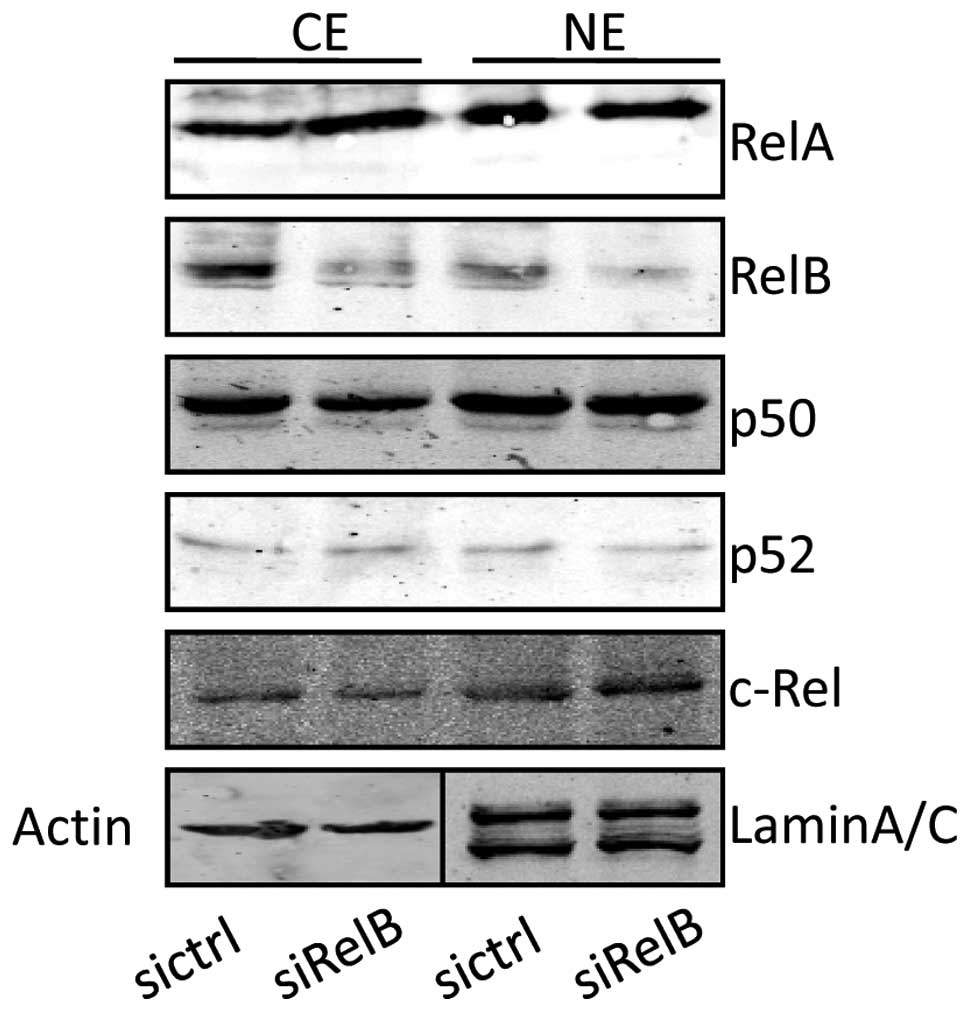
Knockdown of RelB does not affect other
NF-κB subunits
To investigate whether the RelB knockdown affected
the expression of other NF-κB subunits, western blot analysis was
carried out. As shown in Fig. 2,
the expression levels of RelA and p50, contributing to the
canonical NF-κB activity, were comparable between the DU145-siRelB
and DU145-sictrl cells at both cytoplasmic (CE) and nuclear
fractions (NE). Similarly, the expression levels of p52 and c-Rel
were not changed although the RelB expression was clearly reduced
in the DU145-siRelB cells. The band densities for the western blot
analysis were analyzed and compared between the DU145-siRelB and
DU145-sictrl cells at both CE and NE, which further supported that
the NF-κB subunits was not affected by the RelB knockdown (data not
shown). Thus, the data here indicated that the RelB knockdown by
RNAi did not affect the expression of other NF-κB family
members.
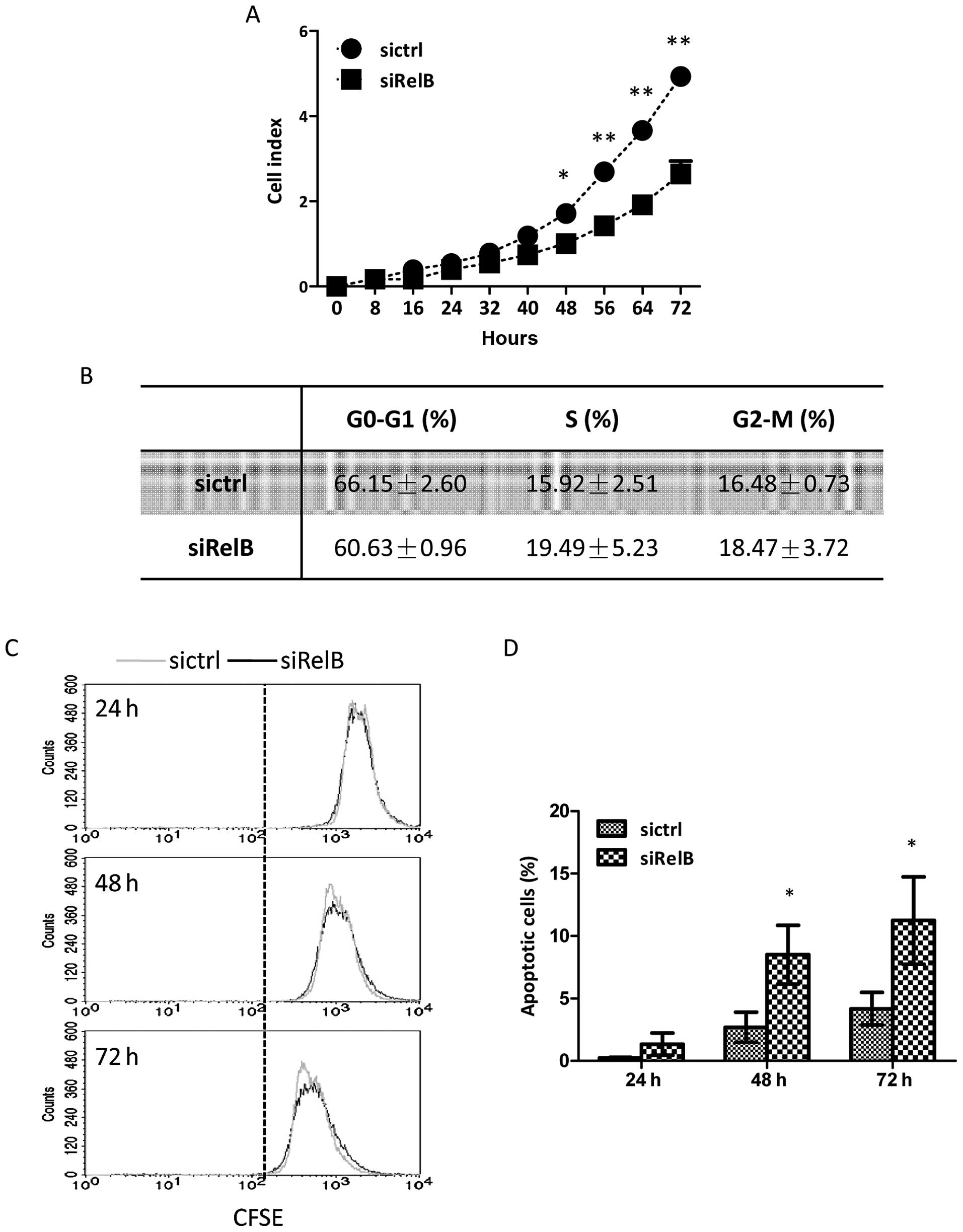
Knockdown of RelB suppresses DU145 cell
growth
Cell growth assay was performed by a real-time
xCelligence system using E-plates to investigate whether RelB might
exert a biological activity on DU145 cell growth. As shown in
Fig. 3A, the DU145-siRelB cells
grew much slower than that of the DU145-sictrl cells, and there was
a statistically significant difference between the two established
cell lines after culturing for 48 h and later. The cell growth
curve clearly indicated that RelB knockdown had suppressing effect
on prostate cancer cell growth. The cell cycle assay and cellular
DNA content measurement were performed by flow cytometry. The
percentages of the DU145-siRelB cells in the three cell cycle
phases (G0-G1, S and G2-M) were 60.63±0.96, 19.49±5.23 and
18.47±3.72%; while those of the DU145-sictrl cells were 66.15±2.60,
15.92±2.51 and 16.48±0.73%, respectively. There were no significant
differences between the two groups in cell cycle progression
(Fig. 3B).
To investigate whether the RelB might affect the
proliferation capability of DU145 cells, a CFSE cell proliferation
assay detected by flow cytometry was carried out. The fluorescence
intensities of CFSE were attenuated in both established cell lines
in a time-dependent manner. However, no significant differences
were observed between the DU145-siRelB and the DU145-sictrl cells
after culturing for 24, 48 and 72 h (Fig. 3C). Thus, RelB knockdown did not
affect the cellular proliferation ability of DU145 prostate cancer
cells.
The Annexin V/PI assay was carried out to
quantitatively analyze the apoptosis. Both the two establish cell
lines underwent spontaneous apoptosis in a time-dependent manner.
As showed in Fig. 3D, the
percentages of apoptotic cells in the DU145-sictrl group were
0.24±0.05, 2.69±1.20 and 4.17±1.30% at 24, 48 and 72 h,
respectively; while those in the DU145-siRelB group were 1.33±0.89,
8.50±2.35 and 11.25±3.50%. Spontaneous apoptosis was markedly
increased in the DU145-siRelB cells, and there were statistically
significant differences between the two cell lines in the apoptosis
rate at both 48- and 72-h time-points. Therefore, the data here
showed that RelB knockdown slowed down the cell growth of DU145
cells. The enhanced apoptosis contributed to the inhibited cell
growth upon downregulation of RelB expression in DU145 prostate
cancer cells.
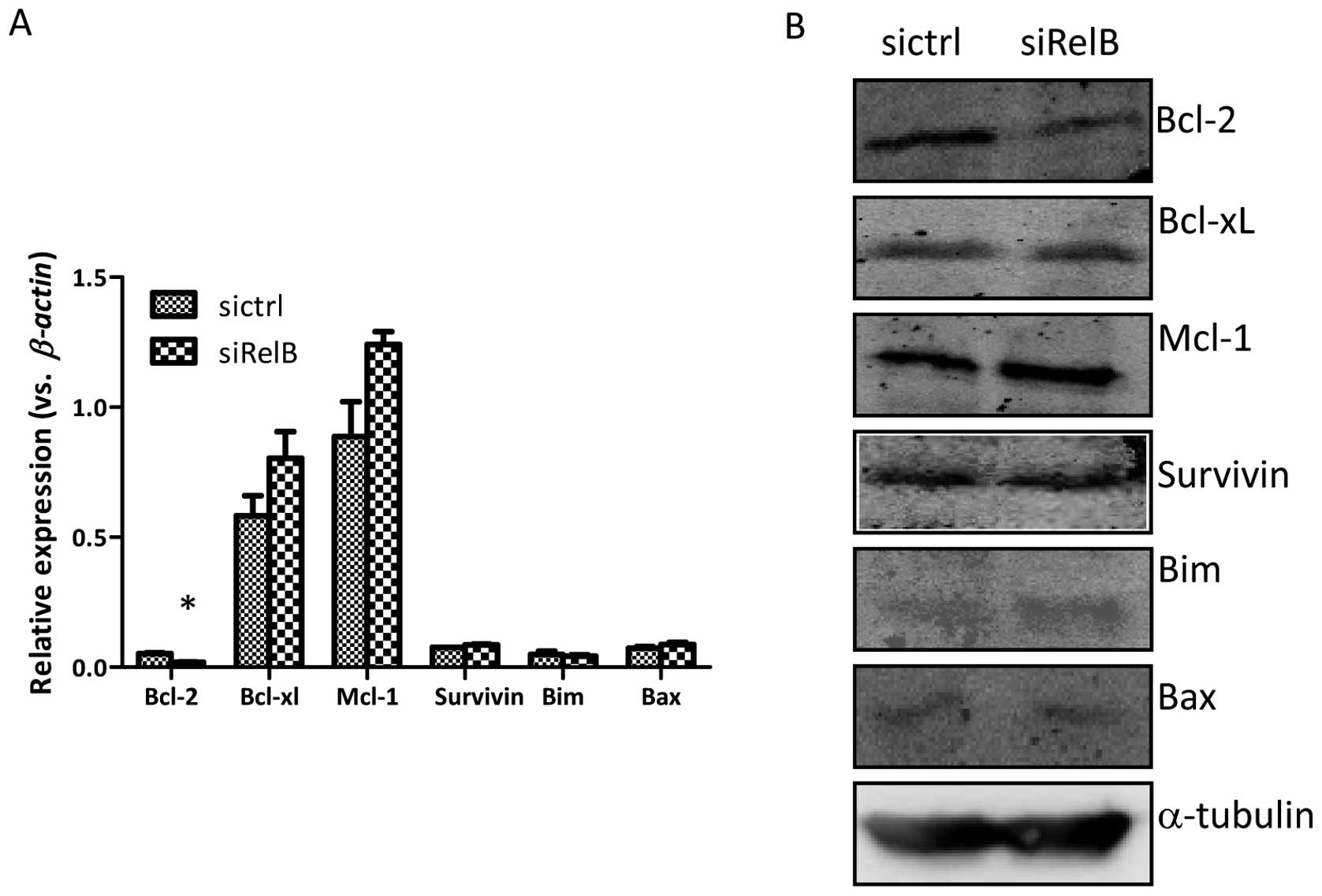
RelB regulates apoptosis-related
protein
To gain insights into the mechanism that RelB
knockdown enhanced prostate cancer cell apoptosis, the mRNA
expressions of several pro-apoptotic genes including Bax and
Bim, as well as anti-apoptotic genes including Bcl-2,
Bcl-xL, Mcl-1 and Survivin were examined by qRT-PCR
analysis. As shown in Fig. 4A, the
mRNA expression of the anti-apoptotic gene, Bcl-2, was
distinctly reduced in the DU145-siRelB cells compared to that of
the DU145-sictrl cells, which was in line with the reduced Bcl-2
protein expression measured by western blot analysis (Fig. 4B). The mRNA and protein expression
of other apoptosis-associated genes such as the pro-apoptotic genes
Bim and Bax, and the anti-apoptotic genes Bcl-xL,
Mcl-1 and Survivin appeared to be independent of RelB
status in the DU145 prostate cancer cells (Fig. 4). Collectively, our data indicate
that RelB exerted a crucial apoptosis resistance function in DU145
cells by regulating an anti-apoptotic NF-κB target gene, such as
Bcl-2.
RelB affects the migration and invasion
of DU145 cells
In order to investigate whether RelB expression
level might affect the migration ability of DU145 prostate cancer
cells, the cell migration assay was measured dynamically using the
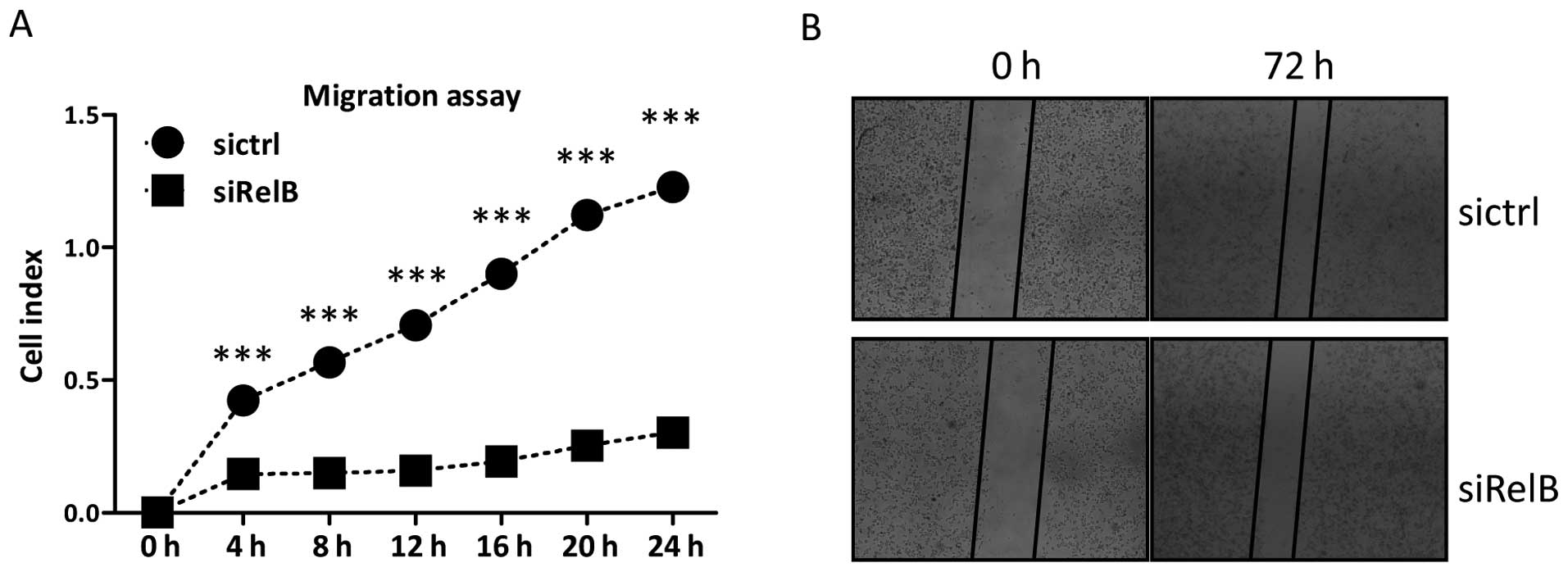
real-time xCelligence system. As shown in Fig. 5A, the DU145-siRelB cells migrated
markedly slower than the DU145-sictrl cells during the 24-h
continuous monitoring. There were significantly statistical
differences in the migration assay between the two established cell
lines. The in vitro scratch assay was also carried out to
evaluate the migration ability of DU145 cells. A scratched cell
monolayer was generated in both cell lines and images were captured
after culturing for 72 h. At the 72-h time-point of the assay, it
was shown that the DU145-siRelB cells migrated from the edge
towards the scratch center much slower than that of the
DU145-sictrl cells (Fig. 5B),
indicating a defected migratory ability of the DU145-siRelB
cells.
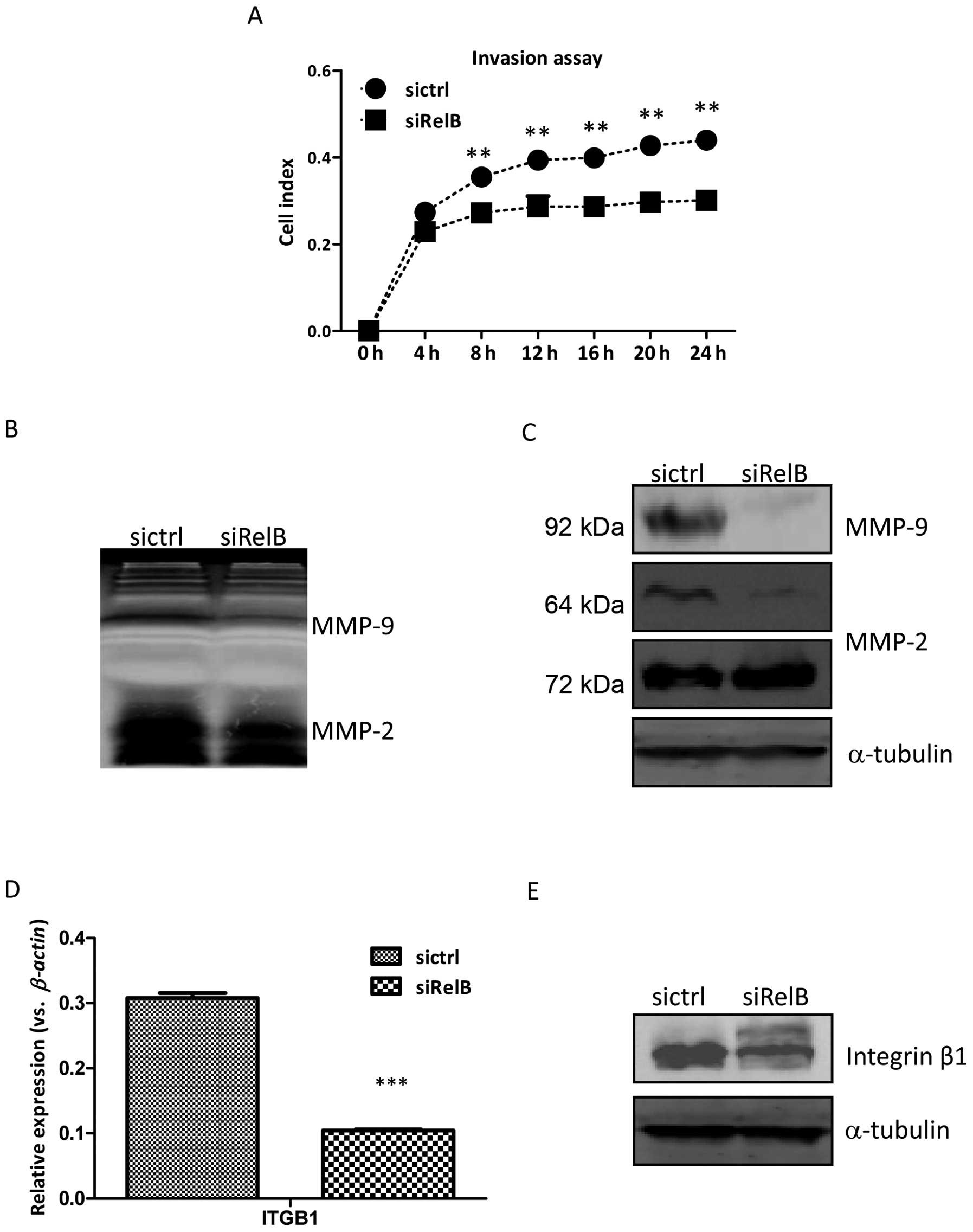
The cell invasion assay was also performed by the
real-time xCelligence system using Matrigel (dilution at
1:40)-coated CIM-plates. As shown in Fig. 6A, the DU145-siRelB cells migrated
and invaded through the Matrigel clearly slower than the
DU145-sictrl cells during the 24-h continuous observation. There
were statistically significant differences between the two
established cell lines at the time-points of 8, 12, 16, 20 and 24
h. Gelatin zymography experiment was further performed to examine
the relative amounts of active and inactive gelatinase MMP-2 or
MMP-9, members of the matrix metalloproteinases family. As shown in
Fig. 6B, the activity of MMP-2 and
MMP-9 was evidently reduced in the DU145-siRelB cells compared to
that of the DU145-sictrl cells, supporting the diminished invasion
ability of cells. Moreover, the expression of MMP-2 and MMP-9 in
the DU145-siRelB at protein level was also evidently lower than the
DU145-sictrl cells (Fig. 6C).
Integrin β-1, encoded by the ITGB1 gene,
belongs to the family of heterodimeric transmembrane cell surface
receptors. Integrin β-1 activation is a key regulator in the switch
from cellular dormancy to metastatic growth in vitro and
in vivo. The ITGB1 expression at mRNA level was
significantly decreased in the DU145-siRelB cells than that of the
DU145-sictrl cells, consistently; the protein level of integrin β-1
was also reduced in the DU145-siRelB cells (Fig. 6D and E). Thus, these results
indicated that RelB knockdown hindered the migration and invasion
abilities of DU145 prostate cancer cells, and the downregulated
integrin β-1 was implicated in these processions.
RelB regulates the radio-sensitivity of
DU145 cells
Several pieces of evidence have demonstrated that
ionizing irradiation can cause cytotoxicity to prostate cancer
cells through diverse mechanisms. In order to investigate the role
of RelB in the radio-sensitivity of prostate cancer cells, DU145
cells were subjected to exposure at 4 and 8 Gy doses of ionizing
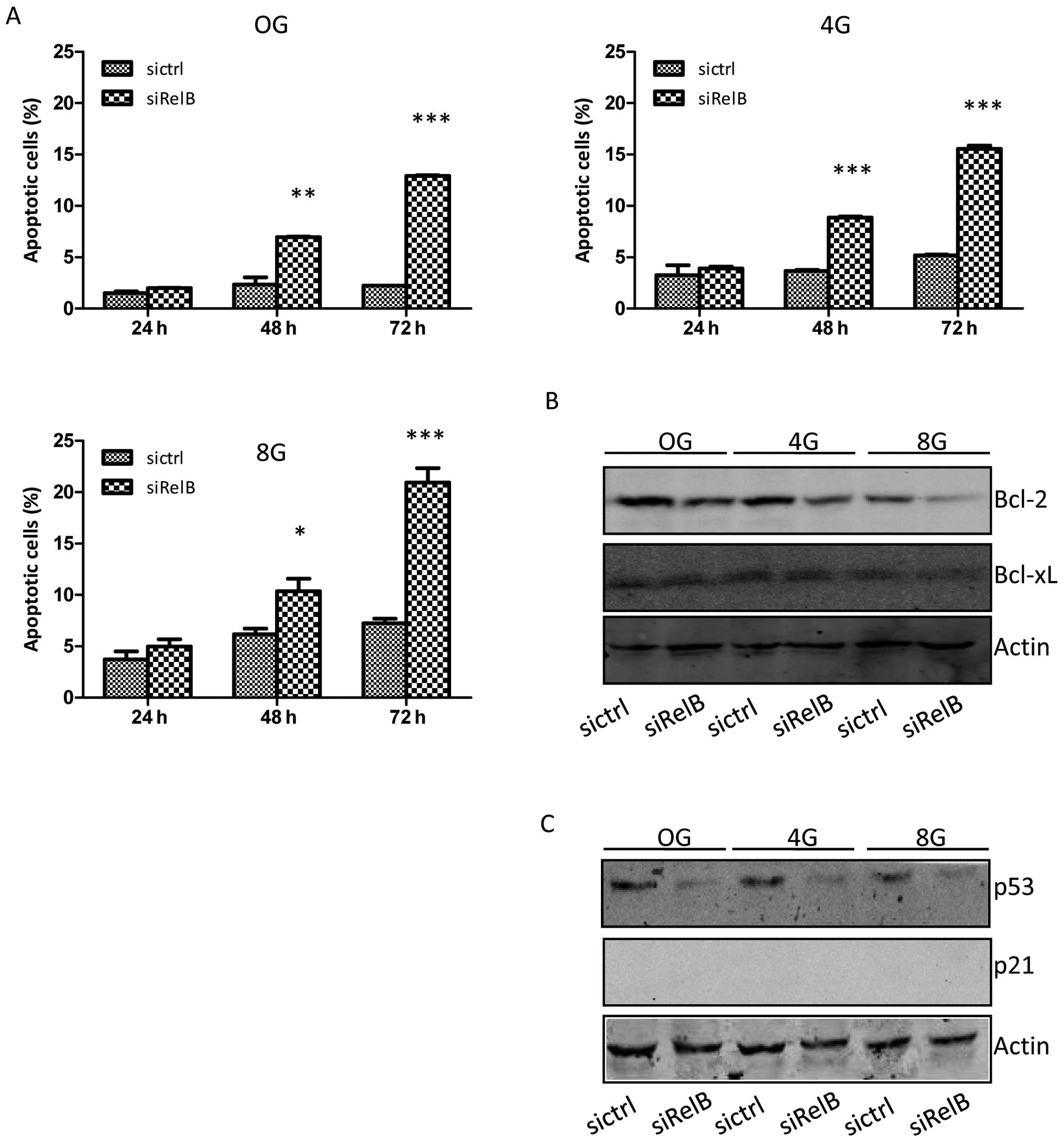
irradiation followed by cell survival assay. As shown in Fig. 7A, the frequencies of apoptotic
cells in the two established cell lines were increased in a
time-dependent and a dose-dependent manner. Response to 4 or 8 Gy
doses of ionizing irradiation, the DU145-siRelB cells had a much
higher apoptosis rate than that of the DU145-sictrl cells. There
were statistically significant differences between the two cell
lines in the apoptosis rate after expose to irradiation for 48 and
72 h.
Consistent with the results shown in Fig. 4B, the expression of one of the
anti-apoptotic proteins, Bcl-2, was clearly reduced in the
DU145-siRelB cells. The expression level of Bcl-2 was unchanged in
response to 4 Gy doses of ionizing irradiation, and was slightly
decreased in response to 8 Gy doses in the DU145-sictrl cells.
However, the expression level of Bcl-2 was decreased in a
dose-dependent manner in the DU145-siRelB cells in response to
ionizing irradiation. Another anti-apoptotic protein, Bcl-xL, was
expressed at a similar level between the DU145-sictrl and
DU145-siRelB cells. Irradiation exposure did not affect the
expression levels of Bcl-xL in the DU145-sictrl or the DU145-siRelB
cells (Fig. 7B). Cell cycle
distribution was not significantly affected in both DU145-sictrl
and DU145-siRelB cells in response to irradiation. The endogenous
expression of p53 was reduced in the DU145-siRelB cells compared to
that in the DU145-sictrl cells. The expression level of p53 was not
changed in DU145-sictrl or DU145-siRelB cells after exposure to
ionizing irradiation. The expression of p21 could not be detected
in DU145-sictrl or DU145-siRelB cells (Fig. 7C).
RelB augments proteasome
inhibitor-induced cell death
In order to detect whether RelB might affect the
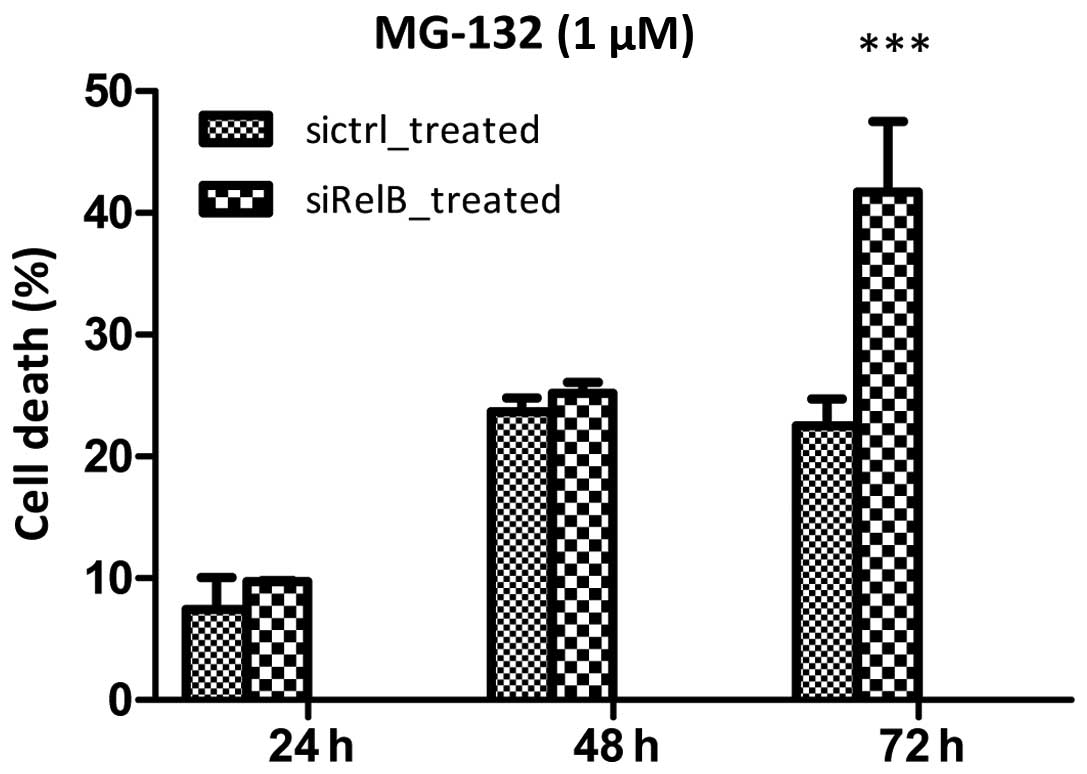
proteasome inhibitor-induced cell death of prostate cancer cells,
both cell lines were treated with MG-132 (1 μM). MG-132, a
proteasome inhibitor, caused considerable cell death in both cell
lines in a time-dependent manner (Fig.
8). After treated with MG-132 for 24 and 48 h, the cell death
in the DU145-siRelB cells was 9.71±0.10 and 25.18±0.93%,
respectively, which were similar to that in DU145-sictrl cells,
7.45±2.61 and 23.69±1.12%. Nevertheless, the percentage of cell
death in the DU145-siRelB cells was 41.75±5.75% after treated with
MG-132 for 72 h, while that in DU145-sictrl cells was only
22.53±2.18%. There was statistically significant difference between
the two established cell lines in the cell death rate after treated
with MG-132 for 72 h. Therefore, shRNA targeting RelB in the DU145
prostate cancer cells sensitized proteasome
inhibitor-treatment.
Discussion
In this study, how the non-canonical NF-κB subunit
RelB, regulates diverse biological behaviors of prostate cancer
cells was extensively examined. RelB knockdown in the DU145 human
androgen-independent prostate cancer cells using shRNA approach
influenced multiple biological processes. The suppressed functions
of the RelB knockdown in prostate cancer cell growth, migration,
invasion, and the acquired chemo-sensitivity were evidently
observed in vitro. We provide several lines of evidence that
RelB plays an important role in prostate cancer progression.
RelA, a canonical NF-κB subunit, is considered as an
independent biomarker of many human malignances including prostate
cancer. While RelA has been thoroughly examined in the initiation
and progression of prostate cancer, RelB, representing the
non-canonical NF-κB activity has been less studied in prostate
cancer. Similar to the highly aggressive prostate cancer PC-3 cell
line, the aggressive DU145 cell line is also androgen-independent
and express both the canonical and the non-canonical NF-κB
signaling molecules.
To determine the relevance of endogenous RelB
activation in diverse aspects in prostate cancer cells, RelB was
knocked down in DU145 cells using shRNA approach in the study. The
stable downregulation of RelB expression in DU145 cells did not
affect the expression of other NF-κB family members including RelA,
c-Rel, NF-κB1, and NF-κB2. We observed significant inhibition of
cell growth upon introducing shRNA-RelB into DU145 cells and found
that the enhanced apoptosis contributed to the slow cell growth.
Nevertheless, the cell cycle distribution and cellular
proliferation were not affected by the knockdown of RelB in DU145
cells. Previously, our lab has reported that not only RelA but also
RelB, subunits of NF-κB family, played an important role in the
cellular behaviors of chronic lymphocytic leukemia (CLL) cells from
bone marrow. The strength of RelB activity positively regulated the
cell survival in CLL cells. High RelB activity, together with RelA
activity, maintains the basal survival of CLL cells. In addition,
the induction of RelA and RelB expression in the nucleus is
responsible for better survival of CLL cells supported by bone
marrow stromal cells (16).
Similar observations were recently reported that RelB-silencing
decreases the viability of Hodgkin lymphoma (HL) cells
significantly while the RelA activity is intact in HL cells
(17). Silencing of RelB
transiently in DU145 cells through RNA interference method, our lab
has previously observed that RelB plays a modest role in regulating
the spontaneous cell death while RelA mediates a selective survival
advantage in DU145 prostate cancer cells (8). It has been reported that inhibiting
RelB in PC-3 cells by small interfering RNA targeting reduces cell
growth and IL-8 expression, indicating a cancer-supportive role of
RelB in prostate cancer cells (18,19).
Therefore, our findings here were consistent with previous reports
that high RelB expression supports the survival of malignant cells
when the endogenous RelA activity is constitutively activated.
However, the detailed underlying mechanism is still undefined.
To further understand the mechanism of enhanced
apoptosis upon introducing of shRNA-RelB into DU145 cells, we
examined a series of apoptosis-related molecules and finally
focused on the NF-κB regulated anti-apoptotic gene, Bcl-2. Both the
western blot and qRT-PCR analysis pointed to a clear downregulation
of the Bcl-2 when RelB expression was blocked. This was expected to
trigger the apoptosis pathway, contributing to enhanced apoptosis
in the DU145-siRelB prostate cancer cells.
The regulation of cell survival by RelB has also
been found in multiple myeloma (MM) cells. RelB-dependent induction
in cIAP2, an anti-apoptotic NF-κB target gene, constitutes a strong
pro-survival signal in MM cell lines and primary cells from MM
patients through its anti-apoptotic role. To a lesser extent, the
expression of other anti-apoptotic genes such as Bcl-xL and
Bcl-2 are also increased in MM cells (20). Abundant RelB protein is expressed
in highly aggressive human breast cancer cell lines, for example
MDA-MB-231 and Hs578T cells. RelB promotes cell survival to
irradiation and doxorubicin, likely due to the induction of
anti-apoptotic genes such as Survivin, MnSOD, and
Bcl-2 in breast cancer cells (12). Here, we presented evidence that
RelB knockdown in DU145 prostate cancer cells inhibited cell
growth, and the dysregulated expression of apoptotic-related
protein such as Bcl-2 played important roles.
Furthermore, we showed that RelB knockdown
suppressed the migration and invasion abilities of DU145 prostate
cancer cells. Very few studies have reported the involvement of
RelB in the migration and invasion of malignant cells. Apigenin
could suppress migration of PC-3 prostate cancer cells by
inhibiting the IKKα/NF-κB activation (13). The nuclear IKKα enhances metastasis
by repressing transcription of the Maspin gene (14), which inhibits cancer metastasis by
suppressing invasion and motility (21). RelB is found to repress the
expression of Maspin in prostate cancer cells, however, the
detailed investigation in this process still demands more efforts
(8). In breast cancer cell lines,
RelB represses the expression of estrogen receptor α (ERα) via the
induction of the B-lymphocyte-induced maturation protein (Blimp1),
which promotes a more migratory phenotype in breast and lung cancer
cells (22,23). Notably, RelB promotes a more
invasive phenotype in ERα-negative cancer via the induction of the
anti-apoptotic gene, Bcl-2. RelB also functions in epithelial to
mesenchymal transition (EMT) in breast cancer cells, and Bcl-2 is
an important downstream mediator (24). Protection from apoptosis by Bcl-2
has been proposed to have an important role in promoting
metastasis. Knockdown of adhesion G protein-coupled receptors
(ADGRG2) in Hs578T and MDA-MB-231 breast cancer cells causes a
strong reduction in cell adhesion and subsequent cell migration,
which is associated with RelB reduction (25). A recent report suggests that RelB
plays a critical role in promoting fibroblast migration prolonged
tumor necrosis factor (TNF)-α treatment. The migration-associated
gene, the matrix metallopeptidase 3 (MMP-3) was identified as a
novel target of RelB (26). For
the first time, we provided evidence that the RelB played a
positive role in regulating the migration and invasion of DU145
prostate cancer cells, and therefore it functioned indeed as an
oncogene in prostate cancer cells.
Diverse molecular regulators, including adhesion
receptor families, receptor tyrosine kinase, cytoskeleton protein,
adaptors, and signaling molecules, are involved in the regulation
of migration and invasion of cancer cells. Interestingly, we found
that the expression of integrin β-1 was downregulated in the
DU145-siRelB cells. Integrin β-1, encoded by the ITGB1 gene,
belongs to the family of heterodimeric transmembrane cell surface
receptors that contain 18α and 8β subunits and bridge the
interaction between cell-cell and cell-extracellular matrix (ECM).
Overexpression of integrin β-1 has been found in many epithelial
malignancies during invasion, angiogenesis, and metastasis.
Integrin β-1 silencing suppresses lung cancer cell invasion and
metastasis in vitro and in vivo (27). Integrin β-1 and integrin-induced
autophosphorylation of focal adhesion kinase (FAK) are increased in
prostate cancer cells, correlating with metastatic potential in
vivo (28). There is a direct
interaction between Maspin and integrin β-1 by the reactive centre
loop of Maspin (29). The
activation of the small cell lung cancer (SCLC) H69 cell line by
lipopolysaccharide (LPS) causes the induction of RelB and p100
expression. Blocking RelB expression prevents the induction of
integrin β-1 and the attachment of H69 cells. Since the
ITGB1 promoter does not contain NF-κB consensus sequences,
it is conceivable that the LPS-mediated induction of integrin β-1
could be through the expression of HIF-1α (30). To understand whether similar
mechanism would occur in the prostate cancer cells is
warranted.
Radiotherapy is one of the most commonly used
therapies for prostate cancer. However, radio-recurrent prostate
cancer and a poor long-term prognosis are experienced by many
prostate cancer patients, as ~30–40% of those individuals treated
with potentially curative doses develop radio-resistance (31). RelB complex, representing the
alternative NF-κB activity, has been shown to confer
radio-resistance in prostate cancer, in part, by stimulating the
expression of MnSOD. Selective blocking RelB activation by SN52
suppresses MnSOD expression and sensitizes PC-3 prostate cancer
cells to radiation (10,32). Bcl-xL, an NF-κB target
anti-apoptotic gene, whose response is widely thought to be
involved in both chemo-resistance and radio-resistance, has also
been modulated by RelB in malignant cells. As shown in a recent
study, RelB-siRNA transfection into murine RM-1 prostate cancer
cells increases radiation-induced apoptosis by inhibiting the
expression of the Bcl-xL gene (33). In our study, RelB knockdown in
DU145 prostate cancer cells sensitized radiation-induced apoptosis,
which was in line with previous studies. Unfortunately, we could
not perform the clonogenic survival assay successfully for
DU145-sictrl or DU145-siRelB cells after irradiation. Though, both
DU145-sictrl and DU145-siRelB cells without irradiation formed
colonies successfully. The number of colonies was evidently low
when cultured using the DU145-siRelB cells, indicating the poor
survival capacity (data not shown). A marked reduction of Bcl-2
expression was observed in response to irradiation in the presence
of RelB knockdown, nevertheless, changes in the expression of
Bcl-xL in response to irradiation were not detect in the
DU145-sictrl or DU145-siRelB cells. The expression of Bcl-2 was
also slightly reduced in the DU145-sictrl cells after exposure to
8-Gy doses of ionizing irradiation, which was correlated with the
increased apoptosis. However, the expression level of Bcl-2 was
decreased in a dose-dependent manner in the DU145-siRelB cells in
response to ionizing irradiation. The reduction of Bcl-2 level also
corresponded to the induction of apoptosis. Bcl-2 has been
previously reported as a direct target of RelB in breast cancer
cells (24). Bcl-2 plays an
important role in radio-resistance of diverse cancer cells. The
Bcl-2 protein expression after radiotherapy correlates with both
response and survival. The Bcl-2 positive cancers show
significantly poorer response than the Bcl-2 negative cancers after
radiation (34). In this study, we
found that the expression of Bcl-2 was regulated by RelB,
indicating that Bcl-2 is a potential target gene of RelB in
prostatic cancer cells. The Bcl-2 expression was reduced both in
the DU145-sictrl and DU145-siRelB cells after irradiation, although
with different levels. The reduced Bcl-2 expression correlated with
the increased apoptosis and the improved radio-sensitivity after
irradiation. Overall, when RelB was silenced in the DU145 cells,
the radio-sensitivity was largely increased due to the reduction of
Bcl-2. However, the potential mechanism underlying the reduction of
Bcl-2 after irradiation in the DU145 cells demands further study.
The cell cycle distribution was not significantly affected in
DU145-sictrl or DU145-siRelB cells. The expression of p53 was
reduced in the DU145-siRelB cells compared to that in the
DU145-sictrl cells. The expression level of p53 was not changed in
DU145-sictrl or DU145-siRelB cells after exposure to ionizing
irradiation. The expression of p21 could not be detected in
DU145-sictrl or DU145-siRelB cells.
Besides its role in carcinogenesis of human cancers,
RelB might play a role in chemo-sensitivity. STI571, a tyrosine
kinase inhibitor, is effective for treating androgen-independent,
but not androgen-responsive prostate cancer cells in the context of
high levels of the RelB activity (35). Targeting of the NF-κB signaling
pathway is proposed as a promising therapeutic choice in cancer. An
indirect approach to target NF-κB is using proteasome inhibitors
such as PS-341 (bortezomib) or MG-132, which induce apoptosis in a
variety of malignant cells. Bortezomib manifests remarkable
anticancer activity in several hematological malignancies such as
MM. Bortezomib is less effective in prostate cancer and other solid
tumors; however, the mechanisms have not been fully understood.
Recent studies suggest that bortezomib treatment in PC-3 and DU145
prostate cancer cells unexpectedly induces the expression of
pro-inflammatory chemokine IL-8, which promotes cancer cell
proliferation, survival, and angiogenesis. Moreover, bortezomib
increases nuclear accumulation of IKKα, and suppression of IKKα
protein levels and enzymatic activity significantly decreases the
bortezomib induced IL-8 expression. Thus, the inactivation of the
non-canonical NF-κB activity is expected to convey better effects
of proteasome inhibitor (36,37).
In this study, we illustrated that RelB knockdown in DU145 cells
enhanced chemo-sensitivity to the proteasome inhibitor MG-132.
Treated with MG-132 for 72 h, the DU145-siRelB cells showed much
more cell death than the DU145-sictrl cells, suggesting a
supporting role of RelB in proteasome inhibitor resistance in
prostate cancer cells. It has been reported that inhibition of RelB
may be one of the principal mechanisms of action of proteasome
inhibitor in MM cells (38).
Therefore, low RelB expression in DU145 prostate cancer cells was
positively correlated with proteasome inhibitor sensitivity. One of
the main functions of proteasome inhibitor is the suppressed
proteasomal degradation of IκBα, resulting in the inhibition of
NF-κB activity and expression of NF-κB target genes. NF-κB activity
is constitutively increased in metastatic prostate cancer cells
through the increased activation of IKK, resulting in the increased
cell survival and resistance to chemotherapy. Proteasome inhibitor
has so far failed to exhibit a significant clinical activity in
prostate cancer patients; the mechanisms are largely unknown. It
has been reported that proteasome inhibitor could increase the
nuclear levels of RelA in prostate cancer cells due to preventing
the proteasomal degradation of nuclear RelA. Targeting both IKKα
may increase the proteasome inhibitor effectiveness in
androgen-independent prostate cancer treatment. IKKα is a critical
molecule in the activation of the non-canonical NF-κB signaling
pathway, leading to the activation of RelB predominantly.
Therefore, it is assumed that targeting RelB might improve
proteasome inhibitor sensitivity. Nevertheless, the molecular
mechanism underling the improved sensitivity to proteasome
inhibitor treatment when RelB is silenced in the prostatic cancer
cells is warranted.
The above findings establish a key tumor-promoting
role of the non-canonical NF-κB activity subunit RelB in the
carcinogenesis of prostate cancer. Our data shed light on
unexplored aspects of RelB in prostate cancer cells. RelB
controlled the spontaneous and radiation-induced cell survival of
prostate cancer cells by regulating Bcl-2 predominantly. RelB
played a supportive role in the migration and invasion abilities of
prostate cancer cells by regulating integrin β-1. Given the potent
effects on cell growth, migration, invasion, and chemo-resistance,
RelB represents an attractive target for cancer therapy. To address
the function of RelB in vivo, further studies will be
performed in the future.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (F.G., grant no. 81172433) and the
Natural Science Foundation of Jiangsu Provincial (F.G., grant no.
BK20151211).
References
|
1
|
Siegel RL, Fedewa SA, Miller KD,
Goding-Sauer A, Pinheiro PS, Martinez-Tyson D and Jemal A: Cancer
statistics for Hispanics/Latinos, 2015. CA Cancer J Clin.
65:457–480. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Napetschnig J and Wu H: Molecular basis of
NF-κB signaling. Annu Rev Biophys. 42:443–468. 2013. View Article : Google Scholar :
|
|
3
|
Weih F and Caamaño J: Regulation of
secondary lymphoid organ development by the nuclear factor-kappaB
signal transduction pathway. Immunol Rev. 195:91–105. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Perkins ND: The diverse and complex roles
of NF-κB subunits in cancer. Nat Rev Cancer. 12:121–132.
2012.PubMed/NCBI
|
|
5
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Suh J and Rabson AB: NF-kappaB activation
in human prostate cancer: Important mediator or epiphenomenon? J
Cell Biochem. 91:100–117. 2004. View Article : Google Scholar
|
|
7
|
Lessard L, Bégin LR, Gleave ME, Mes-Masson
AM and Saad F: Nuclear localisation of nuclear factor-kappaB
transcription factors in prostate cancer: An immunohistochemical
study. Br J Cancer. 93:1019–1023. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo F, Kang S, Zhou P, Guo L, Ma L and Hou
J: Maspin expression is regulated by the non-canonical NF-κB
subunit in androgen-insensitive prostate cancer cell lines. Mol
Immunol. 49:8–17. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lessard L, Saad F, Le Page C, Diallo JS,
Péant B, Delvoye N and Mes-Masson AM: NF-kappaB2 processing and p52
nuclear accumulation after androgenic stimulation of LNCaP prostate
cancer cells. Cell Signal. 19:1093–1100. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Holley AK, Xu Y, St Clair DK and St Clair
WH: RelB regulates manganese superoxide dismutase gene and
resistance to ionizing radiation of prostate cancer cells. Ann NY
Acad Sci. 1201:129–136. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu Y, Fang F, St Clair DK, Josson S,
Sompol P, Spasojevic I and St Clair WH: Suppression of
RelB-mediated manganese superoxide dismutase expression reveals a
primary mechanism for radiosensitization effect of
1alpha,25-dihydroxyvitamin D(3) in prostate cancer cells. Mol
Cancer Ther. 6:2048–2056. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mineva ND, Wang X, Yang S, Ying H, Xiao
ZX, Holick MF and Sonenshein GE: Inhibition of RelB by
1,25-dihydroxyvitamin D3 promotes sensitivity of breast cancer
cells to radiation. J Cell Physiol. 220:593–599. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shukla S, Kanwal R, Shankar E, Datt M,
Chance MR, Fu P, MacLennan GT and Gupta S: Apigenin blocks IKKα
activation and suppresses prostate cancer progression. Oncotarget.
6:31216–31232. 2015.PubMed/NCBI
|
|
14
|
Luo JL, Tan W, Ricono JM, Korchynskyi O,
Zhang M, Gonias SL, Cheresh DA and Karin M: Nuclear
cytokine-activated IKKalpha controls prostate cancer metastasis by
repressing Maspin. Nature. 446:690–694. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guo F, Sun A, Wang W, He J, Hou J, Zhou P
and Chen Z: TRAF1 is involved in the classical NF-kappaB activation
and CD30-induced alternative activity in Hodgkin's lymphoma cells.
Mol Immunol. 46:2441–2448. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu J, Zhou P, Wang W, Sun A and Guo F:
RelB, together with RelA, sustains cell survival and confers
proteasome inhibitor sensitivity of chronic lymphocytic leukemia
cells from bone marrow. J Mol Med Berl. 92:77–92. 2014. View Article : Google Scholar
|
|
17
|
Ranuncolo SM, Pittaluga S, Evbuomwan MO,
Jaffe ES and Lewis BA: Hodgkin lymphoma requires stabilized NIK and
constitutive RelB expression for survival. Blood. 120:3756–3763.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu Y, Josson S, Fang F, Oberley TD, St
Clair DK, Wan XS, Sun Y, Bakthavatchalu V, Muthuswamy A and St
Clair WH: RelB enhances prostate cancer growth: Implications for
the role of the nuclear factor-kappaB alternative pathway in
tumorigenicity. Cancer Res. 69:3267–3271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu Y, Fang F, St Clair DK and St Clair WH:
Inverse relationship between PSA and IL-8 in prostate cancer: An
insight into a NF-κB-mediated mechanism. PLoS One. 7:e329052012.
View Article : Google Scholar
|
|
20
|
Cormier F, Monjanel H, Fabre C, Billot K,
Sapharikas E, Chereau F, Bordereaux D, Molina TJ, Avet-Loiseau H
and Baud V: Frequent engagement of RelB activation is critical for
cell survival in multiple myeloma. PLoS One. 8:e591272013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou J, Hualong Q, Zhou P and Guo F:
Different maspin functions in the lung adenocarcinoma A549 and
SPC-A1 cell lines. Int J Mol Med. 36:1440–1448. 2015.PubMed/NCBI
|
|
22
|
Wang X, Belguise K, O'Neill CF,
Sánchez-Morgan N, Romagnoli M, Eddy SF, Mineva ND, Yu Z, Min C,
Trinkaus-Randall V, et al: RelB NF-kappaB represses estrogen
receptor alpha expression via induction of the zinc finger protein
Blimp1. Mol Cell Biol. 29:3832–3844. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu Z, Sato S, Trackman PC, Kirsch KH and
Sonenshein GE: Blimp1 activation by AP-1 in human lung cancer cells
promotes a migratory phenotype and is inhibited by the lysyl
oxidase propeptide. PLoS One. 7:e332872012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang X, Belguise K, Kersual N, Kirsch KH,
Mineva ND, Galtier F, Chalbos D and Sonenshein GE: Oestrogen
signalling inhibits invasive phenotype by repressing RelB and its
target BCL2. Nat Cell Biol. 9:470–478. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Peeters MC, Fokkelman M, Boogaard B,
Egerod KL, van de Water B, IJzerman AP and Schwartz TW: The
adhesion G protein-coupled receptor G2 (ADGRG2/GPR64)
constitutively activates SRE and NFκB and is involved in cell
adhesion and migration. Cell Signal. 27:2579–2588. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Authier H, Billot K, Derudder E,
Bordereaux D, Rivière P, Rodrigues-Ferreira S, Nahmias C and Baud
V: IKK phosphorylates RelB to modulate its promoter specificity and
promote fibroblast migration downstream of TNF receptors. Proc Natl
Acad Sci USA. 111:14794–14799. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang XM, Li J, Yan MX, Liu L, Jia DS, Geng
Q, Lin HC, He XH, Li JJ and Yao M: Integrative analyses identify
osteopontin, LAMB3 and ITGB1 as critical pro-metastatic genes for
lung cancer. PLoS One. 8:e557142013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee YC, Jin JK, Cheng CJ, Huang CF, Song
JH, Huang M, Brown WS, Zhang S, Yu-Lee LY, Yeh ET, et al: Targeting
constitutively activated β1 integrins inhibits prostate cancer
metastasis. Mol Cancer Res. 11:405–417. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ravenhill L, Wagstaff L, Edwards DR, Ellis
V and Bass R: G-helix of maspin mediates effects on cell migration
and adhesion. J Biol Chem. 285:36285–36292. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Saito T, Sasaki CY, Rezanka LJ, Ghosh P
and Longo DL: p52-independent nuclear translocation of RelB
promotes LPS-induced attachment. Biochem Biophys Res Commun.
391:235–241. 2010. View Article : Google Scholar :
|
|
31
|
Xie BX, Zhang H, Yu L, Wang J, Pang B, Wu
RQ, Qian XL, Li SH, Shi QG, Wang LL, et al: The radiation response
of androgen-refractory prostate cancer cell line C4-2 derived from
androgen-sensitive cell line LNCaP. Asian J Androl. 12:405–414.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu Y, Fang F, St Clair DK, Sompol P,
Josson S and St Clair WH: SN52, a novel nuclear factor-kappaB
inhibitor, blocks nuclear import of RelB:p52 dimer and sensitizes
prostate cancer cells to ionizing radiation. Mol Cancer Ther.
7:2367–2376. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhu L, Zhu B, Yang L, Zhao X, Jiang H and
Ma F: RelB regulates Bcl-xL expression and the irradiation-induced
apoptosis of murine prostate cancer cells. Biomed Rep. 2:354–358.
2014.PubMed/NCBI
|
|
34
|
Harima Y, Harima K, Shikata N, Oka A,
Ohnishi T and Tanaka Y: Bax and Bcl-2 expressions predict response
to radiotherapy in human cervical cancer. J Cancer Res Clin Oncol.
124:503–510. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu Y, Fang F, Sun Y, St Clair DK and St
Clair WH: RelB-dependent differential radiosensitization effect of
STI571 on prostate cancer cells. Mol Cancer Ther. 9:803–812. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Manna S, Singha B, Phyo SA, Gatla HR,
Chang TP, Sanacora S, Ramaswami S and Vancurova I: Proteasome
inhibition by bortezomib increases IL-8 expression in
androgen-independent prostate cancer cells: The role of IKKα. J
Immunol. 191:2837–2846. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Singha B, Gatla HR, Manna S, Chang TP,
Sanacora S, Poltoratsky V, Vancura A and Vancurova I: Proteasome
inhibition increases recruitment of IκB kinase β (IKKβ), S536P-p65,
and transcription factor EGR1 to interleukin-8 (IL-8) promoter,
resulting in increased IL-8 production in ovarian cancer cells. J
Biol Chem. 289:2687–2700. 2014. View Article : Google Scholar
|
|
38
|
Garber K: Gene mutation revelation points
to new target for myeloma treatment, studies say. J Natl Cancer
Inst. 99:1362–1364. 2007. View Article : Google Scholar : PubMed/NCBI
|