Introduction
Lung cancer remains the most frequent cause of
cancer-related death in developed countries (1). Approximately 80% of lung cancers are
categorized as non-small cell lung cancer (NSCLC), and NSCLC is
divided into four major subtypes pathologically: adenocarcinoma,
squamous cell carcinoma (SCC), large cell carcinoma and
neuroendocrine cancer (2).
Development of targeted molecular therapies has remarkably improved
the overall survival of patients with adenocarcinoma (3–6). In
contrast, targeted molecular therapies have offered few benefits in
the management of lung SCC (7).
Therefore, there is a need for effective treatment options based on
current genomic approaches for lung SCC.
After the human genome sequencing era, the discovery
of an extremely large number of non-coding RNAs conceptually
transformed cancer research. MicroRNAs (miRNAs) are small
non-coding RNAs involved in the repression or degradation of target
RNA transcripts in a sequence-dependent manner (1,2). The
ability of miRNAs is unique; a single miRNA may regulate a vast
number of protein-coding or non-coding RNAs in human cells.
Therefore, aberrantly expressed miRNAs may upset tightly regulated
cellular RNA networks. This failing of the RNA network contributes
to cancer development, aggressiveness and drug resistance (3–5). In
fact, aberrantly expressed miRNAs have been reported in various
types of human cancers, including lung SCC (6–8).
Based on miRNA expression signatures, we identified
tumor-suppressive miRNAs and the cancer RNA networks regulated by
these miRNAs (9–11). We hypothesize that elucidation of
tumor-suppressive miRNA-regulated oncogenic networks will provide
new insights into the potential molecular mechanisms of lung
SCC.
In our earlier studies determining miRNA expression
signatures, we showed that miRNA-218 was significantly
downregulated in several types of cancer tissues (12–15).
Our previous studies also demonstrated that downregulation of
miR-218 enhanced overexpression of extracellular matrix
(ECM) protein components or actin-related proteins, and this
promoted cancer cell migration and invasion (16–18).
Tumor-suppressive roles of miR-218 were reported in several
types of cancer. However, the impact of miR-218 on lung SCC
remains ambiguous.
The aim of the present study was to investigate the
functional significance of miR-218 in lung SCC and to
identify molecular targets regulated by this miRNA. We found that
restoration of miR-218 significantly suppressed cancer cell
migration and invasion. Using luciferase reporter assay, tumor
protein D52 (TPD52) was shown to be directly regulated by
miR-218. Overexpression of TPD52 was observed in lung
SCC clinical specimens and downregulation of the TPD52 gene
significantly inhibited cancer cell aggressiveness. miR-218/
TPD52-regulated RNA networks may provide new insights into the
potential mechanisms of lung SCC pathogenesis.
Materials and methods
Clinical specimens, cancer cell lines and
RNA extraction
In all, 31 lung SCC specimens and 24 non-cancerous
lung specimens were obtained from patients who underwent thoracic
surgery at Kagoshima University Hospital from 2010 to 2013. The
present study was approved by the Institutional Review Board for
Clinical Research of the Kagoshima University School of Medicine.
Prior written informed consent and approval were obtained from each
patient. Table I describes the
patient background and clinical characteristics of collected
specimens. Samples were staged using the TNM scheme according to
the International Association for the Study of Lung Cancer and were
graded histologically (19). These
archival formalin-fixed paraffin-embedded (FFPE) samples were
subsequently studied by qRT-PCR analysis and immunohistochemistry
as previously described (9–11).
 | Table ICharacteristics of the lung cancer
and non-cancerous cases. |
Table I
Characteristics of the lung cancer
and non-cancerous cases.
| A, Characteristics
of the lung cancer cases |
|---|
|
|---|
| Lung cancer
patients | n | (%) |
|---|
| Total no | 31 | |
| Median age
(range) | 70 (50–88) | |
| Gender |
| Male | 29 | (93.5) |
| Female | 2 | (6.5) |
| Pathological
stage |
| IA | 4 | (12.9) |
| IB | 9 | (29.0) |
| IIA | 4 | (12.9) |
| IIB | 5 | (16.1) |
| IIIA | 8 | (25.8) |
| IIIB | 1 | (3.2) |
|
| B, Characteristics
of the non-cancerous cases |
|
| Non-cancerous
tissues | n |
|
| Total number | 24 |
| Median age
(range) | 69 (50–88) |
| Gender |
| Male | 24 |
| Female | 0 |
The human lung SCC cell lines EBC-1 and SK-MES-1
were acquired from the Japanese Cancer Research Resources Bank
(Osaka, Japan) and the American Type Culture Collection ATCC;
Manassas, VA, USA), respectively.
Total RNA was isolated using Isogen II (code no:
311-07361; Nippon Gene, Co., Ltd., Tokyo, Japan) as directed by the
manufacturer.
Quantitative reverse transcription PCR
(qRT-PCR)
PCR quantification was performed as previously
described (9–11). The expression of miR-218 was
determined using stem-loop RT-PCR as directed by the manufacturer
(P/N: 000521; Applied Biosystems, Foster City, CA, USA). The TaqMan
probe and TPD52 primers were from Assay-on-Demand™ Gene
Expression products (P/N: Hs00893105_m1; Applied Biosystems). For
quantification, miRNA and mRNA data were normalized against human
RNU48 (P/N: 001006; Applied Biosystems) and GUSB
(P/N: Hs99999908_m1; Applied Biosystems), respectively.
Transfection of mature miRNA and small
interfering RNA (siRNA)
Pre-miR™ miRNA precursors for miR-218
(hsa-miR-218-5p, P/N: AM1 7100; Applied Biosystems) and
negative control miRNA (P/N: AM 17111; Applied Biosystems), Stealth
Select RNAi siRNA, si-TPD52 (P/N: HSS120730 and HSS120731;
Invitrogen, Carlsbad, CA, USA), and negative control siRNA (P/N:
4390843; Invitrogen) were used in this study. EBC-1 and SK-MES-1
cells in Opti-MEM medium (cat. no. 31985070; Thermo Fisher
Scientific, Waltham, MA, USA) were transfected with Lipofectamine
RNAiMAX transfection reagent (P/N: 56532; Invitrogen) with 10 nM
mature miRNA or siRNA.
Cell proliferation, migration and
invasion assays
Cell proliferation was determined by XTT assay using
Cell Proliferation kit (SKU: 20-300-1000; Biological Industries,
Kibbutz Beit Haemek, Israel). Cell migration activity was analyzed
by wound-healing assay, and cell invasion was analyzed using
Corning BioCoat Matrigel Invasion chamber (cat. no. 354480; BD
Biosciences, Bedford, MA, USA). The cell proliferation, migration,
and invasion assays were carried out as previously described
(9–11).
Identification of putative miR-218 target
genes in lung SCC cells
Genome-wide gene expression analysis of
miR-218-transfected EBC-1 cells was performed (accession
number: GSE77790). Putative miR-218-regulated genes were
identified by searching the TargetScan database (http://www.targetscan.org/). We then examined the
expression levels of putative miR-218 targets in lung SCC
clinical expression data from the GEO database (accession number:
GSE19188). Oligo-microarray procedures and data mining methods were
conducted as previously described (20,21).
Western blot analysis
Cells were harvested 96 h after transfection, and
proteins were extracted from lysed cells. Protein lysates (20 μg)
were separated on NuPAGE 4–12% Bis-Tris gels (cat. no. NP0323BOX;
Invitrogen) before transfer of proteins to a polyvinylidene
fluoride membrane. Immunoblotting was performed using diluted
primary anti-TPD52 antibodies (1:250 dilution; Human Protein Atlas
no. HPA028427; Atlas Antibodies, Stockholm, Sweden) and anti-GAPDH
antibodies (1:10,000 dilution; cat. no. MAB374; Chemicon
International, Inc., Temecula, CA, USA). These assays were carried
out as previously described (9–11).
Plasmid construction and dual-luciferase
reporter assay
The procedure for the dual-luciferase reporter assay
was previously described (9–11). A
partial sequence of the wild-type TPD52 3′-UTR containing
the miR-218 target site or the TPD52 3′-UTR partial
sequence lacking the miR-218 target site was cloned into the
psiDHECK-2 vector between the XhoI-PmeI restriction
sites in the 3′-UTR of the hRluc gene (cat. no. C8021;
Promega, Madison, WI, USA).
Immunohistochemistry
The expression status of TPD52 in lung SCC
clinical specimens (BC04002; US Biomax, Inc., Rockville, MD, USA)
was confirmed via immunohistochemistry using an UltraVision
Detection system (cat. no. TP-015-HD, Thermo Fisher Scientific)
according to the manufacturer's protocol. Tissues were incubated
with primary rabbit polyclonal anti-TPD52 antibodies
(1:3,000 dilution; HPA028427) then treated with biotinylated goat
anti-rabbit secondary antibodies. Antibodies were visualized using
diaminobenzidine hydrogen peroxidase as the chromogen, and slides
were counterstained with 0.5% hematoxylin.
Identification of downstream targets
regulated by TPD52 in lung SCC
Gene expression analysis using
si-TPD52-transfected EBC-1 cells identified molecular
targets regulated by TPD52 in lung SCC cells. Microarray
expression profiles of si-TPD52 transfectants were compiled
and deposited into the GEO database (accession number:
GSE82108).
Statistical analysis
RT-PCR results were analyzed using Mann-Whitney U
tests to evaluate the relationships between the 2 groups, while
Bonferroni-adjusted Mann-Whitney U tests were used to analyze the
relationships among three or more variables. All analyses were
performed using Expert StatView (version 5; SAS Institute Inc.,
Cary, NC, USA).
Results
Expression of miR-218 in lung SCC tissues
and cell lines
To confirm the miRNA expression signatures of lung
SCC cells, we evaluated the expression level of miR-218 in
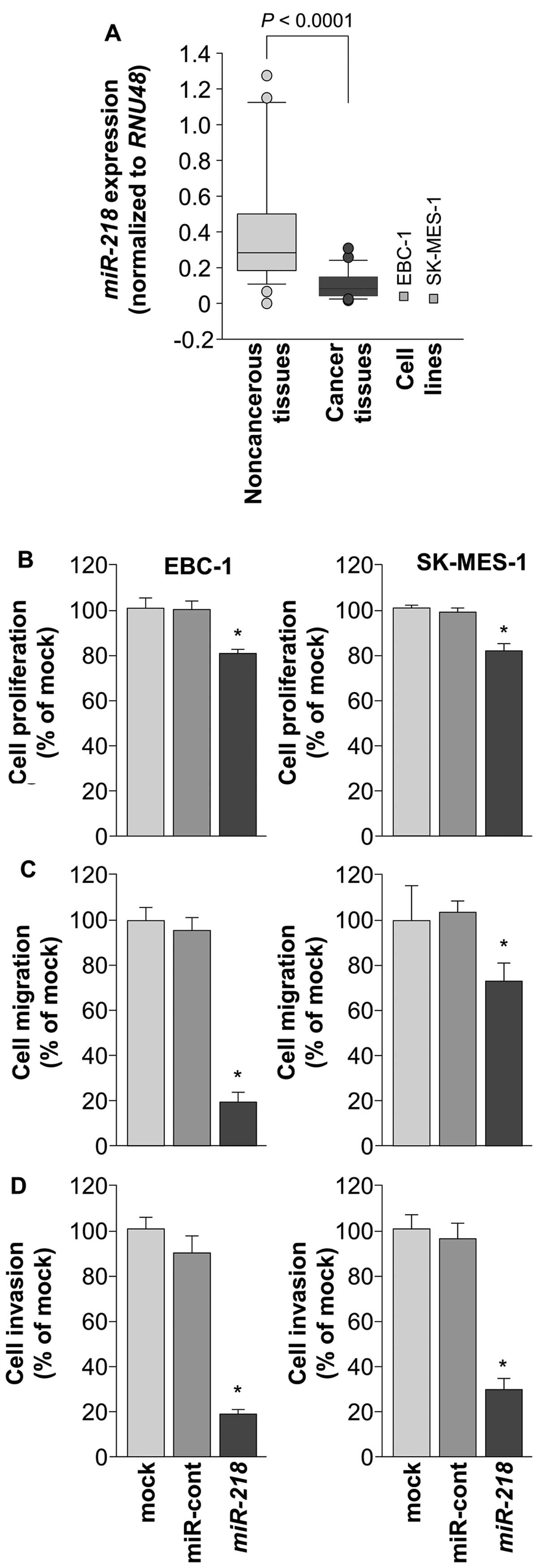
lung SCC tissues (n=31) and non-cancerous lung tissues (n=24). The
patient clinicopathological features are summarized in Table I. The expression level of
miR-218 was significantly downregulated in cancer tissues
compared to non-cancerous tissues (P<0.0001; Fig. 1A). Expression of miR-218 in
EBC-1 and SK-MES-1 cell lines was also lower than in non-cancerous
lung tissues (Fig. 1A).
Effects of miR-218 restoration on the
proliferation, migration and invasion of EBC-1 and SK-MES-1
cells
To investigate the antitumor activity of
miR-218, we performed gain-of-function assays using miRNA
transfection into EBC-1 and SK-MES-1 cells. Cell proliferation
assays showed that cancer cell growth was slightly inhibited in
miR-218 transfectants compared with mock- or miR
control-transfected EBC-1 and SK-MES-1 cells (Fig. 1B). Cancer cell migration and
invasion activities were significantly inhibited in miR-218
transfectants compared with mock- or miR-control-transfectants
(P<0.0001; Fig. 1C and D).
Identification of putative targets
regulated by miR-218 in EBC-1 cells
To identify putative targets of miR-218, we
employed a combination of in silico analysis, genome-wide
gene expression analysis and analysis of gene expression in
clinical specimens using the GEO database. First, we identified 513
genes downregulated by miR-218 transfection into EBC-1 cells
(log2 ratio <−1.0). Among these 513 genes, 163 genes
have putative miR-218 binding sites in their 3′-UTR regions
according to the TargetScan database. Next, we cross-referenced
these genes with genes upregulated (fold change >1.5) in NSCLC
clinical specimens from the GEO gene expression data set (accession
number: GSE19188). A total of 36 candidate genes were identified as
putative target genes of miR-218 regulation in lung SCC
(Table II). Fig. 2 shows our strategy for selecting
putative miR-218 target genes.
| Table IIDownregulated genes in miR-218
tranfectant. |
Table II
Downregulated genes in miR-218
tranfectant.
| Entrez gene ID | Gene symbol | Description | miR-218
target site | EBC-1
miR-218 transfectant (log2 ratio) | GSE: 19188 (fold
change) |
|---|
|
|---|
| Total | Conserved site | Poorly conserved
site |
|---|
| 7163 | TPD52 | Tumor protein
D52 | 4 | 2 | 2 | −1.33 | 2.66 |
| 5563 | PRKAA2 | Protein kinase,
AMP-activated, α 2 catalytic subunit | 4 | 0 | 4 | −1.04 | 2.50 |
| 2673 | GFPT1 |
Glutamine-fructose-6-phosphate
transaminase 1 | 4 | 1 | 3 | −1.17 | 1.92 |
| 332 | BIRC5 | Baculoviral IAP
repeat containing 5 | 2 | 0 | 2 | −2.05 | 12.42 |
| 56938 | ARNTL2 | Aryl hydrocarbon
receptor nuclear translocator-like 2 | 2 | 0 | 2 | −1.09 | 7.85 |
| 112399 | EGLN3 | Egl-9 family
hypoxia-inducible factor 3 | 2 | 1 | 1 | −1.45 | 5.97 |
| 84181 | CHD6 | chromodomain
helicase DNA binding protein 6 | 2 | 1 | 1 | −1.35 | 2.30 |
| 29927 | SEC61A1 | Sec61 α 1 subunit
(S. cerevisiae) | 2 | 1 | 1 | −1.09 | 2.13 |
| 8776 | MTMR1 | Myotubularin
related protein 1 | 2 | 1 | 1 | −1.06 | 1.96 |
| 131566 | DCBLD2 | Discoidin, CUB and
LCCL domain containing 2 | 2 | 2 | 0 | −1.46 | 1.88 |
| 2820 | GPD2 |
Glycerol-3-phosphate dehydrogenase 2
(mitochondrial) | 2 | 0 | 2 | −1.02 | 1.83 |
| 399694 | SHC4 | SHC (Src homology 2
domain containing) family, member 4 | 2 | 1 | 1 | −1.25 | 1.58 |
| 3486 | IGFBP3 | Insulin-like growth
factor binding protein 3 | 1 | 0 | 1 | −2.09 | 4.12 |
| 144406 | WDR66 | WD repeat domain
66 | 1 | 0 | 1 | −1.11 | 2.98 |
| 25907 | TMEM158 | Transmembrane
protein 158 (gene/pseudogene) | 1 | 0 | 1 | −1.12 | 2.96 |
| 10447 | FAM3C | Family with
sequence similarity 3, member C | 1 | 1 | 0 | −2.08 | 2.90 |
| 4017 | LOXL2 | Lysyl oxidase-like
2 | 1 | 0 | 1 | −1.96 | 2.56 |
| 9139 | CBFA2T2 | Core-binding
factor, runt domain, α subunit 2; translocated to, 2 | 1 | 0 | 1 | −1.11 | 2.42 |
| 3691 | ITGB4 | Integrin, β 4 | 1 | 0 | 1 | −1.12 | 2.20 |
| 6745 | SSR1 | Signal sequence
receptor, α | 1 | 1 | 0 | −1.47 | 1.92 |
| 1457 | CSNK2A1 | Casein kinase 2, α
1 polypeptide | 1 | 1 | 0 | −1.30 | 1.90 |
| 157638 | FAM84B | Family with
sequence similarity 84, member B | 1 | 0 | 1 | −1.02 | 1.90 |
| 949 | SCARB1 | Scavenger receptor
class B, member 1 | 1 | 0 | 1 | −1.54 | 1.84 |
| 2011 | MARK2 | MAP/microtubule
affinity-regulating kinase 2 | 1 | 1 | 0 | −1.41 | 1.75 |
| 9289 | GPR56 | G protein-coupled
receptor 56 | 1 | 0 | 1 | −1.53 | 1.70 |
| 55959 | SULF2 | Sulfatase 2 | 1 | 0 | 1 | −1.11 | 1.67 |
| 65985 | AACS | Acetoacetyl-CoA
synthetase | 1 | 0 | 1 | −1.10 | 1.63 |
| 160 | AP2A1 | Adaptor-related
protein complex 2, α 1 subunit | 1 | 1 | 0 | −1.14 | 1.63 |
| 54928 | IMPAD1 | Inositol
monophosphatase domain containing 1 | 1 | 1 | 0 | −1.01 | 1.62 |
| 1889 | ECE1 | Endothelin
converting enzyme 1 | 1 | 0 | 1 | −1.55 | 1.61 |
| 55609 | ZNF280C | Zinc finger protein
280C | 1 | 1 | 0 | −1.10 | 1.61 |
| 6533 | SLC6A6 | Solute carrier
family 6 (neurotransmitter transporter), member 6 | 1 | 1 | 0 | −1.13 | 1.61 |
| 80000 | GREB1L | Growth regulation
by estrogen in breast cancer-like | 1 | 0 | 1 | −1.15 | 1.59 |
| 10776 | ARPP19 | cAMP-regulated
phosphoprotein, 19 kDa | 1 | 1 | 0 | −1.33 | 1.52 |
| 83637 | ZMIZ2 | Zinc finger,
MIZ-type containing 2 | 1 | 1 | 0 | −1.59 | 1.52 |
| 79139 | DERL1 | Derlin 1 | 1 | 0 | 1 | −1.16 | 1.51 |
In the present study, we focused on the TPD52
gene, investigating TPD52 function and downregulated
TPD52-mediated pathways.
TPD52 is directly targeted by miR-218 in
lung SCC cells
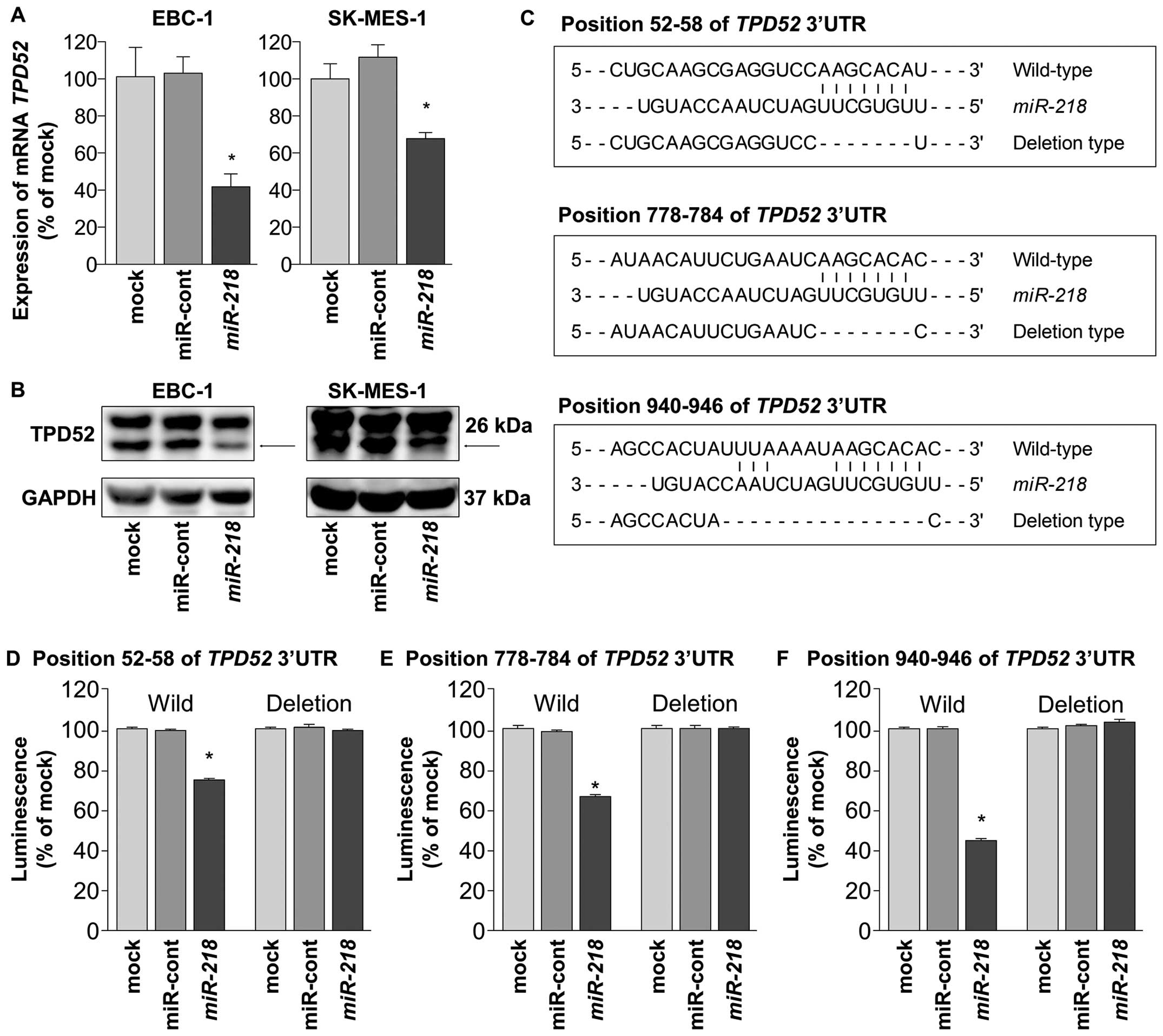
First, we measured the expression of
TPD52/TPD52 using quantitative RT-PCR and western blotting
to determine whether restoration of miR-218 in EBC-1 and
SK-MES-1 cells suppressed the expression of TPD52/TPD52. As
shown in Fig. 3A and B, the
expression levels of TPD52/TPD52 were markedly repressed by
miR-218 transfection compared with mock- or
miR-control-transfected cells.
Next, we used luciferase reporter assays to
determine whether miR-218 directly binds to the 3′-UTR of
TPD52 mRNA. We used vectors encoding either partial
wild-type sequences of the 3′-UTR of TPD52 including the
predicted miR-218 target sites (positions 52–58, 778–784,
940–946 of the TPD52 3′-UTR), or deletion vectors lacking
these sites (Fig. 3C). The
luminescence intensities were significantly reduced by transfection
with miR-218 and the vector carrying the wild-type 3′-UTR of
TPD52, whereas transfection with the deletion vector blocked
the decrease of luminescence in EBC-1 cells (P<0.0001). Thus,
miR-218 bound directly to three sites in the 3′-UTR of
TPD52 (Fig. 3D–F).
Effects of TPD52 silencing on cell
proliferation, migration and invasion in lung SCC cell lines
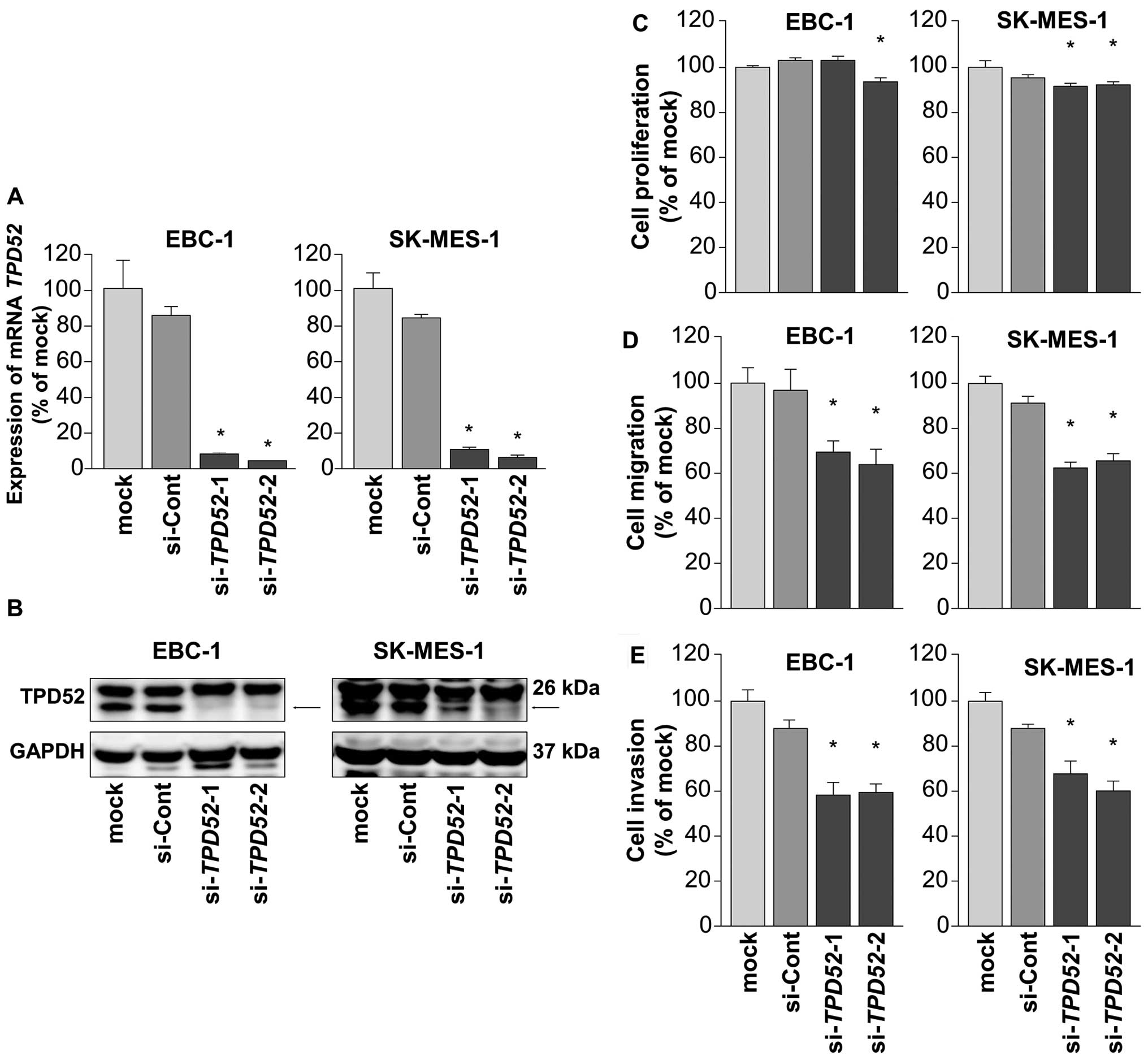
We performed loss-of-function studies using
si-TPD52 to investigate the oncogenic function of
TPD52 in lung SCC cells. First, we measured the knockdown
efficiency of si-TPD52 transfection in EBC-1 and SK-MES-1
cells. RT-PCR and western blotting showed that two siRNAs
(si-TPD52-1 and si-TPD52-2) could effectively reduce
the expression of TPD52/TPD52 in EBC-1 and SK-MES-1 cells
(Fig. 4A and B).
Next, we carried out functional assays using these
two siRNAs. XTT assays demonstrated that cell proliferation was
inhibited by si-TPD52 transfection only in SK-MES-1 cells
(Fig. 4C). Cell migration and
invasion activities were significantly inhibited by si-TPD52
transfection in both EBC-1 and SK-MES-1 cells in comparison with
mock- or negative-control transfectants (Fig. 4D and E).
TPD52 was strongly expressed in clinical
lung SCC specimens
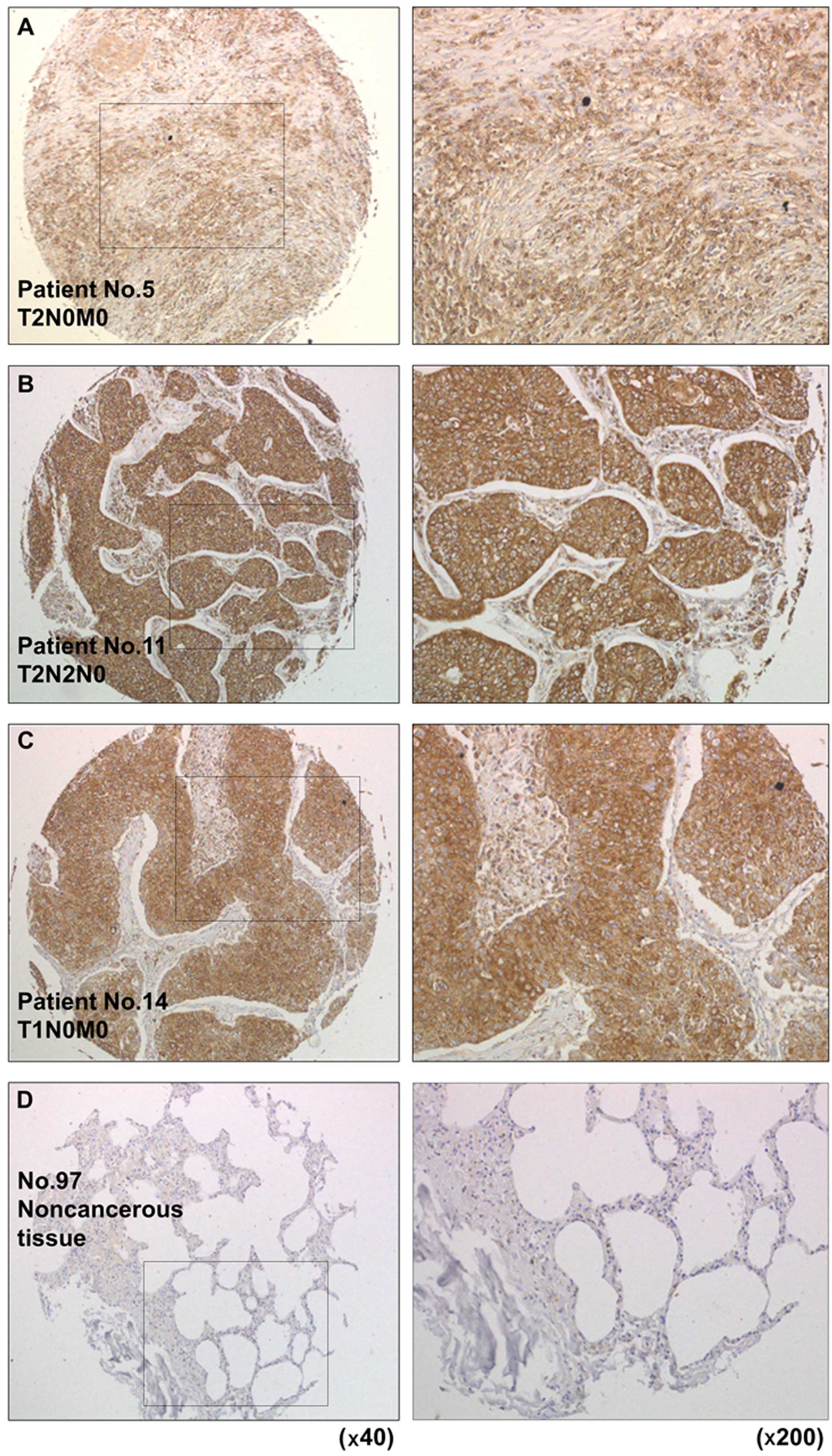
To analyze whether TPD52 was upregulated in
lung SCC clinical specimens, we carried out immunohistochemical
staining of lung SCC and non-cancerous lung tissues. Thirty
specimens were stained in this study (20 lung SCC specimens and 10
non-cancerous tissues); all lung SCC specimens stained moderately
or strongly, whereas all non-cancerous specimens stained weakly or
negatively for TPD52 (Table III
and Fig. 5).
| Table IIIImmunohistochemistry status and
characteristics of the lung cancer and non-cancerous cases. |
Table III
Immunohistochemistry status and
characteristics of the lung cancer and non-cancerous cases.
| A,
Immunohistochemistry status and characteristics of the lung
squamous cell carcinoma cases |
|---|
|
|---|
| Patient no. | Grade | T | N | M | Pathological
stage |
Immunohistochemistry |
|---|
| 1 | 1 | 3 | 1 | 0 | IIIa | (++) |
| 2 | 1 | 3 | 0 | 0 | IIIa | (++) |
| 3 | 2 | 2 | 1 | 0 | II | (++) |
| 4 | 2 | 3 | 0 | 0 | IIIa | (++) |
| 5 | 1 | 2 | 0 | 0 | I | (++) |
| 6 | 1 | 2 | 1 | 0 | II | (+) |
| 7 | 1 | 3 | 1 | 0 | IIIa | (+++) |
| 8 | 1 | 2 | 0 | 0 | I | (++) |
| 9 | 1 | 2 | 1 | 0 | II | (++) |
| 10 | 1 | 2 | 0 | 0 | I | (++) |
| 11 | 2 | 2 | 2 | 0 | IIIa | (+++) |
| 12 | 2 | 2 | 0 | 0 | I | (+) |
| 13 | 2 | 1 | 0 | 0 | I | (++) |
| 14 | 2 | 1 | 0 | 0 | I | (++) |
| 15 | 2 | 2 | 1 | 0 | II | (+) |
| 16 | 2 | 3 | 1 | 0 | III | (++) |
| 17 | 2 | 2 | 0 | 0 | I | (++) |
| 18 | 2 | 2 | 1 | 0 | II | (+) |
| 19 | 2 | 3 | 2 | 0 | IIIa | (+++) |
| 20 | 2 | 2 | 0 | 0 | I | (++) |
|
| B,
Immunohistochemistry status of non-cancerous cases |
|
| Patient no. |
Immunohistochemistry |
|
| 91 | (−) |
| 92 | (+) |
| 93 | (+) |
| 94 | (−) |
| 95 | (−) |
| 96 | (−) |
| 97 | (−) |
| 98 | (+) |
| 99 | (+) |
| 100 | (+) |
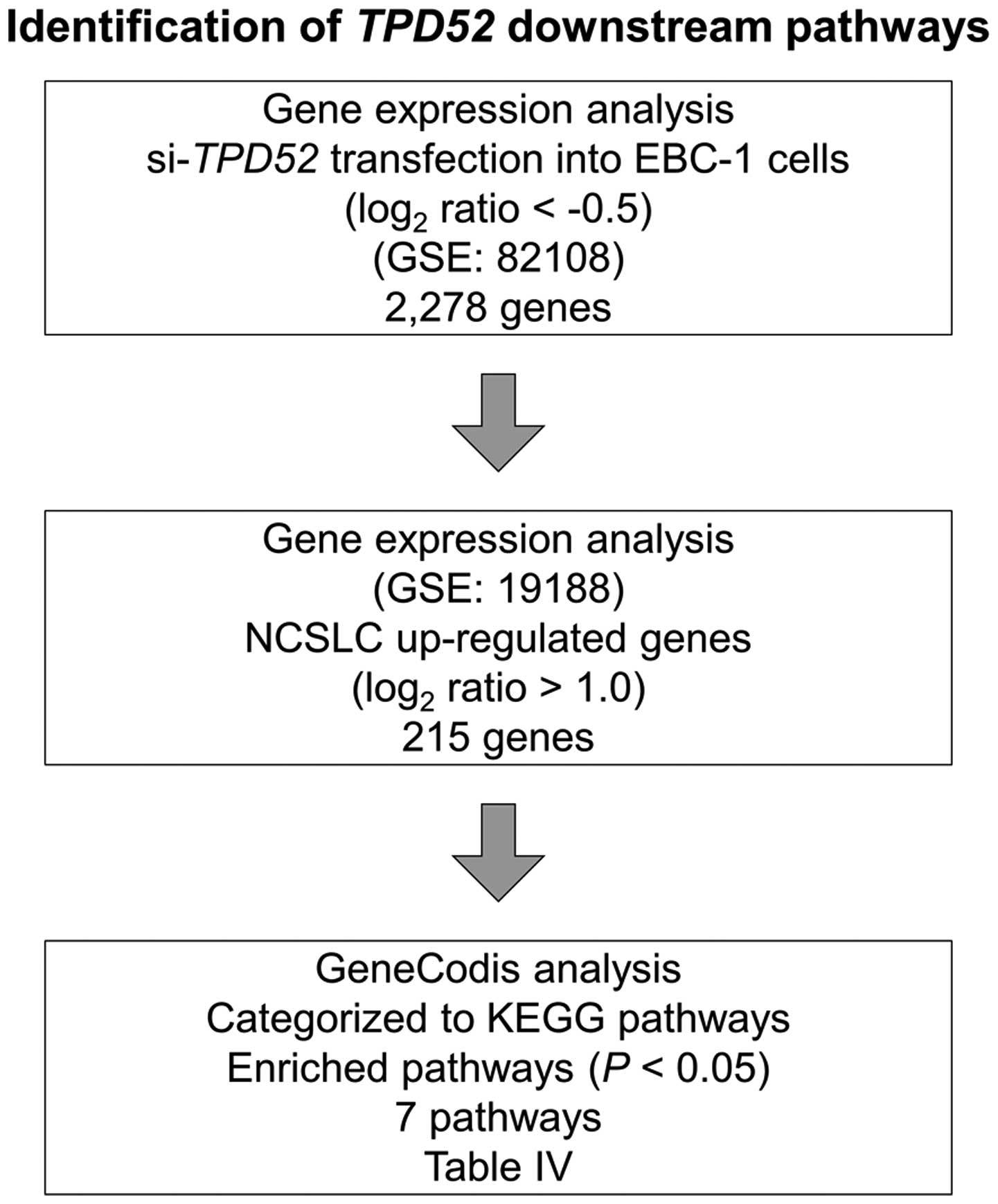
Identification of TPD52-mediated
downstream pathways in lung SCC cells
To identify the downstream genes regulated by
TPD52, genome-wide gene expression analysis and in
silico analysis were performed in lung SCC cells transfected
with si-TPD52. A total of 2,278 genes were identified as
downregulated in si-TPD52-transfected EBC-1 cells compared
with the control (log2 ratio <−0.5). Among them, 215
genes were upregulated in NSCLC specimens in the GEO database
(accession number: GSE19188). We categorized the 215 genes
according to KEGG pathways, and 7 pathways were identified as
significantly enriched pathways (Table IV). Fig. 6 shows our strategy for selecting
TPD52-mediated downstream pathways. Among these pathways, we
focused on the ‘Cell cycle pathway’, ‘DNA replication pathways’ and
‘p53 signaling pathway’. Genes involved in these pathways are
listed in Table V.
| Table IVSignificantly enriched annotations
regulated by si-TPD52-1 and si-TPD52-2 in lung SCC
cells. |
Table IV
Significantly enriched annotations
regulated by si-TPD52-1 and si-TPD52-2 in lung SCC
cells.
| No. of genes | P-value | Annotations |
|---|
| 12 | 2.06E-11 | (KEGG) 04110: Cell
cycle |
| 8 | 8.84E-07 | (KEGG) 04114:
Oocyte meiosis |
| 7 | 2.27E-06 | (KEGG) 05322:
Systemic lupus erythematosus |
| 5 | 5.41E-06 | (KEGG) 03030: DNA
replication |
| 6 | 2.30E-05 | (KEGG) 04914:
Progesterone-mediated oocyte maturation |
| 5 | 9.53E-05 | (KEGG) 04115: p53
signaling pathway |
| 2 | 4.91E-02 | (KEGG) 03410: Base
excision repair |
| Table VRepresentative pathways identified as
downstream of TPD52. |
Table V
Representative pathways identified as
downstream of TPD52.
| Cell cycle
pathway |
|---|
|
|---|
| Gene symbol | Description | GSE:19188
(log2 ratio) |
|---|
| TTK | TTK protein
kinase | 3.23 |
| BUB1 | Budding uninhibited
by benzimidazoles 1 homolog (yeast) | 3.21 |
| CCNB2 | Cyclin B2 | 3.20 |
| MCM4 | Minichromosome
maintenance complex component 4 | 3.13 |
| CCNB1 | Cyclin B1 | 2.82 |
| PLK1 | Polo-like kinase
1 | 2.81 |
| CDC25C | Cell division cycle
25 homolog C (S. pombe) | 2.77 |
| CDK1 | Cyclin-dependent
kinase 1 | 2.40 |
| CCNE2 | Cyclin E2 | 2.04 |
| DBF4 | DBF4 homolog (S.
cerevisiae) | 1.42 |
| MCM6 | Minichromosome
maintenance complex component 6 | 1.31 |
| BUB3 | Budding uninhibited
by benzimidazoles 3 homolog (yeast) | 1.07 |
|
| DNA replication
pathway |
|
| Gene symbol | Description | GSE:19188
(log2 ratio) |
|
| MCM4 | Minichromosome
maintenance complex component 4 | 3.13 |
| FEN1 | Flap
structure-specific endonuclease 1 | 1.66 |
| MCM6 | Minichromosome
maintenance complex component 6 | 1.31 |
| PRIM1 | Primase, DNA,
polypeptide 1 (49 kDa) | 1.25 |
| POLA2 | Polymerase (DNA
directed), α 2 (70 kD subunit) | 1.00 |
|
| p53 signaling
pathway |
|
| Gene symbol | Description | GSE:19188
(log2 ratio) |
|
| CCNB2 | Cyclin B2 | 3.20 |
| CCNB1 | Cyclin B1 | 2.82 |
| GTSE1 | G-2 and S-phase
expressed 1 | 2.79 |
| CDK1 | Cyclin-dependent
kinase 1 | 2.40 |
| CCNE2 | Cyclin E2 | 2.04 |
Discussion
A single miRNA can regulate an extremely large
number of protein-coding or non-coding RNAs. Thus, aberrant
expression of miRNAs disrupts the RNA network in cancer cells.
Identification of aberrantly expressed miRNAs and the novel cancer
networks they regulate is a research trend of the
post-genome-sequencing era. Recent evidence shows that disruption
of normally-regulated RNA networks by aberrantly expressed miRNAs
triggers cancer cell development, progression and metastasis
(3,4). Our recent studies of miRNA expression
signatures in various cancers indicated that miR-218 is
frequently downregulated in cancers (9–11).
Investigating miR-218-regulated RNA networks in lung SCC is
the first step in construction of a new treatment strategy for the
disease.
Our current data showed that expression of
miR218 was significantly reduced in lung SCC specimens. The
mature form of miR-218 is generated from two separate loci,
miR-218-1 and miR-218-2, which are located on
chromosomes 4p15.31 and 5q35.1 within the introns of SLIT2
and SLIT3, respectively (22). Several lines of evidence suggest
that miR-218 is frequently downregulated in various cancers
(17,18,23,24).
In this study, ectopic expression of miR-218 significantly
inhibited cancer cell migration and invasion, suggesting this miRNA
suppresses metastasis-promoting genes. Our previous studies showed
that antitumor effects of miR-218 explicitly contribute to
migration and invasion in head and neck cancer, cervical cancer,
renal cell carcinoma and prostate cancer (17,18,23,24).
Interestingly, miR-218-regulated genes were involved in
ECM-related components such as collagens, laminins and integrins
(17,18,23,24).
Accumulating evidence indicates that the interaction of cancer
cells with their microenvironment influences the initiation,
development and metastasis of cancer cells (25,26).
Overexpression of ECM components and activation of ECM-integrin
signaling were observed in several types of cancers and shown to
activate cancer cell aggressiveness (27). The discovery of tumor-suppressive
miR-218-regulated genes and pathways may provide important
insights into the potential mechanisms of lung SCC metastasis.
To better understand lung SCC development and
metastasis, we identified miR-218 target genes using a
combination of in silico and genome-wide gene expression
analyses. We have identified antitumor miRNA-regulated oncogenes
and novel cancer networks using this strategy (9–11).
In the present study, a total of 36 putative target genes of
miR-218 were identified. Among these genes, we demonstrated
that LOXL2 (lysyl oxidase homolog 2) was directly regulated
by miR-218 in head and neck cancer and prostate cancer
(28). Another group showed direct
regulation of BIRC5 (baculoviral IAP repeat containing 5) by
miR-218 in cervical cancer (29). These findings show the
effectiveness of our strategy in identifying target genes of
miR-218-regulation in cancer cells.
In the present study, we focused on TPD52 and
investigated the functional significance of this gene in lung SCC.
TPD52 was initially cloned through differential screening
using a breast cancer cDNA library (30). This cDNA clone had novel sequences
and was expressed in breast and basal cell carcinomas (30). Interestingly, this gene is located
on the human chromosome 8q21.13, a frequently observed region of
chromosomal amplification in several types of cancers (31–33).
In lung adenocarcinoma, chromosome 8q21.13, which includes
TPD52, was one of the most notable amplified genomic regions
(34). A vast number of studies
showed that TPD52 is overexpressed both at the mRNA and
protein levels in several cancers. Likewise, overexpression of
TPD52 was reported in small cell lung cancer, lung
adenocarcinoma and lung SCC (35–37).
The influence of smoking is important in lung cancer pathogenesis.
Expression levels of TPD52 were enhanced in airway
epithelial cells in smokers with lung cancer compared to smokers
without cancer (38). The
functional roles of TPD52 were investigated using
overexpression or knockdown analytical methods. Ectopic expression
of mouse TPD52 in 3T3 fibroblasts resulted in a transformed
phenotype that progressed to metastasis (39). Several studies have shown that
increased expression of TPD52 enhances the proliferation of
prostate cancer cell lines under both normal and androgen-resistant
conditions (40,41). Other studies showed that
TPD52 knockdown increases apoptotic cell death in
ERBB2-amplified breast cancer cell lines (33). These findings suggest that
over-expression of TPD52 enhances cancer cell aggressiveness
and contributes to several oncogenic pathways.
Recently, several studies demonstrated that
TPD52 expression is regulated by several miRNAs in cancer
cells. Downregulation of TPD52 expression was observed by
miR-107 and miR-185 transfection into non-small cell
lung cancer cells and miR-34a transfection into colorectal
cancer cells (42,43). Our previous data showed that the
tumor-suppressive miR-224 directly regulates oncogenic
TPD52 in prostate cancer cells, and silencing TPD52
results in significant reductions in cancer cell migration and
invasion in prostate cancer cells (44). More recently, miR-218 was
shown to directly regulate TPD52 in prostate cancer cells
(45). These data agree with our
present report in lung SCC and support our conclusions. Control of
TPD52 expression by miRNAs is a novel molecular mechanism of
cancer cells, and further investigation of the miRNA-TPD52 axis is
needed.
In the present study, we identified
TPD52-mediated cancer pathways using genome-wide gene
expression analysis of si-TPD52-transfected lung SCC cells.
Our data showed that several pathways were identified downstream of
TPD52 pathways, such as the ‘Cell cycle pathway’, ‘DNA
replication pathway’ and ‘p53 signaling pathway’. The genes
involved in these pathways were critical regulators of genomic
stability and were mitotic checkpoint genes (46,47).
The functional insights obtained in the current studies indicated
that several cell cycle kinases, such as TTK, BUB1 and PLK1, were
multi-functional enzymes and contributed to cancer cell migration,
invasion and metastasis (48–50).
The protein kinase TTK promoted the cell proliferation and
migration through activation of AKT-mTOR and MDM2-p53 signaling
pathways in hepatocellular carcinoma cells (48). Using the siRNA screen of the human
kinome revealed that a serine/threonine kinase BUB1 acted as an
essential mediator of TGFβ-dependent signaling (49). This study demonstrated that BUB1
interacted with both, TGFBRI and TGFBRII and promoted the
TGFβ-dependent epithelial mesenchymal transition (EMT), cell
migration and invasion (49). PLK1
is a serine/threonine kinase and a pivotal player of cell cycle
regulator (50). Overexpression of
PLK1 was observed in prostate cancer and involved in enhancing EMT
and stimulation of cell migration and invasion (50). In bladder cancer, CCNB2 was
overexpressed in cancer cells and knockdown of CCNB2
inhibited invasion and metastatic abilities (51). These findings have supported our
present data of knockdown of TPD52 in lung cancer cells.
Exploration of novel TPD52-mediated pathways may lead to the
development of new treatment protocols for this disease.
In conclusion, expression of miR-218 was
frequently downregulated in lung SCC clinical specimens and
appeared to function in anti-migration and anti-invasion roles
through targeting of TPD52. Elucidation of
miR-218-regulated cancer networks should provide new
information on potential therapeutic targets in the treatment of
lung SCC development and metastasis.
Acknowledgements
We thank Dr Masami Sato (Department of General
Thoracic Surgery, Graduate School of Medical and Dental Sciences,
Kagoshima University, Kagoshima, Japan) for kindly providing
surgically resected samples. The present study was supported in
part by the Grants-in-Aid for Scientific Research from the Japan
Society for the Promotion of Science (JSPS, KAKENHI grants 15K09179
and 16K19458).
References
|
1
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hobert O: Gene regulation by transcription
factors and microRNAs. Science. 319:1785–1786. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Iorio MV and Croce CM: MicroRNAs in
cancer: Small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rolfo C, Fanale D, Hong DS, Tsimberidou
AM, Piha-Paul SA, Pauwels P, Van Meerbeeck JP, Caruso S, Bazan V,
Cicero G, et al: Impact of microRNAs in resistance to chemotherapy
and novel targeted agents in non-small cell lung cancer. Curr Pharm
Biotechnol. 15:475–485. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yanaihara N, Caplen N, Bowman E, Seike M,
Kumamoto K, Yi M, Stephens RM, Okamoto A, Yokota J, Tanaka T, et
al: Unique microRNA molecular profiles in lung cancer diagnosis and
prognosis. Cancer Cell. 9:189–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Raponi M, Dossey L, Jatkoe T, Wu X, Chen
G, Fan H and Beer DG: MicroRNA classifiers for predicting prognosis
of squamous cell lung cancer. Cancer Res. 69:5776–5783. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Moriya Y, Nohata N, Kinoshita T, Mutallip
M, Okamoto T, Yoshida S, Suzuki M, Yoshino I and Seki N: Tumor
suppressive microRNA-133a regulates novel molecular networks in
lung squamous cell carcinoma. J Hum Genet. 57:38–45. 2012.
View Article : Google Scholar
|
|
9
|
Mataki H, Enokida H, Chiyomaru T, Mizuno
K, Matsushita R, Goto Y, Nishikawa R, Higashimoto I, Samukawa T,
Nakagawa M, et al: Downregulation of the microRNA-1/133a cluster
enhances cancer cell migration and invasion in lung-squamous cell
carcinoma via regulation of Coronin1C. J Hum Genet. 60:53–61. 2015.
View Article : Google Scholar
|
|
10
|
Mataki H, Seki N, Chiyomaru T, Enokida H,
Goto Y, Kumamoto T, Machida K, Mizuno K, Nakagawa M and Inoue H:
Tumor-suppressive microRNA-206 as a dual inhibitor of MET and EGFR
oncogenic signaling in lung squamous cell carcinoma. Int J Oncol.
46:1039–1050. 2015.
|
|
11
|
Mizuno K, Seki N, Mataki H, Matsushita R,
Kamikawaji K, Kumamoto T, Takagi K, Goto Y, Nishikawa R, Kato M, et
al: Tumor-suppressive microRNA-29 family inhibits cancer cell
migration and invasion directly targeting LOXL2 in lung squamous
cell carcinoma. Int J Oncol. 48:450–460. 2016.
|
|
12
|
Kikkawa N, Hanazawa T, Fujimura L, Nohata
N, Suzuki H, Chazono H, Sakurai D, Horiguchi S, Okamoto Y and Seki
N: miR-489 is a tumour-suppressive miRNA target PTPN11 in
hypopharyngeal squamous cell carcinoma (HSCC). Br J Cancer.
103:877–884. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hidaka H, Seki N, Yoshino H, Yamasaki T,
Yamada Y, Nohata N, Fuse M, Nakagawa M and Enokida H: Tumor
suppressive microRNA-1285 regulates novel molecular targets:
Aberrant expression and functional significance in renal cell
carcinoma. Oncotarget. 3:44–57. 2012.PubMed/NCBI
|
|
14
|
Itesako T, Seki N, Yoshino H, Chiyomaru T,
Yamasaki T, Hidaka H, Yonezawa T, Nohata N, Kinoshita T, Nakagawa
M, et al: The microRNA expression signature of bladder cancer by
deep sequencing: The functional significance of the miR-195/497
cluster. PLoS One. 9:e843112014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kojima S, Enokida H, Yoshino H, Itesako T,
Chiyomaru T, Kinoshita T, Fuse M, Nishikawa R, Goto Y, Naya Y, et
al: The tumor-suppressive microRNA-143/145 cluster inhibits cell
migration and invasion by targeting GOLM1 in prostate cancer. J Hum
Genet. 59:78–87. 2014. View Article : Google Scholar
|
|
16
|
Tatarano S, Chiyomaru T, Kawakami K,
Enokida H, Yoshino H, Hidaka H, Yamasaki T, Kawahara K, Nishiyama
K, Seki N, et al: miR-218 on the genomic loss region of chromosome
4p15.31 functions as a tumor suppressor in bladder cancer. Int J
Oncol. 39:13–21. 2011.PubMed/NCBI
|
|
17
|
Kinoshita T, Hanazawa T, Nohata N, Kikkawa
N, Enokida H, Yoshino H, Yamasaki T, Hidaka H, Nakagawa M, Okamoto
Y, et al: Tumor suppressive microRNA-218 inhibits cancer cell
migration and invasion through targeting laminin-332 in head and
neck squamous cell carcinoma. Oncotarget. 3:1386–1400. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamamoto N, Kinoshita T, Nohata N, Itesako
T, Yoshino H, Enokida H, Nakagawa M, Shozu M and Seki N: Tumor
suppressive microRNA-218 inhibits cancer cell migration and
invasion by targeting focal adhesion pathways in cervical squamous
cell carcinoma. Int J Oncol. 42:1523–1532. 2013.PubMed/NCBI
|
|
19
|
Goldstraw P, Crowley J, Chansky K, Giroux
DJ, Groome PA, Rami-Porta R, Postmus PE, Rusch V and Sobin L;
International Association for the Study of Lung Cancer
International Staging Committee; Participating Institutions. The
IASLC Lung Cancer Staging Project: Proposals for the revision of
the TNM stage groupings in the forthcoming (seventh) edition of the
TNM Classification of malignant tumours. J Thorac Oncol. 2:706–714.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fukumoto I, Kinoshita T, Hanazawa T,
Kikkawa N, Chiyomaru T, Enokida H, Yamamoto N, Goto Y, Nishikawa R,
Nakagawa M, et al: Identification of tumour suppressive
microRNA-451a in hypopharyngeal squamous cell carcinoma based on
microRNA expression signature. Br J Cancer. 111:386–394. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Matsushita R, Seki N, Chiyomaru T,
Inoguchi S, Ishihara T, Goto Y, Nishikawa R, Mataki H, Tatarano S,
Itesako T, et al: Tumour-suppressive microRNA-144-5p directly
targets CCNE1/2 as potential prognostic markers in bladder cancer.
Br J Cancer. 113:282–289. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lu YF, Zhang L, Waye MM, Fu WM and Zhang
JF: MiR-218 mediates tumorigenesis and metastasis: Perspectives and
implications. Exp Cell Res. 334:173–182. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yamasaki T, Seki N, Yoshino H, Itesako T,
Hidaka H, Yamada Y, Tatarano S, Yonezawa T, Kinoshita T, Nakagawa
M, et al: MicroRNA-218 inhibits cell migration and invasion in
renal cell carcinoma through targeting caveolin-2 involved in focal
adhesion pathway. J Urol. 190:1059–1068. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nishikawa R, Goto Y, Sakamoto S, Chiyomaru
T, Enokida H, Kojima S, Kinoshita T, Yamamoto N, Nakagawa M, Naya
Y, et al: Tumor-suppressive microRNA-218 inhibits cancer cell
migration and invasion via targeting of LASP1 in prostate cancer.
Cancer Sci. 105:802–811. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Weaver VM, Petersen OW, Wang F, Larabell
CA, Briand P, Damsky C and Bissell MJ: Reversion of the malignant
phenotype of human breast cells in three-dimensional culture and in
vivo by integrin blocking antibodies. J Cell Biol. 137:231–245.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bissell MJ and Radisky D: Putting tumours
in context. Nat Rev Cancer. 1:46–54. 2001. View Article : Google Scholar
|
|
27
|
Hood JD and Cheresh DA: Role of integrins
in cell invasion and migration. Nat Rev Cancer. 2:91–100. 2002.
View Article : Google Scholar
|
|
28
|
Fukumoto I, Kikkawa N, Matsushita R, Kato
M, Kurozumi A, Nishikawa R, Goto Y, Koshizuka K, Hanazawa T,
Enokida H, et al: Tumor-suppressive microRNAs (miR-26a/b,
miR-29a/b/c and miR-218) concertedly suppressed
metastasis-promoting LOXL2 in head and neck squamous cell
carcinoma. J Hum Genet. 61:109–118. 2016. View Article : Google Scholar
|
|
29
|
Kogo R, How C, Chaudary N, Bruce J, Shi W,
Hill RP, Zahedi P, Yip KW and Liu FF: The microRNA-218~Survivin
axis regulates migration, invasion, and lymph node metastasis in
cervical cancer. Oncotarget. 6:1090–1100. 2015. View Article : Google Scholar :
|
|
30
|
Byrne JA, Tomasetto C, Garnier JM, Rouyer
N, Mattei MG, Bellocq JP, Rio MC and Basset P: A screening method
to identify genes commonly overexpressed in carcinomas and the
identification of a novel complementary DNA sequence. Cancer Res.
55:2896–2903. 1995.PubMed/NCBI
|
|
31
|
Byrne JA, Balleine RL, Schoenberg Fejzo M,
Mercieca J, Chiew YE, Livnat Y, St Heaps L, Peters GB, Byth K,
Karlan BY, et al: Tumor protein D52 (TPD52) is overexpressed and a
gene amplification target in ovarian cancer. Int J Cancer.
117:1049–1054. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rubin MA, Varambally S, Beroukhim R,
Tomlins SA, Rhodes DR, Paris PL, Hofer MD, Storz-Schweizer M,
Kuefer R, Fletcher JA, et al: Overexpression, amplification, and
androgen regulation of TPD52 in prostate cancer. Cancer Res.
64:3814–3822. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Roslan N, Bièche I, Bright RK, Lidereau R,
Chen Y and Byrne JA: TPD52 represents a survival factor in
ERBB2-amplified breast cancer cells. Mol Carcinog. 53:807–819.
2014. View Article : Google Scholar
|
|
34
|
Weir BA, Woo MS, Getz G, Perner S, Ding L,
Beroukhim R, Lin WM, Province MA, Kraja A, Johnson LA, et al:
Characterizing the cancer genome in lung adenocarcinoma. Nature.
450:893–898. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rohrbeck A, Neukirchen J, Rosskopf M,
Pardillos GG, Geddert H, Schwalen A, Gabbert HE, von Haeseler A,
Pitschke G, Schott M, et al: Gene expression profiling for
molecular distinction and characterization of laser captured
primary lung cancers. J Transl Med. 6:692008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhu H, Lam DC, Han KC, Tin VP, Suen WS,
Wang E, Lam WK, Cai WW, Chung LP and Wong MP: High resolution
analysis of genomic aberrations by metaphase and array comparative
genomic hybridization identifies candidate tumour genes in lung
cancer cell lines. Cancer Lett. 245:303–314. 2007. View Article : Google Scholar
|
|
37
|
Hanada S, Kakehashi A, Nishiyama N, Wei M,
Yamano S, Chung K, Komatsu H, Inoue H, Suehiro S and Wanibuchi H:
Myristoylated alanine-rich C-kinase substrate as a prognostic
biomarker in human primary lung squamous cell carcinoma. Cancer
Biomark. 13:289–298. 2013.PubMed/NCBI
|
|
38
|
Boelens MC, van den Berg A, Fehrmann RS,
Geerlings M, de Jong WK, te Meerman GJ, Sietsma H, Timens W, Postma
DS and Groen HJ: Current smoking-specific gene expression signature
in normal bronchial epithelium is enhanced in squamous cell lung
cancer. J Pathol. 218:182–191. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lewis JD, Payton LA, Whitford JG, Byrne
JA, Smith DI, Yang L and Bright RK: Induction of tumorigenesis and
metastasis by the murine orthologue of tumor protein D52. Mol
Cancer Res. 5:133–144. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ummanni R, Teller S, Junker H, Zimmermann
U, Venz S, Scharf C, Giebel J and Walther R: Altered expression of
tumor protein D52 regulates apoptosis and migration of prostate
cancer cells. FEBS J. 275:5703–5713. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li L, Xie H, Liang L, Gao Y, Zhang D, Fang
L, Lee SO, Luo J, Chen X, Wang X, et al: Increased PrLZ-mediated
androgen receptor transactivation promotes prostate cancer growth
at castration-resistant stage. Carcinogenesis. 34:257–267. 2013.
View Article : Google Scholar :
|
|
42
|
Takahashi Y, Forrest AR, Maeno E,
Hashimoto T, Daub CO and Yasuda J: MiR-107 and MiR-185 can induce
cell cycle arrest in human non small cell lung cancer cell lines.
PLoS One. 4:e66772009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kaller M, Liffers ST, Oeljeklaus S,
Kuhlmann K, Röh S, Hoffmann R, Warscheid B and Hermeking H:
Genome-wide characterization of miR-34a induced changes in protein
and mRNA expression by a combined pulsed SILAC and microarray
analysis. Mol Cell Proteomics. 10:M111 010462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Goto Y, Nishikawa R, Kojima S, Chiyomaru
T, Enokida H, Inoguchi S, Kinoshita T, Fuse M, Sakamoto S, Nakagawa
M, et al: Tumour-suppressive microRNA-224 inhibits cancer cell
migration and invasion via targeting oncogenic TPD52 in prostate
cancer. FEBS Lett. 588:1973–1982. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Han G, Fan M and Zhang X: microRNA-218
inhibits prostate cancer cell growth and promotes apoptosis by
repressing TPD52 expression. Biochem Biophys Res Commun.
456:804–809. 2015. View Article : Google Scholar
|
|
46
|
Aarts M, Linardopoulos S and Turner NC:
Tumour selective targeting of cell cycle kinases for cancer
treatment. Curr Opin Pharmacol. 13:529–535. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dominguez-Brauer C, Thu KL, Mason JM,
Blaser H, Bray MR and Mak TW: Targeting mitosis in cancer: Emerging
strategies. Mol Cell. 60:524–536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu X, Liao W, Yuan Q, Ou Y and Huang J:
TTK activates Akt and promotes proliferation and migration of
hepatocellular carcinoma cells. Oncotarget. 6:34309–34320.
2015.PubMed/NCBI
|
|
49
|
Nyati S, Schinske-Sebolt K, Pitchiaya S,
Chekhovskiy K, Chator A, Chaudhry N, Dosch J, Van Dort ME,
Varambally S, Kumar-Sinha C, et al: The kinase activity of the
Ser/Thr kinase BUB1 promotes TGF-β signaling. Sci Signal.
8:ra12015. View Article : Google Scholar
|
|
50
|
Wu J, Ivanov AI, Fisher PB and Fu Z:
Polo-like kinase 1 induces epithelial-to-mesenchymal transition and
promotes epithelial cell motility by activating CRAF/ERK signaling.
eLife. 5:52016. View Article : Google Scholar
|
|
51
|
Lei CY, Wang W, Zhu YT, Fang WY and Tan
WL: The decrease of cyclin B2 expression inhibits invasion and
metastasis of bladder cancer. Urol Oncol. 34:237.e1–237.e10. 2016.
View Article : Google Scholar
|