Introduction
Testicular germ cell tumor (TGCT) is the most
frequent solid malignancy occurring in males between the ages of 15
and 34 years (1), with a steadily
rising incidence for the past few decades in the United States and
Europe (2). Histopathologically,
~55% of all TGCTs are classified as seminomas, and the remaining
cases as non-seminomas (3). The
vast majority of TGCTs have an excellent cure rate with
cisplatin-based treatment. Nevertheless, a subset of patients
develops cisplatin resistance resulting in tumor progression and
reduced survival (4). Therefore, a
better understanding of the molecular mechanisms of TGCT
tumorigenesis is needed for identification of new therapeutic
targets and treatment development.
MicroRNAs (miRNAs) are small non-coding RNAs of
~20–24 nucleotides in length, which play important roles in a broad
range of cellular processes, including tumor development and drug
response (5). Genome-wide miRNA
profiling studies have provided evidence of miRNA deregulations in
TGCT. For example, the miR-371-373 cluster is frequently
overexpressed in malignant TGCTs of all histopathological subtypes
(6,7). Other miRNAs, such as the
miR-302 cluster and miR-301, are differentially
expressed based on the cellular differentiation of the tumor
(7,8). To date, very few miRNAs have been
functionally characterized in TGCT. miR-372 and
miR-373 have been shown to play oncogenic roles in TGCT by
targeting the tumor suppressor LATS2 (9). However, the functional roles of other
differentially expressed miRNAs in TGCT have yet to be
characterized.
We previously identified a subset of miRNAs that
were differentially expressed between TGCTs and normal testes (NT)
using a deep sequencing approach (10). Among these, miR-223-3p
expression was higher in TGCTs as compared to NT. miR-223-3p
is known to be deregulated in a broad range of hematological
malignancies and solid tumors (11,12).
However, its role in TGCT remains uncharacterized.
miR-223-3p has been shown to regulate multiple targets in
different cancer types. Among them, F-box/WD repeat-containing
protein (FBXW7) is the most common target, which has been reported
in acute T-cell lymphoblastic leukemia, esophageal squamous cell
carcinoma and gastric cancer (13–15).
FBXW7 is the substrate-recognition component of the SCF-(SKP1,
CUL1, F-box protein)-ubiquitin-ligase complex, which has been
demonstrated to function as a tumor suppressor by promoting the
degradation of several oncoprotein substrates, including c-Myc,
cyclin E, MCL-1, c-JUN, NFkB2 and Notch1 (16,17).
Therefore, suppression of FBXW7 by miR-223-3p can promote
tumor development and progression.
In this study, we investigated the expression and
function of miR-223-3p and FBXW7 in TGCT clinical samples
and cell lines. Our data show that miR-223-3p plays an
oncogenic role in TGCT by promoting cell proliferation and
inhibiting apoptosis via FBXW7.
Materials and methods
Clinical samples and cell lines
Fifteen frozen TGCTs and five NT were provided by
the Cooperative Human Tissue Network, which is funded by the
National Cancer Institute, USA. All samples were included in our
previous small RNA-sequencing study (10). The study was approved by the
Stanford Human Subjects Review Committee.
Two established TGCT cell lines were included in
this study: the TCam-2 seminoma cell line and the 2102Ep
non-seminoma cell line (18,19).
TCam-2 was kindly provided by Dr Leendert H.J. Looijenga
(Department of Pathology, Erasmus MC-University Medical Center
Rotterdam, The Netherlands) and 2102Ep by Dr Peter Andrews
(Department of Biomedical Science, University of Sheffield, UK).
TCam-2 cells were grown in RPMI-1640 and 2102Ep cells were cultured
in DMEM medium, supplemented with 10% fetal bovine serum. All cells
were cultured at 37°C with 5% CO2. Authentication of the
cell lines was verified by short tandem repeat profiling in our
recent study (10).
Data extraction and analysis from
published data and The Cancer Genome Atlas database
For comparison of miR-223-3p expression
between TGCTs and NT, we extracted global TaqMan miRNA profiling
data from the study of Gillis et al (7), which analyzed 61 germ cell tumors,
three NT and five embryonal carcinoma cell lines. We excluded the
10 dysgerminomas (ovarian germ cell tumors), one ovarian embryonal
carcinoma, one ovarian york sac carcinoma and five cell lines, and
re-analyzed the miR-223-3p expression by normalization to
miR-16 in the 49 TGCTs and three NT.
For FBXW7 mRNA, we extracted the microarray
gene expression data of 101 TCGTs and five NT from Gene Expression
Omnibus (GEO accession no. GSE3218; http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE3218).
For analysis of correlation between
miR-223-3p expression and FBXW7 mRNA levels, we
extracted miR-223-3p and FBXW7 mRNA data from The
Cancer Genome Atlas (TCGA) testicular cancer database using the
UCSC Xena browser (http://xena.ucsc.edu/). These miR-223-3p and
FBXW7 expression data had been generated by miRNA expression
Illumina HiSeq and exon expression RNAseq, respectively.
RNA extraction
Total RNA was extracted using the mirVana miRNA
isolation kit (AM1560; Ambion/Thermo Fisher Scientific, Waltham,
MA, USA) and RNA concentration was measured with a NanoDrop ND-1000
spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). All
RNA samples were stored at −80°C until further use.
TaqMan reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed to evaluate the transfection
efficiency of miR-223-3p overexpression or inhibition using
the StepOnePlus Real-Time PCR system (Applied Biosystems/Thermo
Fisher Scientific). cDNA was synthesized from 20 ng of total RNA
and used to quantify miR-223-3p (ID 002295) and RNU48
(ID 001093). All reactions were performed in triplicate. The
relative expression of miR-223-3p was normalized to
RNU48, and the fold change of miR-223-3p in cells
transfected with miR-223-3p mimic/inhibitor relative to
their respective control was reported as 2−ΔΔCt.
Transfection
For miR-223 overexpression and inhibition,
2×105 cells were transfected with 30 nM of miRNA
inhibitor (anti-miR-223, AM12301 or anti-miR negative control no.
1, AM17010; Ambion) or 10 nM of miRNA mimic (pre-miR-223, PM12301
or pre-miR negative control no. 1, AM17110; Ambion) using siPORT
NeoFX transfection agent (AM4511; Ambion).
For co-transfection of miR-223 mimic and
FBXW7-expressing plasmid, 1.5×105 cells were
co-transfected with 500 ng of pCMV-Myc FBXW7 and 10 nM of
pre-miR-223 or pre-miR-NC using Lipofectamine 2000 (no. 11668-019;
Invitrogen/Thermo Fisher Scientific). Cells co-transfected with an
empty vector and pre-miR-NC was used as a control. Cells were
collected 48 h after transfection for subsequent analysis. The
pCMV-Myc FBXW7 plasmid was obtained from Addgene (no. 16652;
Cambridge, MA, USA; https://www.addgene.org/). The empty vector was
prepared by cleavage of pCMV-Myc FBXW7 with BglII and
NotI to remove the full-length coding sequence of
FBXW7.
Annexin V cell apoptosis and EdU
(5-ethynyl-2′-deoxyuridine) cell proliferation assays
Cell apoptosis and proliferation were evaluated in
TCam-2 and 2102Ep cells 72 h after transfection using Annexin V
FITC Apoptosis kit (PHN1018; Invitrogen) and Click-iT EdU Alexa
Fluor 488 flow cytometry assay (C10425; Invitrogen), respectively.
All experimental conditions were according to the manufacturer's
instructions and analyzed by NovoCyte flow cytometer (ACEA
Biosciences, San Diego, CA, USA). At least three independent
experiments were performed in each cell line.
Trypan blue exclusion assay
Trypan blue exclusion assay was performed in TCam-2
and 2102Ep cells 48 or 72 h after transfection. Cells were stained
with 0.4% trypan blue solution and counted by TC10™ Automated Cell
Counter (Bio-Rad, Hercules, CA, USA).
WST-1 assay
Cell growth was measured by WST-1 colorimetric assay
(no. 11644807001; Roche Diagnostics, Indianapolis, IN, USA) in
TCam-2 and 2102Ep cells 72 h after transfection. Cells were plated
into a 96-well plate at a concentration of 5×103/well in
100 μl culture medium. At different time intervals (0, 24,
48 or 72 h after transfection), 10 μl of WST-1 reagent was
added to each well and incubated for 3 h at 37°C. After incubation,
absorbance values were detected at the wavelengths 450 nm
(measurement) and 650 nm (reference) using the VERSAmax ELISA
Microplate Reader (Molecular Devices, Sunnyvale, CA, USA). Each
experimental group was performed in six replicates for each
time-point and all experiments were repeated three times
independently.
Western blotting
Total protein lysates were extracted using NP-40
cell lysis buffer (FNN0021; Invitrogen), supplemented with 1 mM of
phenylmethanesulfonyl fluoride (P7626; Sigma-Aldrich, St. Louis,
MQ, USA) and protease inhibitor (P8340; Sigma-Aldrich). Protein
concentrations were determined using the Pierce™ BCA Protein assay
kit (no. 23227; Pierce Biotechnology, Thermo Fisher Scientific).
Thirty micrograms of protein lysates were separated in NuPAGE Novex
4–12% Bis-Tris gels (NP0321BOX; Invitrogen) and transferred to 0.2
μm nitrocellulose membranes (no. 88024; Invitrogen). After
blocking with 5% skim milk powder (no. 70166; Sigma-Aldrich) in
Tris-buffered saline/0.05% Tween-20, membranes were incubated with
anti-FBXW7 (NBP1-59631; Novus Biologicals, Littleton, CO, USA;
1:1,000 dilution), anti-cleaved PARP (ab32064; Abcam, Cambridge,
UK; 1:1,000 dilution) or anti-Myc-Tag (no. 2276; Cell Signaling
Technologies, Danvers, MA, USA; 1:500 dilution) overnight at 4°C.
Anti-rabbit IgG-HRP (no. 170-6515; Bio-Rad Laboratories; 1:3,000
dilution) or anti-mouse IgG-HRP (sc-2005; Santa Cruz Biotechnology,
Dallas, TX, USA, 1:10,000 dilution) was used as secondary
antibodies. For normalization purpose, the membrane was incubated
with anti-GAPDH (sc-47724; Santa Cruz Biotechnology; 1:1,000
dilution). Signals were detected using the Novex ECL HRP
chemiluminescent substrate reagent (WP20005; Invitrogen) and
LAS-1000 image analyzer (Fujifilm, Tokyo, Japan).
Statistical analyses
All statistical analyses were performed using MS
Office Excel 2007 or SPSS 22.0 (IBM Corp., Armonk, NY, USA).
Comparisons between TGCT and NT were performed by Mann-Whitney U
test, and the transfection experiments were assessed by Student's
paired t-test. Correlation between miR-223-3p and
FBXW7 mRNA expression levels was evaluated using Pearson's
correlation analysis. All statistical tests were two-sided and
P-values <0.05 were considered as statistically significant.
Results
Expression of miR-223-3p and FBXW7 in
TGCTs and NT
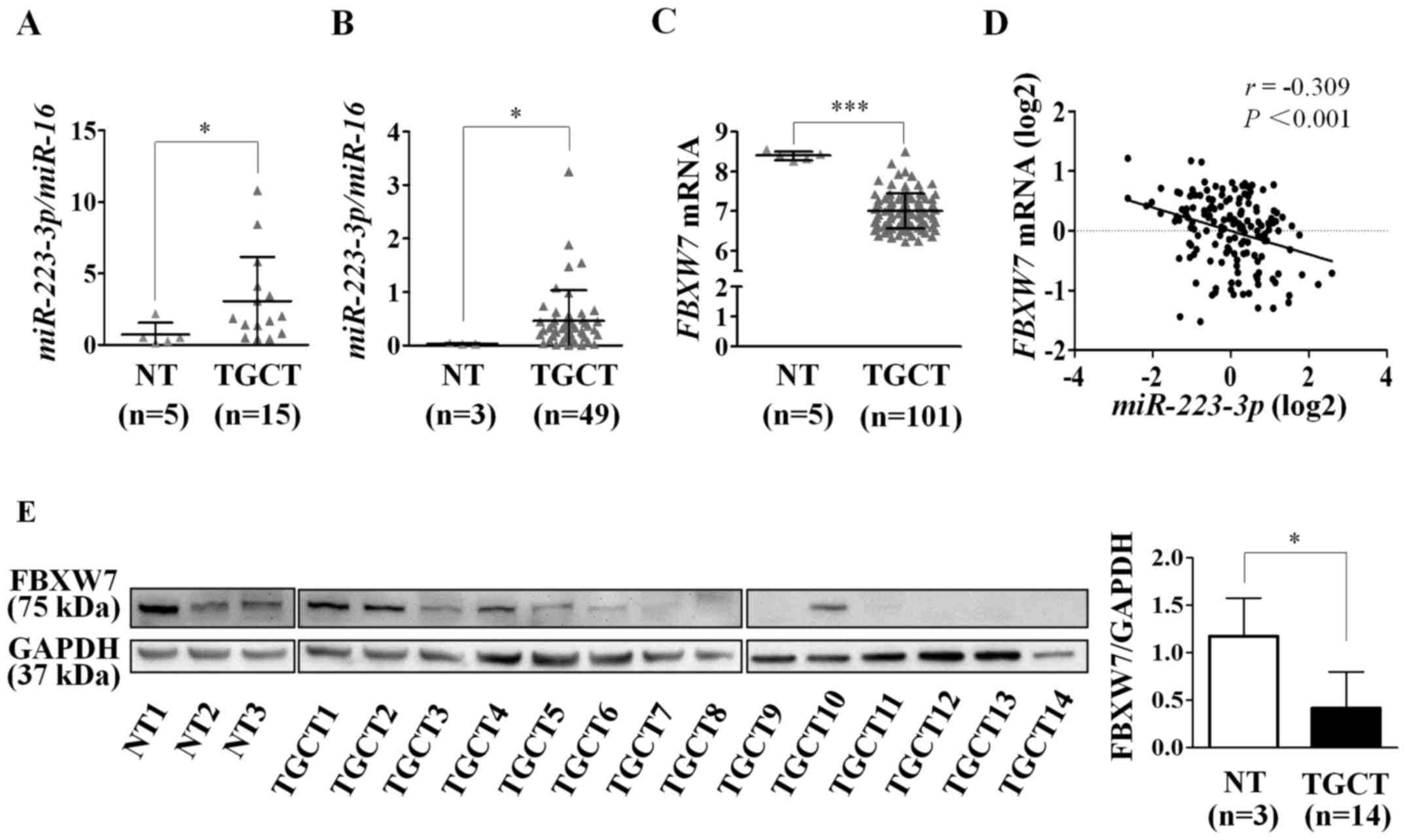
To validate our previous observation of
miR-223-3p overexpression in TGCTs, we re-analyzed
miR-223-3p expression from the miRNA profiling data of
Gillis et al (7), with
inclusion of 49 TGCTs and 3 NT. In agreement with our previous
finding (10) (Fig. 1A), miR-223-3p was
overexpressed in TGCTs compared to NT (P=0.011; Fig. 1B).
To determine whether FBXW7 could be a
candidate target of miR-223-3p in TGCT, we analyzed
FBXW7 expression from the microarray gene expression data of
101 TGCTs and 5 NT in the Gene Expression Omnibus (GEO) database
(accession no. GSE3218). Indeed, we found that FBXW7 mRNA
expression was decreased in TGCTs as compared to NT (P<0.001;
Fig. 1C). We further assessed the
correlation between miR-223-3p and FBXW7 expression
levels using miRNA and gene expression profiles from the TCGA
testicular cancer datasets. The analysis revealed an inverse
correlation (r=−0.309; P<0.001; Fig. 1D), supporting the miRNA-target
relationship.
Additionally, we also quantified FBXW7 protein
expression in 3 NT and 14 TGCT samples by western blot analysis. As
shown in Fig. 1E, the expression
of FBXW7 was low or undetectable in 10/14 TGCTs (71.4%), and
moderate or high in the remaining four tumors. By contrast, all
three NT showed moderate to high expression of FBXW7. Consistent
with the mRNA expression pattern, the FBXW7 protein level in TGCTs
was lower than in NT (P=0.023).
Effect of miR-223-3p modulation on FBXW7
in TGCT cell lines
To further determine whether miR-223-3p could
regulate FBXW7 in TGCT, we performed miR-223-3p
overexpression and inhibition in two TGCT cell lines and evaluated
the effect on FBXW7 protein expression using western blot analysis.
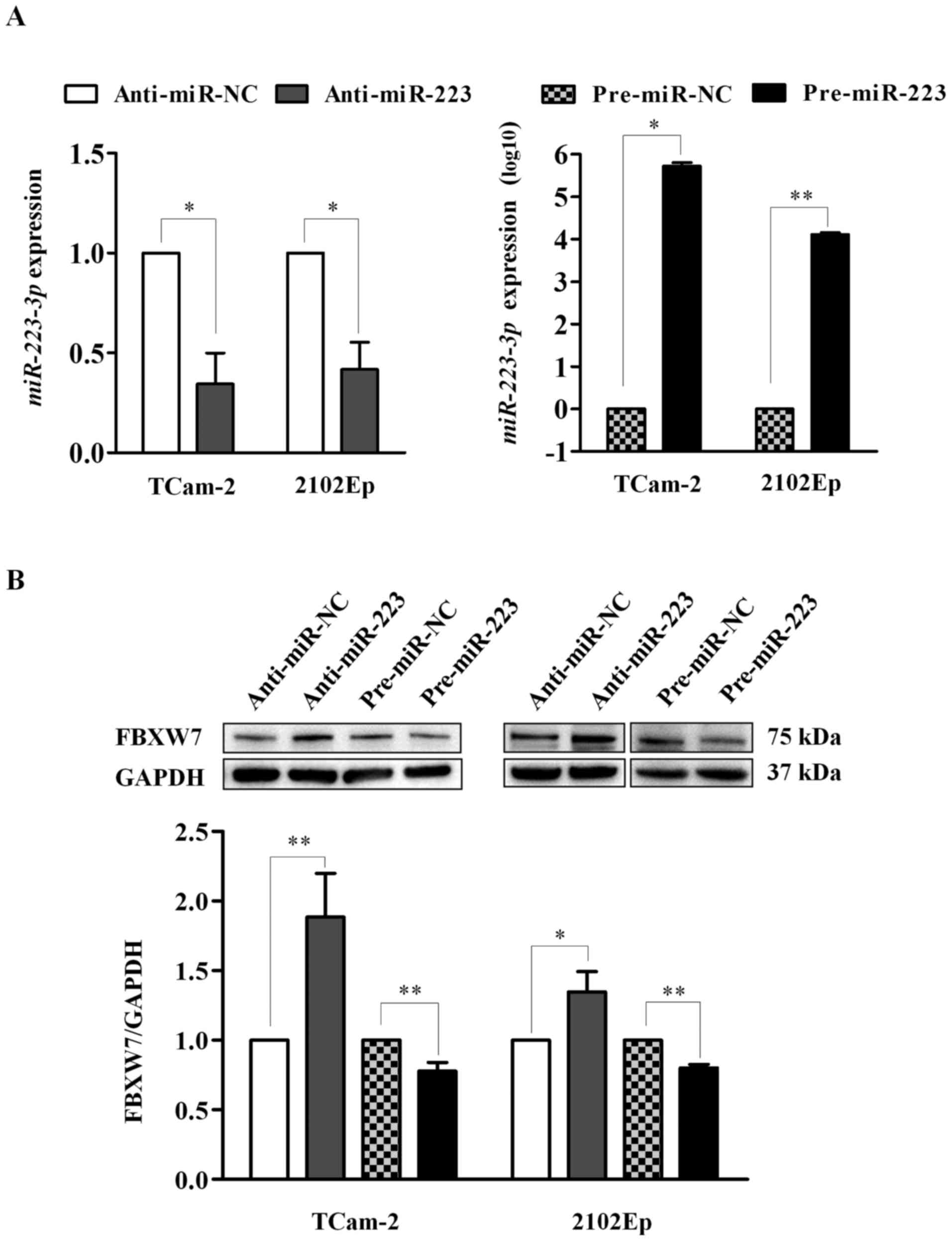
As shown in Fig. 2A, cells
transfected with anti-miR-223 showed significantly lower
miR-223-3p expression than the anti-miR-NC-treated cells in
both cell lines (P=0.018 for both). Similarly, miR-223-3p
expression was significantly increased in cells transfected with
pre-miR-223 relative to its negative control (TCam-2: P=0.014 and
2102Ep: P=0.003). The data support the efficiency of
transfection.
Furthermore, inhibition of miR-223-3p led to
a significant increase of FBXW7 expression in TCam-2 (1.9-fold;
P=0.003) and 2102Ep (1.3-fold, P=0.015) cells. Similarly,
overexpression of miR-223-3p significantly reduced FBXW7
expression in both cell lines (0.8-fold and P<0.01 for both)
(Fig. 2B). The findings indicate
that miR-223-3p suppresses FBXW7 expression in human TGCT
cells.
Functional consequences of miR-223-3p
regulation in TGCT cells
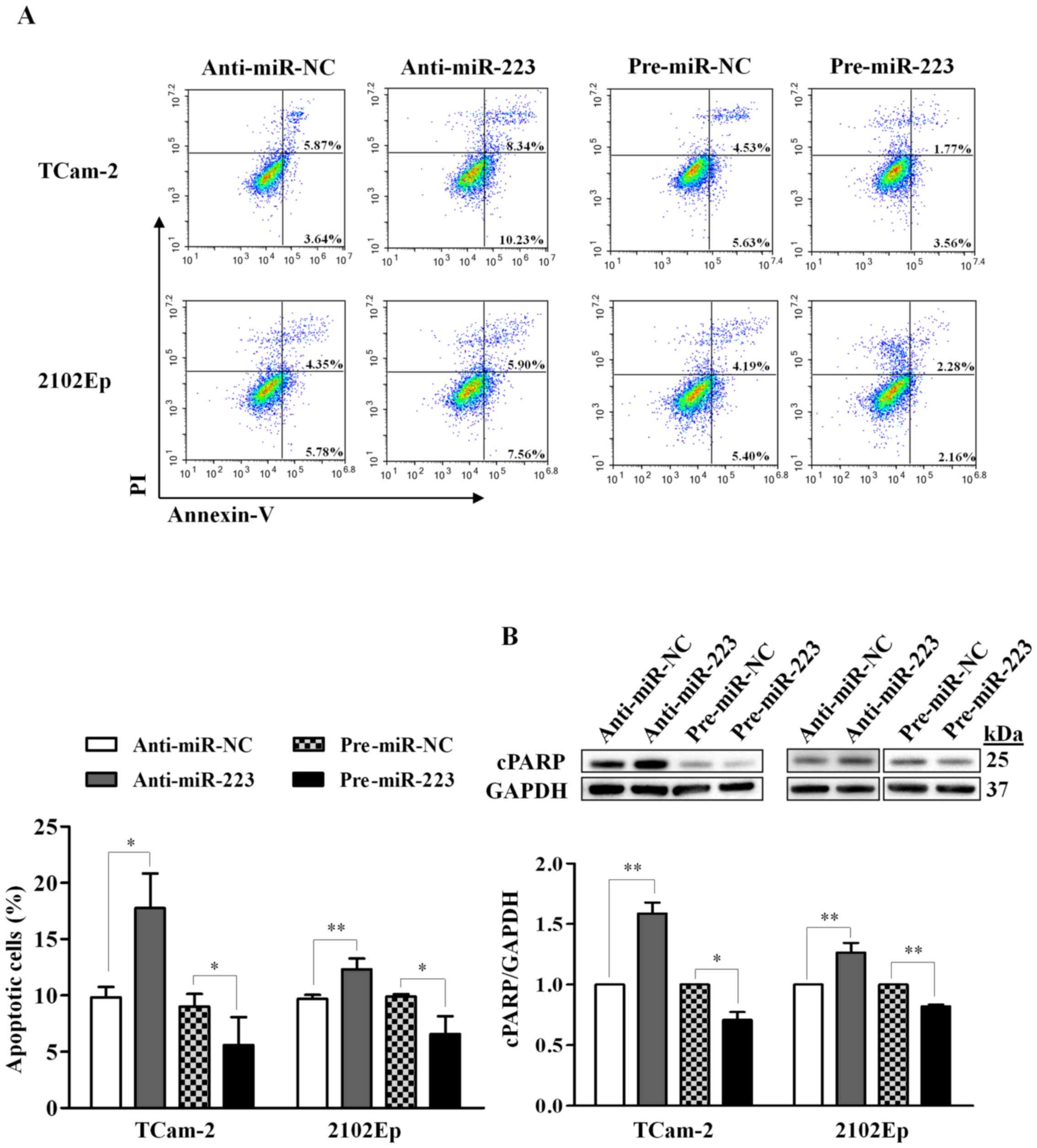
To explore the functional role of miR-223-3p
on apoptosis, we investigated the effect using flow cytometric
detection of Annexin V-stained cells as well as by western blot
analysis of cleaved PARP (cPARP, 25 kDa), which is an apoptosis
marker. For the Annexin V assay, we observed that inhibition of
miR-223-3p in TCam-2 cells significantly increased apoptotic
cells by 80% (P=0.018), while overexpression of miR-223-3p
reduced apoptotic cells by 38% (P=0.038), relative to their
respective negative controls (Fig.
3A). Similar effects were also observed in 2102Ep cells,
however, the effect was less pronounced compared to TCam-2 cells
(27% increase in the miR-223-3p inhibition, P=0.009; 34%
decrease in the miR-223-3p overexpression, P=0.016; Fig. 3A).
For the cPARP detection, silencing of
miR-223-3p led to a significant increase of cPARP expression
in both TCam-2 (1.6-fold, P=0.003) and 2102Ep (1.3-fold, P=0.007)
cells, while overexpressing miR-223-3p resulted in a
significant decrease of cPARP expression (TCam-2: 0.7-fold,
P=0.016; 2102Ep: 0.8-fold, P=0.002) (Fig. 3B). These results indicate that
miR-223-3p inhibits apoptosis in TGCT cells.
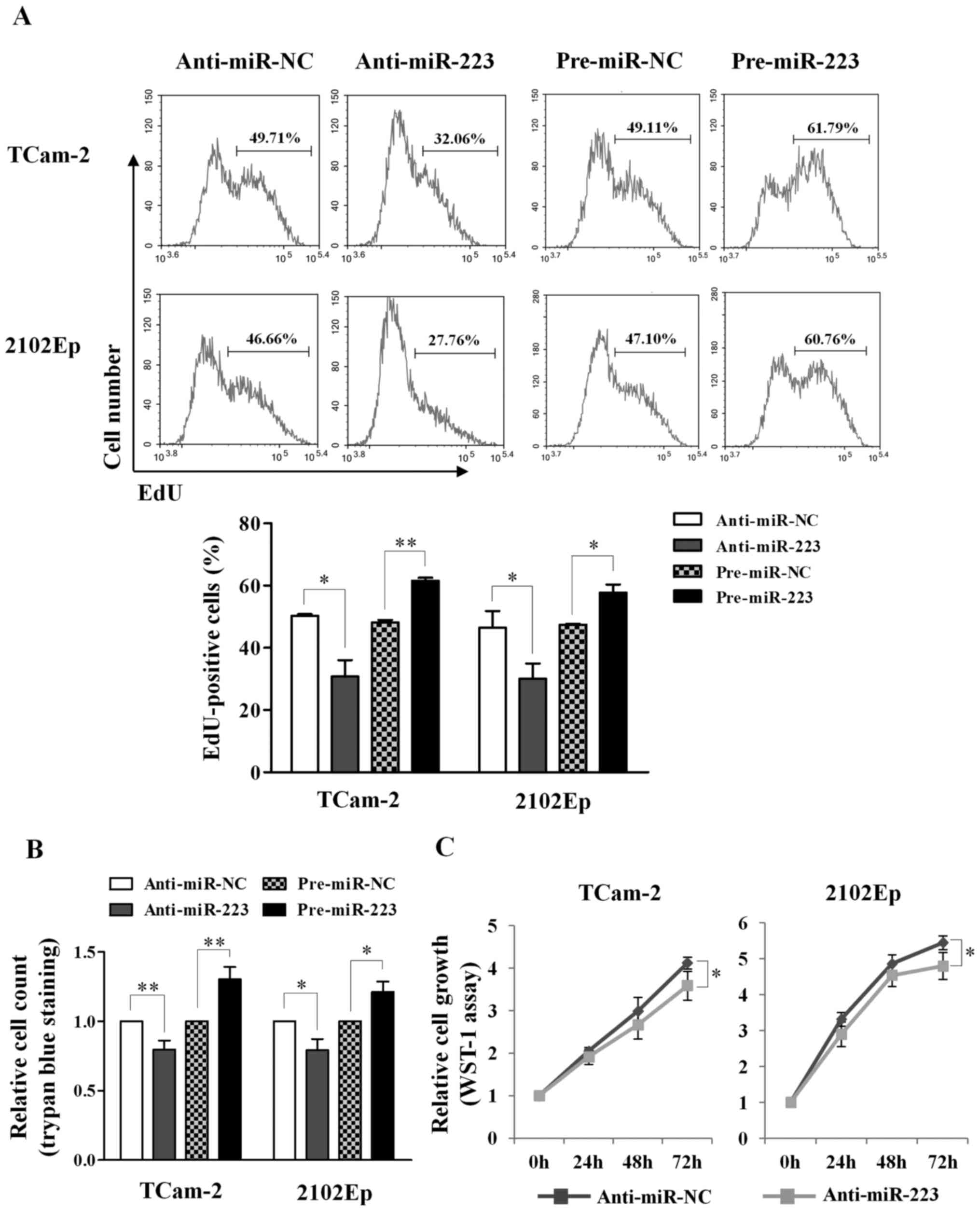
For cell proliferation, we applied three different
assays: Click-iT EdU, WST-1 and trypan blue exclusion. Using the
EdU assay, we observed reduction of EdU-positive cells upon
silencing of miR-223-3p in both TCam-2 (50.3 vs. 30.8%,
P=0.026) and 2102Ep cells (46.5 vs. 30.0%, P=0.045), and increase
of EdU-positive cells upon overexpression of miR-223-3p
(TCam-2: 48.1 vs. 61.6%, P=0.003; 2102Ep: 47.4 vs. 57.7%, P=0.027)
(Fig. 4A). Similarly, the trypan
blue exclusion assay revealed reduction of cell count upon
silencing of miR-223-3p (TCam-2: 0.8-fold, P=0.004; 2102Ep:
0.8-fold, P=0.016) and increase of cell number upon overexpression
of miR-223-3p (TCam-2: 1.3-fold, P=0.006; 2102Ep: 1.2-fold,
P=0.011) (Fig. 4B). The WST-1
assay also showed that silencing of miR-223-3p reduced cell
growth at 72-h post-transfection in both TCam-2 (P=0.043) and
2102Ep (P=0.041) cells (Fig. 4C).
Taken together, the results support that miR-223-3p promotes
cell proliferation in TGCT cell lines.
miR-223-3p mediates regulation of cell
growth and apoptosis through FBXW7 in TGCT
Given that FBXW7 expression is a well-characterized
target of miR-223-3p, we tested whether ectopically
expressed FBXW7 could rescue the miR-223-3p-mediated
apoptotic and proliferative effects. We co-transfected TCam-2 cells
with pre-miR-223 together with a plasmid expressing the entire open
reading frame of FBXW7 without the miR-223-3p binding
site (pCMV-Myc FBXW7) or a vector control. The effects on cell
apoptosis and proliferation were determined using western blot
analysis of cPARP and trypan blue exclusion assay, respectively. As
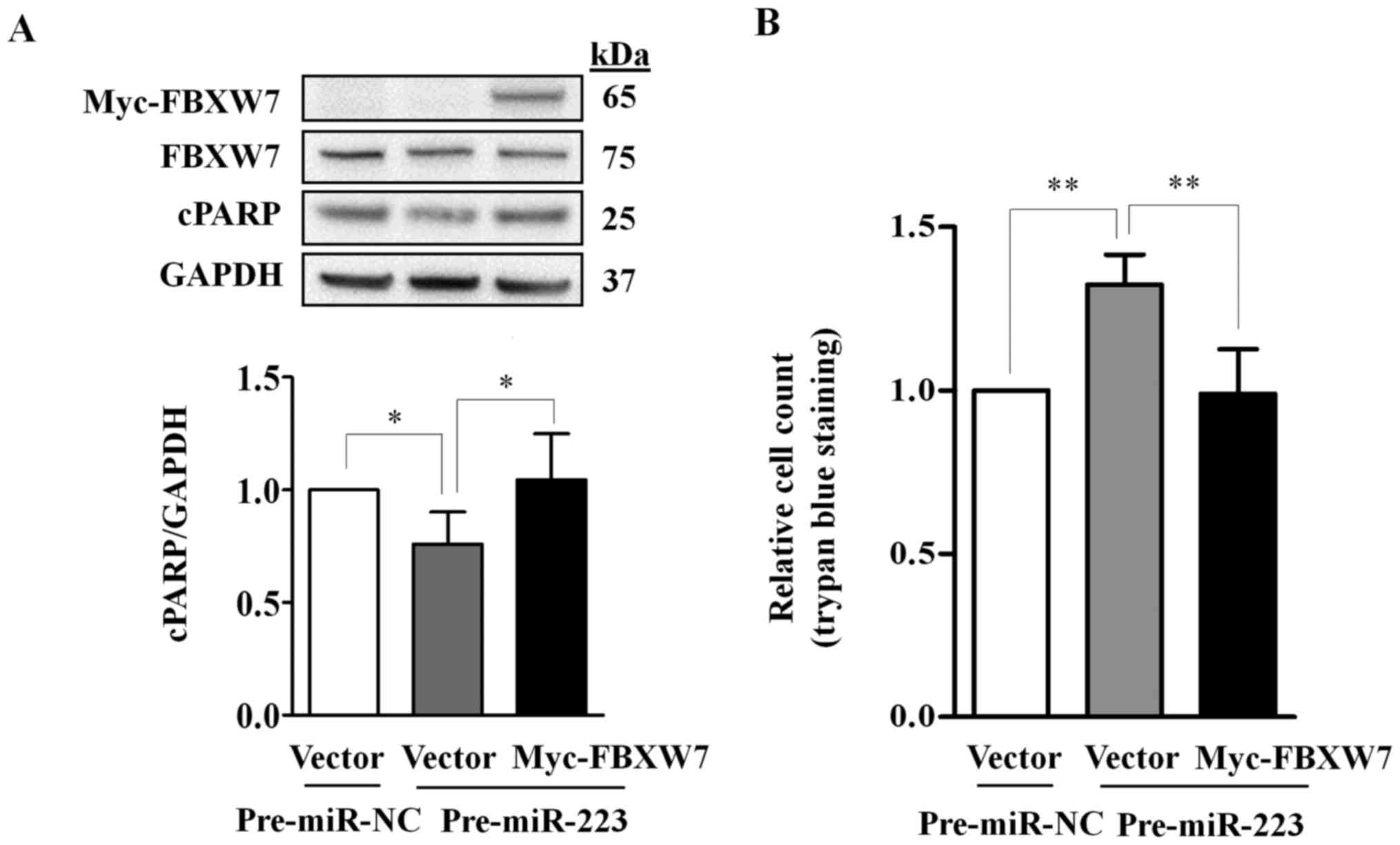
shown in Fig. 5A, the endogenous
FBXW7 was reduced in both cells co-transfected with pre-miR-223 and
Myc-FBXW7 or vector control as compared with the negative
control-transfected cells, indicating the suppression of endogenous
FBXW7 by miR-223-3p overexpression.
Ectopic expression of miR-223-3p
significantly reduced the abundance of cPARP (P=0.043) and
increased the number of live cells (P=0.006) as compared to their
respective controls; the effects were abolished by the ectopically
expressed FBXW7 (Fig. 5).
Together, our data indicate that miR-223-3p regulates cell
growth and apoptosis in TGCT cells through FBXW7.
Discussion
miR-223-3p expression was found higher in
TGCTs than NTs in our previous study (10), and here, we validated the findings
in independent cohorts using previously published dataset (7). Deregulation of miR-223-3p has
been observed in a variety of tumor types. Overexpression was found
in T-cell acute lymphoblastic leukemia (11), oral (12), esophageal (14), gastric (20,21),
bladder (22), and pancreatic
(23) cancers, while its reduced
expression has been reported in osteosarcoma (24), chronic lymphocytic leukemia
(25), and intrahepatic
cholangiocarcinoma (26). These
findings indicate that miR-223-3p plays vital roles in a
variety of tumor types, either as an oncogene or tumor suppressor
depending on the cellular contexts. Consistent with its dual role,
miR-223-3p has been shown to function as an oncogene in
T-cell acute lymphoblastic leukemia, gastric and lung cancers
(13,15,21,27,28),
and as a tumor suppressor in cutaneous T-cell lymphoma and prostate
cancer (29,30). Given its diverse function in
different cancer types, we characterized the functional role of
miR-223-3p in TGCT cells. miR-223-3p was shown to
promote cell proliferation in TGCT cell lines in all three methods
applied and which are based on different principles: the Click-iT
EdU assay allows the detection of the thymidine analog EdU
incorporated into cellular DNA during replication; the WST-1 assay
is based on the metabolic activity of cells for conversion of the
tetrazolium salt WST-1 into a colored dye, and the trypan blue
exclusion assay provides direct counting of the number of live
cells. Our findings support its oncogenic role in TGCT by promoting
cell growth and inhibiting apoptosis in TGCT cell lines.
Additionally, miR-223-3p has been shown to
modulate drug response in several cancer types (31–35).
Importantly, miR-223-3p regulates cisplatin sensitivity in
gastric and esophageal cancers (31,32).
Given that most TGCTs are responsive to cisplatin treatment, it is
intriguing to speculate that miR-223-3p may play an
important role in cisplatin sensitivity in TGCT. Further
investigations are warranted to evaluate the role of
miR-223-3p in cisplatin response in TGCT.
FBXW7 has been demonstrated as a direct target of
miR-223-3p using luciferase reporter assays (13,15).
Here, we show that FBXW7 expression is lower in TGCTs than NT and
inversely correlated with miR-223-3p, and miR-223-3p
regulates FBXW7 protein expression using both gain- and
loss-of-function studies. Most importantly, ectopic expression of
the FBXW7 open reading frame can rescue the cell growth and
apoptosis effects mediated by miR-223-3p. Together, our
findings suggest that miR-223-3p regulates FBXW7 in TGCT and
this regulatory pathway plays an important role in TGCT
pathogenesis.
As afore-mentioned, FBXW7 is an E3 ubiquitin ligase
that degrades several proto-oncogenes involved in cell growth,
apoptosis, cell cycle regulation and differentiation (16). Therefore, the functional phenotypes
observed in this study could due to the loss of FBXW7-mediated
degradation of its substrates. Furthermore, numerous
cancer-associated mutations in FBXW7 have been found in many
cancer types (36), and loss of
FBXW7 function can lead to chromosomal instability and
tumorigenesis (37,38). These findings support the tumor
suppressor function of FBXW7 in human cancers. Although nothing is
known about its role in TGCT, FBXW7 is expressed specifically in
the undifferentiated spermatogonia and suppresses cell
proliferation of spermatogonial stem cell in mice (39). It is tempting to speculate that
loss of FBXW7 expression could lead to uncontrolled cell growth in
TGCT.
In conclusion, we report deregulation of
miR-223-3p and FBXW7 in human TGCT. Our findings also reveal
an oncogenic role of miR-223-3p through repression of the
FBXW7 tumor suppressor, suggesting that this regulation is
important for cell proliferation and apoptosis in TGCT. This study
provides additional evidence of miRNA function in testicular germ
cell tumorigenesis.
Acknowledgments
We thank Dr Leendert H.J. Looijenga and Dr Peter
Andrews for TGCT cell lines; the Cooperative Human Tissue Network
for frozen tissue samples; and the members of the sRNA Group for
valuable discussions and suggestions. This study was supported by
the Swedish Research Council (523-2009-3517 and 521-2010-3518), the
Swedish Cancer Society, the Cancer Research Funds of Radiumhemmet,
Karolinska Institutet and Stockholm County Council. J. Liu and H.
Shi are the recipients of China Scholarship Council training
grants.
References
|
1
|
Mannuel HD, Mitikiri N, Khan M and Hussain
A: Testicular germ cell tumors: Biology and clinical update. Curr
Opin Oncol. 24:266–271. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nigam M, Aschebrook-Kilfoy B, Shikanov S
and Eggener S: Increasing incidence of testicular cancer in the
United States and Europe between 1992 and 2009. World J Urol.
33:623–631. 2015. View Article : Google Scholar
|
|
3
|
McGlynn KA and Cook MB: Etiologic factors
in testicular germ-cell tumors. Future Oncol. 5:1389–1402. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Koychev D, Oechsle K, Bokemeyer C and
Honecker F: Treatment of patients with relapsed and/or
cisplatin-refractory metastatic germ cell tumours: An update. Int J
Androl. 34:e266–e273. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Palmer RD, Murray MJ, Saini HK, van Dongen
S, Abreu-Goodger C, Muralidhar B, Pett MR, Thornton CM, Nicholson
JC, Enright AJ, et al Children's Cancer and Leukaemia Group:
Malignant germ cell tumors display common microRNA profiles
resulting in global changes in expression of messenger RNA targets.
Cancer Res. 70:2911–2923. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gillis AJ, Stoop HJ, Hersmus R, Oosterhuis
JW, Sun Y, Chen C, Guenther S, Sherlock J, Veltman I, Baeten J, et
al: High-throughput microRNAome analysis in human germ cell
tumours. J Pathol. 213:319–328. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suh MR, Lee Y, Kim JY, Kim SK, Moon SH,
Lee JY, Cha KY, Chung HM, Yoon HS, Moon SY, et al: Human embryonic
stem cells express a unique set of microRNAs. Dev Biol.
270:488–498. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Voorhoeve PM, le Sage C, Schrier M, Gillis
AJ, Stoop H, Nagel R, Liu YP, van Duijse J, Drost J, Griekspoor A,
et al: A genetic screen implicates miRNA-372 and miRNA-373 as
oncogenes in testicular germ cell tumors. Cell. 124:1169–1181.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Özata DM, Li X, Lee L, Liu J, Warsito D,
Hajeri P, Hultman I, Fotouhi O, Marklund S, Ährlund-Richter L, et
al: Loss of miR-514a-3p regulation of PEG3 activates the NF-kappa B
pathway in human testicular germ cell tumors. Cell Death Dis. In
press.
|
|
11
|
Chiaretti S, Messina M, Tavolaro S, Zardo
G, Elia L, Vitale A, Fatica A, Gorello P, Piciocchi A, Scappucci G,
et al: Gene expression profiling identifies a subset of adult
T-cell acute lymphoblastic leukemia with myeloid-like gene features
and over-expression of miR-223. Haematologica. 95:1114–1121. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Manikandan M, Deva Magendhra Rao AK,
Arunkumar G, Manickavasagam M, Rajkumar KS, Rajaraman R and
Munirajan AK: Oral squamous cell carcinoma: microRNA expression
profiling and integrative analyses for elucidation of
tumourigenesis mechanism. Mol Cancer. 15:282016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mavrakis KJ, Van Der Meulen J, Wolfe AL,
Liu X, Mets E, Taghon T, Khan AA, Setty M, Rondou P, Vandenberghe
P, et al: A cooperative microRNA-tumor suppressor gene network in
acute T-cell lymphoblastic leukemia (T-ALL). Nat Genet. 43:673–678.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kurashige J, Watanabe M, Iwatsuki M,
Kinoshita K, Saito S, Hiyoshi Y, Kamohara H, Baba Y, Mimori K and
Baba H: Overexpression of microRNA-223 regulates the ubiquitin
ligase FBXW7 in oesophageal squamous cell carcinoma. Br J Cancer.
106:182–188. 2012. View Article : Google Scholar :
|
|
15
|
Li J, Guo Y, Liang X, Sun M, Wang G, De W
and Wu W: MicroRNA-223 functions as an oncogene in human gastric
cancer by targeting FBXW7/hCdc4. J Cancer Res Clin Oncol.
138:763–774. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Welcker M and Clurman BE: FBW7 ubiquitin
ligase: A tumour suppressor at the crossroads of cell division,
growth and differentiation. Nat Rev Cancer. 8:83–93. 2008.
View Article : Google Scholar
|
|
17
|
Wang L, Ye X, Liu Y, Wei W and Wang Z:
Aberrant regulation of FBW7 in cancer. Oncotarget. 5:2000–2015.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mizuno Y, Gotoh A, Kamidono S and Kitazawa
S: Establishment and characterization of a new human testicular
germ cell tumor cell line (TCam-2). Nippon Hinyokika Gakkai Zasshi.
84:1211–1218. 1993.In Japanese.
|
|
19
|
Andrews PW, Goodfellow PN, Shevinsky LH,
Bronson DL and Knowles BB: Cell-surface antigens of a clonal human
embryonal carcinoma cell line: Morphological and antigenic
differentiation in culture. Int J Cancer. 29:523–531. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Petrocca F, Visone R, Onelli MR, Shah MH,
Nicoloso MS, de Martino I, Iliopoulos D, Pilozzi E, Liu CG, Negrini
M, et al: E2F1-regulated microRNAs impair TGFbeta-dependent
cell-cycle arrest and apoptosis in gastric cancer. Cancer Cell.
13:272–286. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li X, Zhang Y, Zhang H, Liu X, Gong T, Li
M, Sun L, Ji G, Shi Y, Han Z, et al: miRNA-223 promotes gastric
cancer invasion and metastasis by targeting tumor suppressor
EPB41L3. Mol Cancer Res. 9:824–833. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gottardo F, Liu CG, Ferracin M, Calin GA,
Fassan M, Bassi P, Sevignani C, Byrne D, Negrini M, Pagano F, et
al: Micro-RNA profiling in kidney and bladder cancers. Urol Oncol.
25:387–392. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bloomston M, Frankel WL, Petrocca F,
Volinia S, Alder H, Hagan JP, Liu CG, Bhatt D, Taccioli C and Croce
CM: MicroRNA expression patterns to differentiate pancreatic
adenocarcinoma from normal pancreas and chronic pancreatitis. JAMA.
297:1901–1908. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu J, Yao Q, Hou Y, Xu M, Liu S, Yang L,
Zhang L and Xu H: MiR-223/Ect2/p21 signaling regulates osteosarcoma
cell cycle progression and proliferation. Biomed Pharmacother.
67:381–386. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fazi F, Racanicchi S, Zardo G, Starnes LM,
Mancini M, Travaglini L, Diverio D, Ammatuna E, Cimino G, Lo-Coco
F, et al: Epigenetic silencing of the myelopoiesis regulator
microRNA-223 by the AML1/ETO oncoprotein. Cancer Cell. 12:457–466.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Karakatsanis A, Papaconstantinou I,
Gazouli M, Lyberopoulou A, Polymeneas G and Voros D: Expression of
microRNAs, miR-21, miR-31, miR-122, miR-145, miR-146a, miR-200c,
miR-221, miR-222, and miR-223 in patients with hepatocellular
carcinoma or intrahepatic cholangiocarcinoma and its prognostic
significance. Mol Carcinog. 52:297–303. 2013. View Article : Google Scholar
|
|
27
|
Mansour MR, Sanda T, Lawton LN, Li X,
Kreslavsky T, Novina CD, Brand M, Gutierrez A, Kelliher MA,
Jamieson CH, et al: The TAL1 complex targets the FBXW7 tumor
suppressor by activating miR-223 in human T cell acute
lymphoblastic leukemia. J Exp Med. 210:1545–1557. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liang H, Yan X, Pan Y, Wang Y, Wang N, Li
L, Liu Y, Chen X, Zhang CY, Gu H, et al: MicroRNA-223 delivered by
platelet-derived microvesicles promotes lung cancer cell invasion
via targeting tumor suppressor EPB41L3. Mol Cancer. 14:582015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
McGirt LY, Adams CM, Baerenwald DA,
Zwerner JP, Zic JA and Eischen CM: miR-223 regulates cell growth
and targets proto-oncogenes in mycosis fungoides/cutaneous T-cell
lymphoma. J Invest Dermatol. 134:1101–1107. 2014. View Article : Google Scholar :
|
|
30
|
Kurozumi A, Goto Y, Matsushita R, Fukumoto
I, Kato M, Nishikawa R, Sakamoto S, Enokida H, Nakagawa M, Ichikawa
T, et al: Tumor-suppressive microRNA-223 inhibits cancer cell
migration and invasion by targeting ITGA3/ITGB1 signaling in
prostate cancer. Cancer Sci. 107:84–94. 2016. View Article : Google Scholar
|
|
31
|
Streppel MM, Pai S, Campbell NR, Hu C,
Yabuuchi S, Canto MI, Wang JS, Montgomery EA and Maitra A: MicroRNA
223 is upregulated in the multistep progression of Barrett's
esophagus and modulates sensitivity to chemotherapy by targeting
PARP1. Clin Cancer Res. 19:4067–4078. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou X, Jin W, Jia H, Yan J and Zhang G:
MiR-223 promotes the cisplatin resistance of human gastric cancer
cells via regulating cell cycle by targeting FBXW7. J Exp Clin
Cancer Res. 34:282015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Eto K, Iwatsuki M, Watanabe M, Ishimoto T,
Ida S, Imamura Y, Iwagami S, Baba Y, Sakamoto Y, Miyamoto Y, et al:
The sensitivity of gastric cancer to trastuzumab is regulated by
the miR-223/FBXW7 pathway. Int J Cancer. 136:1537–1545. 2015.
View Article : Google Scholar
|
|
34
|
Kumar V, Palermo R, Talora C, Campese AF,
Checquolo S, Bellavia D, Tottone L, Testa G, Miele E, Indraccolo S,
et al: Notch and NF-κB signaling pathways regulate miR-223/FBXW7
axis in T-cell acute lymphoblastic leukemia. Leukemia.
28:2324–2335. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li R, Wu S, Chen X, Xu H, Teng P and Li W:
miR-223/FBW7 axis regulates doxorubicin sensitivity through
epithelial mesenchymal transition in non-small cell lung cancer. Am
J Transl Res. 8:2512–2524. 2016.PubMed/NCBI
|
|
36
|
Akhoondi S, Sun D, von der Lehr N,
Apostolidou S, Klotz K, Maljukova A, Cepeda D, Fiegl H, Dafou D,
Marth C, et al: FBXW7/hCDC4 is a general tumor suppressor in human
cancer. Cancer Res. 67:9006–9012. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rajagopalan H, Jallepalli PV, Rago C,
Velculescu VE, Kinzler KW, Vogelstein B and Lengauer C:
Inactivation of hCDC4 can cause chromosomal instability. Nature.
428:77–81. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mao JH, Perez-Losada J, Wu D, Delrosario
R, Tsunematsu R, Nakayama KI, Brown K, Bryson S and Balmain A:
Fbxw7/Cdc4 is a p53-dependent, haploinsufficient tumour suppressor
gene. Nature. 432:775–779. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kanatsu-Shinohara M, Onoyama I, Nakayama
KI and Shinohara T: Skp1-Cullin-F-box (SCF)-type ubiquitin ligase
FBXW7 negatively regulates spermatogonial stem cell self-renewal.
Proc Natl Acad Sci USA. 111:8826–8831. 2014. View Article : Google Scholar : PubMed/NCBI
|