Introduction
Suppressor of Fused (SuFu) is a key regulator of
Hedgehog (Hh) signaling and Gli activities. Ablation of SuFu in
mouse embryos leads to severe neural tube defects and embryonic
lethality (1,2). It is widely speculated that SuFu
inhibits Gli-mediated Hh pathway through sequestering Gli2 protein
in the cytoplasm and promoting proteolytic degradation of
full-length (FL) Gli3 (3–5). SuFu also traverses into the nucleus
to repress transcription by recruiting the SAP18-mSin3 complex to
Gli-binding regions (6). However,
recent study showed that SuFu can also play a positive role in the
maximal activation of Hh signaling likely through protecting FL
Gli2 and Gli3 proteins from SPOP-mediated ubiquitination and
degradation (7–9). Other studies delineate the role of
SuFu in tumorigenesis and implicate SUFU as a tumor suppressor.
Taylor et al found that germline mutations or deletions of
SUFU lead to medulloblastoma in a subset of children (10). SUFU is also known to be a rare
cause of Gorlin syndrome, of which the patients always harbor
mutations in PTCH1 (11).
Furthermore, deletion of SUFU has been identified in other human
tumors, including rhabdomyosarcoma (12), basal cell carcinoma (13) and prostate cancer (14), which further support SUFU as a
tumor suppressor gene. Despite the central and conserved roles of
SuFu in Hh signaling pathway and tumor, little is known about its
regulation. Limited studies showed that SuFu undergoes
ubiquitin-proteasomal degradation in response to Shh signaling in
freshly isolated mouse embryonic fibroblasts and in embryonic
tissues (15). In addition, the
recombinant human SuFu was found to be phosphorylated and
stabilized by purified PKA (16).
In our previous study (17), we employed a yeast two-hybrid
approach to identify human SuFu interacting proteins. We found NIMA
(never in mitosis A)-related expressed kinase 2A (Nek2A) as one of
the SuFu-interacting proteins. Nek2A belongs to the Nek family of
serine/threonine kinases, and is expressed in vertebrates as two
main splice variants, Nek2A and Nek2B. The C terminus of Nek2A, but
not Nek2B, contains the binding site for protein phosphatase 1 and
motifs targeting the protein for ubiquitin-mediated degradation
after mitotic entry. As a cell cycle-regulated kinase, Nek2A
localizes to centrosomes and exhibits increased activity in S and
G2 phases (18). During mitosis,
Nek2 contributes to spindle pole formation through phosphorylation
of centriolar cohesion proteins, including C-Nap1, rootletin, and
Nep which is required for microtubule anchoring and spindle
assembly (19–23).
Nek2 has emerged as an important oncogene due to its
regulatory role in mitosis and cancer-related signaling pathways.
Increased Nek2 expression has been tied to serosal invasion,
lymphatic invasion, peritoneal dissemination and poor prognosis of
colorectal cancer (24), for which
the reason may be that Nek2 was associated with beta-catenin
relocalization from membrane to cytoplasma and nucleus (25). In breast cancer studies, high Nek2
expression correlates with poor prognosis, and in various human
breast cancer cell lines, Nek2 knockdown induces aneuploidy and
cell cycle arrest that leads to cell death (26). In addition, analysis of the gene
expression profiles of breast cancer samples revealed that
co-elevated levels of Hec1 and Nek2 correlate with the shortest
survival (27). Moreover in
ovarian cancer, Nek2 mRNA expression is upregulated, especially in
drug-resistant cells. The bioinformatic analysis revealed that Nek2
may directly or indirectly interact with a number of genes,
proteins, microRNAs associated with drug resistance in ovarian and
other types of cancer (28).
Aberrant Nek2 expression has also been found in other cancers, such
as non-small cell lung cancer and malignant peripheral nerve sheath
tumor (29,30).
Given the importance of SuFu and Nek2A in
tumorigenesis, we further dissected the functional relationship of
these two proteins. We show here that Nek2A impairs
ubiquitin/proteasome-mediated SuFu degradation, thus negatively
modulates Hh transduction. Interestingly, in response to SuFu
stabilization, Hh adjusts the transcription and expression of Nek2A
in mammalian cells. Therefore, Nek2A functions as part of a
negative feedback loop that modulates Hh activity, which may
provide new insights into a dynamic process of Hh/Gli signaling
regulation driven by feedback adaptation mechanisms.
Materials and methods
Reagents, antibodies and small molecular
inhibitors
Lubrol-PX and other chemicals were purchased from
Sigma-Aldrich (St. Louis, MO, USA). Lipofectamine 2000 was obtained
from Invitrogen Life Technologies (Carlsbad, CA, USA). The sources
for monoclonal antibodies are as follows: Sigma-Aldrich (anti-Flag
M2, F3165; anti-c-Myc, M4439), Abcam (anti-Pathed/PTCH1, ab55629;
anti-SuFu, ab52913), Santa Cruz Biotechnology (normal rabbit IgG,
sc-2027; anti-Gli2, sc-271786), BD Pharmingen (anti-Nek2 MAB,
610594), Covance (anti-ubiquitin P4G7, MMS-258R), and Millipore
(anti-GAPDH, MAB374). Polyclonal antibodies were purchased from
Abcam (anti-SuFu, ab28083; anti-Smo, ab38686; anti-Gli1, ab92611).
The working concentrations for small molecular inhibitors and
chemicals are as follows: GANT61 (20 mM, G9048; Sigma-Aldrich),
MG-132 (25 mM, S2619; Selleck), purmorphamine (20 mM, S3042;
Selleck), CHX (100 mg/ml, C7689; Solarbio). DMSO (0231; Amresco)
was used as the solvent for the inhibitors and the vehicle
control.
Plasmids
For mammalian cell expression, human cDNAs of SuFu
(NM_016169) from human fetal brain cDNA library (Takara, Otsu,
Japan) were amplified by PCR and cloned into mammalian expression
vectors tagged with Flag. C-terminal Myc/his tagged human cDNA of
Nek2A (NM_002497) was generated by PCR using the same strategy as
described above and subcloned in pcDNA3.1. The miRNAi expression
vectors that suppress Nek2A expression were generated by the
BLOCK-iT™ Pol II miR-RNAi Expression system (Invitrogen). The
oligonucleotide sequences for miRNAi constructs miR-Nek2A-80, -202
and -603 were: 5′-GGA AGA GTG ATG GCA AGA TAT-3′; 5′-GTT CGT TAC
TAT GAT CGG ATT-3′; 5′-ATT GGG CTG CTT GCT GTA TGA-3′. The
authenticity of the constructs was verified by DNA sequencing.
Cell culture, transfection and Bcl-2
reporter assay
HEK293T, H4 (American Type Culture Collection,
Manassas, VA, USA) and CaES-17 (Shanghai Institute of Cell Biology,
Shanghai, China) cells were cultured in DMEM, supplemented with 10%
fetal calf serum, penicillin (100 U/ml) and streptomycin (100
mg/ml), and maintained at 37°C in an atmosphere of 5%
CO2 and 95% humidity. Transient transfection of cells
was performed with the standard calcium phosphate technique or with
Lipofectamine 2000 according to instructions of the manufacturer.
For Bcl-2 luciferase reporter assay, HEK293T cells were seeded in
triplicate in 12-well plates and allowed to settle for 12 h. Three
hundred nanograms of Bcl-2 luciferase reporter plasmid and the
internal control plasmid pRL-TK (10 ng/well) were co-transfected
into HEK293T cells. Plasmids for the expression of Gli1 and Gli2
(0.7 mg/well) were introduced to exaggerate the luciferase signal.
The luciferase and Renilla signals were measured 48 h after
transfection using Dual Luciferase Reporter assay kit (Promega,
Madison, WI, USA). Each reporter gene assay was performed in
triplicate.
Ubiquitination assay
To assay the effects of Nek2A on the ubiquitination
of SuFu, Flag-tagged SuFu and Myc-tagged Nek2A were co-transfected
into HEK293T cells. After 48 h of transfection, cells were treated
with MG-132 for indicated times (0, 4 and 8 h) before they were
lysed in Co-IP buffer (50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 50 mM
NaF, 2 mM EDTA, 10% glycerol, 0.5% NP-40). Then
immuno-precipitation was carried out with anti-Flag M2 Magnetic
Beads (M8823; Sigma-Aldrich), and the immunoprecipitated proteins
were subjected to 8% SDS-PAGE, followed by western blot
analysis.
Western blotting
Cells were harvested 24-48 h after transfection and
subjected to western blot analysis as described previously
(31). Cells were washed with cold
PBS and then suspended in protein lysis buffer at 4°C for 30 min
and centrifuged (12,000 rpm, 15 min at 4°C) to maintain the
supernatant. Protein concentrations were determined using Pierce™
BCA Protein assay kit (Thermo Scientific™, Rockford, IL, USA)
according to the standard protocol of the manufacturer. Proteins
were subsequently resolved by SDS-PAGE and transferred to a
polyvinylidene difluoride membrane (Bio-Rad), and then probed with
the specified primary antibody followed by appropriate secondary
antibody. The immunostaining was visualized by using Kodak X-ray
film, which were subsequently scanned with an Epson 1680 scanner.
Quantitative analysis was performed on scanned images of blots
using ImageJ software (NIH Image analysis website http://rsb.info.nih.gov/ij/)
N-Shh conditioned medium
HEK293T cells were transfected with 10 mg of the
Flag-N-Shh plasmid per 10-cm cell culture dish. Twelve hours after
addition of the plasmids, the media were replaced with 10 ml of
DMEM with 2% fetal calf serum per dish. The culture media were
harvested 24 h after the medium change. Then, cells were incubated
for an additional 24 h with 10 ml of DMEM containing 2% fetal calf
serum. After that, the media were harvested and combined with the
previously collected media. The combined medium was termed N-Shh
conditioned medium and stored at 4°C until used.
Quantitative real-time PCR analysis
Total RNA of HEK293T cells was harvested using
TRIzol® reagent (Life Technologies™) and evaluated by
quantitative real-time PCR. Briefly, 1 mg of total RNA was employed
to prepare cDNA via reverse transcription using a
PrimeScript® RT reagent kit with gDNA Eraser (DRR047A;
Takara). Quantitative real-time PCR was performed using
SYBR® Premix Ex Taq™ II (Tli RnaseH Plus) (DRR820A;
Takara) in an ABI StepOnePlus™ Real-Time PCR system (Applied
Biosystem, Inc.). GAPDH was used as an internal control to
normalize the variability in expression levels. The primers are
shown in Table I.
 | Table IThe real-time PCR primers. |
Table I
The real-time PCR primers.
| PCR primer | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| NEK2A |
TCTTAATCTTCCATCCTCAG |
CTATACAGAAAGGCATGGCT |
| SUFU |
CCCAAGCTTGATGGCGGAGCTGCGGCCTAG |
CGCGGATCCCTAGTGTAGCGGACTGTCGAAC |
| GLI1 |
TCCTACCAGAGTCCCAAGTT |
CCCTATGTGAAGCCCTATTT |
| GLI2 |
CCTGGCATGACTACCACTATGAG |
GGCTTGGCTGGCATGTTG |
| GAPDH |
CAGGGCTGCTTTTAACTCTGGT |
GATTTTGGAGGGATCTCGCT |
In silico promoter analysis and chromatin
immunoprecipitation (ChIP) assay
Identification of transcription factor binding sites
in the NEK2A promoter was performed using the MatInspector module
of the Genomatix database, together with Matrix Family Library
version 8.3 (32). ChIP assay was
performed using the EZ-ChIP kit (Upstate Biotechnology) according
to the manufacturer's manual. Briefly, cells were cross-linked in
1% formaldehyde, and DNA was sonicated into a range of 200–1000
base pairs in size using a Bioruptor Sonicator (Diagenode) for five
cycles of 3 sec on/3 sec off. The extracts were pre-cleared in
BSA-blocked protein A/G beads and incubated with antibodies or IgG
control overnight. After washes, DNA was eluted and
reverse-cross-linked overnight at 65°C, and then purified and
amplified by PCR. The primers for PCR are shown in Table II.
| Table IIThe PCR primers in ChIP assay. |
Table II
The PCR primers in ChIP assay.
| Promotor
region | Sequence | Forward
(5′-3′) | Reverse
(5′-3′) | Length (bp) |
|---|
| −3062 to −3054 | AGCCACCCA |
GTCTGTTGAGCAGGCTGGAGT |
TGTGGTGGCACGTCTGTAGTC | 124 |
| −2602 to −2594 | GCCCACTCA |
CAGCCTCCTGTGCTACTCTTT |
CCAACCCACTCCCTTATCCA | 560 |
| −1880 to −1872 | GACCTCCCA |
CTCCTGACCTCGTAATCCACC |
AGGCTGCTGCCAGATGCTAC | 213 |
| −449 to −441 | GCCCACCCG |
ACTCCTGGGCTCAAGCGACC |
GAGCTGAATACAAATTAGAAATACAGAC | 114 |
Statistical analysis
All data were expressed as mean ± SD for experiments
performed at least three times. The difference between 2 groups was
analyzed using Student's t-test or one-way ANOVA. Significance was
defined at P<0.05.
Results
Nek2A prevents SuFu from
proteasome-dependent degradation
SuFu was shown to be degraded rapidly in some cancer
cells (15). In a previous study,
we demonstrated that Nek2A is an interacting protein of SuFu and
Nek2A stabilizes SuFu (17). Here
we further investigated the detailed mechanism of SuFu
stabilization induced by Nek2A. We transfected HEK293T cells with
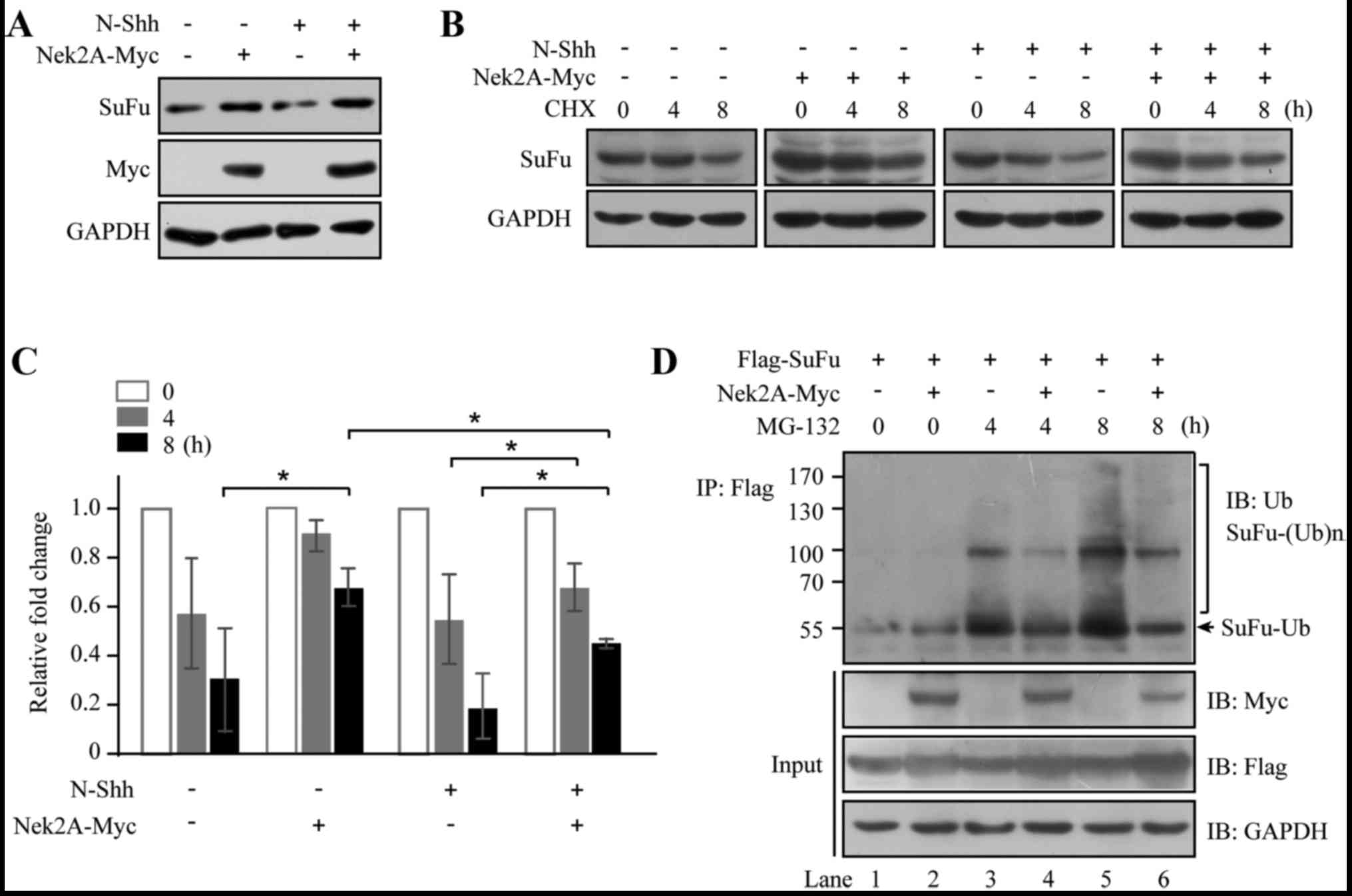
Nek2A and then stimulated with N-Shh conditioned medium. As shown
in Fig. 1A, SuFu abundance
decreased upon the treatment of N-Shh, consistent with a previous
study (15). However, Nek2A
reversed the effect of N-Shh on SuFu degradation, suggesting that
Nek2A stabilizes SuFu. Furthermore, half-life of SuFu was evaluated
in the presence of CHX, which inhibits protein synthesis (Fig. 1B). SuFu turnover was attenuated in
Nek2A-transfected cells; however, when N-Shh was added, SuFu
rapidly degraded and Nek2A reversed the stimulatory effect of N-Shh
on SuFu turnover (Fig. 1C), which
further supporting that Nek2A may stabilize SuFu. To delineate the
mechanism for the interaction of these two proteins, we treated
HEK293T cells with Nek2A alone or together with MG-132, an
inhibitor of 26S proteasome, for 4 and 8 h. Lysates were
immunoprecipitated with Flag-tagged SuFu and the precipitants were
probed for ubiquitin. As shown in Fig.
1D, lanes 1 and 2 exhibited basal ubiquitination profiles of
SuFu without or with Nek2A respectively, and the polyubiquitinated
proteins are hardly detected. Pretreatment of MG-132 in the absence
of Nek2A (lanes 3 and 5) resulted in significant accumulation of
ubiquitinated SuFu. However, Nek2A treatment diminished SuFu
ubiquitination (lanes 4 and 6) in comparison with the control.
These results indicate a role of Nek2A in SuFu regulation through
ubiquitin/proteasome pathway.
Suppression of Nek2A activates Hh
signaling pathway
Considering the critical repressive role of SuFu in
Hh signaling, we reasoned that Nek2A may regulate activation of Hh
pathway through stabilizing SuFu protein. Therefore, we generated
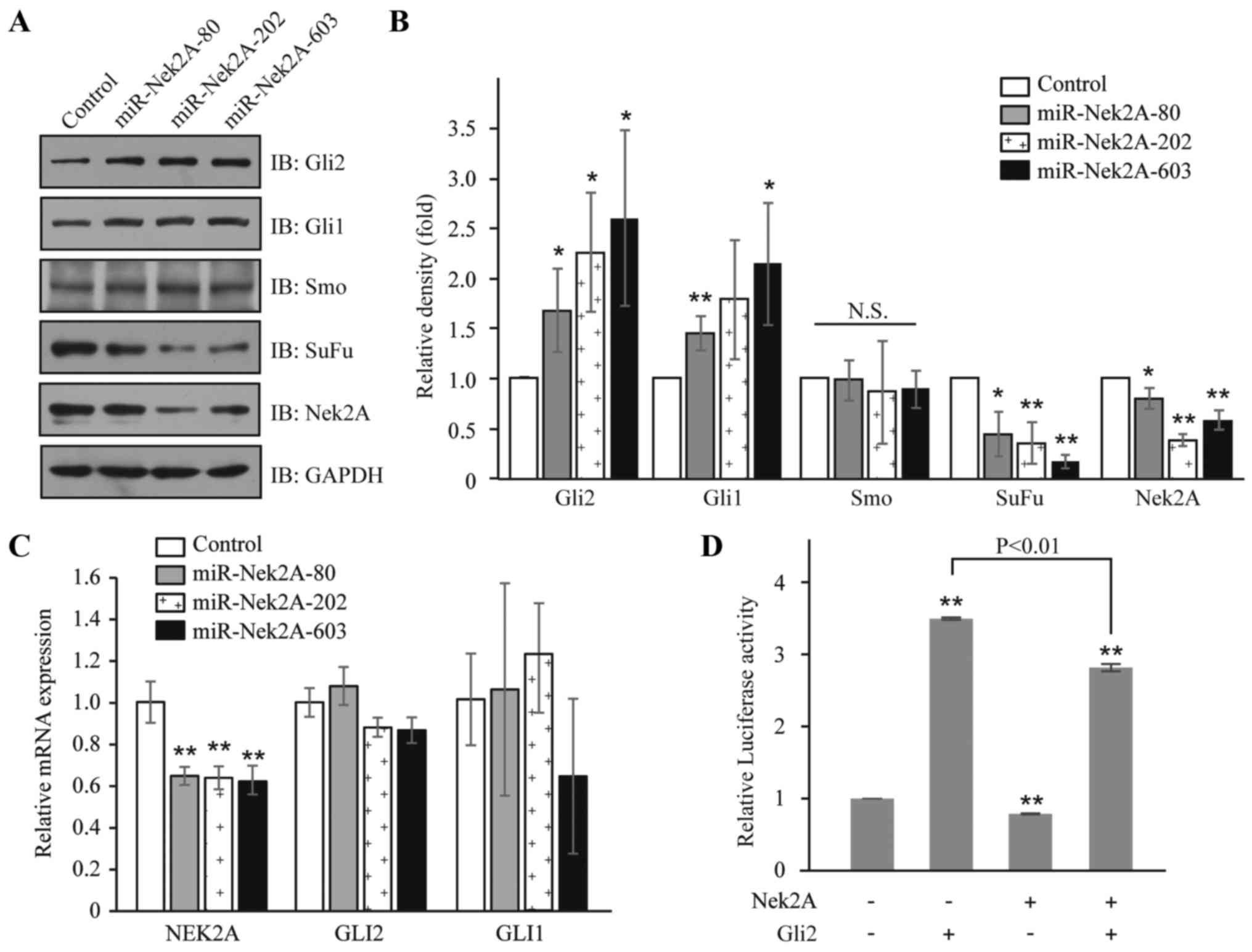
Nek2A miRNAi constructs that inhibit Nek2A expression (17). In HEK293T cells, repression of
Nek2A by miR-Nek2A-80, -202 or -603 resulted in elevated protein
levels of Gli2 and its target Gli1, but no obvious change of Smo,
which is a member of the frizzled family of seven-pass
transmembrane receptors (33)
(Figs. 2A and B). Interestingly,
upon the introduction of miR-Nek2A, the mRNA levels of Gli2 and
Gli1 had not been affected (Fig.
2C), indicating that Nek2A acts as a modulator for SuFu at
post-transcriptional level. Furthermore, to assess the effects of
Nek2A on the transcriptional activity of Hh, we transfected Nek2A
in HEK293T cells and found that in cells overexpressing Nek2A, the
transcription of Bcl-2 (a target of Hh signaling), was
significantly decreased compared to control cells with or without
Gli2 expression (Fig. 2D). These
results suggest that Nek2A negatively regulates Hh signaling.
Nek2A is regulated by Hh signaling
Nek2 has been found expressed aberrantly in multiple
cancers and emerged as a possible oncogene due to its regulatory
role in mitosis and cancer-related signaling pathways. Given the
important roles of SuFu and Nek2 in cancer and the fact that these
two proteins interact with each other, we set out to investigate
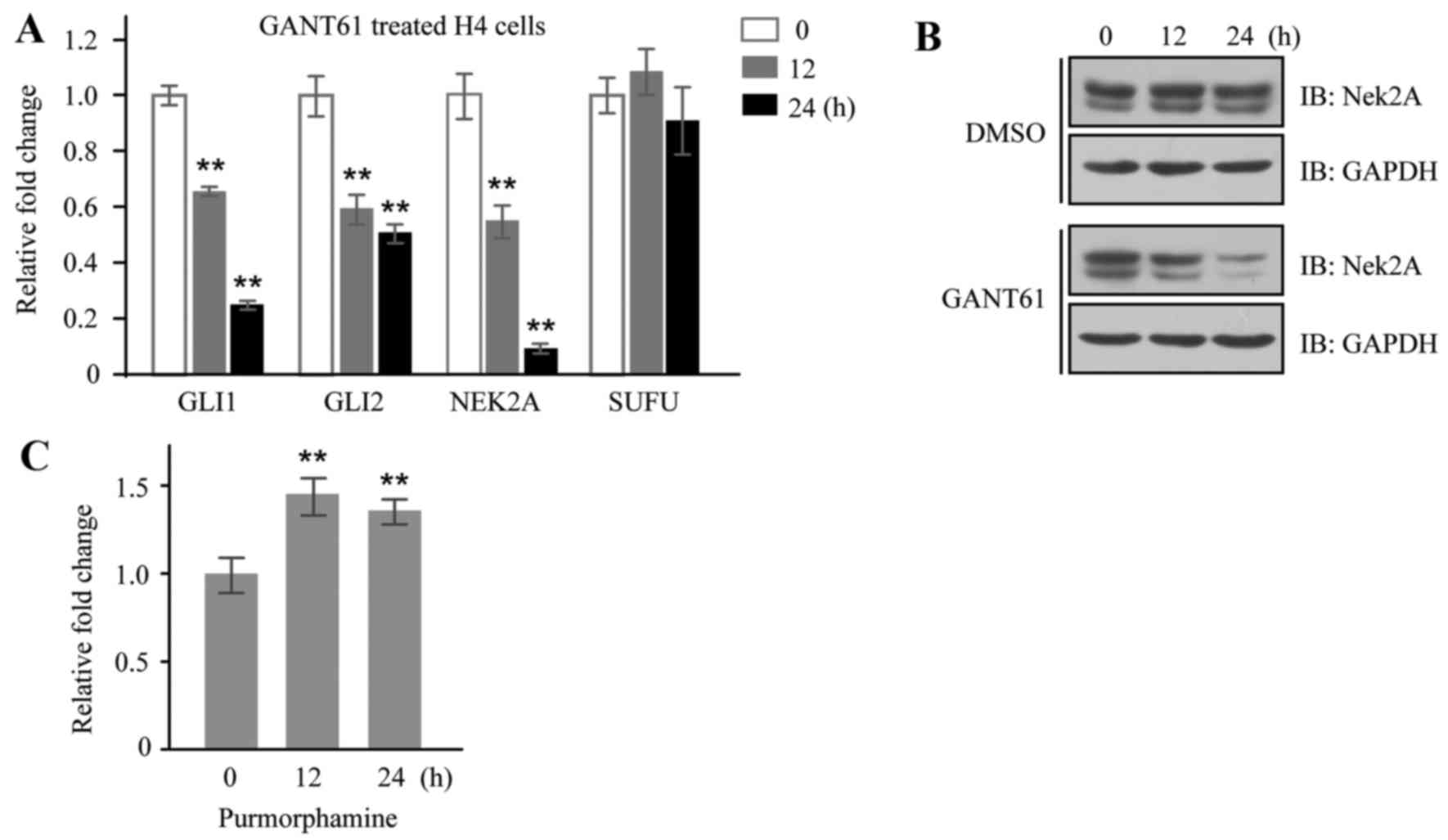
the relationship of Nek2A and Hh signaling. GANT61, an inhibitor of
Gli transcription factors, was employed to inhibit Hh signaling in
H4 cells, and mRNA expressions of GLI1 and GLI2 were downregulated
as expected; interestingly, the expression of NEK2A was also
reduced (Fig. 3A). Moreover, the
inhibitory effect of GANT61 on Nek2A expression was further
verified in H4 cells by western blotting (Fig. 3B). On the other hand, we detected
NEK2A mRNA expression in the same cells stimulated by
purmorphamine, a purine derivative found to activate the Hh pathway
by directly targeting Smo (34),
and found that mRNA level of NEK2A was significantly increased in
treated cells (Fig. 3C). The fact
that Nek2A can be regulated by Hh signaling indicates that Nek2A
may be a new target gene of Hh pathway.
NEK2A is a target gene of transcription
factor Gli
To further elucidate the upstream regulation of
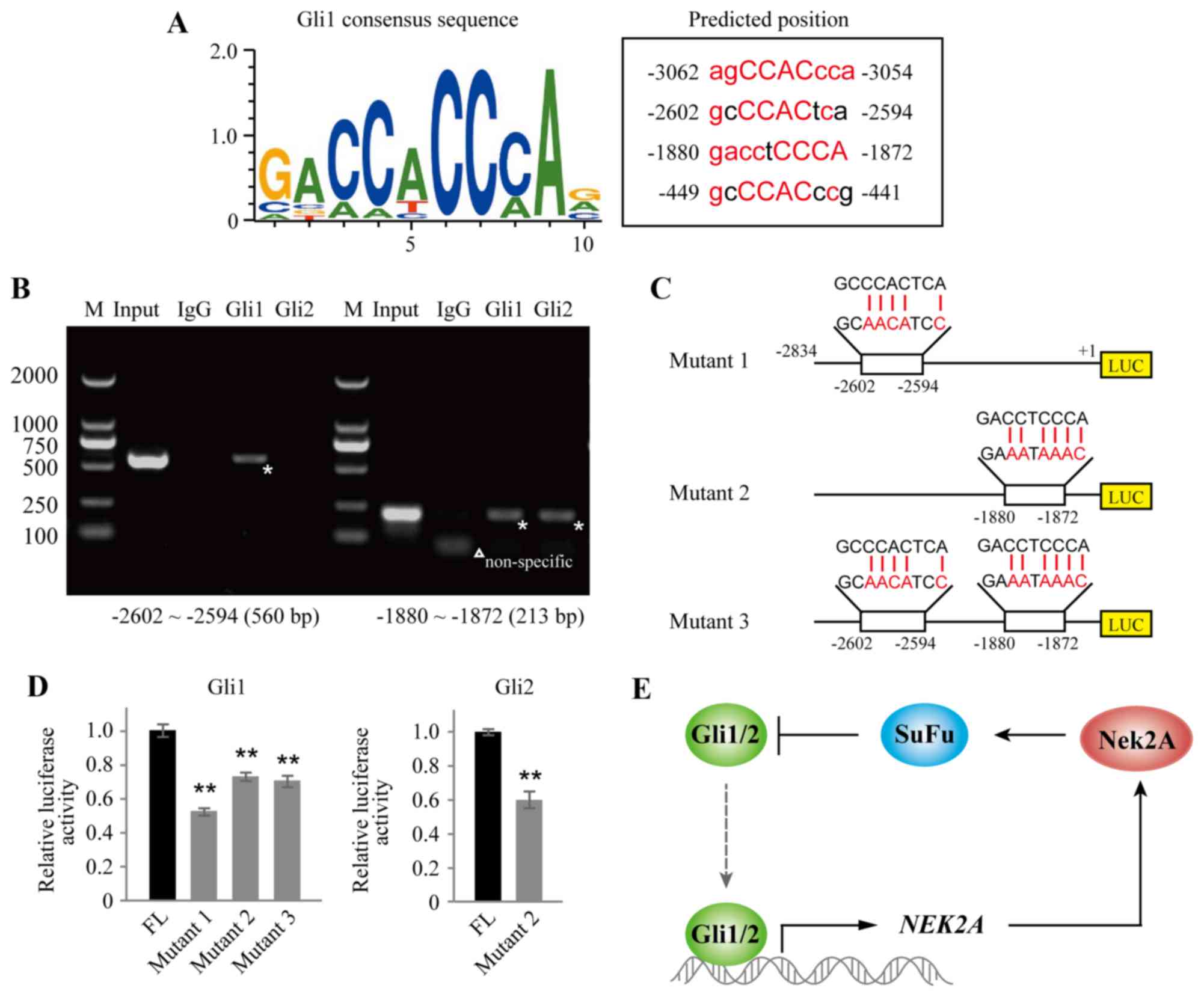
Nek2A and its relationship with Hh signaling, we used the Genomatix
software MatInspector module for in silico analysis of
putative transcription factor Gli binding sites, which were
presented within the −3500/+1 region of the human Nek2A promoter.
Four possible binding sites were found in the promoter region of
Nek2A as shown in Fig. 4A. We next
conducted a set of ChIP assays in CaES-17 cells and found that Gli1
displayed a strong binding enrichment to the specific regions:
−2602 to −2594, and −1880 to −1872, of the Nek2A promoter, whereas
Gli2 only bound to region of −1880 to −1872, which are indicated
with stars in Fig. 4B.
To verify these binding domains, we constructed
luciferase reporter vectors driven by FL or mutant Nek2A promoter
(Fig. 4C). The FL or mutants of
Nek2A (mutants 1, 2 and 3) were co-transfected with Gli1 or Gli2 in
HEK293T cells as indicated. As shown in Fig. 4D, luciferase activities of single
mutants (mutant 1 or 2) and double mutant (mutant 3) diminished
significantly when compared with FL control in Gli1 overexpressed
cells. In Gli2 transfected cells, mutant 2 also showed attenuated
luciferase activity. Together, these data support that Nek2A is the
target gene of Gli transcription factors. A working model is
proposed to illustrate the feedback regulatory loop between Nek2A
and SuFu (Fig. 4E). In brief,
Gli1/2 directly activates NEK2A. Increased Nek2A protein then
interacts with SuFu and promotes its stabilization. Stabilized SuFu
subsequently inhibits Gli1/2 entering into the nucleus and
suppresses Gli1/2 transcriptional function on NEK2A.
Discussion
The present study casts new insight into the
regulation of Hh/Gli signaling. In particular, Nek2A stabilizes
SuFu partly through the inhibition of SuFu ubiquitination;
moreover, Nek2A impedes protein expression and transcriptional
activity of Gli2. We also demonstrate that Nek2A is a target gene
of Gli transcription activity, revealing a feedback loop of Nek2A
in regulating Gli-mediated Hh signaling (Fig. 4E).
Hh signaling pathway was first identified by a
genetic screen in Drosophila for mutants affecting body
patterning in the early 1980s (35). Additional proteins have been found
to be involved in regulating the Hh pathway since then (36). Here, our study introduces Nek2A as
an important regulator of Hh signaling. Moreover, we demonstrate
that besides their role in the activation of Hh downstream
signaling, Gli1 and Gli2 both bind to the promoter region of Nek2A
gene and activate Nek2A transcription. These findings reveal a new
regulatory mechanism of Hh/SuFu/Gli signaling pathway.
Feedback loops are the building blocks of system
dynamics in biological systems. It may operate via an indirect and
integrated inhibitor/stimulator feedback loop influencing the
production of the other. Many feedback loops have been reported in
different signaling pathways and biological processes. For
instance, Hh-dependent GLI1 induction constitutes a positive
feedback loop, while Hh-dependent regulation of PTCH1, HHIP1, CDON
and BOC constitutes negative feedback loops (37,38).
We present here a novel, negative feedback loop in which Nek2A
stabilizes SuFu, generally a key inhibitory regulator in Hh
signaling, through impairing ubiquitin/proteasome degradation of
SuFu, and simultaneously requires for the activating effects of
Gli1/2, positive regulators in Hh signaling. Since the binding of
SuFu and Glis prevents Gli1/2 from turning on transcription in the
nucleus, resulting in diminished expression of Gli target genes
(e.g. Nek2A), our results uncover a negative feedback loop. Under
the normal physiology condition, constituents of Hh signaling and
Nek2A possess a homeostasis condition. When Hh signaling is
aberrantly activated by excessive Shh stimulation under
pathological condition, suppression effect of SuFu to Gli1/2 would
be diminished resulting from SuFu degradation, and we considered
this process a positive feedback regulation of Hh signaling
pathway. In order to prevent further dysfunction of the biosystem,
this positive feedback loop is supposed to be rescued with some
intrinsic mechanisms, of which stabilization effect of Nek2A on
SuFu may be one.
Many proteins are variously modified, and one
prominent intersection between post-translational modifications is
phosphorylation and ubiquitination. In our previous study (17), SuFu is demonstrated to be
stabilized by Nek2A through phosphorylation on two specific sites
of T225 and S352. In the present study, we identify a new mechanism
for SuFu regulation as Nek2A impedes ubiquitin/proteasome
proteolysis of SuFu. It is known that phosphorylation can promote
or block ubiquitination. In this study, we propose that SuFu
phosphorylation inhibits its ubiquitination, likely through the
following possible mechanisms: one modification masks the
recognition site for a second post-transcriptional modification;
phosphorylation negatively regulates the activity of the E3 ligase
responsible for ubiquitin transfer; phosphorylation influences
ubiquitination by regulating the substrate-ligase interaction at
the level of subcellular compartmentalization.
Similar to other kinases involved in spindle
assembly or duplication (39),
overexpression of Nek2 has been implicated in several neoplastic
diseases, such as breast carcinomas (40), lung adenocarcinomas (41), testicular seminomas (42) and diffuse large B cell lymphomas
(43). On the other hand, in Hh
driven cancers, the therapies of Gli inhibitors are often reported
to be related to drug resistance (44,45).
However, the underlying mechanism is unclear. Our results on the
relationship of Nek2A and Hh signaling may provide one possible
explanation for the drug resistance of Gli inhibitors. We found
that GANT61 (Gli1/2 inhibitor) inhibited the mRNA expression and
the protein synthesis of Nek2A (Fig.
3A and B). Therefore, the diminished Nek2A renders a more
fragile SuFu suspectable to degradation, and thus releases the
inhibition of Hh signaling, which may counteract the tumor
suppressive effect of Gli inhibitors. Future studies will be
required to determine precisely whether or how Nek2A may contribute
to drug-resistance in certain cancer cells. Furthermore, given that
Nek2A is a marker of cell cycle and its abundance increases through
S and G2 and reaches peak in late G2/M, we propose that Nek2A may
be a key signal indicating the events of excessive proliferation,
thus initiating the feedback regulatory process to attenuate the
aberrant cell proliferation.
In conclusion, our results identify Nek2A as both a
regulator of Hh signaling pathway through impairing
ubiquitin/proteasome proteolysis of SuFu and a target gene of
Gli1/2. Therefore, we illustrated that Nek2A fine-tunes Hh
patterning through a feedback loop, which may be a critical player
for the rigorous maintenance of cellular homeostasis.
Acknowledgments
This study was supported in part by grants from the
National Natural Science Foundation of China (no. 81560457 to Y.W.,
no. 31460305 to S.L.), the Natural Science Foundation of Jiangxi
Province (no. 20151BAB205037 to Y.W.).
References
|
1
|
Cooper AF, Yu KP, Brueckner M, Brailey LL,
Johnson L, McGrath JM and Bale AE: Cardiac and CNS defects in a
mouse with targeted disruption of suppressor of fused. Development.
132:4407–4417. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Svärd J, Heby-Henricson K, Persson-Lek M,
Rozell B, Lauth M, Bergström A, Ericson J, Toftgård R and Teglund
S: Genetic elimination of Suppressor of fused reveals an essential
repressor function in the mammalian Hedgehog signaling pathway. Dev
Cell. 10:187–197. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stone DM, Murone M, Luoh S, Ye W, Armanini
MP, Gurney A, Phillips H, Brush J, Goddard A, de Sauvage FJ, et al:
Characterization of the human suppressor of fused, a negative
regulator of the zinc-finger transcription factor Gli. J Cell Sci.
112:4437–4448. 1999.PubMed/NCBI
|
|
4
|
Barnfield PC, Zhang X, Thanabalasingham V,
Yoshida M and Hui CC: Negative regulation of Gli1 and Gli2
activator function by Suppressor of fused through multiple
mechanisms. Differentiation. 73:397–405. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kise Y, Morinaka A, Teglund S and Miki H:
Sufu recruits GSK3beta for efficient processing of Gli3. Biochem
Biophys Res Commun. 387:569–574. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cheng SY and Bishop JM: Suppressor of
Fused represses Gli-mediated transcription by recruiting the
SAP18-mSin3 corepressor complex. Proc Natl Acad Sci USA.
99:5442–5447. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu J, Heydeck W, Zeng H and Liu A: Dual
function of suppressor of fused in Hh pathway activation and mouse
spinal cord patterning. Dev Biol. 362:141–153. 2012. View Article : Google Scholar
|
|
8
|
Chen MH, Wilson CW, Li YJ, Law KK, Lu CS,
Gacayan R, Zhang X, Hui CC and Chuang PT: Cilium-independent
regulation of Gli protein function by Sufu in Hedgehog signaling is
evolutionarily conserved. Genes Dev. 23:1910–1928. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang C, Pan Y and Wang B: Suppressor of
fused and Spop regulate the stability, processing and function of
Gli2 and Gli3 full-length activators but not their repressors.
Development. 137:2001–2009. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Taylor MD, Liu L, Raffel C, Hui CC,
Mainprize TG, Zhang X, Agatep R, Chiappa S, Gao L, Lowrance A, et
al: Mutations in SUFU predispose to medulloblastoma. Nat Genet.
31:306–310. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pastorino L, Ghiorzo P, Nasti S,
Battistuzzi L, Cusano R, Marzocchi C, Garrè ML, Clementi M and
Scarrà GB: Identification of a SUFU germline mutation in a family
with Gorlin syndrome. Am J Med Genet A. 149A:1539–1543. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tostar U, Malm CJ, Meis-Kindblom JM,
Kindblom LG, Toftgård R and Undén AB: Deregulation of the hedgehog
signalling pathway: A possible role for the PTCH and SUFU genes in
human rhabdomyoma and rhabdomyosarcoma development. J Pathol.
208:17–25. 2006. View Article : Google Scholar
|
|
13
|
Sharpe HJ, Pau G, Dijkgraaf GJ,
Basset-Seguin N, Modrusan Z, Januario T, Tsui V, Durham AB, Dlugosz
AA, Haverty PM, et al: Genomic analysis of smoothened inhibitor
resistance in basal cell carcinoma. Cancer Cell. 27:327–341. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sheng T, Li C, Zhang X, Chi S, He N, Chen
K, McCormick F, Gatalica Z and Xie J: Activation of the hedgehog
pathway in advanced prostate cancer. Mol Cancer. 3:292004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yue S, Chen Y and Cheng SY: Hedgehog
signaling promotes the degradation of tumor suppressor Sufu through
the ubiquitin-proteasome pathway. Oncogene. 28:492–499. 2009.
View Article : Google Scholar
|
|
16
|
Chen Y, Yue S, Xie L, Pu XH, Jin T and
Cheng SY: Dual phosphorylation of suppressor of fused (Sufu) by PKA
and GSK3beta regulates its stability and localization in the
primary cilium. J Biol Chem. 286:13502–13511. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Y, Li Y, Hu G, Huang X, Rao H, Xiong
X, Luo Z, Lu Q and Luo S: Nek2A phosphorylates and stabilizes SuFu:
A new strategy of Gli2/Hedgehog signaling regulatory mechanism.
Cell Signal. 28:1304–1313. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hayward DG and Fry AM: Nek2 kinase in
chromosome instability and cancer. Cancer Lett. 237:155–166. 2006.
View Article : Google Scholar
|
|
19
|
Fry AM, Meraldi P and Nigg EA: A
centrosomal function for the human Nek2 protein kinase, a member of
the NIMA family of cell cycle regulators. EMBO J. 17:470–481. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mayor T, Hacker U, Stierhof YD and Nigg
EA: The mechanism regulating the dissociation of the centrosomal
protein C-Nap1 from mitotic spindle poles. J Cell Sci.
115:3275–3284. 2002.PubMed/NCBI
|
|
21
|
Fry AM, Mayor T, Meraldi P, Stierhof YD,
Tanaka K and Nigg EA: C-Nap1, a novel centrosomal coiled-coil
protein and candidate substrate of the cell cycle-regulated protein
kinase Nek2. J Cell Biol. 141:1563–1574. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bahe S, Stierhof YD, Wilkinson CJ, Leiss F
and Nigg EA: Rootletin forms centriole-associated filaments and
functions in centrosome cohesion. J Cell Biol. 171:27–33. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rapley J, Baxter JE, Blot J, Wattam SL,
Casenghi M, Meraldi P, Nigg EA and Fry AM: Coordinate regulation of
the mother centriole component nlp by nek2 and plk1 protein
kinases. Mol Cell Biol. 25:1309–1324. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takahashi Y, Iwaya T, Sawada G, Kurashige
J, Matsumura T, Uchi R, Ueo H, Takano Y, Eguchi H, Sudo T, et al:
Up-regulation of NEK2 by microRNA-128 methylation is associated
with poor prognosis in colorectal cancer. Ann Surg Oncol.
21:205–212. 2014. View Article : Google Scholar
|
|
25
|
Neal CP, Fry AM, Moreman C, McGregor A,
Garcea G, Berry DP and Manson MM: Overexpression of the Nek2 kinase
in colorectal cancer correlates with beta-catenin relocalization
and shortened cancer-specific survival. J Surg Oncol. 110:828–838.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cappello P, Blaser H, Gorrini C, Lin DC,
Elia AJ, Wakeham A, Haider S, Boutros PC, Mason JM, Miller NA, et
al: Role of Nek2 on centrosome duplication and aneuploidy in breast
cancer cells. Oncogene. 33:2375–2384. 2014. View Article : Google Scholar
|
|
27
|
Hu CM, Zhu J, Guo XE, Chen W, Qiu XL, Ngo
B, Chien R, Wang YV, Tsai CY, Wu G, et al: Novel small molecules
disrupting Hec1/Nek2 interaction ablate tumor progression by
triggering Nek2 degradation through a death-trap mechanism.
Oncogene. 34:1220–1230. 2015. View Article : Google Scholar
|
|
28
|
Liu X, Gao Y, Lu Y, Zhang J, Li L and Yin
F: Upregulation of NEK2 is associated with drug resistance in
ovarian cancer. Oncol Rep. 31:745–754. 2014.
|
|
29
|
Zhong X, Guan X, Liu W and Zhang L:
Aberrant expression of NEK2 and its clinical significance in
non-small cell lung cancer. Oncol Lett. 8:1470–1476.
2014.PubMed/NCBI
|
|
30
|
Stricker TP, Henriksen KJ, Tonsgard JH,
Montag AG, Krausz TN and Pytel P: Expression profiling of 519
kinase genes in matched malignant peripheral nerve sheath
tumor/plexiform neurofibroma samples is discriminatory and
identifies mitotic regulators BUB1B, PBK and NEK2 as overexpressed
with transformation. Mod Pathol. 26:930–943. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luo SW, Zhang C, Zhang B, Kim CH, Qiu YZ,
Du QS, Mei L and Xiong WC: Regulation of heterochromatin
remodelling and myogenin expression during muscle differentiation
by FAK interaction with MBD2. EMBO J. 28:2568–2582. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Joshi H, Nord SH, Frigessi A,
Børresen-Dale AL and Kristensen VN: Overrepresentation of
transcription factor families in the genesets underlying breast
cancer subtypes. BMC Genomics. 13:1992012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen Y and Struhl G: In vivo evidence that
Patched and Smoothened constitute distinct binding and transducing
components of a Hedgehog receptor complex. Development.
125:4943–4948. 1998.PubMed/NCBI
|
|
34
|
Sinha S and Chen JK: Purmorphamine
activates the Hedgehog pathway by targeting Smoothened. Nat Chem
Biol. 2:29–30. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nüsslein-Volhard C and Wieschaus E:
Mutations affecting segment number and polarity in Drosophila.
Nature. 287:795–801. 1980. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Huangfu D and Anderson KV: Signaling from
Smo to Ci/Gli: Conservation and divergence of Hedgehog pathways
from Drosophila to vertebrates. Development. 133:3–14. 2006.
View Article : Google Scholar
|
|
37
|
Holtz AM, Peterson KA, Nishi Y, Morin S,
Song JY, Charron F, McMahon AP and Allen BL: Essential role for
ligand-dependent feedback antagonism of vertebrate hedgehog
signaling by PTCH1, PTCH2 and HHIP1 during neural patterning.
Development. 140:3423–3434. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tenzen T, Allen BL, Cole F, Kang JS,
Krauss RS and McMahon AP: The cell surface membrane proteins Cdo
and Boc are components and targets of the Hedgehog signaling
pathway and feedback network in mice. Dev Cell. 10:647–656. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fukasawa K: Oncogenes and tumour
suppressors take on centrosomes. Nat Rev Cancer. 7:911–924. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hayward DG, Clarke RB, Faragher AJ, Pillai
MR, Hagan IM and Fry AM: The centrosomal kinase Nek2 displays
elevated levels of protein expression in human breast cancer.
Cancer Res. 64:7370–7376. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Landi MT, Dracheva T, Rotunno M, Figueroa
JD, Liu H, Dasgupta A, Mann FE, Fukuoka J, Hames M, Bergen AW, et
al: Gene expression signature of cigarette smoking and its role in
lung adenocarcinoma development and survival. PLoS One.
3:e16512008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Barbagallo F, Paronetto MP, Franco R,
Chieffi P, Dolci S, Fry AM, Geremia R and Sette C: Increased
expression and nuclear localization of the centrosomal kinase Nek2
in human testicular seminomas. J Pathol. 217:431–441. 2009.
View Article : Google Scholar
|
|
43
|
Andréasson U, Dictor M, Jerkeman M,
Berglund M, Sundström C, Linderoth J, Rosenquist R, Borrebaeck CA
and Ek S: Identification of molecular targets associated with
transformed diffuse large B cell lymphoma using highly purified
tumor cells. Am J Hematol. 84:803–808. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kobune M, Takimoto R, Murase K, Iyama S,
Sato T, Kikuchi S, Kawano Y, Miyanishi K, Sato Y, Niitsu Y, et al:
Drug resistance is dramatically restored by hedgehog inhibitors in
CD34+ leukemic cells. Cancer Sci. 100:948–955. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Amable L, Fain J, Gavin E and Reed E: Gli1
contributes to cellular resistance to cisplatin through altered
cellular accumulation of the drug. Oncol Rep. 32:469–474.
2014.PubMed/NCBI
|