The human genome project has demonstrated that
<2% nucleic acid sequences are protein-coding. In 2002, Okazaki
et al (1) first identified
a class of longer transcripts by sequencing a mouse cDNA library,
and termed them long non-coding RNAs (lncRNAs). Due to limitations
in our understanding of gene regulation and expression, these
non-coding RNAs were considered to be transcriptional noise until
HOX transcript antisense RNA (HOTAIR) was identified. HOTAIR is a
lncRNA located on the antisense strand of the HOXC gene cluster,
which regulates the expression of the HOXD gene and the occurrence
of H3K4me2 methylation modifications in certain gene regions, and
silences expression at this site (2,3).
Although lncRNAs do not encode proteins, they participate in
important cellular physiological processes. Numerous studies have
reported that lncRNAs are abnormally expressed in various types of
tumor, and affect the occurrence and development of tumors through
different signaling pathways. lncRNAs are generally defined as
long-stranded RNAs with >200 nucleotides, but this is not
absolute. For example, BC1 and snaR are <200 nucleotides in
length (4). lncRNAs may also be
classified according to location, object of action and functional
mechanism (5). The progress in
lncRNA research is associated with the progress in research
technology. It is worth mentioning that RNA-sequencing technology,
proposed by the Snyder team at Yale University, has low background
noise and is able to detect single-base differences between similar
genes or different transcripts caused by variable cleavage in the
gene family, as well as low abundance transcripts (6). At present, next-generation sequencing
technologies have provided the largest experimental evidence on
lncRNA, with the advantages of high flux, short duration, high
accuracy and abundant information.
lncRNAs participate in the regulation of various
biological activities, including genomic imprinting, X-chromosome
inactivation, chromosome modification and telomere biology. They
are also associated with the occurrence and development of various
human diseases, particularly with the occurrence and development of
malignant tumors. Their long ribbon-like molecular structure
interacts with multiple types of molecules, thus affecting cell
biological activities. Signaling pathways participate in the
transduction of cell signals through a series of enzymatic
reactions, which are regulated by various types of molecules.
lncRNA is an important regulator of signaling pathways, and
regulates gene expression at a transcriptional,
post-transcriptional and epigenetic level. This review focused on
the function of lncRNA in several classical signaling pathways.
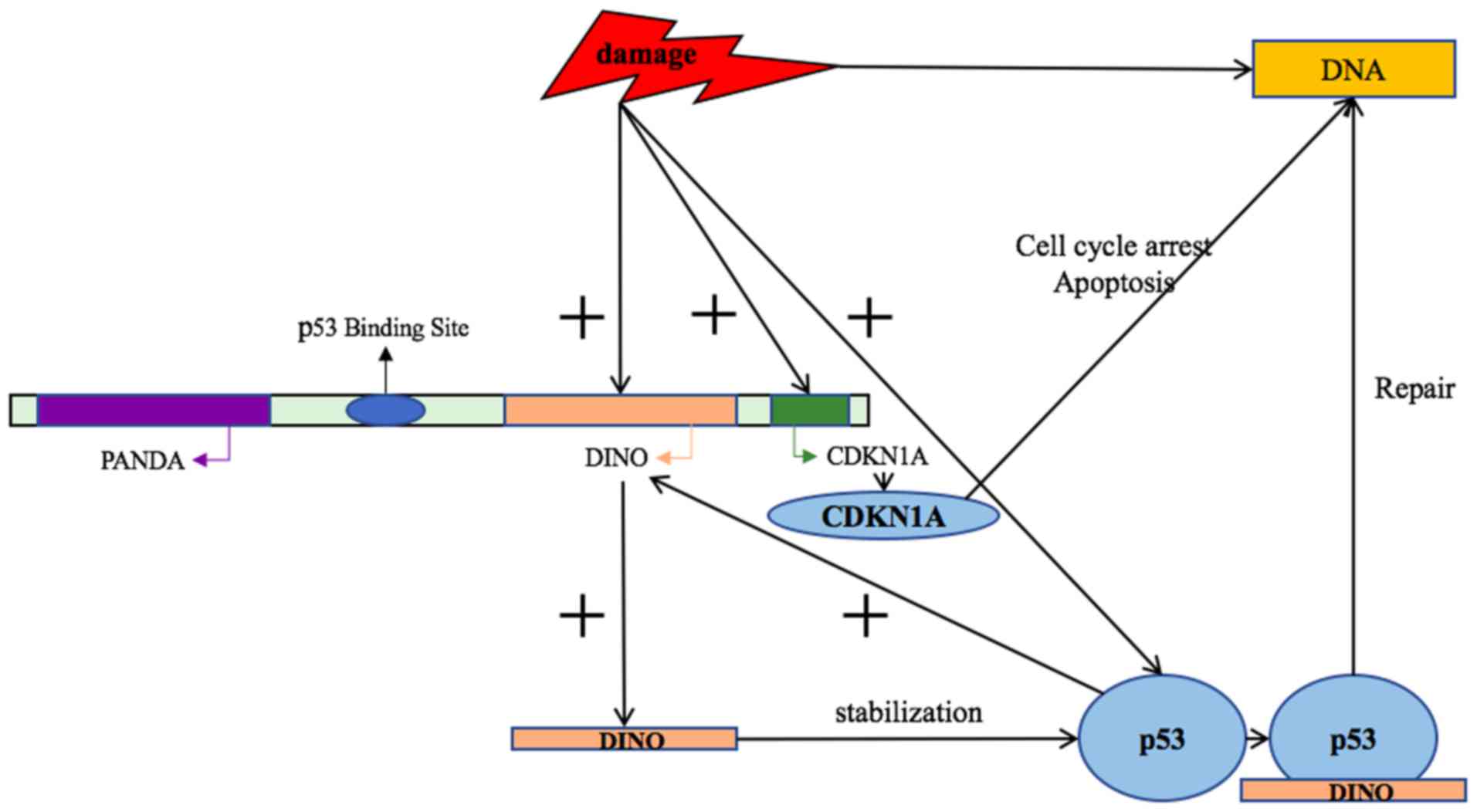
The damage induced noncoding (lncRNA-DINO) is a DNA
damage-activated transcriptional lncRNA involved in the
lncRNA-directed biological DNA damage response. Numerous abnormal
changes in the p53 pathway may increase the probability of
tumorigenesis. DINO interacts with the p53 protein to promote its
stability, which causes protein accumulation, activation of p53
target proteins and regulates a p53 auto-enhancement loop (Fig. 1). DINO is involved in p53-dependent
gene expression, cell cycle arrest and apoptosis. In the absence of
DNA damage, artificially upregulating DINO expression can
effectively activate the DNA damage response pathway and cell cycle
arrest. However, when artificially inhibiting the expression of
DINO, the response of cells to p53 signaling is weakened. In mice,
inactivation of the DINO gene or the promoter blocks the p53
pathway and improves acute radiation syndrome in vivo
(8). Thus, DNA damage-induced
lncRNAs form a feedback loop with homologous transcription factors
to enhance cell signaling networks. A genome-wide transcriptome
analysis of human diploid fibroblasts revealed that lncRNA
metastasis associated lung adenocarcinoma transcript 1 (MALAT1) is
involved in regulating the expression of cell cycle-associated
genes, and is required for G1/S and mitotic progression. Low
expression of MALAT1 leads to the activation of p53 and its target
genes, and cell cycle defects observed in MALAT1-depleted cells are
sensitive to p53 expression levels, indicating that p53 is a key
downstream mediator of MALAT1. Furthermore, reduced expression of
an oncogenic transcription factor, B-MYB, which is involved in G2/M
progression, is expressed in MALAT1-depleted cells due to altered
binding of splicing factors on B-MYB pre-mRNA and aberrant
alternative splicing (9). These
findings reveal the mechanism by which lncRNA regulates cell
proliferation.
lncRNAs also regulate p53, subsequently affecting
cell cycle progression, for example via lncRNA regulator of
reprogramming (lncRNA-ROR). Unlike MDM2, which leads to p53
degradation through the ubiquitin-proteasome pathway, it directly
inhibits p53 through interactions with heterogeneous nuclear
ribonucleoprotein I (hnRNP I) at the translational level. ROR
carries an hnRNP I binding domain that acts on the 5'-untranslated
region of p53 mRNA to suppress the translation of p53, thereby
inhibiting p53-mediated cell cycle arrest and apoptosis. The
induction of p53 following treatment with doxorubicin increases ROR
expression levels, and the ectopic expression of p53 induces ROR
production, resulting in a ROR-p53 auto-regulatory feedback loop
(10). Similarly to mRNA, the
major transcriptional processes of lncRNA are regulated by various
factors, including p53, NF-κB, Sox2, Oct4 and Nanog (11). For example, the transcriptional
promoter region of ROR is directly affected by pluripotency
factors, OCT4, SOX2 and KLF4 (12). lncRNA-p21 binding to hnRNP-K
assists hnRNP-K to localize and inhibit p53-regulated genes, and is
a p53-dependent transcriptional response inhibitor (13). Competing endogenous RNAs (ceRNAs)
regulate gene expression through competitive binding of microRNA.
By providing ceRNA or natural microRNA sponge-like functions,
lncRNAs are important post-transcriptional regulators of gene
expression, regulating microRNAs and competitively inhibiting
important receptors involved in various cellular biological
activities (14). For example,
lncRNA-loc285194, growth-arrest specific 5 (GAS5) and HOTAIR appear
to function as competitive endogenous RNAs participating in the
inhibition of tumor development (15-17).
GAS5 also has the ability to simulate DNA and compete with steroid
receptors in cells, which regulates steroid-mediated
transcriptional regulation, growth arrest and apoptosis (18). In human meningioma cells, lncRNA
maternally expressed gene 3 (MEG3) stimulates p53-mediated
transactivation and inhibits tumor proliferation by inducing
apoptosis (19), which may be
associated with its ability to downregulate MDM2 expression as
demonstrated by Zhou et al (20). Certain natural antisense
transcripts are important lncRNAs that may be paired with
complementary RNAs to form sense-antisense double-stranded RNAs.
This causes the degradation or translational inhibition of target
mRNA. For example, lncRNA-Wrap53 is a natural antisense transcript
of p53. The highly conserved Wrap53 has been demonstrated to
regulate endogenous p53 mRNA levels and further induce p53 protein
expression by targeting the 5'-untranslated region of p53 mRNA
(21). p21-associated lncRNA
DNA-damage activated (PANDA) is an lncRNA located ~5 kB upstream of
the CDKN1A TSS gene, and is induced by specific DNA damage. PANDA
is also a p53-associated regulatory lncRNA involved in cell cycle
progression and apoptosis (22).
The ectopic expression of lncRNA is involved in numerous disease
processes, but the underlying molecular mechanism is not yet fully
understood. Table I lists the
lncRNAs that interact with the p53 pathway in various types of
human cancer.
NF-κB is a widely expressed pleiotropic
transcription factor. It is a heterotrimer composed of p50, p65 and
IκB. The NF-κB signaling pathway is activated by a variety of
extracellular stimuli, serving a central role in transcriptional
regulation, and in the expression and regulation of various genes.
It is a marker of cell activation, and mediates a variety of
biological processes, including inflammation, cell proliferation
and tumor metastasis. The NF-κB pathway is regulated by various
lncRNAs.
lncRNA-Lethe was the first lncRNA identified to be
involved in regulation of the NF-κB signaling pathway. In human 293
cells, Lethe is selectively induced by pro-inflammatory cytokines
through the NF-κB pathway, and acts on RelA (the p65 subunit of
NF-κB) to inhibit RelA-DNA binding and subsequently target gene
activation (23). Lethe has an
active role in regulating the NF-κB pathway and is involved in
inflammation through a typical negative feedback mechanism.
Activated NF-κB promotes the expression of Lethe, and Lethe
directly inhibits this pathway by interacting with RelA, thereby
reducing the production of various inflammatory proteins (23). A novel lncRNA, namely nuclear
transcription factor NF-κB interacting lncRNA (NKILA), which
participates in the inhibition of breast cancer metastasis through
the NF-κB signaling pathway, was discovered by analyzing a large
number of lncRNAs in human breast cancer (24). NF-κB upregulates the expression of
NKILA by binding to the promoter of NKILA, and a large quantity of
NKILA is able to bind to NF-κB/IκB to form a stable complex, thus
directly masking the phosphorylation site of IκB to prevent
IKK-mediated phosphorylation of IκB and activation of NF-κB. The
negative feedback of NKILA and NF-κB serves a key role in
preventing overactivation of the NF-κB pathway in
inflammation-stimulated mammary epithelial cells. However, highly
expressed microRNA-103/107 mediates the degradation of NKILA, which
results in overactivation of the NF-κB pathway, thus contributing
to cancer metastasis and poor patient outcome (24). As a key enzyme in prostaglandin
synthesis, COX-2 serves an important role in the prognosis of
tumors, particularly neoangiogenesis and tumor progression
(25,26). lncRNA p50-associated COX-2
extragenic RNA (PACER), localized in the nucleus, binds directly to
free p50, inhibits the formation of homodimers, promotes the
formation of heterodimers, increases the transcription enhancement
effect and increases the expression of COX-2 (27). Pearson et al (28) achieved results, which support these
observations, whereby primary human osteoarthritic chondrocytes
treated with IL-1β rapidly induced the formation of PACER,
indicating that PACER expression is associated with COX2. In
addition to PACER, the lncRNA cyclooxygenase 2 (COX2) locus near
the COX2 gene is essential for the transcription of
NF-κB-associated late inflammatory genes in endotoxin-treated mouse
macrophages (29). In the nucleus,
MALAT1 interacts with the P65-p50 heterodimer to inhibit downstream
TNF/ IL-6 expression. Bioinformatics analysis has revealed that
MALAT1 affects NF-κB/RelA activity in the epithelial-mesenchymal
transition process (30). The
abnormal expression of HOTAIR in ovarian cancer induces NF-κB
activation by inhibiting Iκ-Bα in the DNA damage response. It also
increases the expression of MMP-9 and IL-6, key target genes of
NF-κB, and serves an important role in DNA damage response, cell
senescence and chemotherapy resistance (31). In addition, certain lncRNAs
regulate signaling pathways indirectly through signaling elements
or associated upstream molecules in the NF-κB pathway. For example,
MIR31HG is frequently deleted in glioblastoma, and significantly
reduces the levels of micro (mi)RNA-31, while loss of miRNA-31
expression enhances NF-κB signaling by targeting TRADD, its
upstream activator, and promotes tumor growth (32). C2dat1 promotes neural cell survival
by enhancing CAMK2D expression and activating the NF-κB pathway
(33). Expression of IL1β-eRNA,
IL1β-RBT46, AS-IL1α, ANRIL, THRIL and IL7R is induced by the NF-κB
signaling pathway, and regulates the expression of NF-κB-associated
target genes (34-38). Table
II lists the lncRNAs that interact with the NF-κB pathway in
cancer.
The Wnt gene was first named Int1, because its
activation is dependent on the insertion of the mouse breast
cancer-associated virus gene (39). Since then, a large number of
studies have reported that Int1 serves an important role in the
normal embryonic development of mice and is equivalent to the
Wingless gene of drosophila. Therefore, combining Wingless with
Int1 it was named Wnt. The Wnt signaling pathway is an
evolutionarily highly conserved signaling system involved in
important physiological processes, including embryonic development,
tissue differentiation and cell homeostasis (40,41).
Specific lncRNAs associated with the Wnt signaling pathway are
discussed below.
Numerous lncRNAs target and affect the accumulation
of β-catenin, a key molecule that acts on the Wnt pathway, thereby
regulating the expression of Wnt target genes and the function of
cancer cells (42,43). Numerous lncRNAs regulate target
genes to promote tumorigenesis and development. In breast cancer,
patients exhibiting high CCAT2 expression have a significantly
poorer prognosis and lower overall survival rate compared with
patients with low CCAT2 expression (44). A previous study on MCF-7 and
MDA-MB-231 breast cancer cells demonstrated that the suppression of
CCAT2 expression decreases the levels of β-catenin in the cytoplasm
and nucleus, and reduces the expression of CCND1 and c-myc, which
are classic downstream genes of the Wnt/β-catenin signaling
pathway. These results indicate the possible role of CCAT2 in
breast cancer (44). The abnormal
expression of HOTAIR has been demonstrated in certain types of
human cancer. In colorectal cancer, Wnt/0205-catenin signaling is
inhibited by HOTAIR-knockdown and miR-203a-3p overexpression
(45). The downregulation of
HOTAIR expression is essential for reducing cell proliferation and
chemoresistance via modulation of miR-203a-3p expression levels and
the activity of the Wnt/0205-catenin signaling pathway (45). lncRNA-CRNDE is significantly
overexpressed in renal cell carcinoma. Elevated CRNDE expression
may regulate the PI3K/Akt/GSK3b signaling pathway, increase the
level of β-catenin in the nucleus, and ultimately regulate the
expression of the Wnt target gene to regulate the proliferation of
renal cancer cells (46).
lncRNA-TCF7 promotes the self-renewal and proliferation of
hepatocellular carcinoma cells by activating the Wnt signaling
pathway. It may be that TCF7 recruits SWI/SNF complexes to act on
its own promoter to regulate expression and activate the Wnt
pathway (47). Similarly, numerous
studies have reported that lncRNAs inhibit tumorigenesis. In a
large number of non-small cell lung cancer tissue samples, low
expression of AK126698 often predicts larger tumor diameter and
advanced tumor stage. Conversely, an increase in AK126698
expression targets Frizzled-8, one of the receptors acting of the
Wnt/β-catenin signaling pathway, inhibiting the proliferation and
metastasis of cancer cells and inducing apoptosis (48). The expression of lncRNA-CTD903 is
significantly upregulated in human colorectal cancer and may be
used as an independent prognostic factor for colorectal cancer.
When CTD903 was knocked down in the RKO and SW480 cell lines, the
invasion and metastasis of the tumor cells is increased, and
epithelial-mesenchymal transition-like characteristics are
exhibited (49). Further results
demonstrated that downregulated CTD903 expression allows the
Wnt/β-catenin signaling pathway to be more easily activated,
increasing the expression of transcription factors, Twist and
Snail, as well as the protein expression of the mesenchymal marker,
Vimentin, and reducing that of the epithelial marker, ZO-1
(49). Chemotherapy resistance can
result in poor prognosis. Furthermore, lncRNAs serve a role in cell
resistance. The level of lncRNA-Meg3 is significantly lower in the
cisplatin-resistant lung cancer cell line, A549/DDP, compared with
that in the A549 cell line (50).
Further results demonstrated that the upregulation of Meg3
expression in vitro increases the sensitivity of the
A549/DDP lung cancer cell line to cisplatin. However, the
sensitivity of A549 cells to cisplatin decreased following RNA
interference downregulation of Meg3 expression. Researchers
hypothesized that Meg3 mediates increased cell cycle arrest and
apoptosis by regulating p53, β-catenin and survivin, resulting in
increased sensitivity to chemotherapy (50). In osteosarcoma, HOTTIP induces
tumor occurrence and resistance by activating the Wnt signaling
pathway, which may be a potential target for the treatment of
osteosarcoma (51). Table III lists the lncRNAs that
interact with the Wnt signaling pathway and their general roles in
cancer.
The notch signaling pathway is a highly
evolutionarily conserved cell signaling pathway. It mediates direct
cell-cell contact and affects the normal morphogenesis of cells. It
regulates differentiation of multipotent progenitor cells,
apoptosis, cell proliferation and the formation of cell borders
(52). lncRNA also interacts with
the Notch signaling pathway and affects tumor progression.
It has been reported that the non-coding sequences
that accompany the genes are often involved in the regulation of
peripheral genes. In the human genome, lncRNA-NALT was identified
at a distance of ~100 bases from the NOTCH1 gene. In T-cell acute
lymphoblastic leukemia cells, increased expression of NALT
significantly promotes cell proliferation. It may be that NALT
activates transcription in the Notch pathway and regulatory
elements in the nucleus (53).
lncRNA-SNHG12 expression is significantly elevated in osteosarcoma
tissues and cell lines, and predicts poor prognosis. A study of the
biological function of SNHG12 in the 143B and U2OS cell lines
revealed that the downregulation of SNHG12 inhibits cell
proliferation by blocking the G0/G1 phase of the cell cycle, and
reduces the potential for cell invasion and metastasis. SNHG12
sponges miR-195-5p in osteosarcoma cells, similarly to
endogenously-competent RNA, to regulate Notch2 gene expression,
thereby affecting signal transduction in the Notch pathway
(54). In a study of CRC
specimens, and matched adjacent normal tissues and CRC cell lines,
the expression of FOXD2-AS1 was significantly increased in CRC
tissues as well as in CRC cell lines, and downregulation of
FOXD2-AS1 expression suppressed proliferation, invasion and
migration in vitro. Further study examined associated
markers of the epithelial-mesenchymal transition and Notch
signaling pathways, and confirmed that the two pathways are
inactivated in CRC cells following FOXD2-AS1-knockdown (55). When studying the expression profile
of lncRNA-MEG3 and the two Notch pathway-associated molecules,
Notch1 and Hes1, in human endometrial tissues and cell lines, Guo
et al (56) reported that,
compared with the normal control group, MEG3 expression is
significantly downregulated in cancer tissue. However, the
expression levels of Notch1 and Hes1 are significantly increased.
Upregulation of MEG3 in tumor cells in vivo inhibits the
growth of tumors by inhibiting the Notch pathway (56). The Notch pathway serves an
important role in maintaining the pluripotency of glioma stem
cells. The activation of Notch1 induces lncRNA-TUG1 expression in
specific glioma cell lines. In the cytoplasm, TUG1 promotes cell
self-renewal by decoying miR-145. In the nucleus, TUG1 binds YY1 by
recruiting multi-comb complexes to allow site-specific H3K27
histones to methylate, thereby inhibiting gene mutation. In
addition, intravenous administration of TSG1 with antisense
oligonucleotides induces the differentiation of glioma stem cell
lines in vivo, and effectively inhibits their proliferation
(57). Table IV lists the lncRNAs that interact
with the Notch pathway in cancer.
AKT, also known as protein kinase B, is an important
crossover molecule in multiple signaling pathways and is activated
by various substances, including hormones, growth factors,
cytokines and intercellular matrix. The PI3K/AKT signaling pathway
is involved in important physiological activities, including cell
proliferation and survival, and is associated with cancer,
diabetes, rheumatoid arthritis and other diseases (58,59).
In recent years, the regulatory mechanism of lncRNAs in the
PI3K/AKT pathway and the impact on diseases, have gradually been
revealed.
The MAPK pathway is a classical signaling pathway,
and is involved in the regulation of cell proliferation,
differentiation, transformation and apoptosis. The MAPK pathway is
associated with the occurrence of various diseases, including
inflammation and tumors, by phosphorylating nuclear transcription
factors, cytoskeletal proteins, and enzymes. MAPK signaling
primarily includes four pathways: ERK, JNK/SAPK, P38 and ERK5/BMK1.
The ERK, JNK, P38 and ERK5/BMK1 pathways can be activated by
different stimuli, and there is wide crosstalk between these
pathways, resulting in mutual synergy or inhibition between them.
As important regulatory genes, numerous lncRNAs have been
identified to regulate the MAPK pathway.
An important role of the SNHG12-miR-181a-MAPK/Slug
axis was described in multidrug resistance in non-small cell lung
cancer cells. In non-small cell lung cancer cells, silencing of
lncRNA-SNHG12 upregulates miR-181a expression, and inhibits MAPK1
and MAP2K1 expression. This is accompanied by decreased
phosphorylation of MAPK1 and MAP2K1, and decreased Slug expression
levels. Ultimately, lncRNA-SNHG12 inhibits the activation of the
MAPK/Slug pathway and increases the drug sensitivity of tumor cells
(71). Similarly, lncRNA-XIST acts
via microRNA to influence the activation of pathways. XIST
regulates MAPK1 by targeting miR-194-5p-like ceRNA and reducing its
expression. Silenced XIST inhibits the proliferation of hepatoma
cells and reduces their invasiveness (72). In previous in vitro and
in vivo studies of gallbladder carcinoma, the knockdown of
MALAT1 was demonstrated to reduce the phosphorylation of MEK1/2,
ERK1/2, MAPK and JNK 1/2/3, resulting in significant inhibition of
the ERK/MAPK pathway. The metastasis and invasiveness of cancer
cells is also weakened simultaneously (73). In a cecal ligation and puncture
(CLP) rat model, the expression levels of p38, MAPK and NF-κB
protein were significantly higher in the CLP group compared with
that in the sham group (74).
Overexpression of MALAT1 alone significantly increases the levels
of p38/MAPK and NF-κB. An elaborative study demonstrated that
MALAT1 mediates the development of heart failure and the
inflammatory response through the activation of p38/MAPK/NF-κB.
Furthermore, in the inflammatory response to sepsis, p38/MAPK/NF-κB
may be downstream of MALAT1 signaling (74). In neuro-genic hyperplasia of
neuroblastoma-derived Neuro-2a cells, silencing of MALAT1 results
in significant activation of the MAPK pathway, and abnormal
activation of the peroxisome proliferator-activated receptor and
the p53 pathway (75). Thus, the
same lncRNA appears to exhibit different effects in different types
of tumors. When lncRNA-BANCR is deleted in lung cancer cells, the
proliferation and metastasis of cancer cells is increased. In
addition, when the expression of BANCR is increased, the
proliferation of lung cancer cells is inhibited, acting as a tumor
suppressor gene (76). In in
vivo and in vitro experiments on malignant melanoma,
tumor growth is inhibited following BANCR-knockout. The underlying
mechanism may involve the silencing of BANCR making it difficult to
activate the MAPK pathway, and to inhibit important components of
the pathway, ERK1/2 and JNK (77).
Table VI lists the lncRNAs that
interact with the MAPK pathway in cancer.
The regulatory roles of lncRNAs are diverse and
complex, and the current understanding of them is limited. The
ENCODE research project identified <10,000 lncRNAs in the human
genome (78), but only ~100
lncRNAs are known to be biologically functional. This is not only
due to limited research techniques, but also the nature of lncRNAs.
The intracellular abundance of lncRNA is typically low, and the
function between different species is not conserved. For example,
the human HOTAIR gene has a transregulatory effect on the HOXD
gene, but this function is not observed in mice (79). Previous studies have demonstrated
that lncRNAs primarily act as signal transduction molecules,
decoys, transcription factors and scaffolds, interacting with
proteins, DNA, RNA or other molecules to participate in
transcriptional regulation, post-transcriptional regulation and
epigenetic regulation of genes. The transcriptional regulation and
posttranscriptional regulation of lncRNAs have been detailed, but
their regulatory role in epigenetics is also an important function,
and lncRNA-Xist is an interesting example (80,81).
lncRNA-Xist is able to extensively bind to the X-chromosome through
an interaction with polycomb repressive complex 2, leading to the
inactivation of the X-chromosome (82,83).
Recently, the novel silencing effect of Xist on the X-chromosome
identified by Chen et al (84) has been debated. In the experiments
by Chen et al (84), it was
demonstrated that Xist interacts with Lamin B Receptor (LBR), a
nuclear inner membrane protein involved in the establishment of the
three-dimensional structures of chromosomes by regulating the
anchoring of chromatin in the nucleus. It was hypothesized that the
combination of Xist and LBR triggers alterations to the
three-dimensional structure of the X-chromosome and thus
participates in the silencing of the X-chromosome. This hypothesis
was validated by constructing stable cell lines, LBS-Xist and
LBS-Xist-BoxB, associated with Xist and LBR (84). However, a study by Wang et
al questioned these findings. According to the analysis of the
original sequencing data, it was considered that the
Xist-associated stable cell lines, LBS-Xist and LBS-Xist-BoxB, were
unsuccessfully constructed, and the sequencing depth and data
normalization were also problematic (85). In response, the original research
team provided more detailed evidence to confirm the results of
their own research (86). Similar
disputes reflect that, although lncRNAs have been recognized as
having important biological functions, our understanding remains
far from thorough. However, rational academic controversy makes
lncRNA-associated research more rigorous.
This grant was supported by National Science
Foundation Committee of China (grant no. 81502482 and 81672474),
Zhejiang Provincial National Science Foundation Committee of China
(grant no. LQ14H160017) and Zhejiang Provincial research projects
of medical and healthy industries (grant nos. 2015KYB026, 2017KY193
and 2017KY210).
Not applicable.
YS, JYZ and HHC consulted and preliminarily
summarized the literature required for the manuscript. WS was the
major contributor in writing the manuscript. ZFW contributed to the
writing of the manuscript. DSH and JGZ approved the final version
of the manuscript. All authors read and approved the final
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
|
1
|
Okazaki Y, Furuno M, Kasukawa T, Adachi J,
Bono H, Kondo S, Nikaido I, Osato N, Saito R, Suzuki H, et al:
Analysis of the mouse transcriptome based on functional annotation
of 60,770 full-length cDNAs. Nature. 420:563–573. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rinn JL, Kertesz M, Wang JK, Squazzo SL,
Xu X, Brugmann SA, Goodnough LH, Helms JA, Farnham PJ, Segal E, et
al: Functional demarcation of active and silent chromatin domains
in human HOX loci by noncoding RNAs. Cell. 129:1311–1323. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsai MC, Manor O, Wan Y, Mosammaparast N,
Wang JK, Lan F, Shi Y, Segal E and Chang HY: Long noncoding RNA as
modular scaffold of histone modification complexes. Science.
329:689–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Amaral PP, Clark MB, Gascoigne DK, Dinger
ME and Mattick JS: lncRNAdb: A reference database for long
noncoding RNAs. Nucleic Acids Res. 39(Suppl 1): D146–D151. 2011.
View Article : Google Scholar :
|
|
5
|
St Laurent G, Wahlestedt C and Kapranov P:
The Landscape of long noncoding RNA classification. Trends Genet.
31:239–251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Z, Gerstein M and Snyder M: RNA-Seq:
A revolutionary tool for transcriptomics. Nat Rev Genet. 10:57–63.
2009. View
Article : Google Scholar
|
|
7
|
Lane DP and Crawford LV: T antigen is
bound to a host protein in SV40-transformed cells. Nature.
278:261–263. 1979. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schmitt AM, Garcia JT, Hung T, Flynn RA,
Shen Y, Qu K, Payumo AY, Peres-da-Silva A, Broz DK, Baum R, et al:
An inducible long noncoding RNA amplifies DNA damage signaling. Nat
Genet. 48:1370–1376. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tripathi V, Shen Z, Chakraborty A, Giri S,
Freier SM, Wu X, Zhang Y, Gorospe M, Prasanth SG, Lal A, et al:
Long noncoding RNA MALAT1 controls cell cycle progression by
regulating the expression of oncogenic transcription factor B-MYB.
PLoS Genet. 9:e10033682013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang A, Zhou N, Huang J, Liu Q, Fukuda K,
Ma D, Lu Z, Bai C, Watabe K and Mo YY: The human long non-coding
RNA-RoR is a p53 repressor in response to DNA damage. Cell Res.
23:340–350. 2013. View Article : Google Scholar :
|
|
11
|
Guttman M, Amit I, Garber M, French C, Lin
MF, Feldser D, Huarte M, Zuk O, Carey BW, Cassady JP, et al:
Chromatin signature reveals over a thousand highly conserved large
non-coding RNAs in mammals. Nature. 458:223–227. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Loewer S, Cabili MN, Guttman M, Loh YH,
Thomas K, Park IH, Garber M, Curran M, Onder T, Agarwal S, et al:
Large intergenic non-coding RNA-RoR modulates reprogramming of
human induced pluripotent stem cells. Nat Genet. 42:1113–1117.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huarte M, Guttman M, Feldser D, Garber M,
Koziol MJ, Kenzelmann-Broz D, Khalil AM, Zuk O, Amit I, Rabani M,
et al: A large intergenic noncoding RNA induced by p53 mediates
global gene repression in the p53 response. Cell. 142:409–419.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu Q, Huang J, Zhou N, Zhang Z, Zhang A,
Lu Z, Wu F and Mo YY: LncRNA loc285194 is a p53-regulated tumor
suppressor. Nucleic Acids Res. 41:4976–4987. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang XF, Ye Y and Zhao SJ: LncRNA Gas5
acts as a ceRNA to regulate PTEN expression by sponging miR-222-3p
in papillary thyroid carcinoma. Oncotarget. 9:3519–3530. 2017.
|
|
17
|
Cai H, Yao J, An Y, Chen X, Chen W, Wu D,
Luo B, Yang Y, Jiang Y, Sun D, et al: LncRNA HOTAIR acts a
competing endogenous RNA to control the expression of notch3 via
sponging miR-613 in pancreatic cancer. Oncotarget. 8:32905–32917.
2017.PubMed/NCBI
|
|
18
|
Hudson WH, Pickard MR, de Vera IM, Kuiper
EG, Mourtada-Maarabouni M, Conn GL, Kojetin DJ, Williams GT and
Ortlund EA: Conserved sequence-specific lincRNA-steroid receptor
interactions drive transcriptional repression and direct cell fate.
Nat Commun. 5:53952014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang X, Gejman R, Mahta A, Zhong Y, Rice
KA, Zhou Y, Cheunsuchon P, Louis DN and Klibanski A: Maternally
expressed gene 3, an imprinted noncoding RNA gene, is associated
with meningioma pathogenesis and progression. Cancer Res.
70:2350–2358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou Y, Zhong Y, Wang Y, Zhang X, Batista
DL, Gejman R, Ansell PJ, Zhao J, Weng C and Klibanski A: Activation
of p53 by MEG3 non-coding RNA. J Biol Chem. 282:24731–24742. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mahmoudi S, Henriksson S, Corcoran M,
Méndez-Vidal C, Wiman KG and Farnebo M: Wrap53, a natural p53
antisense transcript required for p53 induction upon DNA damage.
Mol Cell. 33:462–471. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hung T, Wang Y, Lin MF, Koegel AK, Kotake
Y, Grant GD, Horlings HM, Shah N, Umbricht C, Wang P, et al:
Extensive and coordinated transcription of noncoding RNAs within
cell-cycle promoters. Nat Genet. 43:621–629. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rapicavoli NA, Qu K, Zhang J, Mikhail M,
Laberge RM and Chang HY: A mammalian pseudogene lncRNA at the
interface of inflammation and anti-inflammatory therapeutics.
eLife. 2:e007622013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu B, Sun L, Liu Q, Gong C, Yao Y, Lv X,
Lin L, Yao H, Su F, Li D, et al: A cytoplasmic NF-κB interacting
long noncoding RNA blocks IκB phosphorylation and suppresses breast
cancer metastasis. Cancer Cell. 27:370–381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rozic JG, Chakraborty C and Lala PK:
Cyclooxygenase inhibitors retard murine mammary tumor progression
by reducing tumor cell migration, invasiveness and angiogenesis.
Int J Cancer. 93:497–506. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Subbaramaiah K and Dannenberg AJ:
Cyclooxygenase 2: A molecular target for cancer prevention and
treatment. Trends Pharmacol Sci. 24:96–102. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Krawczyk M and Emerson BM: p50-associated
COX-2 extragenic RNA (PACER) activates COX-2 gene expression by
occluding repressive NF-κB complexes. eLife. 3:e017762014.
View Article : Google Scholar
|
|
28
|
Pearson MJ, Philp AM, Heward JA, Roux BT,
Walsh DA, Davis ET, Lindsay MA and Jones SW: Long intergenic
noncoding RNAs mediate the human chondrocyte inflammatory response
and are differentially expressed in osteoarthritis cartilage.
Arthritis Rheumatol. 68:845–856. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu G, Gong A-Y, Wang Y, Ma S, Chen X, Chen
J, Su CJ, Shibata A, Strauss-Soukup JK, Drescher KM, et al:
LincRNA-Cox2 promotes late inflammatory gene transcription in
macrophages through modulating SWI/SNF-mediated chromatin
remodeling. J Immunol. 196:2799–2808. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gutschner T, Hämmerle M and Diederichs S:
MALAT1 - a paradigm for long noncoding RNA function in cancer. J
Mol Med (Berl). 91:791–801. 2013. View Article : Google Scholar
|
|
31
|
Özeş AR, Miller DF, Özeş ON, Fang F, Liu
Y, Matei D, Huang T and Nephew KP: NF-κB-HOTAIR axis links DNA
damage response, chemoresistance and cellular senescence in ovarian
cancer. Oncogene. 35:5350–5361. 2016. View Article : Google Scholar
|
|
32
|
Rajbhandari R, McFarland BC, Patel A,
Gerigk M, Gray GK, Fehling SC, Bredel M, Berbari NF, Kim H, Marks
MP, et al: Loss of tumor suppressive microRNA-31 enhances
TRADD/NF-κB signaling in glioblastoma. Oncotarget. 6:17805–17816.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu Q, Deng F, Xing Z, Wu Z, Cen B, Xu S,
Zhao Z, Nepomuceno R, Bhuiyan MI, Sun D, et al: Long non-coding RNA
C2dat1 regulates CaMKIIδ expression to promote neuronal survival
through the NF-κB signaling pathway following cerebral ischemia.
Cell Death Dis. 7:e21732016. View Article : Google Scholar
|
|
34
|
IIott NE, Heward JA, Roux B, Tsitsiou E,
Fenwick PS, Lenzi L, Goodhead I, Hertz-Fowler C, Heger A, Hall N,
et al: Long non-coding RNAs and enhancer RNAs regulate the
lipopolysac-charide-induced inflammatory response in human
monocytes. Nat Commun. 5:39792014. View Article : Google Scholar
|
|
35
|
Chan J, Atianand M, Jiang Z, Carpenter S,
Aiello D, Elling R, Fitzgerald KA and Caffrey DR: Cutting edge: A
natural antisense transcript, AS-IL1α, controls inducible
transcription of the proin-flammatory cytokine IL-1α. J Immunol.
195:1359–1363. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou X, Han X, Wittfeldt A, Sun J, Liu C,
Wang X, Gan LM, Cao H and Liang Z: Long non-coding RNA ANRIL
regulates inflammatory responses as a novel component of NF-κB
pathway. RNA Biol. 13:98–108. 2016. View Article : Google Scholar
|
|
37
|
Li Z, Chao TC, Chang KY, Lin N, Patil VS,
Shimizu C, Head SR, Burns JC and Rana TM: The long noncoding RNA
THRIL regulates TNFα expression through its interaction with
hnRNPL. Proc Natl Acad Sci USA. 111:1002–1007. 2014. View Article : Google Scholar
|
|
38
|
Cui H, Xie N, Tan Z, Banerjee S,
Thannickal VJ, Abraham E and Liu G: The human long noncoding RNA
lnc-IL7R regulates the inflammatory response. Eur J Immunol.
44:2085–2095. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nusse R and Varmus HE: Many tumors induced
by the mouse mammary tumor virus contain a provirus integrated in
the same region of the host genome. Cell. 31:99–109. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Miki T, Yasuda SY and Kahn M:
Wnt/β-catenin signaling in embryonic stem cell self-renewal and
somatic cell reprogramming. Stem Cell Rev. 7:836–846. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Serio RN: Wnt of the two horizons: Putting
stem cell self-renewal and cell fate determination into context.
Stem Cells Dev. 23:1975–1990. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ma Y, Yang Y, Wang F, Moyer MP, Wei Q,
Zhang P, Yang Z, Liu W, Zhang H, Chen N, et al: Long non-coding RNA
CCAL regulates colorectal cancer progression by activating
Wnt/β-catenin signalling pathway via suppression of activator
protein 2α. Gut. 65:1494–1504. 2015. View Article : Google Scholar
|
|
43
|
Fan Y, Shen B, Tan M, Mu X, Qin Y, Zhang F
and Liu Y: Long non-coding RNA UCA1 increases chemoresistance of
bladder cancer cells by regulating Wnt signaling. FEBS J.
281:1750–1758. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cai Y, He J and Zhang D: Long noncoding
RNA CCAT2 promotes breast tumor growth by regulating the Wnt
signaling pathway. Onco Targets Ther. 8:2657–2664. 2015.PubMed/NCBI
|
|
45
|
Xiao Z, Qu Z, Chen Z, Fang Z, Zhou K,
Huang Z, Guo X and Zhang Y: LncRNA HOTAIR is a prognostic biomarker
for the proliferation and chemoresistance of colorectal cancer via
MiR-203a-3p-mediated Wnt/β-catenin signaling pathway. Cell Physiol
Biochem. 46:1275–1285. 2018. View Article : Google Scholar
|
|
46
|
Shao K, Shi T, Yang Y, Wang X, Xu D and
Zhou P: Highly expressed lncRNA CRNDE promotes cell proliferation
through Wnt/β-catenin signaling in renal cell carcinoma. Tumour
Biol. 37:15997–16004. 2016. View Article : Google Scholar
|
|
47
|
Wang Y, He L, Du Y, Zhu P, Huang G, Luo J,
Yan X, Ye B, Li C, Xia P, et al: The long noncoding RNA lncTCF7
promotes self-renewal of human liver cancer stem cells through
activation of Wnt signaling. Cell Stem Cell. 16:413–425. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fu X, Li H, Liu C, Hu B, Li T and Wang Y:
Long noncoding RNA AK126698 inhibits proliferation and migration of
non-small cell lung cancer cells by targeting Frizzled-8 and
suppressing Wnt/β-catenin signaling pathway. OncoTargets Ther.
9:3815–3827. 2016. View Article : Google Scholar
|
|
49
|
Yuan Z, Yu X, Ni B, Chen D, Yang Z, Huang
J, Wang J, Chen D and Wang L: Overexpression of long non-coding
RNA-CTD903 inhibits colorectal cancer invasion and migration by
repressing Wnt/β-catenin signaling and predicts favorable
prognosis. Int J Oncol. 48:2675–2685. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Xia Y, He Z, Liu B, Wang P and Chen Y:
Downregulation of Meg3 enhances cisplatin resistance of lung cancer
cells through activation of the WNT/β-catenin signaling pathway.
Mol Med Rep. 12:4530–4537. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li Z, Zhao L and Wang Q: Overexpression of
long non-coding RNA HOTTIP increases chemoresistance of
osteosarcoma cell by activating the Wnt/β-catenin pathway. Am J
Transl Res. 8:2385–2393. 2016.
|
|
52
|
High FA and Epstein JA: The multifaceted
role of Notch in cardiac development and disease. Nat Rev Genet.
9:49–61. 2008. View Article : Google Scholar
|
|
53
|
Wang Y, Wu P, Lin R, Rong L, Xue Y and
Fang Y: LncRNA NALT interaction with NOTCH1 promoted cell
proliferation in pediatric T cell acute lymphoblastic leukemia. Sci
Rep. 5:137492015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhou S, Yu L, Xiong M and Dai G: LncRNA
SNHG12 promotes tumorigenesis and metastasis in osteosarcoma by
upregu-lating Notch2 by sponging miR-195-5p. Biochem Biophys Res
Commun. 495:1822–1832. 2018. View Article : Google Scholar
|
|
55
|
Yang X, Duan B and Zhou X: Long non-coding
RNA FOXD2-AS1 functions as a tumor promoter in colorectal cancer by
regulating EMT and Notch signaling pathway. Eur Rev Med Pharmacol
Sci. 21:3586–3591. 2017.PubMed/NCBI
|
|
56
|
Guo Q, Qian Z, Yan D, Li L and Huang L:
LncRNA-MEG3 inhibits cell proliferation of endometrial carcinoma by
repressing Notch signaling. Biomed Pharmacother. 82:589–594. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Katsushima K, Natsume A, Ohka F, Shinjo K,
Hatanaka A, Ichimura N, Sato S, Takahashi S, Kimura H, Totoki Y, et
al: Targeting the Notch-regulated non-coding RNA TUG1 for glioma
treatment. Nat Commun. 7:136162016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Osaki M, Oshimura M and Ito H: PI3K-Akt
pathway: Its functions and alterations in human cancer. Apoptosis.
9:667–676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Bellacosa A, Kumar CC, Di Cristofano A and
Testa JR: Activation of AKT kinases in cancer: Implications for
therapeutic targeting. Adv Cancer Res. 94:29–86. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Koirala P, Huang J, Ho TT, Wu F, Ding X
and Mo YY: LncRNA AK023948 is a positive regulator of AKT. Nat
Commun. 8:144222017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cheng Z, Guo J, Chen L, Luo N, Yang W and
Qu X: A long noncoding RNA AB073614 promotes tumorigenesis and
predicts poor prognosis in ovarian cancer. Oncotarget.
6:25381–25389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hu L, Lv QL, Chen SH, Sun B, Qu Q, Cheng
L, Guo Y, Zhou HH and Fan L: Up-regulation of long non-coding RNA
AB073614 predicts a poor prognosis in patients with glioma. Int J
Environ Res Public Health. 13:4332016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li J, Wang YM and Song YL: Knockdown of
long noncoding RNA AB073614 inhibits glioma cell proliferation and
migration via affecting epithelial-mesenchymal transition. Eur Rev
Med Pharmacol Sci. 20:3997–4002. 2016.PubMed/NCBI
|
|
64
|
Wang Y, Kuang H, Xue J, Liao L, Yin F and
Zhou X: LncRNA AB073614 regulates proliferation and metastasis of
colorectal cancer cells via the PI3K/AKT signaling pathway. Biomed
Pharmacother. 93:1230–1237. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Song W, Mei JZ and Zhang M: lncRNA
PlncRNA-1 promotes colorectal cancer cell progression by regulating
PI3K/Akt signaling pathway. Oncol Res. 26:261–268. 2018. View Article : Google Scholar
|
|
66
|
Peng W, Wang Z and Fan H: lncRNA NEAT1
impacts cell proliferation and apoptosis of colorectal cancer via
regulation of Akt signaling. Pathol Oncol Res. 23:651–656. 2017.
View Article : Google Scholar
|
|
67
|
Li C, Liang G, Yang S, Sui J, Yao W, Shen
X, Zhang Y, Peng H, Hong W, Xu S, et al: Dysregulated lncRNA-UCA1
contributes to the progression of gastric cancer through regulation
of the PI3K-Akt-mTOR signaling pathway. Oncotarget. 8:93476–93491.
2017.PubMed/NCBI
|
|
68
|
Xu S, Sui S, Zhang J, Bai N, Shi Q, Zhang
G, Gao S, You Z, Zhan C, Liu F, et al: Downregulation of long
noncoding RNA MALAT1 induces epithelial-to-mesenchymal transition
via the PI3K-AKT pathway in breast cancer. Int J Clin Exp Pathol.
8:4881–4891. 2015.PubMed/NCBI
|
|
69
|
Jin Y, Feng SJ, Qiu S, Shao N and Zheng
JH: LncRNA MALAT1 promotes proliferation and metastasis in
epithelial ovarian cancer via the PI3K-AKT pathway. Eur Rev Med
Pharmacol Sci. 21:3176–3184. 2017.PubMed/NCBI
|
|
70
|
Yang W, Li X, Qi S, Li X, Zhou K, Qing S,
Zhang Y and Gao MQ: lncRNA H19 is involved in TGF-β1-induced
epithelial to mesenchymal transition in bovine epithelial cells
through PI3K/AKT Signaling Pathway. PeerJ. 5:e39502017. View Article : Google Scholar
|
|
71
|
Wang P, Chen D, Ma H and Li Y: LncRNA
SNHG12 contributes to multidrug resistance through activating the
MAPK/Slug pathway by sponging miR-181a in non-small cell lung
cancer. Oncotarget. 8:84086–84101. 2017.PubMed/NCBI
|
|
72
|
Kong Q, Zhang S, Liang C, Zhang Y, Kong Q,
Chen S, Qin J and Jin Y: lncRNA XIST functions as a molecular
sponge of miR-194-5p to regulate MAPK1 expression in hepatocellular
carcinoma cell. J Cell Biochem. 119:4458–4468. 2018. View Article : Google Scholar
|
|
73
|
Wu XS, Wang XA, Wu WG, Hu YP, Li ML, Ding
Q, Weng H, Shu YJ, Liu TY, Jiang L, et al: MALAT1 promotes the
proliferation and metastasis of gallbladder cancer cells by
activating the ERK/MAPK pathway. Cancer Biol Ther. 15:806–814.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Chen H, Wang X, Yan X, Cheng X, He X and
Zheng W: LncRNA MALAT1 regulates sepsis-induced cardiac
inflammation and dysfunction via interaction with miR-125b and p38
MAPK/NFκB. Int Immunopharmacol. 55:69–76. 2018. View Article : Google Scholar
|
|
75
|
Chen L, Feng P, Zhu X, He S, Duan J and
Zhou D: Long non-coding RNA Malat1 promotes neurite outgrowth
through activation of ERK/MAPK signalling pathway in N2a cells. J
Cell Mol Med. 20:2102–2110. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Jiang W, Zhang D, Xu B, Wu Z, Liu S, Zhang
L, Tian Y, Han X and Tian D: Long non-coding RNA BANCR promotes
proliferation and migration of lung carcinoma via MAPK pathways.
Biomed Pharmacother. 69:90–95. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Li R, Zhang L, Jia L, Duan Y, Li Y, Bao L
and Sha N: Long non-coding RNA BANCR promotes proliferation in
malignant melanoma by regulating MAPK pathway activation. PLoS One.
9:e1008932014. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Rosenbloom KR, Dreszer TR, Long JC,
Malladi VS, Sloan CA, Raney BJ, Cline MS, Karolchik D, Barber GP,
Clawson H, et al: ENCODE whole-genome data in the UCSC Genome
Browser: Update 2012. Nucleic Acids Res. 40:D912–D917. 2012.
View Article : Google Scholar :
|
|
79
|
Schorderet P and Duboule D: Structural and
functional differences in the long non-coding RNA hotair in mouse
and human. PLoS Genet. 7:e10020712011. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Brown CJ, Ballabio A, Rupert JL,
Lafreniere RG, Grompe M, Tonlorenzi R and Willard HF: A gene from
the region of the human X inactivation centre is expressed
exclusively from the inactive X chromosome. Nature. 349:38–44.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Brown CJ, Hendrich BD, Rupert JL,
Lafrenière RG, Xing Y, Lawrence J and Willard HF: The human XIST
gene: Analysis of a 17 kb inactive X-specific RNA that contains
conserved repeats and is highly localized within the nucleus. Cell.
71:527–542. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Engreitz JM, Pandya-Jones A, McDonel P,
Shishkin A, Sirokman K, Surka C, Kadri S, Xing J, Goren A, Lander
ES, et al: The Xist lncRNA exploits three-dimensional genome
architecture to spread across the X chromosome. Science.
341:12379732013. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Zhao J, Sun BK, Erwin JA, Song J-J and Lee
JT: Polycomb proteins targeted by a short repeat RNA to the mouse X
chromosome. Science. 322:750–756. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Chen CK, Blanco M, Jackson C, Aznauryan E,
Ollikainen N, Surka C, Chow A, Cerase A, McDonel P and Guttman M:
Xist recruits the X chromosome to the nuclear lamina to enable
chromosome-wide silencing. Science. 354:468–472. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang CY, Froberg JE, Blum R, Jeon Y and
Lee JT: Comment on ‘Xist recruits the X chromosome to the nuclear
lamina to enable chromosome-wide silencing’. Science. 356:3562017.
View Article : Google Scholar
|
|
86
|
Chen CK, Chow A, Lai M and Guttman M:
Response to Comment on ‘Xist recruits the X chromosome to the
nuclear lamina to enable chromosome-wide silencing’. Science.
356:54392017. View Article : Google Scholar
|
|
87
|
Suemori H and Noguchi S: Hox C cluster
genes are dispensable for overall body plan of mouse embryonic
development. Dev Biol. 220:333–342. 2000. View Article : Google Scholar
|
|
88
|
Amândio AR, Necsulea A, Joye E, Mascrez B
and Duboule D: Hotair is dispensible for mouse development. PLoS
Genet. 12:e10062322016. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Lai KM, Gong G, Atanasio A, Rojas J,
Quispe J, Posca J, White D, Huang M, Fedorova D, Grant C, et al:
Diverse phenotypes and specific transcription patterns in twenty
mouse lines with ablated LincRNAs. PLoS One. 10:e01255222015.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Tano K, Onoguchi-Mizutani R, Yeasmin F,
Uchiumi F, Suzuki Y, Yada T and Akimitsu N: Identification of
Minimal p53 Promoter Region Regulated by MALAT1 in Human Lung
Adenocarcinoma Cells. Front Genet. 8:2082018. View Article : Google Scholar :
|
|
91
|
Yang P, Yang Y, An W, Xu J, Zhang G, Jie J
and Zhang Q: The long noncoding RNA-ROR promotes the resistance of
radiotherapy for human colorectal cancer cells by targeting the
p53/miR-145 pathway. J Gastroenterol Hepatol. 32:837–845. 2017.
View Article : Google Scholar
|
|
92
|
Chen RP, Huang ZL, Liu LX, Xiang MQ, Li
GP, Feng JL, Liu B and Wu LF: Involvement of endoplasmic reticulum
stress and p53 in lncRNA MEG3-induced human hepatoma HepG2 cell
apoptosis. Oncol Rep. 36:1649–1657. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Li J, Bian EB, He XJ, Ma CC, Zong G, Wang
HL and Zhao B: Epigenetic repression of long non-coding RNA MEG3
mediated by DNMT1 represses the p53 pathway in gliomas. Int J
Oncol. 48:723–733. 2016. View Article : Google Scholar
|
|
94
|
Lu KH, Li W, Liu XH, Sun M, Zhang ML, Wu
WQ, Xie WP and Hou YY: Long non-coding RNA MEG3 inhibits NSCLC
cells proliferation and induces apoptosis by affecting p53
expression. BMC Cancer. 13:4612013. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Zhu J, Liu S, Ye F, Shen Y, Tie Y, Zhu J,
Wei L, Jin Y, Fu H, Wu Y, et al: Long noncoding RNA MEG3 interacts
with p53 protein and regulates partial p53 target genes in hepatoma
cells. PLoS One. 10:e01397902015. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Sun L, Li Y and Yang B: Downregulated long
non-coding RNA MEG3 in breast cancer regulates proliferation,
migration and invasion by depending on p53's transcriptional
activity. Biochem Biophys Res Commun. 478:323–329. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Zhang EB, Yin DD, Sun M, Kong R, Liu XH,
You LH, Han L, Xia R, Wang KM, Yang JS, et al: P53-regulated long
non-coding RNA TUG1 affects cell proliferation in human non-small
cell lung cancer, partly through epigenetically regulating HOXB7
expression. Cell Death Dis. 5:e12432014. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Huang J, Zhou N, Watabe K, Lu Z, Wu F, Xu
M and Mo YY: Long non-coding RNA UCA1 promotes breast tumor growth
by suppression of p27 (Kip1). Cell Death Dis. 5:e10082014.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Zhai N, Xia Y, Yin R, Liu J and Gao F: A
negative regulation loop of long noncoding RNA HOTAIR and p53 in
non-small-cell lung cancer. OncoTargets Ther. 9:5713–5720. 2016.
View Article : Google Scholar
|
|
100
|
Su P, Wang F, Qi B, Wang T and Zhang S:
p53 regulation-association long non-coding RNA (LncRNA PRAL)
inhibits cell proliferation by regulation of P53 in human lung
cancer. Med Sci Monit. 23:1751–1758. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Gong Z, Zhang S, Zeng Z, Wu H, Yang Q,
Xiong F, Shi L, Yang J, Zhang W, Zhou Y, et al: LOC401317, a
p53-regulated long non-coding RNA, inhibits cell proliferation and
induces apoptosis in the nasopharyngeal carcinoma cell line HNE2.
PLoS One. 9:e1106742014. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Zhai W, Li X, Wu S, Zhang Y, Pang H and
Chen W: Microarray expression profile of lncRNAs and the
upregulated ASLNC04080 lncRNA in human endometrial carcinoma. Int J
Oncol. 46:2125–2137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Thorenoor N, Faltejskova-Vychytilova P,
Hombach S, Mlcochova J, Kretz M, Svoboda M and Slaby O: Long
non-coding RNA ZFAS1 interacts with CDK1 and is involved in
p53-dependent cell cycle control and apoptosis in colorectal
cancer. Oncotarget. 7:622–637. 2016. View Article : Google Scholar :
|
|
104
|
Li P, Zhang X, Wang L, Du L, Yang Y, Liu
T, Li C and Wang C: lncRNA HOTAIR Contributes to 5FU Resistance
through Suppressing miR-218 and Activating NF-κB/TS Signaling in
Colorectal Cancer. Mol Ther Nucleic Acids. 8:356–369. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Liao Z, Zhao J and Yang Y: Downregulation
of lncRNA H19 inhibits the migration and invasion of melanoma cells
by inactivating the NF κB and PI3K/Akt signaling pathways. Mol Med
Rep. 17:7313–7318. 2018.PubMed/NCBI
|
|
106
|
Yang T, Li S, Liu J, Yin D, Yang X and
Tang Q: lncRNA-NKILA/ NF-κB feedback loop modulates laryngeal
cancer cell proliferation, invasion, and radioresistance. Cancer
Med. 7:2048–2063. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Bian D, Gao C, Bao K and Song G: The long
non-coding RNA NKILA inhibits the invasion-metastasis cascade of
malignant melanoma via the regulation of NF-ĸB. Am J Cancer Res.
7:28–40. 2017.
|
|
108
|
Huang W, Cui X, Chen J, Feng Y, Song E, Li
J and Liu Y: Long non-coding RNA NKILA inhibits migration and
invasion of tongue squamous cell carcinoma cells via suppressing
epithelial-mesenchymal transition. Oncotarget. 7:62520–62532.
2016.PubMed/NCBI
|
|
109
|
Ma Y, Zhou G, Li M, Hu D, Zhang L, Liu P
and Lin K: Long noncoding RNA DANCR mediates cisplatin resistance
in glioma cells via activating AXL/PI3K/Akt/NF-κB signaling
pathway. Neurochem Int. 118:233–241. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Liang S, Zhang S, Wang P, Yang C, Shang C,
Yang J and Wang J: LncRNA, TUG1 regulates the oral squamous cell
carcinoma progression possibly via interacting with Wnt/β-catenin
signaling. Gene. 608:49–57. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Yang YT, Wang YF, Lai JY, Shen SY, Wang F,
Kong J, Zhang W and Yang HY: Long non-coding RNA UCA1 contributes
to the progression of oral squamous cell carcinoma by regulating
the WNT/β-catenin signaling pathway. Cancer Sci. 107:1581–1589.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Ge XS, Ma HJ, Zheng XH, Ruan HL, Liao XY,
Xue WQ, Chen YB, Zhang Y and Jia WH: HOTAIR, a prognostic factor in
esophageal squamous cell carcinoma, inhibits WIF-1 expression and
activates Wnt pathway. Cancer Sci. 104:1675–1682. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Wu KF, Liang WC, Feng L, Pang JX, Waye MM,
Zhang JF and Fu WM: H19 mediates methotrexate resistance in
colorectal cancer through activating Wnt/β-catenin pathway. Exp
Cell Res. 350:312–317. 2017. View Article : Google Scholar
|
|
114
|
Cao Y, Shi H, Ren F, Jia Y and Zhang R:
Long non-coding RNA CCAT1 promotes metastasis and poor prognosis in
epithelial ovarian cancer. Exp Cell Res. 359:185–194. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Fu Z, Chen C, Zhou Q, Wang Y, Zhao Y, Zhao
X, Li W, Zheng S, Ye H, Wang L, et al: LncRNA HOTTIP modulates
cancer stem cell properties in human pancreatic cancer by
regulating HOXA9. Cancer Lett. 410:68–81. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Yue B, Liu C, Sun H, Liu M, Song C, Cui R,
Qiu S and Zhong M: A Positive Feed-Forward Loop between
LncRNA-CYTOR and Wnt/β-Catenin Signaling Promotes Metastasis of
Colon Cancer. Mol Ther. 26:1287–1298. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Liu B, Pan CF, He ZC, Wang J, Wang PL, Ma
T, Xia Y and Chen YJ: Long noncoding RNA-LET suppresses tumor
growth and EMT in lung adenocarcinoma. Biomed Res Int.
2016:46934712016. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Wang X, Lu X, Geng Z, Yang G and Shi Y:
lncRNA PTCSC3/ miR-574-5p governs cell proliferation and migration
of papillary thyroid carcinoma via Wnt/β-catenin signaling. J Cell
Biochem. 118:4745–4752. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Zhang Z, Zhou C, Chang Y, Zhang Z, Hu Y,
Zhang F, Lu Y, Zheng L, Zhang W, Li X, et al: Long non-coding RNA
CASC11 interacts with hnRNP-K and activates the WNT/β-catenin
pathway to promote growth and metastasis in colorectal cancer.
Cancer Lett. 376:62–73. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Zhang F, Wan M, Xu Y, Li Z, Leng K, Kang
P, Cui Y and Jiang X: Long noncoding RNA PCAT1 regulates
extrahepatic cholangiocarcinoma progression via the
Wnt/β-catenin-signaling pathway. Biomed Pharmacother. 94:55–62.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Wang Y, Zhou J, Xu YJ and Hu HB: Long
non-coding RNA LINC00968 acts as oncogene in NSCLC by activating
the Wnt signaling pathway. J Cell Physiol. 233:3397–3406. 2018.
View Article : Google Scholar
|
|
122
|
Hang Q, Sun R, Jiang C and Li Y: Notch 1
promotes cisplatin-resistant gastric cancer formation by
upregulating lncRNA AK022798 expression. Anticancer Drugs.
26:632–640. 2015.PubMed/NCBI
|
|
123
|
Lu S, Dong W, Zhao P and Liu Z: lncRNA
FAM83H-AS1 is associated with the prognosis of colorectal carcinoma
and promotes cell proliferation by targeting the Notch signaling
pathway. Oncol Lett. 15:1861–1868. 2018.PubMed/NCBI
|
|
124
|
Chen H, Liu JZ, Hu GJ, Shi LL and Lan T:
Promotion of proliferation and metastasis of hepatocellular
carcinoma by LncRNA00673 based on the targeted-regulation of notch
signaling pathway. Eur Rev Med Pharmacol Sci. 21:3412–3420.
2017.PubMed/NCBI
|
|
125
|
Zhang L, Liang X and Li Y: Long non-coding
RNA MEG3 inhibits cell growth of gliomas by targeting miR-93 and
inactivating PI3K/AKT pathway. Oncol Rep. 38:2408–2416. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Li T, Xiao Y and Huang T: HIF-1α-induced
upregulation of lncRNA UCA1 promotes cell growth in osteosarcoma by
inactivating the PTEN/AKT signaling pathway. Oncol Rep.
39:1072–1080. 2018.PubMed/NCBI
|
|
127
|
Yun-Bo F, Xiao-Po L, Xiao-Li L, Guo-Long
C, Pei Z and Fa-Ming T: LncRNA TUG1 is upregulated and promotes
cell proliferation in osteosarcoma. Open Med (Wars). 11:163–167.
2016.
|
|
128
|
Yan J, Dang Y, Liu S, Zhang Y and Zhang G:
LncRNA HOTAIR promotes cisplatin resistance in gastric cancer by
targeting miR-126 to activate the PI3K/AKT/MRP1 genes. Tumour Biol.
37:16345–16355. 2016. View Article : Google Scholar
|
|
129
|
Wang H, Li Q, Tang S, Li M, Feng A, Qin L,
Liu Z and Wang X: The role of long noncoding RNA HOTAIR in the
acquired multidrug resistance to imatinib in chronic myeloid
leukemia cells. Hematology. 22:208–216. 2017. View Article : Google Scholar
|
|
130
|
Jiang N, Wang X, Xie X, Liao Y, Liu N, Liu
J, Miao N, Shen J and Peng T: lncRNA DANCR promotes tumor
progression and cancer stemness features in osteosarcoma by
upregulating AXL via miR-33a5p inhibition. Cancer Lett. 405:46–55.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Yuan Q, Liu Y, Fan Y, Liu Z, Wang X, Jia
M, Geng Z, Zhang J and Lu X: LncRNA HOTTIP promotes papillary
thyroid carcinoma cell proliferation, invasion and migration by
regulating miR-637. Int J Biochem Cell Biol. 98:1–9. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Xue D, Zhou C, Lu H, Xu R, Xu X and He X:
LncRNA GAS5 inhibits proliferation and progression of prostate
cancer by targeting miR-103 through AKT/mTOR signaling pathway.
Tumour Biol. 37:16187–16197. 2016. View Article : Google Scholar
|
|
133
|
Huang YS, Chang CC, Lee SS, Jou YS and
Shih HM: Xist reduction in breast cancer upregulates AKT
phosphorylation via HDAC3-mediated repression of PHLPP1 expression.
Oncotarget. 7:43256–43266. 2016.PubMed/NCBI
|
|
134
|
Lu Y, Li Y, Chai X, Kang Q, Zhao P, Xiong
J and Wang J: Long noncoding RNA HULC promotes cell proliferation
by regulating PI3K/AKT signaling pathway in chronic myeloid
leukemia. Gene. 607:41–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Wang J, Ma W and Liu Y: Long non-coding
RNA HULC promotes bladder cancer cells proliferation but inhibits
apoptosis via regulation of ZIC2 and PI3K/AKT signaling pathway.
Cancer Biomark. 20:425–434. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
He XS, Guo LC, Du MZ, Huang S, Huang RP,
Zhan SH, Gu DM, Liu WS, Wang XM, Wu H, et al: The long non-coding
RNA NONHSAT062994 inhibits colorectal cancer by inactivating Akt
signaling. Oncotarget. 8:68696–68706. 2017.PubMed/NCBI
|
|
137
|
Han W and Liu J: LncRNA-p21 inhibited the
proliferation of osteosarcoma cells via the miR-130b/PTEN/AKT
signaling pathway. Biomed Pharmacother. 97:911–918. 2018.
View Article : Google Scholar
|
|
138
|
Wang L, Wang F, Na L, Yu J, Huang L, Meng
ZQ, Chen Z, Chen H, Ming LL and Hua YQ: LncRNA AB209630 inhibits
gemcitabine resistance cell proliferation by regulating PI3K/ AKT
signaling in pancreatic ductal adenocarcinoma. Cancer Biomark.
22:169–174. 2018. View Article : Google Scholar
|
|
139
|
Wang B, Jiang H, Wang L, Chen X, Wu K,
Zhang S, Ma S and Xia B: Increased MIR31HG lncRNA expression
increases gefitinib resistance in non-small cell lung cancer cell
lines through the EGFR/PI3K/AKT signaling pathway. Oncol Lett.
13:3494–3500. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Chen S, Wang Y, Zhang JH, Xia QJ, Sun Q,
Li ZK, Zhang JG, Tang MS and Dong MS: Long non-coding RNA PTENP1
inhibits proliferation and migration of breast cancer cells via AKT
and MAPK signaling pathways. Oncol Lett. 14:4659–4662. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Li Q, Feng Y, Chao X, Shi S, Liang M, Qiao
Y, Wang B, Wang P and Zhu Z: HOTAIR contributes to cell
proliferation and metastasis of cervical cancer via targetting
miR-23b/MAPK1 axis. Biosci Rep. 38:BSR201715632018. View Article : Google Scholar :
|
|
142
|
Gao R, Zhang R, Zhang C, Zhao L and Zhang
Y: Long noncoding RNA CCAT1 promotes cell proliferation and
metastasis in human medulloblastoma via MAPK pathway. Tumori.
104:43–50. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Liu S, Yan G, Zhang J and Yu L: Knockdown
of long noncoding RNA (lncRNA) metastasis-associated lung
adenocarcinoma transcript 1 (MALAT1) inhibits proliferation,
migration, and invasion and promoted apoptosis by targeting miR-124
in retinoblastoma. Oncol Res. May 21–2017.Epub ahead of print.
View Article : Google Scholar
|
|
144
|
Huang JL, Ren TY, Cao SW, Zheng SH, Hu XM,
Hu YW, Lin L, Chen J, Zheng L and Wang Q: HBx-related long
non-coding RNA DBH-AS1 promotes cell proliferation and survival by
activating MAPK signaling in hepatocellular carcinoma. Oncotarget.
6:33791–33804. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Peng WX, Huang JG, Yang L, Gong AH and Mo
YY: Linc-RoR promotes MAPK/ERK signaling and confers
estrogen-independent growth of breast cancer. Mol Cancer.
16:1612017. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Wu F, Mo Q, Wan X, Dan J and Hu H:
NEAT1/has-mir-98 5p/MAPK6 axis is involved in non-small-cell lung
cancer (NSCLC) development. J Cell Biochem. Nov 2–2017.Epub ahead
of print. View Article : Google Scholar
|
|
147
|
Peng W and Fan H: Long noncoding RNA CCHE1
indicates a poor prognosis of hepatocellular carcinoma and promotes
carcinogenesis via activation of the ERK/MAPK pathway. Biomed
Pharmacother. 83:450–455. 2016. View Article : Google Scholar : PubMed/NCBI
|