Introduction
Breast cancer (BC) is the most frequently diagnosed
cancer and the second leading cause of cancer-associated mortality
among women worldwide (1). It was
estimated that there were ~2.1 million newly diagnosed BC cases
among women worldwide in 2018 (2).
Despite significant improvements in early diagnosis and therapeutic
strategies, patients with BC have a poor prognosis (3), which is predominantly due to tumor
metastasis (4). Therefore,
understanding the mechanisms underlying BC metastasis is crucial
for developing novel therapeutic strategies for BC.
Epithelial-to-mesenchymal transition (EMT), as a
vital process for tumor metastasis, is essential for the
transformation of early tumors into aggressive malignancies
(5). During EMT, the cells lose
their epithelial characteristics, including cell polarity and
specialized cell-cell adhesions, and exhibit migratory and invasive
behavior (6). During the process
of EMT, the expression levels of the epithelial marker E-cadherin
are significantly downregulated, while those of mesenchymal cell
markers, such as Snai1 and N-cadherin, are upregulated (7-9).
Accumulating evidence has demonstrated that the transforming growth
factor (TGF)-β/SMAD signaling pathway plays a key role in EMT
(10,11). TGF-β/SMAD signaling is initiated by
the binding of the TGF-β1/2 to the TGF-β receptor (TGFβR) II. The
ligand-receptor complex then recruits TGFβR1 to activate downstream
SMAD2/3. Phosphorylated (p-)SMAD2/3 further forms trimers with
SMAD4, which translocate to the nucleus, subsequently leading to
the activation of crucial transcription factors regulating EMT
(12). Of note, previous studies
have indicated that numerous microRNAs (miRNAs/miRs) play pivotal
roles in TGF-β/SMAD signaling-mediated EMT and tumor metastasis
(13-15).
miRs are a class of endogenous, highly conserved,
small non-coding RNAs that are ~22 nucleotides in length, and
negatively regulate gene expression by binding to the
3′-untranslated region (UTR) of target mRNAs, suppressing
translation and inducing mRNA degradation (16-18).
Recent studies demonstrated that miR-133b mainly acts as a tumor
suppressor in numerous types of human cancer (19-21).
In colorectal cancer, it has been reported that patients with low
levels of miR-133b expression have a poor prognosis in terms of
overall survival and positive lymph node metastasis (22). Overexpression of miR-133b inhibits
the clonogenicity and invasion of BC cells (23); however, whether miR-133b affects
TGF-β-induced EMT and metastasis of BC cells and the underlying
mechanism remain unknown.
The aim of the present study was to investigate the
association of miR-133b expression with BC metastasis and clinical
stage, as well as the correlation between the expression of
miR-133b and TGFβR1 in BC, hoping that the findings will provide
further insight into the mechanisms by which miRNA regulates
TGF-β-induced EMT and BC metastasis, and to determine whether
miR-133b may be considered as a potential prognostic predictor and
therapeutic target for the clinical diagnosis and treatment of
BC.
Materials and methods
Patients and tissue samples
A total of 78 paired BC and adjacent normal tissues
(≥2 cm away from the edge of cancer tissue, with no cancer cells
identified via microscopy) were obtained from The First Affiliated
Hospital of Nantong University between June 2013 and December 2018.
Among the 78 patients enrolled, 42 had BC (mean age 60.9 years;
range, 32-77 years), while 36 had metastatic BC (mean age, 66.3
years; range, 48-87 years). Pathological analysis of patients with
BC was performed according to the Revised International System for
Staging Breast Cancer (24). None
of the patients had been treated with chemotherapy, radiotherapy,
or any other therapy, prior to surgical intervention. All tissues
were immediately frozen in liquid nitrogen following surgery, and
were then stored at -80°C until use.
Cell lines and cell culture
The human BC cell line MDA-MB-468 and the
immortalized normal breast epithelial cell line MCF-10A were
purchased from the American Type Culture Collection. BC cells
MCF-7, MDA-MB-231 and MDA-MB-453 were obtained from the Cell Bank
of Chinese Academy of Sciences. MDA-MB-231, MDA-MB-453 and
MDA-MB-468 cells were cultured in RPMI-1640 medium (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(HyClone; GE Healthcare Life Sciences). MCF-7 cells were cultured
in Dulbecco's Modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc.) with 10 µg/ml human recombinant insulin
(Sigma-Aldrich; Merck KGaA) and 10% fetal bovine serum (HyClone; GE
Healthcare Life Sciences). MCF-10A cells were cultured in RPMI-1640
medium with 10% fetal bovine serum and 100 ng/ml cholera toxin
(Sigma-Aldrich; Merck KGaA). Cells were cultured at 37°C in a
humidified atmosphere containing 5% CO2.
RNA extraction, cDNA synthesis, and
reverse transcription- quantitative polymerase chain reaction
(RT-qPCR) analysis
Cells (2×106) and tissues (200 mg) were
employed for total RNA extraction using TRIzol® reagent
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocols. The concentration and purity of total RNA were detected
using a NanoDrop2000 spectrophotometer (NanoDrop Technologies;
Thermo Fisher Scientific, Inc.). Pre-designed stem-loop RT primers
were employed to synthesize the cDNA of miR-133b and U6 (25), and random primers were used for the
cDNA synthesis of TGFβR1. The synthesis of cDNA using reverse
transcriptase was performed using the M-MLV First Strand kit
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. Briefly, the reaction conditions were as follows: 25°C
for 10 min, 42°C for 45 min, and 85°C for 5 min. qPCR was performed
using SYBR Green qPCR SuperMix-UDG kits (Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. The
thermocy-cling conditions were as follows: Initial preincubation at
95°C for 10 min, followed by 40 cycles of amplification at 95°C for
5 sec and at 60°C for 30 sec. qPCR was performed on an ABI Prism
7500 Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The primer sequences used for RT and qPCR were
pre-synthesized (Genewiz Inc.) and are summarized in Table I. U6 was used for the normalization
of miR-133b expression, whereas β-actin served as an endogenous
control for TGFβR1 mRNA expression. RT-qPCR analysis was conducted
in triplicate; relative expression levels were evaluated using the
2-ΔΔCq method (26).
 | Table IPrimer sequences for RT and
RT-qPCR. |
Table I
Primer sequences for RT and
RT-qPCR.
| Name | Sequence
(5′–3′) |
|---|
| RT primers | |
| U6 |
CGAGCACAGAATCGCTTCACGAATTTGCGTGTCAT |
| miR-940 |
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGATAGCTGGTCGTA |
| RT-qPCR
primers | |
| U6 | F:
CGAGCACAGAATCGCTTCA; R: CTCGCTTCGGCAGCACATAT |
| miR-940 | F:
CAGTGCAGGGTCCGAGGTAT; R: CGTCTTTGGTCCCCTTCAAC |
| TGFβR1 | F:
GAGGAAAGTGGCGGGGAG; R: CCAACCAGAGCTGAGTCCAAGTA |
| β-actin | F:
CACAGAGCCTCGCCTTTGCC; R: CATGCCGGAGCCGTTGTCG |
Establishment of stable cell lines
overexpressing miR-133b
To stably overexpress miR-133b (LV-miR-133b) in
MDA-MB-231 cells, the miR-133b sequence was cloned into the
pLKO.1-puro-vector (InovoGen Tech. Co.) via the AgeI and
EcoRI (Fermentas; Thermo Fisher Scientific, Inc.) sites to
generate the pLKO.1-puro miR-133b plasmid. Then, the aforementioned
construct or control vector with the packaging plasmids psPAX2 and
pMD2.G (Geneseed Biotech) were co-transfected into 293T cells to
produce the lentivirus. MDA-MB-231 cells were infected with the
lentivirus and selected with 1 µg/ml puromycin
(Sigma-Aldrich; Merck KGaA) to obtain stable cell lines
(LV-miR-133b). Finally, the stable cell lines were used for in
vivo experiments of metastasis. The sequence of the miR-133b
segment was 5′-CCG GUC AGA AGA AAG AUG CCC CCU GCU CUG GCU GGU CAA
ACG GAA CCA AGU CCG UCU UCC UGA GAG GUU UGG UCC CCU UCA ACC AGC UAC
AGC AGG GCU GGC AAU GCC CAG UCC UUG GAG A-3′.
RNA oligonucleotides and cell
transfection
In the present study, two small interfering
(si)RNAs) were employed, which were designed using BLOCK-iT™ RNAi
Designer (https://rnaid-esigner.thermofisher.com/rnaiexpress/design.do)
and synthesized by Shanghai GenePharma Co., Ltd., to specifically
target TGFβR1 (si-TGFβR1-1 and si-TGFβR1-2). Scramble siRNA (si-NC)
was used as a negative control. miR-133b mimic, inhibitor
(anti-miR-133b) and negative controls (NCs) were obtained from
Shanghai GenePharma Co., Ltd. MCF-7 and MDA-MB-231 cells were
cultured on 6-well plates and transiently transfected using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) when 70-80% confluence was attained, according to
the manufacturer's protocols. At 48 h post-transfection, the cells
were harvested for further analysis. The sequences were as follows:
si-TGFβR1-1, 5′-CCA UUG AUA UUG CUC CAA A-3′ (sense), si-TGFβR1-2,
5′-GCA GCU AGG CUU ACA GCA U-3′ (sense), si-NC, 5′-UUC UCC GAA CGU
GUC ACG U-3′ (sense); miR-133b mimics, 5′-UUU GGU CCC CUU CAA CCA
GCU A-3′ (sense), and 5′-GCU GGU UGA AGG GGA CCA AAU U-3′
(antisense); miR-133b NC (miR-NC), 5′-UUC UCC GAA CGU GUC ACG U-3′
(sense), and 5′-ACG UGA CAC GUU CGG AGA A-3′ (antisense);
anti-miR-133b, 5′-GCU GGU UGA AGG GGA CCA AAU U-3′; inhibitor NC
(anti-miR-NC), 5′-CAG UAC UUU UGU GUA GUA CAA-3′.
Dual-luciferase reporter assay
The bioinformatic tools TargetScan (http://www.targetscan.org) and miRPathDB (https://mpd.bioinf.uni-sb.de/mirnas.html) were used to
predict whether miR-133b targets TGFβR1. The psiCHECK-2
dual-luciferase vector (Promega Corporation) was used to generate a
construct containing the TGFβR1 3′-UTR fused to the 3′-end of the
luciferase reporter. The wild-type (WT) fragment containing
predicted miR-133b target sites (positions 2,161-2,167) and mutant
(MUT) fragment were directly synthesized (Genewiz, Inc.) and
respectively subcloned into the psiCHECK-2 vector to create
corresponding constructs. To investigate the effects of miR-133b on
luciferase activity, MCF-7 and MDA-MB-231 cells were inoculated in
24-well plates, and were then co-transfected with WT or MUT 3′-UTR
luciferase reporter plasmids, and miR-133b mimics or miR-NC using
Lipofectamine 3000. Following trans-fection for 48 h, the cells
were harvested, and the relative luciferase activity was determined
using a Dual-Luciferase Reporter Assay System (Promega Corporation)
on a TD20/20 Luminometer (Turner Designs) according to the
manufacturer's protocols. The relative luciferase activity was
presented as the ratio of Renilla/firefly in accordance with
the Dual-Luciferase Assay Manual (27). Each experiment was repeated in
triplicate.
Western blot assay
Cell protein lysates were prepared using
radioimmunoprecipitation assay buffer (Cell Signaling Technology,
Inc.) with protease and phosphatase inhibitor cocktail (Beyotime
Institute of Biotechnology), according to the manufacturers'
instructions. The protein concentration was determined using a
Bicinchoninic Acid Protein Assay kit (Beyotime Institute of
Biotechnology) and separated by 10% SDS-PAGE. Proteins were
subsequently transferred to polyvinylidene difluoride membranes
(EMD Millipore). The membranes were blocked with 3% bovine serum
albumin (Beyotime Institute of Biotechnology) for 2 h at room
temperature, and then incubated overnight at 4°C with primary
antibodies against TGFβR1 (1:3,000; cat. no. AF3025; R&D
Systems, Inc.), N-cadherin (1:3,000; cat. no. 610920; BD
Biosciences), Snail (1:1,000; cat. no. 3895s; Cell Signaling
Technology, Inc.), p-SMAD3 (1:3,000; cat. no. Ab52903; Abcam),
SMAD3 (1:3,000; cat. no. 9523S; Cell Signaling Technology, Inc.)
and β-actin (1:5,000; cat. no. af5172; Affinity) according to the
manufacturer's protocols. Then, the membranes were incubated at
room temperature for 1.5 h with goat anti-rabbit IgG (1:4,000; cat.
no. BA1054; Beyotime Institute of Biotechnology) or goat anti-mouse
IgG (1:4,000; cat. no. BA1050; Beyotime Institute of Biotechnology)
secondary antibodies. The protein bands were visualized with an
electrochemiluminescence kit (Vazyme) and analyzed with a ChemiDoc™
XRS system (Bio-Rad Laboratories, Inc.). Western blotting was
performed in triplicate and representative images are
presented.
Transwell migration and invasion
assays
For the migration assay, 8-µm Transwell
chambers (Corning Incorporated) were prepared, whereas the
membranes of chambers coated with Matrigel (200 ng/ml; BD
Biosciences) were used for the invasion experiments. According to
the manufacturer's protocols, at 48 h following transfection, MCF-7
and MDA-MB-231 cells were trypsinized and counted. A total of
5×104 cells in 100 µl serum-free RPMI-1640 medium
were added to the upper chamber, and 600 µl RPMI-1640 medium
supplemented with 20% fetal bovine serum was added to the lower
chamber. After 6 h, TGF-β1 (5 ng/ml) was added to the lower
chambers. After culturing for 24 h (migration) or 36 h (invasion)
in a 5% CO2 incubator at 37°C, the upper chamber was
removed, and the non-migrating or non-invading cells were wiped
away using a cotton swab. The cells in the lower chamber were fixed
with 100% methanol for 20 min and stained with 0.1% crystal violet
solution for 1 h at room temperature. Cells in five random fields
(magnification, ×100) were imaged and counted under a light
microscope. Experiments were performed in triplicate.
In vivo metastasis assays
To assess lung metastasis, 5-week-old female BALB/c
nude mice were purchased from the Laboratory Animal Center of
Nantong University. The mice were divided into two groups (5 mice
per group), including the LV-miR-133b and control (LV-miR-NC)
groups. The mice were bred and housed under specific pathogen-free
conditions. LV-miR-133b and LV-miR-NC MDA-MB-231 cells
(3×105 cells/mouse) in PBS were injected into the tail
vein of mice. At 8 weeks following the injection of MDA-MB-231
cells, the mice were euthanized; the lungs were collected and fixed
in Bouin's solution for metastatic nodule analysis, or fixed with
4% polyoxymethylene for H&E staining.
H&E staining
According to the H&E staining protocols
(15), the lung tissues were fixed
in 4% paraformaldehyde overnight at room temperature, and
dehydrated through an ethanol gradient [60, 70, 80, 95, 100 (I) and
100% (II)] for 20 min per concentration. Then, the samples were
embedded in paraffin for 40 min at 60°C and cut into 5-µm
sections. The lung tissue sections were dewaxed with xylene (I) and
xylene (II) for 10 min, followed by rehydration via an ethanol
gradient (100, 95, 80, 70 and 60%) for 5 min per concentration.
Subsequently, the tissue sections were washed with distilled water
for 2 min, followed by hematoxylin staining for 5 min, and washing
in distilled water for 30 sec. Subsequently, the samples were
treated with 1% hydrochloric acid ethanol for 10 sec and washed for
15 min. Next, the lung tissue sections were stained with 1% eosin
for 2 min and washed for 2 min. The tissue sections were dehydrated
via an ethanol gradient (95, 95, 100 and 100%), and then cleared
with xylene (I) and xylene (II) for 10 min. The sections were
sealed with neutral gum and viewed under a light microscope
(Olympus Corporation).
Statistical analysis
SPSS 23.0 (IBM Corp.) and GraphPad Prism 7.01
(GraphPad Software, Inc.) were used for all statistical analyses.
In clinical samples of BC, differences between two groups were
assessed using paired or unpaired Student's t-tests (two-tailed).
The association between the expression levels of miR-133b or
TGFβR1, and the clinico-pathological parameters of patients with BC
were analyzed by non-parametric tests (Mann-Whitney U test for two
groups, and a Kruskal-Wallis test for multiple groups). The
correlation between the miR-133b and TGFβR1 mRNA expression level
ratios (T/N) in the BC tissues was analyzed by Spearman's rank
correlation analysis. Differences among multiple groups were
determined by one-way ANOVA followed by Tukey's post hoc test.
Prognostic analysis was performed using the Kaplan-Meier method
with log-rank test. The results from tissues are presented as box
and whiskers with minimum and maximal values, while other data are
expressed as mean ± standard deviation. All data were generated
from three independent experiments. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-133b expression is downregulated and
is inversely associ- ated with TGFβR1 expression in BC cells and
tissues
In order to investigate the association between
miR-133b and TGFβR1, RT-qPCR was performed using four human BC cell
lines. As shown in Fig. 1A,
miR-133b expression levels were notably decreased in the MCF-7,
MDA-MB-231, MDA-MB-453 and MDA-MB-468 cell lines compared with
those in the immortalized normal breast epithelial cell line
MCF-10A. Additionally, the four BC cell lines exhibited
significantly increased TGFβR1 mRNA expression levels (P<0.01;
Fig. 1B). To further confirm
whether there is an inverse association between miR-133b and TGFβR1
mRNA expression in patients with BC, and whether it affects the
development of BC, RT-qPCR was performed. From the analysis of 78
BC tissues and paired non-cancerous tissues, a significant
reduction in miR-133b expression (P<0.001; 61/78, 78.2%) and a
significant increase in TGFβR1 mRNA expression (P<0.001; 59/78,
75.6%) were detected in BC tissues when compared with non-cancerous
breast tissues (Fig. 1C and D).
Furthermore, clinicopathological analysis revealed that miR-133b
expression (T/N) was markedly reduced in BC tissues from patients
with advanced tumor-node-metastasis (TNM) stage and lymph node
metastasis compared with those with earlier TNM stage and no lymph
node metastasis (P<0.05; Table
II). On the contrary, the mRNA expression levels of TGFβR1
(T/N) were higher in BC tissues from patients with advanced TNM
stage and lymph node metastasis compared with those with earlier
TNM stage and no lymph node metastasis (P<0.05; Table II). Of note, a significant inverse
correlation was observed between miR-133b expression (T/N) and
TGFβR1 mRNA expression (T/N) in 78 paired tissues (r=-0.495,
P<0.001; Fig. 1E).
Additionally, in order to verify these findings, datasets
containing the expression data of BC tissues of the Gene Expression
Omnibus and The Cancer Genome Atlas (TCGA) were analyzed. Data of
the GSE19536 dataset revealed that the expression of miR-133b is
downregulated in BC tissues (P<0.001; Fig. 1F); TCGA (NM_004612_1_2228)
indicated that TGFβR1 mRNA expression was upregulated in invasive
BC (P<0.001; Fig. 1G). These
results support the findings of the present study on expression,
and the association between miR-133b and TGFβR1 expression in BC
cells and tissues. Collectively, these results suggest an inverse
association between the expression of miR-133b and TGFβR1, which
indicates a potential role for miR-133b in regulating TGFβR1
expression in BC.
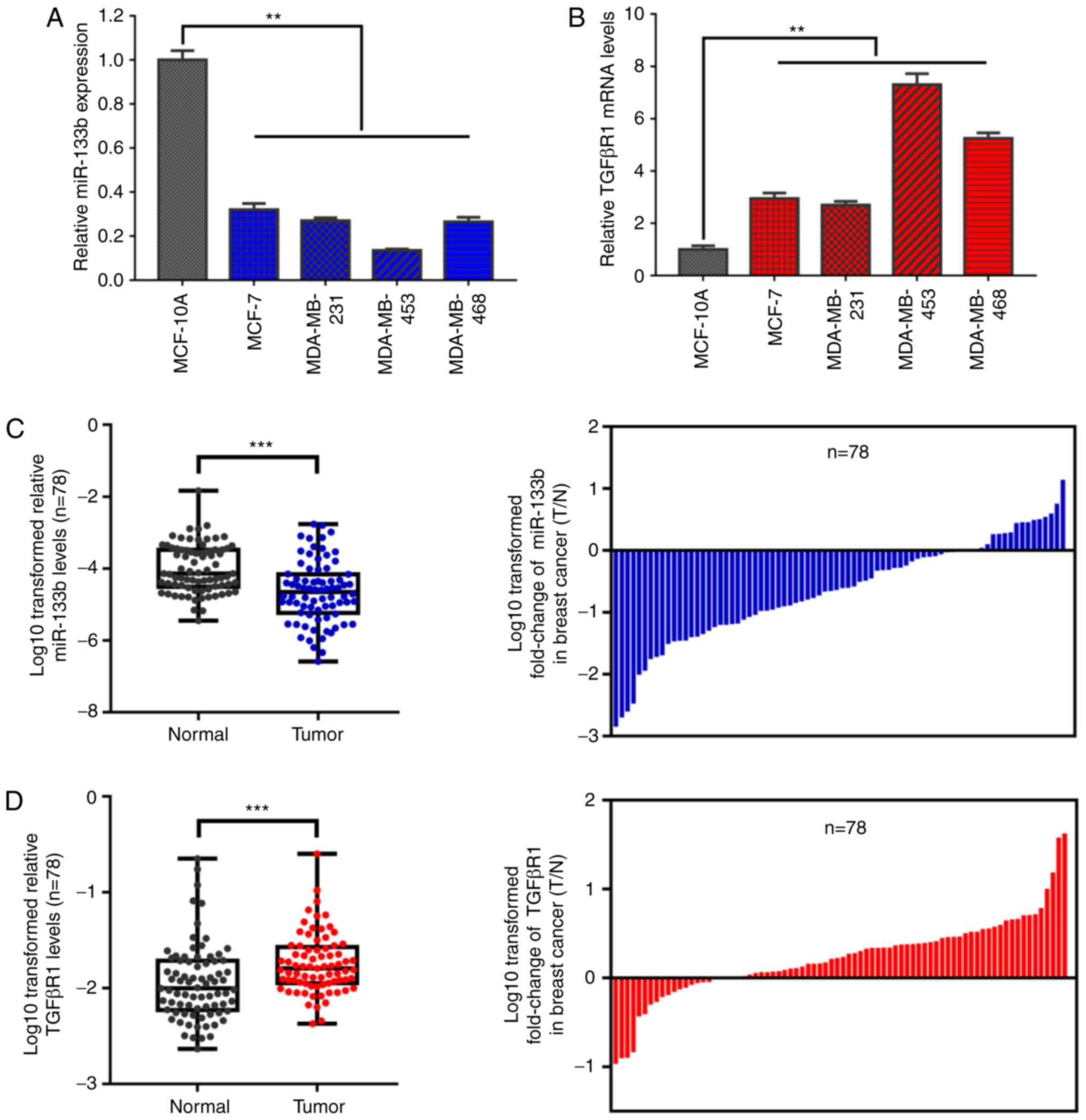 | Figure 1miR-133b expression is downregulated
and inversely correlated with TGFβR1 expression in BC cell lines
and tissues. RT-qPCR analysis of (A) miR-133b and (B) TGFβR1 mRNA
expression levels in human breast epithelial and BC cells. U6 and
β-actin were used as internal controls. Each RT-qPCR analysis was
performed in triplicate. RT-qPCR analysis of (C) miR-133b and (D)
TGFβR1 mRNA expression in 78 human T and N samples (left panel).
Mean values are indicated by solid bars, and values are expressed
as mean ± standard deviation. The log10 transformed fold change
(T/N) of miR-133b and TGFβR1 expression levels of each sample are
presented (right panel). U6 and β-actin were used as internal
controls. (E) Correlation between miR-133b and TGFβR1 mRNA
expression in 78 paired T samples. X and Y axes represent the log10
transformed fold change of T/N expression ratios of miR-133b and
TGFβR1 mRNA levels, respectively. The differential expression of
(F) miR-133b and (G) TGFβR1 were analyzed using the GSE19536
dataset of the Gene Expression Omnibus and The Cancer Genome Atlas
(NM_004612_1_2228), respectively. Data are presented as box and
whiskers with minimum and maximal values. **P<0.01,
***P<0.001. BC, breast cancer; miR, microRNA;
RT-qPCR, reverse transcription-quantitative polymerase chain
reaction; T, tumor tissues; N, paired non-cancerous breast tissues;
TGFβR1, transforming growth factor β receptor I. |
| Table IIDifferential expression ratios (T/N)
of miR-133b and TGFβR1 mRNA between various clinicopathological
parameters in breast cancer tissues. |
Table II
Differential expression ratios (T/N)
of miR-133b and TGFβR1 mRNA between various clinicopathological
parameters in breast cancer tissues.
| Parameters | n | miR-133b | TGFβR1 mRNA |
|---|
| Age, years | | | |
| <65 | 38 | 1.044±0.383 | 2.220±0.403 |
| ≥65 | 40 | 0.766±0.191 | 2.376±0.316 |
| aP-value | | 0.317 | 0.541 |
| Pathological
type | | | |
| Non-invasive
carcinoma | 5 | 1.668±0.632 | 2.312±0.853 |
| Early invasive
carcinoma | 12 | 1.561±0.531 | 1.756±0.377 |
| Invasive
carcinoma | 61 | 0.673±0.110 | 2.406±0.912 |
| bP-value | | 0.034 | 0.683 |
| TNM stage | | | |
| I | 28 | 1.227±0.479 | 1.964±0.260 |
| II | 19 | 0.841±0.225 | 1.859±0.351 |
| III and IV | 31 | 0.460±0.141 | 2.874±0.543 |
| bP-value | | 0.041 | 0.024 |
| Lymph node
metastasis | | | |
| No | 42 | 1.102±0.415 | 1.966±0.210 |
| Yes | 36 | 0.729±0.162 | 2.689±0.486 |
| aP-value | | 0.013 | 0.039 |
miR-133b suppresses TGFβR1 expression by
directly targeting TGFβR1 3′-UTR in BC cells
miRs play an important role in negatively regulating
gene expression by binding to the 3′-UTR of target mRNAs (16). Therefore, it was hypothesized that
miR-133b suppresses TGFβR1 expression by directly binding to the
3′-UTR of TGFβR1. To verify this hypothesis, two mRNA
target-predicting tools (TargetScan 7.1 and miRPathDB) were used to
identify the candidate target genes of miR-133b. As expected,
TGFβR1 was validated as a putative target gene of miR-133b, as the
3′-UTR of TGFβR1 was observed to contain one conserved binding site
for miR-133b (Fig. 2A). To further
confirm the prediction, a dual-luciferase reporter assay was
conducted (Fig. 2A) to determine
whether miR-133b can recognize and target the seed region in the
3′-UTR of TGFβR1. The results revealed that miR-133b significantly
inhibited the luciferase activity in MCF-7 (P<0.01; Fig. 2B) and MDA-MD-231 cells (P<0.01;
Fig. 2C) transfected with the
TGFβR1 3′-UTR-WT reporter; however, this suppression was not
observed in cells transfected with the MUT reporter (Fig. 2B and C). Additionally,
overexpression of miR-133b (Fig. 2D
and G) markedly decreased the expression levels of TGFβR1 mRNA
(Fig. 2E and H) and protein
(Fig. 2F and I) in MCF-7 and
MDA-MB-231 cells, whereas cells transfected with miR-133b inhibitor
(Fig. 2D and G) exhibited
increased TGFβR1 expression at the mRNA (Fig. 2E and H) and protein (Fig. 2F and I) levels. In conclusion,
these findings indicated that TGFβR1 is a direct target gene of
miR-133b in BC cells.
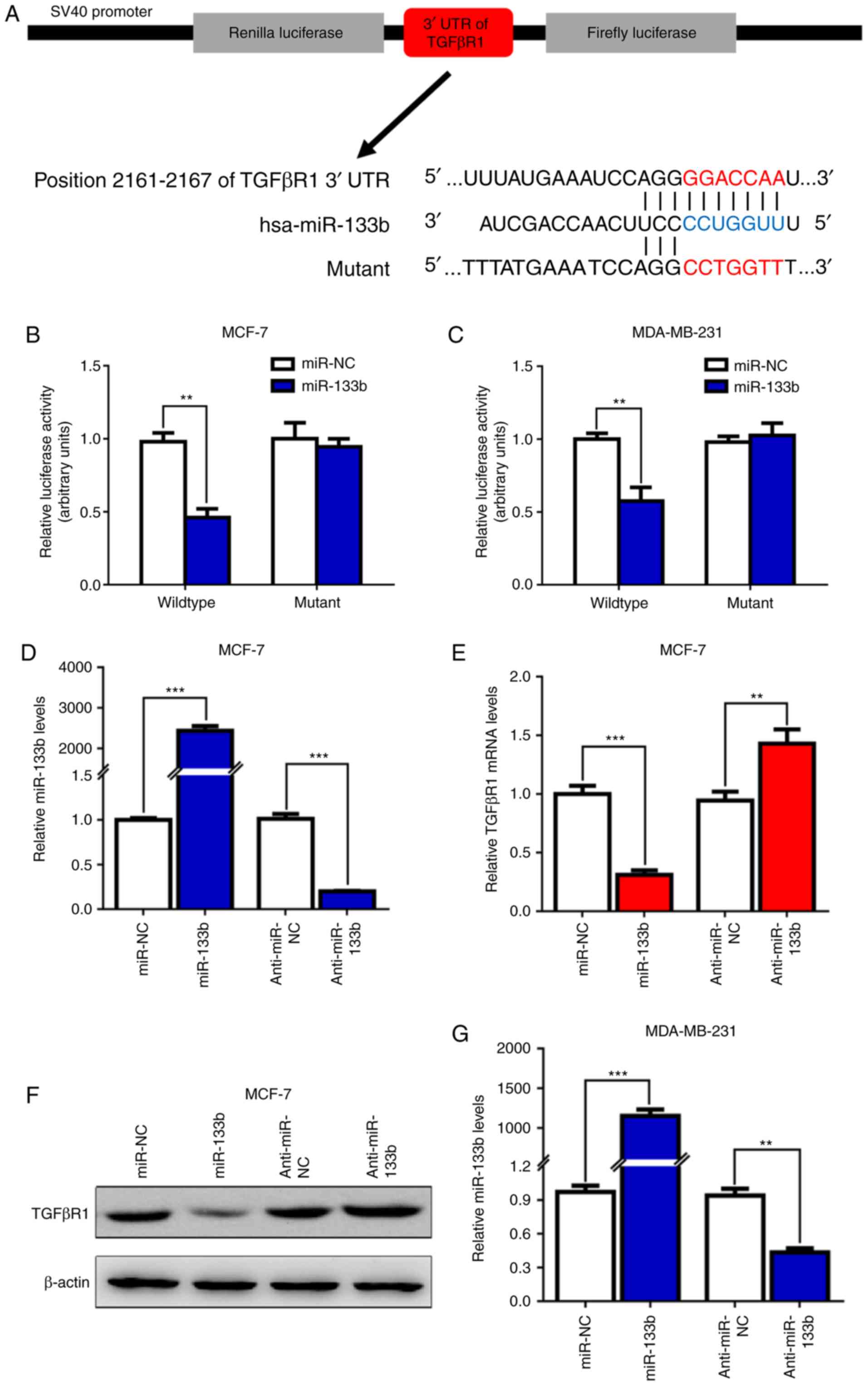 | Figure 2miR-133b inhibits TGFβR1 expression
by targeting the 3′-UTR of TGFβR1 in breast cancer cells. (A)
Schematic of the subcloning performed of the predicted miR-133b
binding sites of TGFβR1 3′-UTR into the psiCHECK-2 luciferase
vector. Predicted duplex formation between miR-133b and the
wild-type/mutant of miR-133b binding sites were presented. (B and
C) Relative luciferase activities of the wild-type or mutant TGFβR1
3′-UTR reporter gene in MCF-7 and MDA-MB-231 cells transfected with
miR-NC or miR-133b. Relative Renilla luciferase activity was
calculated after normalizing to firefly luciferase activity. (D)
The expression levels of miR-133b were detected by RT-qPCR in MCF-7
cells after transfection with miR-133b mimics or inhibitor for 48
h. Then, the expression levels of TGFβR1 (E) mRNA and (F) protein
were detected by RT-qPCR and western blotting, respectively. (G)
The expression levels of miR-133b were detected by RT-qPCR in
MDA-MB-231 cells after transfection with miR-133b mimics or
inhibitor for 48 h. **P<0.01,
***P<0.001. miR-133b inhibits TGFβR1 expression by
targeting the 3′-UTR of TGFβR1 in breast cancer cells. Then, the
expression levels of TGFβR1 (H) mRNA and (I) protein were detected
by RT-qPCR and western blotting, respectively.
**P<0.01, ***P<0.001. miR, microRNA;
NC, negative control; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; TGFβR1, transforming growth factor β
receptor I; 3′-UTR, 3′-untranslated region. |
miR-133b inactivates the TGF-β/SMAD
pathway, and suppresses TGF-β-induced EMT and cell invasion
TGFβR1 is an important receptor of the TGF-β/SMAD
signaling axis, and plays a key role in TGF-β-induced EMT and
cancer metastasis (28-30). It was observed that miR-133b
negatively regulates the expression of TGFβR1 (Fig. 2F and I) in BC cells. In order to
determine the biological effects of decreased miR-133b expression
on the progression and metastasis of BC, western blot and Transwell
assays were performed to analyze the effects of miR-133b on
TGF-β-induced EMT and BC cell migration and invasion. As shown in
Fig. 3A and B, overexpres-sion of
miR-133b significantly inhibited the protein expression of TGFβR1
in MCF-7 and MDA-MB-231 cells. Furthermore, upon treatment with
TGF-β1, BC cells transfected with miR-133b exhibited reduced
expression levels of p-SMAD3, which is an indispensable downstream
effector in canonical TGF-β/SMAD signaling; Snail and N-cadherin
expression was reduced compared with that in miR-NC-transfected
cells. Of note, the protein expression levels of SMAD3 were
markedly unchanged. Conversely, the ectopic expression of miR-133b
significantly suppressed the migration and invasion abilities of
MCF-7 (P<0.01; Fig. 3C and D)
and MDA-MB-231 (P<0.01; Fig. 3E and
F) cells induced by TGF-β in vitro. Collectively, these
results indicate that miR-133b overexpression may inhibit the
expression of TGFβR1, and inhibit TGF-β1-induced EMT and cell
migration and invasion via suppression of the TGF-β/SMAD signaling
pathway in BC cells.
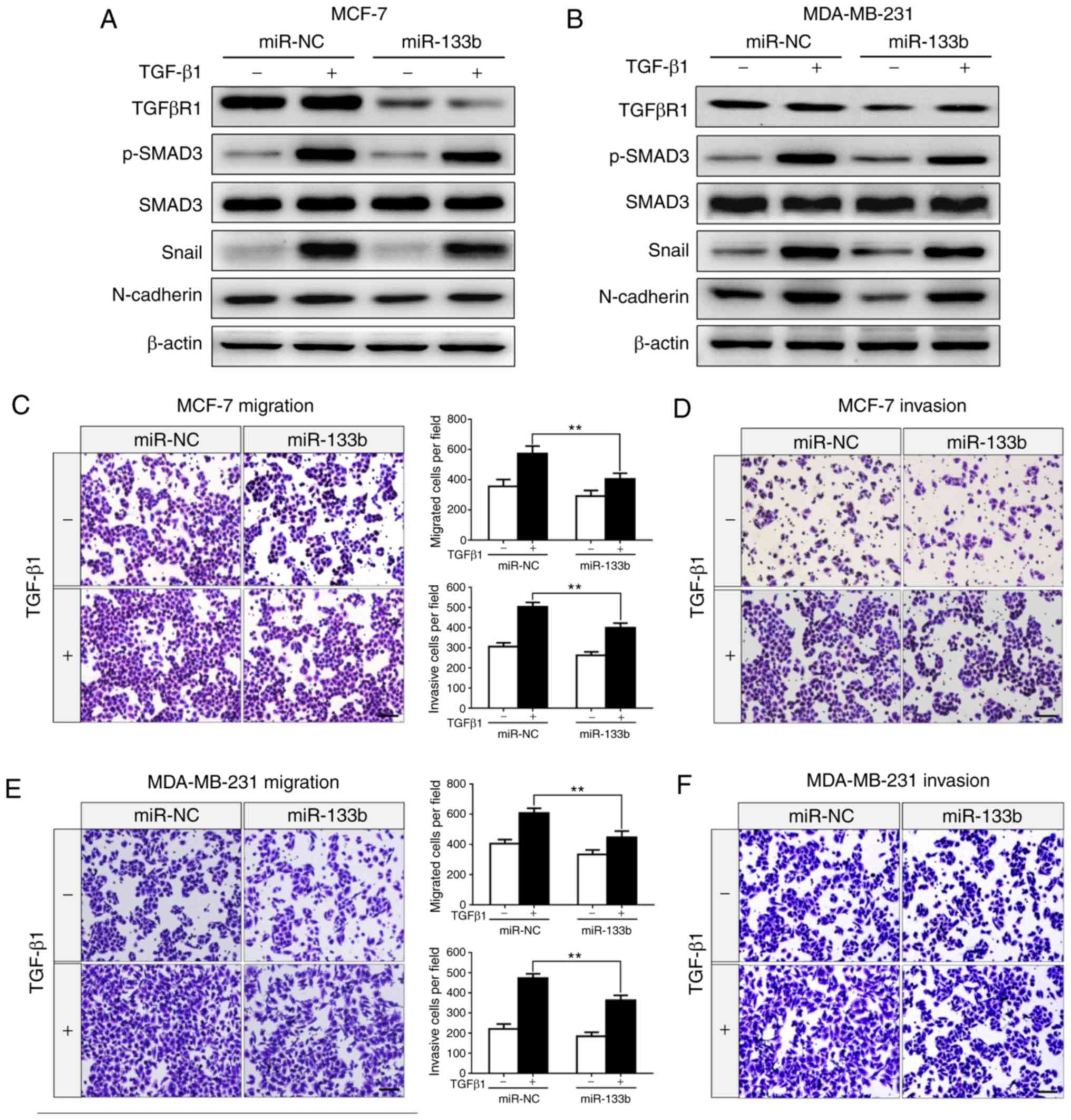 | Figure 3miR-133b suppresses TGF-β-induced
epithelial-to-mesenchymal transition and invasion of breast cancer
cells. (A) Western blot analysis for the expression of TGFβR1,
p-SMAD3, SMAD3, Snail and N-cadherin in MCF-7 cells transfected
with miR-NC or miR-133b mimics in the absence or presence of TGF-β1
(5 ng/ml) for 24 h. (B) Expression of TGFβR1, p-SMAD3, SMAD3, Snail
and N-cadherin in MDA-MB-231 cells transfected with miR-NC or
miR-133b mimics in the absence or presence of TGF-β1 (5 ng/ml) for
24 h. β-actin was used as an internal control. Transwell assays for
MCF-7 cells transfected with miR-NC or miR-133b mimics in the
absence or presence of TGF-β1 (5 ng/ml) for 24 h (migration) or 36
h (invasion). (C) Migrating and (D) invading cells were stained and
counted in at least three light microscopic fields. Scale bar, 100
µm. Transwell assays for MDA-MB-231 cells transfected with
miR-NC or miR-133b mimics in the absence or presence of TGF-β1 (5
ng/ml) for 24 h (migration) or 36 h (invasion). (E) Migrating and
(F) invading cells were stained and counted in at least three light
microscopic fields. Scale bar, 100 µm.
**P<0.01. miR, microRNA; NC, negative control;
TGFβR1, transforming growth factor β receptor I. |
Knockdown of TGFβR1 suppresses
TGF-β-induced EMT and BC cell invasion
To determine the functional role of TGFβR1, and
examine whether TGFβR1 knockdown can mimic the phenotype generated
by miR-133b overexpres-sion, two siRNAs (si-TGFβR1-1 and
si-TGFβR1-2) were used to specifically silence TGFβR1 expression.
The results demonstrated that TGFβR1 expression was markedly
down-regulated in the MCF-7 and MDA-MB-231 cells transfected with
TGFβR1 siRNA (Fig. 4A-C).
Subsequently, the protein expression levels of EMT markers were
detected in the presence or absence of TGF-β1. As shown in Fig. 4B and C, knockdown of TGFβR1
significantly inhibited TGF-β-induced upregulation of p-SMAD3,
Snail and N-cadherin in MCF-7 (Fig.
4B) and MDA-MB-231 (Fig. 4C)
cells. On the contrary, the protein expression levels of SMAD3 were
markedly unaltered. Additionally, the results of the Transwell
assays revealed that knockdown of TGFβR1 suppressed the migration
(P<0.01, Fig. 4D and F) and
invasion (P<0.01, Fig. 4E and
G) abilities of the cells. Our results indicated that TGFβR1
knockdown mimicked the phenotype induced by miR-133b
overexpression, which further suggested that miR-133b inhibits
TGF-β-induced EMT, cell migration and invasion by directly
targeting TGFβR1 in BC cells.
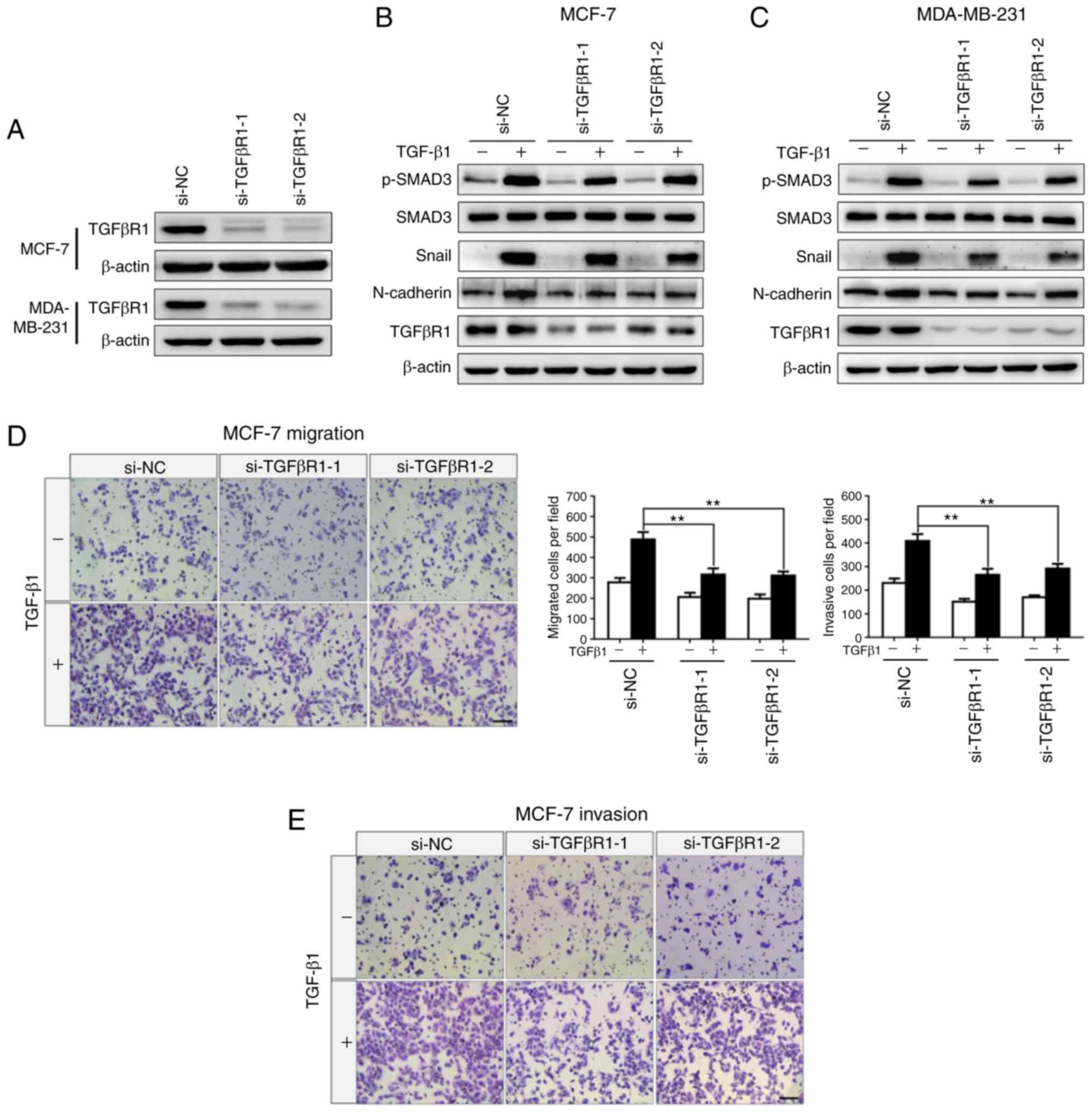 | Figure 4Knockdown of TGFβR1 suppresses
TGF-β-induced epithelial-to-mesenchymal transition and invasion of
breast cancer cells. (A) Western blot analysis for the expression
of TGFβR1 in MCF-7 and MDA-MB-231 cells transfected with si-NC,
si-TGFβR1-1 or si-TGFβR1-2. β-actin was used as an internal
control. Expression of TGFβR1, p-SAMD3, SMAD3, Snail, and
N-cadherin in (B) MCF-7 and (C) MDA-MB-231 cells transfected with
si-NC, si-TGFβR1-1 or si-TGFβR1-2 in the absence or presence of
TGF-β1 (5 ng/ml) for 24 h. Transwell assays for MCF-7 cells
transfected with si-NC, si-TGFβR1-1 or si-TGFβR1-2 in the absence
or presence of TGF-β1 (5 ng/ml) for 24 h (migration) or 36 h
(invasion). (D) Migrating and (E) invading cells were stained and
counted in at least three light microscopic fields. Scale bar, 100
µm. Transwell assays for MDA-MB-231 cells transfected with
si-NC, si-TGFβR1-1 or si-TGFβR1-2 in the absence or presence of
TGF-β1 (5 ng/ml) for 24 h (migration) or 36 h (invasion). (F)
Migrating and (G) invading cells were stained and counted in at
least three light microscopic fields. Scale bar, 100 µm.
**P<0.01. siRNA, small interfering RNA; TGFβR1,
transforming growth factor β receptor I. |
miR-133b attenuates BC cell metastasis in
vivo
To further investigate the role of miR-133b
overexpression in BC cell metastasis in vivo, LV-miR-133b
MDA-MB-231 cells were established. A ~50-fold increased expression
of miR-133b in LV-miR-133b MDA-MB-231 cells was observed
(P<0.001; Fig. 5A). Western
blot analysis confirmed the protein expression levels of TGFβR1 to
be significantly downregulated in MDA-MB-231 cells stably
overexpressing miR-133b compared with control cells (Fig. 5A). Then, LV-miR-133b and LV-miR-NC
MDA-MB-231 cells were intravenously injected into BALB/c nude mice
via the tail vein (Fig. 5B). At 8
weeks post-inoculation, the mice were euthanized and the lungs were
collected for the analysis of metastases and histological
examination. As expected, mice injected with LV-miR-133b MDA-MB-231
cells developed fewer metastatic nodules in the lungs (Fig. 5C and D). Consistently, H&E
staining of the lungs demonstrated that the number and size of
micrometastases were markedly deceased in LV-miR-133b mice compared
with control mice (Fig. 5C and E).
In addition, Kaplan-Meier survival analysis (31) revealed that patients with BC and
low levels of miR-133b expression had significantly shorter overall
survival compared with those with high miR-133b expression levels
(Fig. 5F). Conversely, the
expression levels of TGFβR1 were negatively associated with overall
survival in patients with BC (Fig.
5G). Based on these findings, it may be concluded that low
expression levels of miR-133b and high expression levels of TGFβR1
may be associated with poor prognosis in patients with BC.
Collectively, these data indicate that the ectopic expression of
miR-133b is associated with downregulated TGFβR1 expression, which
may suppress the invasion and metastasis of BC cells in
vivo.
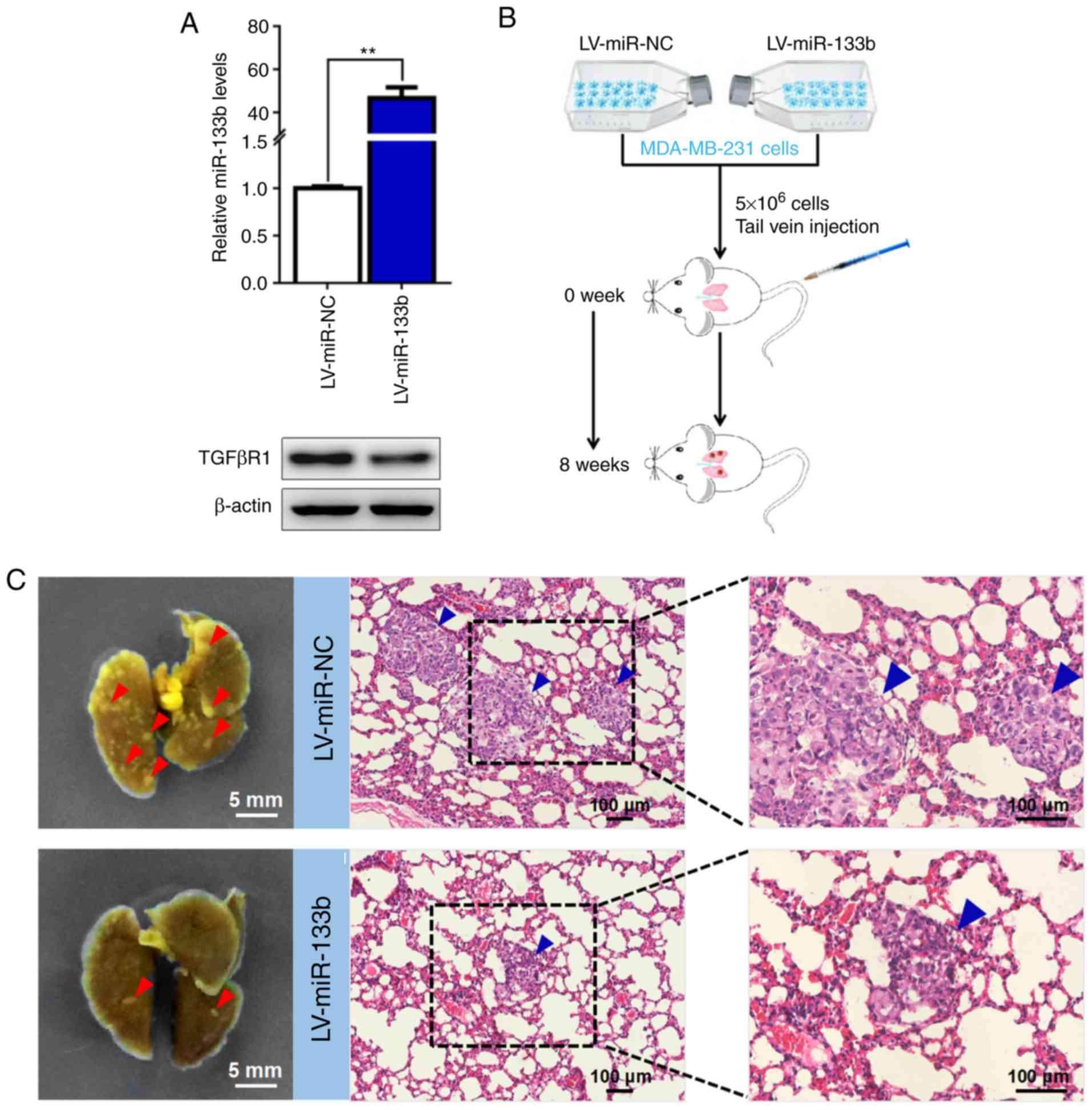 | Figure 5miR-133b attenuates BC cell
metastasis in vivo. (A) MDA-MB-231 cells stably
overexpressing miR-133b were generated. miR-133b expression was
determined by reverse transcription-quantitative polymerase chain
reaction analysis (upper panel). TGFβR1 expression was determined
by western blot analysis (lower panel). U6 and β-actin were used as
internal controls. (B) Schematic flowchart of the in vivo
metastasis experiments with MDA-MB-231 cells, which were
intravenously injected into BALB/c nude mice (n=5 mice per group).
(C) Representative images of metastatic nodules in the lungs
obtained from mice injected with miR-133b-overexpressing MDA-MB-231
cells or control cells (left panel). Scale bar, 5 mm. H&E
staining was performed for the histological analysis of metastatic
tumor cells in the lung. Scale bar, 100 µm. Red arrowheads,
metastatic nodules; blue arrowheads, micrometastases. (D) Visible
metastatic nodules and (E) micrometastases were counted and
analyzed. *P<0.05. Survival analysis was performed
using a Kaplan-Meier plotter to determine the association between
the expression of (F) miR-133b and (G) TGFβR1 and the overall
survival of patients with BC. **P<0.01. BC, breast
cancer; LV, lentivirus; miR, microRNA; NC, negative control;
TGFβR1, transforming growth factor β receptor I. |
Discussion
Despite advances in clinical therapy, metastasis
remains the leading cause of mortality among patients with BC
(32). It has been well documented
that the TGF-β/SMAD signaling pathway plays an important role in
EMT and tumor metastasis. Furthermore, the crucial upstream
receptor of the TGF-β/SMAD pathway, TGFβR1, plays a pivotal role in
TGF-β-induced EMT and tumor metastasis (28-30).
miR-133b was recently reported to be involved in the development
and progression of BC (23). The
effects of the crosstalk between miR-133b and the TGF-β/SMAD
pathway on tumor metastasis and the underlying mechanisms remain
unclear, particularly in BC. To the best of our knowledge, the
present study is the first to demonstrate that miR-133b directly
targets TGFβR1 to inhibit TGF-β-induced EMT and metastasis via
suppression of TGF-β/SMAD signaling.
Accumulating evidence has demonstrated that miRNAs
play a pivotal regulatory role in the initiation and development of
various cancers (33). Therefore,
an in-depth understanding of the biological function of specific
miRNAs in cancer may aid with the evaluation of potential
therapeutic targets. Recently, dysregulated miR-133b was reported
to be involved in the initiation and progression of BC (23,34);
however, the effects of miR-133b on TGF-β-induced EMT and tumor
metastasis and the underlying mechanisms require further
investigation, particularly in BC. The aim of the present study was
to investigate the biological behavior of miR-133b and the possible
mechanism of action in BC. As demonstrated by the findings,
miR-133b expression was found to be downregulated in BC cell lines
and tissues; this decrease was significantly associated with poor
prognosis of patients with BC. Furthermore, the expression of
miR-133b was inversely associated with lymph node metastasis and
TNM stage in patients with BC. Functional analysis indicated that
miR-133b significantly inhibited TGF-β-induced EMT and BC cell
invasion in vitro and in vivo. The findings of the
clinical analysis revealed that miR-133b may serve as a prognostic
marker in BC patients; however, whether miR-133b expression levels
are correlated with BC recurrence and whether they may serve as a
predictor of clinical outcome in BC is yet to be determined.
Consistent with our findings, recent studies have suggested that
miR-133b is also downregulated in gastric, lung and colorectal
cancer (35-37). On the contrary, Qin et al
(38) reported that miR-133b acted
as an oncogene, promoting the progression of cervical carcinoma.
This discrepancy may be attributed to differences in the tumor
microenvironment, tumor heterogeneity or target genes.
miRNAs may act as oncogenes or tumor-suppressor
genes, which is largely dependent on their targets in a variety of
human cancers (39). Bioinformatic
algorithms were utilized to predict potential targets, and
ultimately identified a novel miR-133b-targeted gene, TGFβR1, which
is a critical receptor in the TGFβ1/SMAD signaling pathway. TGFβR1
has been confirmed to play a pivotal role in TGF-β-induced EMT and
tumor metastasis (28-30). It has been well-documented that
TGFβ1/SMAD signaling plays a dual role in cancer development and
progression. Of note, activated TGF-β/SMAD signaling serves as a
tumor suppressor in early-stage tumors, but acts as a tumor
promoter in late-stage tumors (40,41).
Consistently, Fukai et al (42) demonstrated that the expression of
TGFRβ1 was downregulated in early-stage human esophageal squamous
cell carcinoma; however, our results were consistent with those of
a recent study by Li et al (43) reporting that TGFβR1 mRNA expression
was upregulated in hepatocellular carcinoma, with high mRNA levels
of TGFβR1 being associated with advanced TNM stage (43). Furthermore, in silico
analysis demonstrated that increased TGFβR1 expression was
associated with poor prognosis of BC patients. Most importantly, we
observed that siRNA-mediated knockdown of TGFβR1 suppressed
TGF-β-induced EMT and BC cell invasion, whereas TGFβR1 silencing
mimicked the effects and phenotype of miR-133b overexpression. The
findings of the present study may provide insight into the
association among miRNAs, the TGF-β/SMAD pathway and cancer
metastasis. However, there were certain limitations to the present
study, including insufficient materials, such as the small number
of BC cell lines and BALB/c nude mice used. Moreover, the present
study focused on the function of miR-133b in TGF-β-induced EMT,
which is an important event promoting metastasis in advanced-stage
cancers. Since TGF-β/SMAD signaling plays a key role in regulating
cell proliferation in terms of cell cycle progression during the
early stages of BC, it would be interesting to address the issue of
whether miR-133b regulates the cell cycle of BC cells in future
studies.
In summary, to the best of our knowledge, the
present study was the first to identify that miR-133b inhibits
TGF-β-induced EMT and BC metastasis via suppression of TGFβR1
expression in vitro and in vivo. These findings are
further supported by the inverse association observed between
miR-133b and TGFβR1 expression in BC cell lines and tissues. The
results of the present study suggest a novel mechanism by which
miRNA regulates TGF-β-mediated EMT and tumor metastasis, and
indicate that overexpression of miR-133b may be considered as a
promising strategy for treating advanced BC in the future.
Funding
The present study was supported by the grants from
Science and Technology Plan of Nantong (no. YYZ16044), the Students
Innovation and Entrepreneurship Training Program of Nantong
University (no. 2018138), the Research and Development program of
Kangda College of Nanjing Medical University (no. KD2018KYJJZD002)
and the ‘521 High-level Talent Cultivation Project' Research
program of Lianyungang (no. LYG521037).
Availability of data and materials
The datasets generated and analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
CL and SW designed the research and analyzed the
data. SW prepared the figures and drafted the manuscript. SW, MH,
ZW, WW and SQ performed the experiments. All authors have read and
approved the final version of this manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The First Hospital of Nantong University (Nantong,
China), and all enrolled patients provided written informed
consent. The animal experiments were conducted with the approval of
the Experimental Animal Ethical Committee of Nantong University,
and were conducted in accordance with the Guide for the Care and
Use of Laboratory Animals by US National Institutes of Health.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Ferlay J, Colombet M, Soerjomataram I,
Mathers C, Parkin DM, Piñeros M, Znaor A and Bray F: Estimating the
global cancer incidence and mortality in 2018: GLOBOCAN sources and
methods. Int J Cancer. 144:1941–1953. 2019.
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hartman EK and Eslick GD: The prognosis of
women diagnosed with breast cancer before, during and after
pregnancy: A meta-analysis. Breast Cancer Res Treat. 160:347–360.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kang Y and Massagué J:
Epithelial-mesenchymal transitions: Twist in development and
metastasis. Cell. 118:277–279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu J, Lamouille S and Derynck R:
TGF-beta-induced epithelial to mesenchymal transition. Cell Res.
19:156–172. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nieman MT, Prudoff RS, Johnson KR and
Wheelock MJ: N-cadherin promotes motility in human breast cancer
cells regardless of their E-cadherin expression. J Cell Biol.
147:631–644. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Martin TA, Goyal A, Watkins G and Jiang
WG: Expression of the transcription factors snail, slug, and twist
and their clinical significance in human breast cancer. Ann Surg
Oncol. 12:488–496. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Islam SS, Mokhtari RB, El Hout Y, Azadi
MA, Alauddin M, Yeger H and Farhat WA: TGF-β1 induces EMT
reprogramming of porcine bladder urothelial cells into collagen
producing fibroblasts-like cells in a Smad2/Smad3-dependent manner.
J Cell Commun Signal. 8:39–58. 2014. View Article : Google Scholar
|
|
11
|
Divya T, Velavan B and Sudhandiran G:
Regulation of Transforming Growth Foctor-β/Smad mediated
epithelial-mesenchymal transition by celastrol provides protection
against bleomycin-induced pulmonary fibrosis. Basic Clin Pharmacol
Toxicol. 123:122–129. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shi Y and Massagué J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang G, Du MY, Zhu H, Zhang N, Lu ZW,
Qian LX, Zhang W, Tian X, He X and Yin L: MiRNA-34a reversed
TGF-β-induced epithelial-mesenchymal transition via suppression of
SMAD4 in NPC cells. Biomed Pharmacother. 106:217–224. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu H, Xu Z, Li C, Xu C, Lei Z, Zhang HT
and Zhao J: MiR-145 and miR-203 represses TGF-β-induced
epithelial-mesenchymal transition and invasion by inhibiting SMAD3
in non-small cell lung cancer cells. Lung Cancer. 97:87–94. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang L, Tong X, Zhou Z, Wang S, Lei Z,
Zhang T, Liu Z, Zeng Y, Li C, Zhao J, et al: Circular RNA
hsa_circ_0008305 (circPTK2) inhibits TGF-β-induced
epithelial-mesenchymal transition and metastasis by controlling
TIF1γ in non-small cell lung cancer. Mol Cancer. 17:1402018.
View Article : Google Scholar
|
|
16
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zamore PD and Haley B: Ribo-gnome: The big
world of small RNAs. Science. 309:1519–1524. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nazarov PV, Reinsbach SE, Muller A, Nicot
N, Philippidou D, Vallar L and Kreis S: Interplay of microRNAs,
transcription factors and target genes: Linking dynamic expression
changes to function. Nucleic Acids Res. 41:2817–2831. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li X, Wan X, Chen H, Yang S, Liu Y, Mo W,
Meng D, Du W, Huang Y, Wu H, et al: Identification of miR-133b and
RB1CC1 as independent predictors for biochemical recurrence and
potential therapeutic targets for prostate cancer. Clin Cancer Res.
20:2312–2325. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kano M, Seki N, Kikkawa N, Fujimura L,
Hoshino I, Akutsu Y, Chiyomaru T, Enokida H, Nakagawa M and
Matsubara H: miR-145, miR-133a and miR-133b: Tumor-suppressive
miRNAs target FSCN1 in esophageal squamous cell carcinoma. Int J
Cancer. 127:2804–2814. 2010. View Article : Google Scholar
|
|
21
|
Guo L, Bai H, Zou D, Hong T, Liu J, Huang
J, He P, Zhou Q and He J: The role of microRNA-133b and its target
gene FSCN1 in gastric cancer. J Exp Clin Cancer Res. 33:992014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nohata N, Hanazawa T, Enokida H and Seki
N: microRNA-1/133a and microRNA-206/133b clusters: Dysregulation
and functional roles in human cancers. Oncotarget. 3:9–21. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang QY, Zhou CX, Zhan MN, Tang J, Wang
CL, Ma CN, He M, Chen GQ, He JR and Zhao Q: MiR-133b targets Sox9
to control pathogenesis and metastasis of breast cancer. Cell Death
Dis. 9:7522018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Thor A: A revised staging system for
breast cancer. Breast J. 10(Suppl 1): S15–S18. 2015. View Article : Google Scholar
|
|
25
|
Varkonyi-Gasic E, Wu R, Wood M, Walton EF
and Hellens RP: Protocol: A highly sensitive RT-PCR method for
detection and quantification of microRNAs. Plant Methods. 3:122007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
27
|
Wolter JM, Kotagama K, Pierre-Bez AC,
Firago M and Mangone M: 3′LIFE: A functional assay to detect miRNA
targets in high-throughput. Nucleic Acids Res. 42:e1322014.
View Article : Google Scholar
|
|
28
|
Liu RY, Zeng Y, Lei Z, Wang L, Yang H, Liu
Z, Zhao J and Zhang HT: JAK/STAT3 signaling is required for
TGF-β-induced epithelial-mesenchymal transition in lung cancer
cells. Int J Oncol. 44:1643–1651. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fang Y, Chen Y, Yu L, Zheng C, Qi Y, Li Z,
Yang Z, Zhang Y, Shi T, Luo J and Liu M: Inhibition of breast
cancer metastases by a novel inhibitor of TGFβ receptor 1. J Natl
Cancer Inst. 105:47–58. 2013. View Article : Google Scholar
|
|
30
|
Cortez VS, Ulland TK, Cervantes-Barragan
L, Bando JK, Robinette ML, Wang Q, White AJ, Gilfillan S, Cella M
and Colonna M: SMAD4 impedes the conversion of NK cells into
ILC1-like cells by curtailing non-canonical TGF-β signaling. Nat
Immunol. 18:995–1003. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nagy Á, Lánczky A, Menyhárt O and Győrffy
B: Validation of miRNA prognostic power in hepatocellular carcinoma
using expression data of independent datasets. Sci Rep. 8:92272018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dawood S, Broglio K, Buzdar AU, Hortobagyi
GN and Giordano SH: Prognosis of women with metastatic breast
cancer by HER2 status and trastuzumab treatment: An
institutional-based review. J Clin Oncol. 28:92–98. 2010.
View Article : Google Scholar :
|
|
33
|
Nicoloso MS, Spizzo R, Shimizu M, Rossi S
and Calin GA: MicroRNAs-the micro steering wheel of tumour
metastases. Nat Rev Cancer. 9:293–302. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chan M, Liaw CS, Ji SM, Tan HH, Wong CY,
Thike AA, Tan PH, Ho GH and Lee AS: Identification of circulating
microRNA signatures for breast cancer detection. Clin Cancer Res.
19:4477–4487. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wen D, Li S, Ji F, Cao H, Jiang W, Zhu J
and Fang X: miR-133b acts as a tumor suppressor and negatively
regulates FGFR1 in gastric cancer. Tumour Biol. 34:793–803. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Crawford M, Batte K, Yu L, Wu X, Nuovo GJ,
Marsh CB, Otterson GA and Nana-Sinkam SP: MicroRNA 133B targets
pro-survival molecules MCL-1 and BCL2L2 in lung cancer. Biochem
Biophys Res Commun. 388:483–489. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hu G, Chen D, Li X, Yang K, Wang H and Wu
W: miR-133b regulates the MET proto-oncogene and inhibits the
growth of colorectal cancer cells in vitro and in vivo. Cancer Biol
Ther. 10:190–197. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Qin W, Dong P, Ma C, Mitchelson K, Deng T,
Zhang L, Sun Y, Feng X, Ding Y, Lu X, et al: MicroRNA-133b is a key
promoter of cervical carcinoma development through the activation
of the ERK and AKT1 pathways. Oncogene. 31:4067–4075. 2012.
View Article : Google Scholar
|
|
39
|
Ventura A and Jacks T: MicroRNAs and
cancer: Short RNAs go a long way. Cell. 136:586–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lei Z, Xu G, Wang L, Yang H, Liu X, Zhao J
and Zhang HT: MiR-142-3p represses TGF-β-induced growth inhibition
through repression of TGFβR1 in non-small cell lung cancer. FASEB
J. 28:2696–2704. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Akhurst RJ and Derynck R: TGF-beta
signaling in cancer-a double-edged sword. Trends Cell Biol.
11:S44–S51. 2001.PubMed/NCBI
|
|
42
|
Fukai Y, Fukuchi M, Masuda N, Osawa H,
Kato H, Nakajima T and Kuwano H: Reduced expression of transforming
growth factor-beta receptors is an unfavorable prognostic factor in
human esophageal squamous cell carcinoma. Int J Cancer.
104:161–166. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li Y, Liu G, Li X, Dong H, Xiao W and Lu
S: Long non-coding RNA SBF2-AS1 promotes hepatocellular carcinoma
progression through regulation of miR-140-5p-TGFβR1 pathway.
Biochem Biophys Res Commun. 503:2826–2832. 2018. View Article : Google Scholar : PubMed/NCBI
|














