Introduction
Mucin-type O-glycosylation is a fundamental
post-translational modification in secreted or membrane-associated
proteins. The first step in O-glycan biosynthesis is
catalyzed by a large family of polypeptide
N-acetylgalactosaminyltransferases (GALNTs) that transfer
N-acetylgalactosamine (GalNAc) to serine or threonine residues on
the target proteins (1-4). Aberrant O-glycosylation of
cancer cells affects their growth and survival and their ability to
invade and metastasize (5).
Accumulating evidence has demonstrated that dysregulation of GALNT
expression results in aberrant O-glycosylation in breast
cancer cells (6). Among the
GALNTs, GALNT6 is upregulated in breast (7-11),
ovarian (12,13), renal cell (14), lung (15) and pancreatic (16) cancers. Notably, an increasing
number of studies have demonstrated that GALNT6 controls cancer
cell proliferation, migration and invasion abilities through the
regulation of mucin (MUC)1 (10),
fibronectin (17), MUC4 (18), glucose-regulated protein 78 (GRP78)
(19), estrogen receptor (ER)
(20), and epidermal growth factor
receptor signaling molecules (12)
as its substrates. However, the mechanism through which GALNT6
promotes carcinogenesis is incompletely understood, since its
O-glycan substrates have not been fully elucidated.
The extracellular matrix protein lectin
galactoside-binding soluble 3 binding protein (LGALS3BP) is
initially purified and cloned as a ligand of the lactose-specific
S-type lectin galectin-3 (formerly Mac-2), which was identified as
a heavy glycosylated molecule in the tumor tissues and serum of
patients with lung, breast and ovarian cancer (21). LGALS3BP was also identified as a
tumor-associated antigen (90K) that is secreted in the serum in
various types of cancer (22-24).
Accumulating studies have revealed that elevated serum and tissue
levels of LGALS3BP are associated with poor prognosis in several
human cancers, including breast cancer, lymphoma, pleural
mesothelioma and non-small-cell lung cancer (25-32).
However, the molecular mechanism underlying glycosylated LGALS3BP
secretion in the serum and tissue of cancer patients has not yet
been fully elucidated.
The aim of the present study was to investigate
whether GALNT6 directly binds and O-glycosylates LGALS3BP,
thereby enhancing the secretion of LGALS3BP in breast cancer cells,
hoping that the findings may lead to the development of
GALNT6-based therapies and novel biomarkers for mammary tumors.
Materials and methods
Cell lines and culture conditions
Human breast cancer cell lines (ZR-75-1, BT-474,
SK-BR-3, HCC1569, HCC1954, MDA-MB-231, MDA-MB-468, BT-20, HCC38,
HCC70, HCC1395, HCC1806 and HCC1937), a mammary epithelial cell
line (MCF-10A) and HeLa cells were obtained from the American Type
Culture Collection. MCF-7 cells were obtained from the Health
Science Research Resources Bank. 293T cells and MDA-MB-453 cells
were provided by the RIKEN BioResource Research Center. All cell
lines were cultured under the corresponding depositors’
recommendations. The cell line stocks used in the present study had
been properly stored in liquid nitrogen. The morphology of these
cell lines was monitored by microscopy and confirmed that it was
maintained by comparing our images with the original morphological
images. No Mycoplasma contamination was detected in any of
the cultures using a Mycoplasma Detection kit (Takara Bio) and
MycoAlert™ Mycoplasma Detection Kit (Lonza).
Plasmids
The pCAGGSnHC vectors expressing wild-type (WT) and
inactivated mutant (H271D) GALNT6 in frame with an HA-tag at the
carboxyl terminus (GALNT6 WT-HA and GALNT6 H271D-HA) were described
previously (10). To construct the
LGALS3BP expression vector, the entire coding sequence of LGALS3BP
complementary DNA was amplified by reverse transcription
(RT)-polymerase chain reaction (PCR) analysis using KOD -Plus- DNA
polymerase (Toyobo Life Science) and cloned into the pCAGGSn3FC
expression vector in frame with a FLAG-tag at the carboxyl terminus
(LGALS3BP-FLAG). The following primer set was used for LGALS3BP-WT
amplification: 5′-ATA AGA ATG CGG CCG CAT GAC CCC TCC GAG
GCT C-3′ (forward) and 5′-CCGCTC GAG GTC CAC ACC TGA GGA GTT
G-3′ (reverse). The underlined sequences indicate the NotI
and XhoI sites, respectively. In addition to full-length
LGALS3BP (1-585), expression plasmids for the LGALS3BP deletion
mutants Del-1 (1-538) and Del-2 (1-560) were prepared by PCR. The
amplification conditions were as follows: 94°C for 2 min, followed
by 30 cycles of denaturing, annealing and extension (94°C for 15
sec, 55°C for 30 sec and 68°C for 2 min, respectively). The reverse
primers 5′-CCG CTC
GAG CTC GAA ATC GGT GAC GTC TGC CA-3′ (Del-1) and
5′-CCGCTC GAG GGG
GAA GGA GGA GGT GCT CTT CG-3′ (Del-2) were used with the forward
primer 5′-ATA AGA ATG CGG
CCG CAT GAC CCC TCC GAG GCT C-3′. The LGALS3BP alanine
substitution mutants Mut-1 (S546A), Mut-2 (T550A/N551A), Mut-3
(S552A/S553A), Mut-4 (S555A/T556A), Mut-5 (S557A/S558A), Mut-6
(S555A) and Mut-7 (T556A), which were point mutants of
LGALS3BP-Del-2 (1-560)-FLAG, and the T556A, T571A, T556A/T571A,
T579A, N580A, S581A, S582A and T556A/T571A/S582A mutants of
full-length LGALS3BP-FLAG, were generated by conventional
two-step mutagenesis PCR.
LGALS3BP-FLAG- and GALNT6-FLAG-expressing pQCXIP
(Takara Bio) retroviral vectors were generated using PCR primers.
The following primer set was used to amplify LGALS3BP-FLAG: 5′-ATA
AGA ATG CGG CCG CAT
GAC CCC TCC GAG GCT C-3′ (forward) and 5′-CCG GAT CCT TAC TTG TCG TCA TCG TCT TTG TAG TCG TCC
ACA CCT GAG GAG TTG-3′ (reverse). The underlined sequences
indicate the NotI and BamHI sites, respectively; the
double underlining indicates the FLAG-tag sequence. The following
primer set was used to amplify GALNT6-FLAG: 5′-GGG GCG GCC GCC ACC
ATG AGG CTC CTC CGC AGA CGC CA-3′ (forward) and 5′-CCG AAT TCT
CAC TAC TTG TCG TCA
TCG TCT TTG TAG TCG ACA AAG AGC CAC AAC TGA TGG G-3′
(reverse). The underlined sequences indicate the NotI and
BamHI sites, respectively; the double underlining indicates
the FLAG-tag sequence.
Transfection and viral infection
293T and HeLa cells were transfected with expression
vectors using FuGENE 6 (Promega Corporation). Plat-E cells were
kindly provided by Dr Toshio Kitamura (University of Tokyo) and
were co-transfected with retroviral vectors and pVSV-G using FuGENE
HD (Promega Corporation), according to the manufacturer’s
instructions. Culture supernatants were collected 48 h after
transfection. ZR-75-1 and HCC1806 cells were infected with the
culture supernatants in the presence of polybrene. Infected cells
were then selected in the presence of puromycin.
For gene silencing via RNA interference, ZR-75-1 and
BT-20 cells were transfected with siRNA oligonucleotides
(Sigma-Aldrich Japan KK) using Lipofectamine RNAiMAX (Thermo Fisher
Scientific, Inc.). The sequences targeting enhanced green
fluorescent protein (EGFP), GALNT6 #1,
LGALS3BP #1 or LGALS3BP #2 were as follows: 5′-GCA
GCA CGA CUU CUU CAA G-3′ for EGFP, 5′-GAG AAA UCC UUC GGU
GAC A-3′ for GALNT6 #1, 5′-GCG UGA ACG AUG GUG ACA U-3′ for
LGALS3BP #1, and 5′-UCA CCC UGU CGU CAG UCA A-3′ for
LGALS3BP #2.
Immunoprecipitation (IP)
Cells were lysed with lysis buffer [50 mM Tris-HCl
(pH 7.5), 150 mM NaCl, 5 mM 2-mercaptoethanol, 0.1% NP-40 and 0.5%
CHAPS]. Cell supernatants and cell lysates were immunoprecipitated
with anti-FLAG M2 affinity gel (Sigma-Aldrich; Merck KGaA).
Immunoprecipitated protein complexes were washed three times with
lysis buffer. Immunoprecipitates were separated using 10% SDS-PAGE
and analyzed by immunoblotting.
Immunoblotting analysis and silver
staining
The protein solutions were mixed with SDS-lysis
buffer [125 mM Tris-HCl (pH 6.8), 4% SDS, 20% glycerol, 0.04%
bromophenol blue and 10% 2-mercaptoethanol] to prepare the samples
for SDS-PAGE analysis. To denature the proteins, the mixed solution
was heated at 95°C for 5 min. Equal volumes of cell lysate (5
µl; 1% of whole-cell lysates, 10% of immune-precipitates)
were loaded and resolved on 10% SDS-PAGE gels. Lysates were
electrophoretically separated, blotted onto nitrocellulose
membranes, and blocked with 4% Block Ace solution (Dainippon
Pharmaceutical) for 3 h at room temperature. The membranes were
subsequently incubated for 12 h at 4°C with antibodies against the
following proteins: FLAG-tag (M2, mouse monoclonal, 1:5,000, cat.
no. F3165, Sigma-Aldrich; Merck KGaA), HA tag (3F10; rat
monoclonal, 1:3,000, cat. no. 11867423001, Roche Diagnostics GmbH),
LGALS3BP (rabbit polyclonal, 1:1,000, cat. no. NBP1-89346, Novus
Biologicals), GALNT6 (rabbit polyclonal, 1:500) (10) and actin (AC-15, mouse monoclonal,
1: 5,000, cat. no. A1978, Sigma-Aldrich; Merck KGaA). Following
incubation with a horseradish peroxidase (HRP)-conjugated secondary
antibody (anti-mouse IgG-HRP, polyclonal, 1:5,000, sc-2005;
anti-rat IgG-HRP, polyclonal, 1:5,000, sc-2006; anti-rabbit
IgG-HRP, polyclonal, 1:1,000, sc-2004; Santa Cruz Biotechnology,
Inc.), the blots were developed using an enhanced chemiluminescence
system (GE Healthcare) and scanned using an Image Reader LAS-3000
mini (Fujifilm). Vicia villosa agglutinin (VVA) lectin blot
analysis was performed using biotin-conjugated VVA lectin (cat. no.
BA-3601-1, EY Laboratories) and streptavidin-HRP (cat. no. 554066,
BD Biosciences), as described previously (10). Silver staining for 30 sec at room
temperature was performed using a Silver Stain MS Kit (Wako Pure
Chemical Industries, Ltd.). Full-length images of the immunoblots
are shown in the Supplementary
Figures.
2-Dimensional image-converted analysis of
liquid chromatography-mass spectrometry (LC-MS) (2DICAL)
Cell lines stably expressing GALNT6-HA or mock
vector were established as described previously (10). Cells were lysed in 0.5% NP-40
immunoprecipitation lysis buffer at 4°C for 30 min and were then
incubated with anti-HA agarose beads (Sigma-Aldrich; Merck KGaA) at
4°C for 16 h. After washing with the same buffer, the proteins
bound to the beads were eluted with 1% sodium deoxycholate buffer.
Subsequently, samples were prepared for LC-MS as described
previously (33).
Immunocytochemistry
Cells were fixed with 4% paraformaldehyde for 30 min
at 4°C and incubated with mouse anti-FLAG-tag M2 (Sigma-Aldrich;
Merck KGaA), rat anti-HA-tag (Roche Diagnostics GmbH), rabbit
anti-FLAG-tag (Sigma-Aldrich; Merck KGaA), mouse anti-Golgi-58k
(Sigma-Aldrich; Merck KGaA), and rabbit anti-GALNT6 (10) antibodies. The nuclei were
counterstained with 4′,6-diamidino-2-phenylindole (DAPI).
Fluorescent images were obtained with Olympus IX71 and FV3000
microscopes (Olympus Corporation).
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Expression of the GALNT6 and LGALS3BP
genes was evaluated by RT-qPCR. Total RNA was extracted, and
complementary DNA was subsequently synthesized. The complementary
DNA was analyzed by qPCR using a 7500 Real-Time PCR system (Thermo
Fisher Scientific, Inc.) and SYBR Premix ExTaq (Takara Bio)
according to the manufacturer’s instructions. The thermocycling
conditions were as follows: 10 min at 94°C, followed by 40 cycles
of denaturation at 94°C for 15 sec, 1 min annealing and extension
at 60°C, and reading of the fluorescence. Following
threshold-dependent cycling, melting curve analysis was performed
from 60 to 94°C. Gene expression in each sample was normalized to
the β2-MG mRNA content and quantified using the 2−∆∆Ct
method. The following primers were used: GALNT6, 5′-GCA GAG
GTC TGG ATG GAC AGC TA-3′ and 5′-GGT AGA CAT TGT GCA GGT ACC AG-3′;
LGALS3BP, 5′-GCC CCT TCT ACC TGA CCA A-3′ and 5′-TGA GTA GGG
CGA CAT CTG G-3′; GALNT3, 5′-ATG CCT TGC TCT GTT GTT GG-3′
and 5′-TGG TTT GCC TCC TTG ATT GT-3′; and β2-MG, 5′-AAC TTA
GAG GTG GGG AG CAG-3′ and 5′-CAC AAC CAT GCC TTA CTT TAT C-3′.
Determination of LGALS3BP in the culture
supernatant by immunoblotting analysis
The extracellular release of LGALS3BP from the cells
were evaluated by immunoblotting. Briefly, ZR-75-1, BT-20 and
SK-BR-3 cells were transfected with siRNA against GALNT6 or
LGALS3BP, and the culture medium was changed to 0.1% FBS in medium
at 48 h post-transfection. After 24 h, the culture medium was
collected and filtered, followed by acetone precipitation at −30°C
for 2 h, and subsequently centrifuged at 22,000 × g for 30 min. The
pellets were dried and lysed with SDS-lysis buffer [125 mM Tris-HCl
(pH 6.8), 4% SDS, 20% glycerol, 0.04% bromophenol blue and 10%
2-mercaptoethanol]. LGALS3BP in the culture medium was determined
using immunoblotting.
Effect of soluble LGALS3BP on the
proliferation of HCC1806 cells
HeLa and 293T cells were transfected with GALNT6-HA
construct (WT and H271D mutant) and LGALS3BP-FLAG construct. After
24 h, the culture medium was changed to 1% FBS or 2% FBS in medium
of HeLa and 293T cells, respectively, and cultured for 72 h. The
culture medium was collected and filtered (pore size, 0.22
µm), followed by replacement of each culture medium with the
medium of HCC1806 cells. After further culture for 72 and 96 h,
cell proliferation was assessed as described below.
LGALS3BP ELISA
The LGALS3BP protein levels in the culture
supernatants were quantified using a human galectin-3BP/MAC-2BP
Quantikine ELISA Kit (R&D Systems, Inc.) according to the
manufacturer’s instructions.
Cell proliferation assay
WST-8 assays were performed using a Cell Counting
Kit-8 (Dojindo Molecular Technologies, Inc.). A 1:10 dilution of
Cell Counting Kit-8 solution was added to the cells and incubated
for 1 h. Subsequently, the absorbance at 450 nm was measured to
calculate the number of viable cells per well.
In vitro glycosylation assay
In vitro GALNT6 activity toward LGALS3BP as a
substrate was assessed as follows (34): Various concentrations of
recombinant GALNT6 protein (codons 35-622) were incubated with 2
µM biotin-tagged LGALS3BP peptide (568-585, WT, T571A, S582A
and T571A/T582A) and UDP-GalNAc for 3 h at room temperature. The
reactions were terminated by the addition of 0.5 M EDTA. These
reaction mixtures were then transferred into NeutrAvidin-coated
plates (Thermo Fisher Scientific, Inc.) and incubated for 1 h at
room temperature. After the samples were washed with PBS, the
α-GalNAc-specific lectin Helix pomatia agglutinin conjugated
to HRP (HPA-HRP, EY Laboratories) was added and subsequently
incubated for 30 min at room temperature. After washing, bound
HPA-HRP was quantified by incubation with
3,3′,5,5′-tetramethylbenzidine peroxide solution (Thermo Fisher
Scientific, Inc.) for 30 min in the dark at room temperature. After
HRP activity was terminated with sulfuric acid, the absorbance at
415 nm was measured using a microplate reader (Infinite M200,
Tecan).
Proteasome-dependent degradation of
LGALS3BP
ZR-75-1 cells were transfected with EGFP or GALNT6
#1 siRNA. After 48 h of transfection, cells were treated with 10
µM MG132 (EMD/Merck KGaA), and harvested at 6 and 12 h after
treatment. Immunoblotting analyses were performed using the
indicated antibodies.
Analysis of public database
The data for GALNT6 and LGALS3BP mRNA expression
(RNA Seq V2 RSEM) in primary invasive breast carcinoma samples were
downloaded from The Cancer Genome Atlas (TCGA) network (http://cancergenome.nih.gov/). Kaplan-Meier plots for
breast cancer patients were analyzed by the Kaplan-Meier plotter
(KM plotter) database (http://kmplot.com/analysis/). Breast cancer patients
in the KM plotter database were stratified into 'low’ and 'high’
GALNT6 mRNA expression groups (Affymetrix ID: 219956_at) based on
an autoselected best cutoff value.
Statistical analysis
Data are expressed as the means ± standard deviation
(SD). The Student’s t-test was used for comparisons between two
groups, and one-way analysis of variance with Tukey or Dunnett’s
post hoc test was applied for multiple comparison. P-values
<0.05 were considered to indicate a statistically significant
difference. The statistical significance of TCGA datasets was
calculated using the Kruskal-Wallis test and Dunnett’s post-hoc
test with SPSS version 20.0 (IBM Corp.). GALNT6 and
LGALS3BP expression in ER-positive or human epidermal growth
factor receptor 2 (HER2)-positive samples and a corresponding
normal sample were compared using two-tailed Student’s t-test.
Results
GALNT6 expression is correlated with
breast cancer prognosis
GALNT6 expression has been reported to be
upreg-ulated in breast cancer cells (7-10).
The association between theGALNT6 expression level and
prognosis for each breast cancer subtype was further investigated
via analysis of TCGA public datasets. GALNT6 expression was
found to be higher in patients with ER-positive and HER2-positive
invasive breast cancer compared with that in patients with
ER-negative or triple-negative breast cancer (Fig. S1A). Moreover, it was confirmed
that GALNT6 was expressed at high levels in breast cancer
compared with corresponding normal breast control samples (n=110;
Fig. S1B), as reported previously
(7,9,10).
Of note, GALNT6 was significantly upregulated in ER- (n=34)
and HER2-positive (n=9) breast cancer compared with the
corresponding normal controls (Fig.
S1C). More importantly, KM plotter database analysis revealed
that high expression levels of GALNT6 were correlated with shorter
duration of relapse-free survival (P=0.006), distant
metastasis-free survival (P=0.00084) and overall survival
(P=0.0036) across all breast tumor types (Fig. S1D). Notably, high GALNT6
expression was positively correlated with reduced relapse-free
survival duration compared with that of patients with low
GALNT6 expression in patients with the ER-positive
(P=0.0038) and HER2-positive (P=0.032) subtypes of breast cancer
(Fig. S1E). In addition, in the
ER-negative subtype, high GALNT6 expression was also
associated with reduced relapse-free survival duration (P=0.017;
Fig. S1E). These findings
strongly support the results of previous studies (7-10)
reporting that GALNT6 plays a crucial role in mammary
carcinogenesis and progression of breast cancer, regardless of the
ER or HER2 expression status, although GALNT6 expression was
found to be higher in patients with ER-positive and HER2-positive
status among all breast tumor types.
Identification of LGALS3BP as a substrate
of GALNT6
To further investigate unknown effects of GALNT6 on
mammary carcinogenesis and progression of breast cancer,
GALNT6-binding protein(s) as substrate(s) were investigated by
2DICAL analysis (35) using a
previously established stable cell line (HeLa-GALNT6-WT) (10,17)
(Fig. S2A and B). LGALS3BP (also
referred to as Mac-2-binding protein or tumor-associated antigen
90K) was identified as a candidate GALNT6-binding protein, as four
different peptides derived from the LGALS3BP protein were detected
along with three peptides from the GALNT6 protein (Table I). Moreover, LGAL3BP has been
identified as a heavy glycosylated molecule in tumor tissues and
serum from patients with lung, breast and ovarian cancers (21). Therefore, we focused on LGALS3BP as
a strong candidate GLANT6-binding protein.
 | Table I2DICAL analysis for GALNT6-binding
proteins in GALNT6-HA-expressing HeLa cells vs. mock-HeLa
cells. |
Table I
2DICAL analysis for GALNT6-binding
proteins in GALNT6-HA-expressing HeLa cells vs. mock-HeLa
cells.
| Protein name | Expected value | Peptide
sequence |
|---|
| BANF1 | 6.E-09 |
HRDFVAEPMGEKPVGSLAGIGEVLGK |
| BANF1 | 5.E-08 |
DFVAEPMGEKPVGSLAGIGEVLGK |
| TPP1 | 8.E-08 | LYQQHGAGLFDVTR |
| GALNT6 | 4.E-07 |
NSQVPKDEEWELAQDQLIR |
| KPNB1 | 4.E-07 | VLANPGNSQVAR |
| VIM | 4.E-06 | SLYASSPGGVYATR |
| GALNT6 | 9.E-06 | LAEVWMDSYKK |
| VIM | 1.E-05 | LQDEIQNMKEEMAR |
| TPP1 | 1.E-05 | LFGGNFAHQASVAR |
| JUP | 2.E-05 | ALMGSPQLVAAVVR |
| VIM | 2.E-05 | NLQEAEEWYK |
| VIM | 9.E-05 | ILLAELEQLK |
| JUP | 0.00011 | AAMIVNQLSK |
| JUP | 0.00023 | LLNDEDPVVVTK |
| TPP1 | 0.00028 | ILSGRPPLGFLNPR |
| JUP | 0.00031 | EGLLAIFK |
| GALNT6 | 0.00044 | SFGDISER |
| LGALS3BP | 0.00066 | AVDTWSWGER |
| LGALS3BP | 0.00075 | SDLAVPSELALLK |
| KPNB1 | 0.0025 | WLAIDANAR |
| IPO7 | 0.0026 | AIFQTIQNR |
| LGALS3BP | 0.0031 | ASHEEVEGLVEK |
| JUP | 0.0059 | LAEPSQLLK |
| BANF1 | 0.0067 | KDEDLFR |
| KPNB1 | 0.008 | VAAGLQIK |
| BHLHA9 | 0.024 | GAPGLGLTAR |
| SCFD1 | 0.027 | IMSKTTLDK |
| LGALS1 | 0.029 | SFVLNLGK |
| JUP | 0.039 | SGGIPALVR |
| LGALS3BP | 0.045 | VEIFYR |
The expression levels of GALNT6 and
LGALS3BP were initially examined in 15 breast cancer cell
lines and the normal epithelial cell line MCF 10A by qPCR analysis,
and high expression levels of both genes were found in the ZR-75-1,
BT-20 and SK-BR-3 cell lines (Fig.
S2C). To validate the 2DICAL results, LGALS3BP-FLAG was
transfected into ZR-75-1 cells, and cell lysates were
immunoprecipitated with an anti-FLAG antibody. Subsequent western
blot analysis with the indicated antibodies demonstrated that
exogenous LGALS3BP-FLAG coimmunoprecipitated with endogenous GALNT6
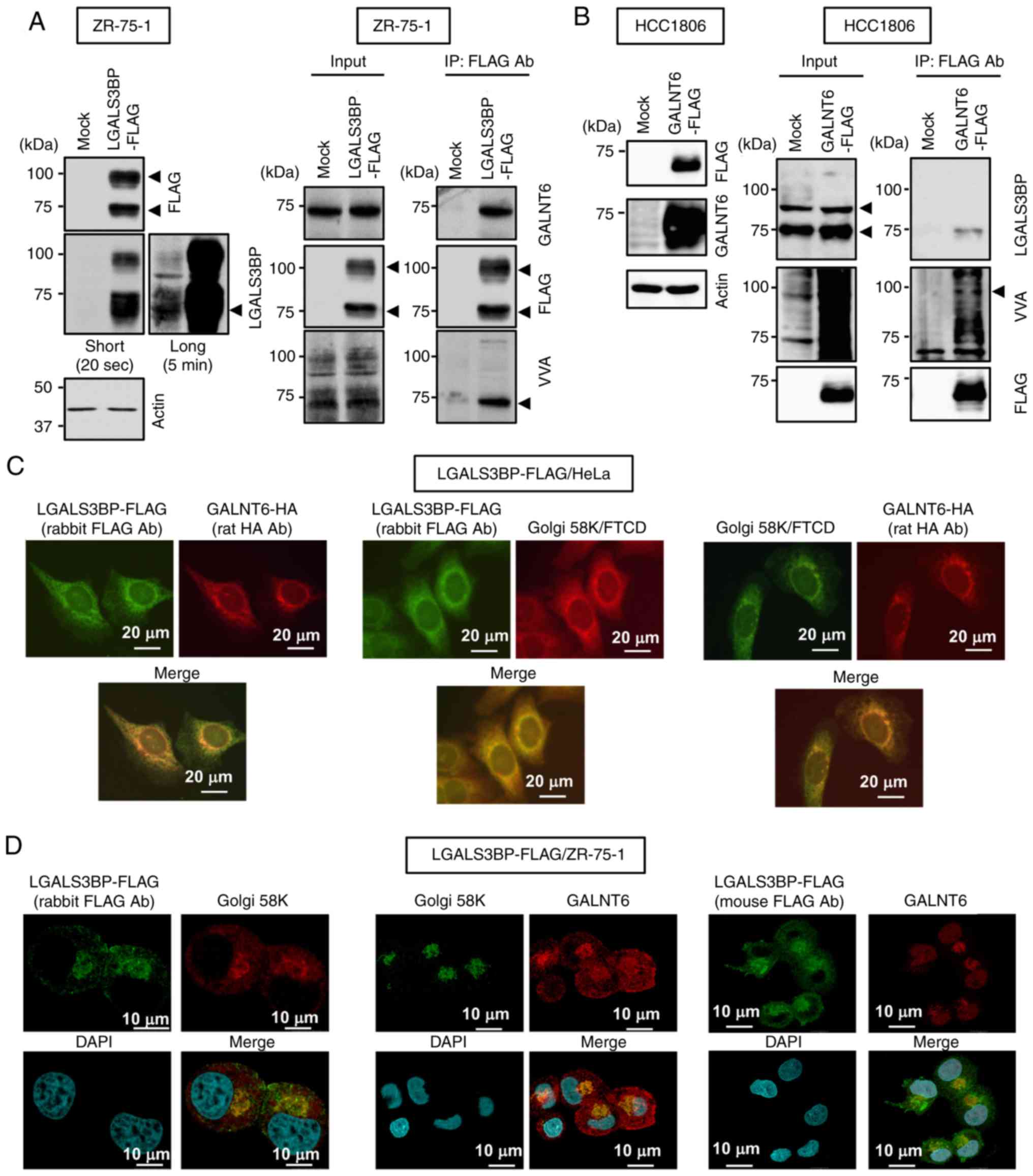
in ZR-75-1 cells (Fig. 1A).
Conversely, it was confirmed that GALNT6-FLAG coimmunoprecipitated
with endogenous LGALS3BP in HCC1806 cells, which express low levels
of GALNT6 (Fig. 1B). Notably, the
~75-kDa band of O-glycosylation of coimmunoprecipitated
LGALS3BP-FLAG with endogenous GALNT6 was observed by VVA lectin
blot analysis (Fig. 1A). By
contrast, the ~90-kDa band of O-glycosylated LGALS3BP when
GALNT6-FLAG was exogenously overexpressed was observed by VVA
lectin blot analysis (Fig. 1B).
These findings suggest that GALNT6 interacts with and
O-glycosylates LGALS3BP in breast cancer cells.
To further validate the GALNT6-LGALS3BP interaction,
the subcellular localization of these proteins in breast cancer
cells was examined by immunocytochemistry. Since GALNT6 is
reportedly localized in the Golgi apparatus of breast cancer cells
(10,17) and LGALS3BP is reported to be a
heavy glycosylated secreted protein (21-24),
their colocalization in the Golgi apparatus of cancer cells was
investigated. Both LGALS3BP-FLAG and GALNT6-HA were clearly
observed in the Golgi apparatus in HeLa cells, as evaluated by the
Golgi marker Golgi-58k (Fig. 1C).
Moreover, it was confirmed that stably expressed LGALS3BP-FLAG
colocalized with endogenous GALNT6 in the Golgi apparatus in
ZR-75-1 cells (Fig. 1D), which
suggests that GALNT6 interacts and colocalizes with LGALS3BP in the
Golgi apparatus in breast cancer cells.
Effects of GALNT6 on the secretion and
O-glycosylation of LGALS3BP
As LGALS3BP is reported to be a secreted galectin-3
ligand glycoprotein in several tumors, including breast cancer
(21-25,31,32),
the effect of GALNT6-dependent LGALS3BP O-glycosylation on
LGALS3BP secretion was examined by VVA lectin and western blot
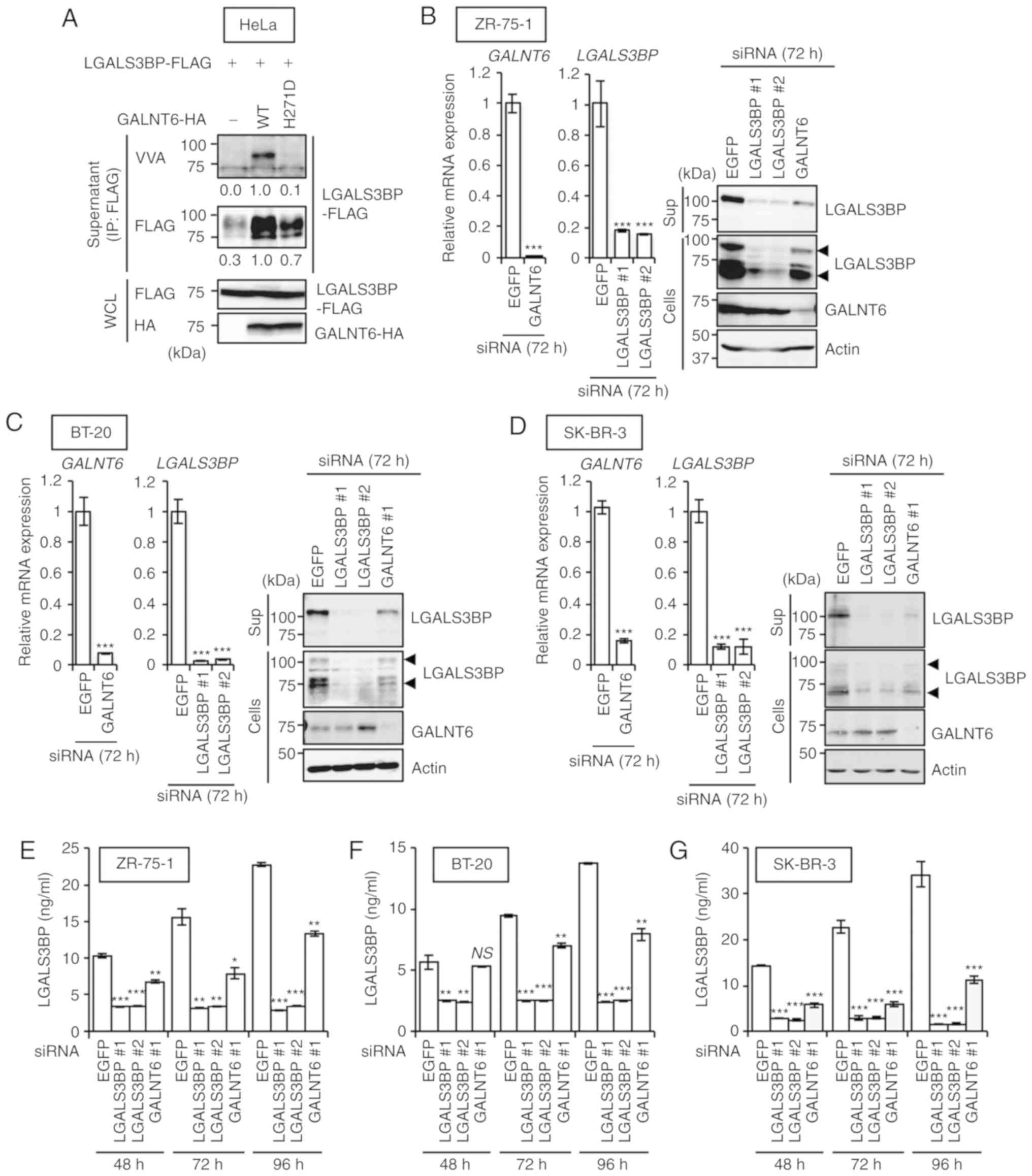
analyses. It was confirmed that both GALNT6 WT-HA and the
enzyme-dead GALNT6 H271D-HA mutant interacted with LGALS3BP-FLAG in
293T cells (Fig. S3A). However,
overexpression of GALNT6 WT-HA, but not GALNT6 H271D-HA, induced
the O-glycosylation of LGALS3BP-FLAG in the supernatant of
transfected HeLa cells (Fig.
2A).
 | Figure 2Roles of GALNT6 in the secretion and
O-glycosylation of LGALS3BP. (A) GALNT6-HA (WT or the
enzyme-dead H271D mutant) stably transfected HeLa cells were
transfected with the LGALS3BP-FLAG expression plasmid vector. After
72 h, cell supernatants and cell lysates were harvested and
immuno-precipitated with anti-FLAG affinity gel. Then,
immunoblotting analyses were performed using the indicated
antibodies and VVA-lectin. The band densities on the immunoblots
are expressed relative to those of cells cotransfected with the
LGALS3BP-FLAG and GALNT6 WT-HA expression plasmids, which were
defined as 1.0. (B-D) The human breast cancer cell lines (B)
ZR-75-1, (C) BT-20 and (D) SK-BR-3 were transfected with EGFP,
GALNT6 #1, LGALS3BP #1 or LGALS3BP #2 siRNA. The GALNT6 and
LGALS3BP mRNA expression levels were evaluated by
quantitative PCR at 72 h after transfection. The levels were
expressed relative to those in EGFP siRNA-transfected cells, which
were defined as 1.0 (left). The protein levels of LGALS3BP, GALNT6
and actin in the extracellular (Sup: Acetone precipitation) or
intracellular (Cells) compartment at 72 h after transfection were
determined by immunoblot analysis (right). The data are presented
as the means ± standard deviation of three independent experiments
(***P<0.001 vs. EGFP siRNA-transfected cells). (E-G)
The LGALS3BP protein levels in the culture supernatants of (E)
ZR-75-1, (F) BT-20 and (G) SK-BR-3 cells at 48, 72 and 96 h after
transfection were determined by LGALS3BP ELISA (Materials and
methods). The data are presented at the means ± standard deviation
of three independent experiments (**P<0.01,
***P<0.001 vs. EGFP siRNA-transfected cells in each
time). NS, not significant. GALNT6, polypeptide
N-acetylgalactosaminyltransferase 6; LGALS3BP, lectin
galactoside-binding soluble 3 binding protein; WT, wild-type; VVA,
Vicia villosa agglutinin; WCL, whole-cell lysates; EGFP,
enhanced green fluorescent protein. |
To further validate the results mentioned above, the
effects of siRNA-mediated GALNT6 knockdown on the
O-glycosylation and secretion of LGALS3BP in the ZR-75-1,
BT-20 and SK-BR-3 cell lines were examined by immunoblot-ting and
ELISA. As GALNT6 exhibits the highest structural and amino acid
sequence similarity with GALNT3 among GALNT family members, we
first examined the specificity of GALNT6 knockdown on
GALNT3 expression in BT-20 cells by qPCR. The results
demonstrated that GALNT6 knockdown does not affect
GALNT3 expression at the transcriptional level (Fig. S3B). Subsequently, immunoblotting
and ELISA demonstrated that silencing GALNT6 markedly
reduced both the intracellular and extracellular release of
LGALS3BP into the culture supernatant of ZR-75-1 (Fig. 2B and E), BT-20 (Fig. 2C and F) and SK-BR-3 (Fig. 2D and G) cells, compared with
siEGFP-treated cells. In addition, it was confirmed that the
knockdown of GALNT6 did not affect LGALS3BP
expression at the transcriptional level in ZR-75-1 cells (Fig. S3C). It was further confirmed that
degradation of the LGALS3BP protein was still observed in
GALNT6-depleted cells, despite the presence of MG132 (Fig. S3D). These data suggest that the
reduction of GALNT6-mediated O-glycosylation may cause
degradation of the LGALS3BP protein through a
proteasome-independent proteolysis pathway. Therefore, GALNT6 may
regulate LGALS3BP protein stabilization through
O-glycosylation in breast cancer cells.
Oncogenic effects of the GALNT6-LGALS3BP
axis on the proliferation of breast cancer cells
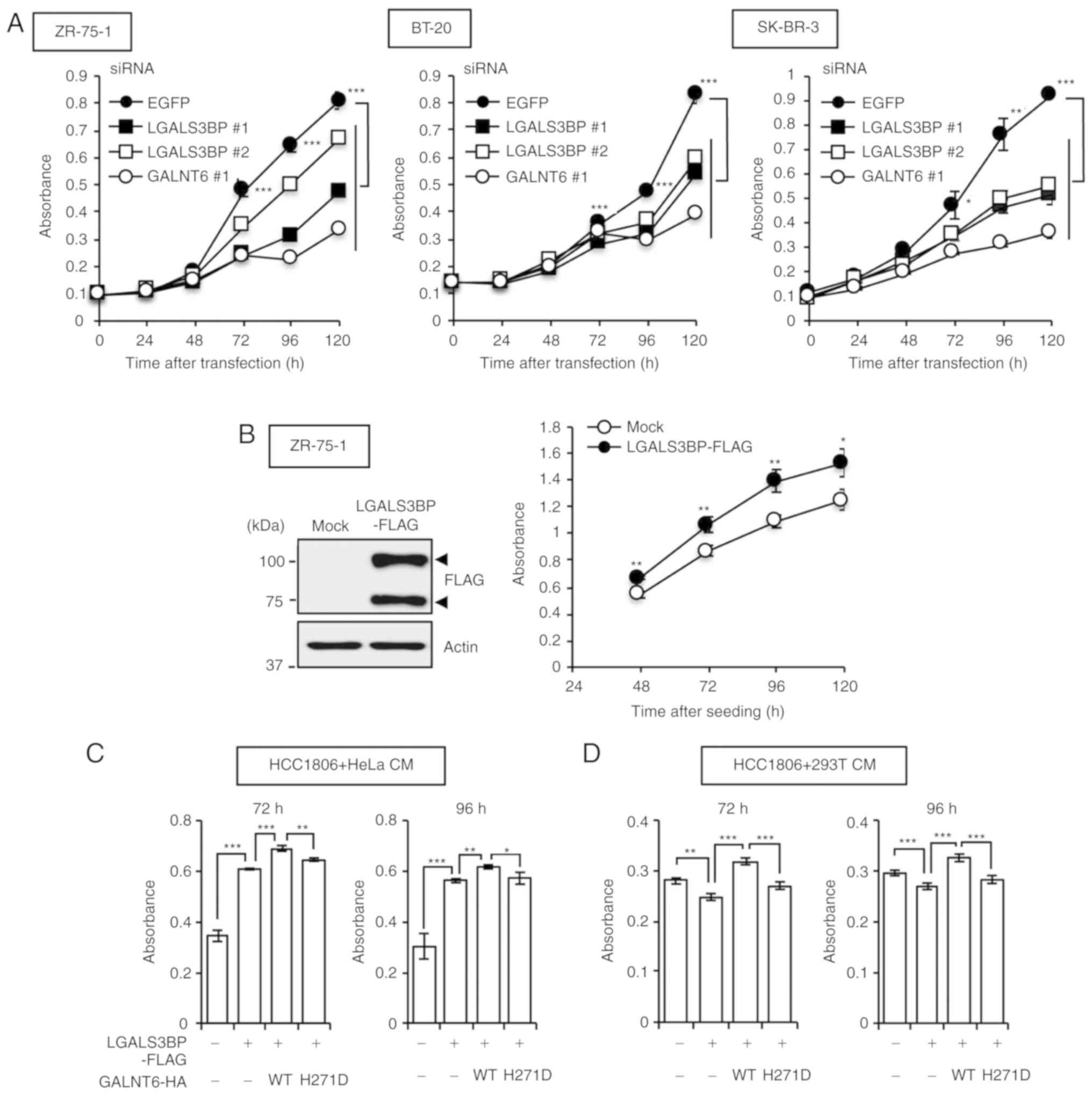
We then sought to investigate the effect of GALNT6
and LGALS3BP on the proliferation of breast cancer cells. siRNAs
were first used to knock down endogenous GALNT6 and LGALS3BP
expression in ZR-75-1, BT-20 and SK-BR-3 cells. Silencing of
GALNT6 and LGALS3BP inhibited the proliferation of
ZR-75-1, BT-20 and SK-BR-3 cells (Fig.
3A) from 72 h after each siRNA transfection. Conversely, stable
expression of LGALS3BP-FLAG in ZR-75-1 cells led to marked
enhancement of cell growth compared with that in mock-transfected
ZR-75-1 cells from 48 h after seeding (Fig. 3B). Subsequently, it was
hypothesized that GALNT6 overexpression enhances the release of
O-glycosylated LGALS3BP from breast cancer cells, thereby
promoting the proliferation of these cells. Therefore, we next
investigated the effect of soluble LGALS3BP on cell proliferation
in breast cancer cell lines. We further analyzed cell proliferation
in breast cancer cell lines using conditioned medium (CM) from
LGALS3BP-FLAG-overexpressing (LGAL3SBP+) and
GALNT6-HA-overexpressing (GALNT6 WT+) HeLa or 293T
cells. Treatment with CM from LGAL3SBP+ HeLa cells
significantly promoted HCC1806 cell proliferation (Fig. 3C). Notably, CM from
LGAL3SBP+/GALNT6 WT+ HeLa and 293T cells
significantly enhanced the proliferation of HCC1806 cells in a
time-dependent manner compared with the effect of CM from
LGAL3SBP+/GALNT6− or
LGAL3SBP+/GALNT6 H271D+ HeLa and 293T cells
(Fig. 3C and D). These findings
indicate that the O-glycosylation of LGALS3BP by GALNT6 is
crucial for the enhancement of breast cancer cell
proliferation.
 | Figure 3Effect of GALNT6-dependent LGALS3BP
O-glycosylation on cell growth. (A) Cell proliferation
following transfection with EGFP, GALNT6 #1, LGALS3BP #1 or
LGALS3BP #2 siRNA was assessed in ZR-75-1 (left), BT-20 (middle)
and SK-BR-3 (right) cells at 0, 24, 48, 72, 96 or 120 h after
transfection via a WST-8 assay. The data are presented as the means
± standard deviation of three independent experiments
(*P<0.05, **P<0.01,
***P<0.001 vs. EGFP siRNA-transfected cells). (B) The
proliferation of ZR-75-1 cells stably expressing Mock or
LGALS3BP-FLAG vectors (left) was assessed at 48, 72, 96 and 120 h
after seeding via a WST-8 assay (right). The data are presented as
the means ± standard deviation of three independent experiments
(*P<0.05, **P<0.01 vs. mock-transfected
ZR-75-1 cells). (C) HeLa or (D) 293T cells were cotransfected with
the LGALS3BP-FLAG and GALNT6-HA expression plasmid vectors. After
24 h, the medium was replaced with (C) 1% FBS/RPMI or (D) 2%
FBS/RPMI medium, and CM was harvested at 72 h after the medium
change. Next, the medium of HCC1806 cells was replaced with CM from
(C) HeLa or (D) 293T cells, and proliferation was assessed at 72
and 96 h after the medium change via a WST-8 assay. The data are
presented as the means ± standard deviation of three independent
experiments (*P<0.05, **P<0.01,
***P<0.001). NS, not significant; CM, conditioned
medium; GALNT6, polypeptide N-acetylgalactosaminyltransferase 6;
LGALS3BP, lectin galactoside-binding soluble 3 binding protein;
EGFP, enhanced green fluorescent protein. |
The carboxyl-terminal region of LGALS3BP
regulates GALNT6-dependent LGALS3BP O-glycosylation and
release
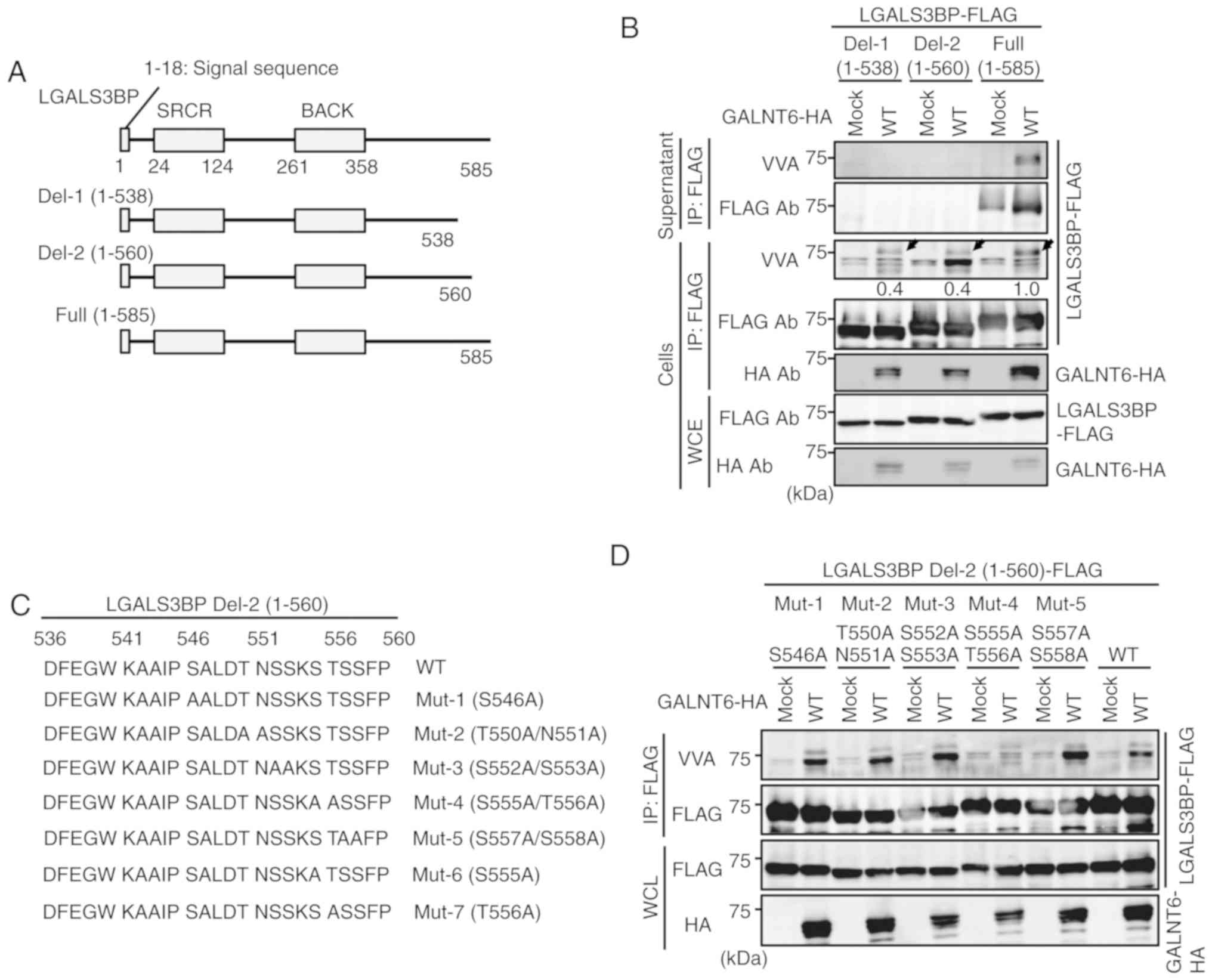
To investigate the detailed biological significance
of GALNT6-dependent LGALS3BP O-glycosylation, we first
attempted to determine the critical region(s) required for the
O-glycosylation and extracellular release of LGALS3BP.
Accumulating functional and structural studies have demonstrated
that the LGALS3BP protein contains an N-terminal signal sequence, a
scavenger receptor cysteine-rich (SRCR) domain for protein-protein
interaction, and a BTB and C-terminal Kelch (BACK) domain for
interaction with the Cullin3 ubiquitin ligase complex and
homo-oligomerization (Fig. 4A)
(21,22,36,37).
As the function of the C-terminal region of LGALS3BP remains
unclear, we focused on this region. Plasmids expressing LGALS3BP
C-terminal region deletion mutants [Del-1 (1-538) and Del-2 (1-560)
(Fig. 4A)] were generated and both
mutants were first cotransfected independently with GALNT6-HA into
293T cells. Next, immunoprecipitation with an anti-FLAG antibody
followed by immunoblotting with the indicated antibodies and VVA
lectin blot analyses were performed. Both LGALS3BP deletion mutants
interacted with GALNT6-HA and were O-glycosylated in the
intracellular fraction of 293T cells (Fig. 4B). Importantly, extracellular
release of LGALS3BP was not observed in the supernatant fraction of
the cells transfected with both LGALS3BP deletion mutants and
GALNT6-HA (Fig. 4B), indicating
that the region between amino acids 561 and 585 of LGALS3BP is the
minimal requirement for the extracellular release of LGALS3BP.
Moreover, the band of O-glycosylated LGAL3SBP was observed
in the supernatant from only the cells transfected with full-length
LGALS3BP and GALNT6-HA by VVA lectin blot analysis (Fig. 4B).
 | Figure 4The carboxyl-terminal region of
LGALS3BP regulates GALNT6-dependent LGALS3BP O-glycosylation
and release. (A) Schematic diagram of LGALS3BP and deletion mutants
(Del-1, Del-2 and Full). (B) 293T cells were cotransfected with the
LGALS3BP deletion mutant-FLAG (Del-1, Del-2 or Full) and GALNT6-HA
expression plasmid vectors. After 72 h, cell supernatants and cell
lysates (Cells) were harvested and immunoprecipitated with
anti-FLAG affinity gel. Immunoblotting was performed using the
indicated antibodies and VVA-lectin. Arrowheads, potential
O-glycosylation of intracellular LGALS3BP-FLAG protein. (C)
Schematic diagram of the LGALS3BP Del-2 (1-560) point mutants
(Mut-1, -2, -3, -4, -5, -6 and -7). (D) 293T cells were
cotransfected with the LGALS3BP Del-2 (1-560) point mutant-FLAG
(Mut-1, -2, -3, -4 or -5) or the WT (1-560)-FLAG expression plasmid
and GALNT6-HA expression plasmids. After 72 h, cell lysates were
harvested and immunoprecipitated with anti-FLAG affinity gel. (E)
293T cells were cotransfected with either the LGALS3BP Del-2
(1-560) point mutant-FLAG (Mut-4, -6 or -7) or the WT (1-560)-FLAG
expression plasmid and GALNT6-HA expression plasmids. After 72 h,
cell lysates were harvested and immunoprecipitated with anti-FLAG
affinity gel. (F) 293T cells were cotransfected with either the
LGALS3BP Del-2 (1-560) T556A, Del-2 (1-560) WT, full-length T556A
or full-length WT-FLAG and the GALNT6-HA expression plasmid
vectors. After 72 h, cell supernatants and cell lysates (Cells)
were harvested and immunoprecipitated with anti-FLAG affinity gel.
The band densities on the immunoblot are expressed relative to
those of cells cotransfected with the LGALS3BP full-length WT-FLAG
and GALNT6 WT-HA expression plasmid vectors, which were defined as
1.0. GALNT6, polypeptide N-acetylgalactosaminyltransferase 6;
LGALS3BP, lectin galactoside-binding soluble 3 binding protein;
WCL, whole-cell lysates; SRCR, scavenger receptor cysteine-rich;
BACK, BTB and C-terminal Kelch; WT, wild-type; VVA, Vicia
villosa agglutinin. |
The region(s) required for GALNT6-dependent LGALS3BP
O-glycosylation were next investigated. As shown in Fig. 4B, the GALNT6-dependent
O-glycosylation level of Del-1 and Del-2 determined by VVA
in the intracellular fraction (cells) was obviously reduced (60%)
compared with that of WT, as indicated by the arrowheads. This
result indicates that both the regions between amino acids 1-560
and 561-585 may be required for GALNT6-dependent LGALS3BP
O-glycosylation. Therefore, we first focused on the Del-2
mutant (amino acids 1-560) of LGALS3BP to identify GALNT6-dependent
O-glycosylation sites. Because the Del-2 region contains
several candidate serine and threonine residues for
O-glycosylation, which are highly conserved among several
species (Fig. S4), Del-2 mutant
constructs were prepared, in which each of these candidate residues
was separately substituted with alanine (Fig. 4C). Notably, Mut-4 (S555A/T556A)
markedly reduced GALNT6-dependent O-glycosylation of Del-2
(Fig. 4D). Next, to further narrow
down the GALNT6-dependent O-glycosylation site(s) within
Del-2 region, Mut-6 (S555A) and Mut-7 (T556A) constructs were
generated (Fig. 4C). It was
confirmed that Mut-7 (T556A), as well as Mut-4 (S555A/T556A),
mostly abolished GALNT6-dependent O-glycosylation of
LGALS3BP (Fig. 4E), suggesting
that T556 is a strong candidate site for GALNT6-dependent
O-glycosylation of LGALS3BP.
Finally, we attempted to validate this result using
the T556A mutant plasmid vector from the full-length LGALS3BP
sequence. Unexpectedly, a strong band representing GALNT6-dependent
LGALS3BP O-glycosylation was observed in the supernatant
fraction from GALNT6 WT-transfected cells via VVA lectin blot
analysis (Fig. 4F), suggesting
that the region encompassing amino acids 561-585 contains at least
one other site putatively O-glycosylated by GALNT6.
Furthermore, a slight reduction of the LGALS3BP protein secretion
level was observed in the supernatant from both LGALS3BP-T556A-and
mock-transfected cells, suggesting that, in addition to T556, at
least one other putative GALNT6-dependent O-glycosylation
site may be located within the region encompassing amino acids
561-585 of LGALS3BP.
Identification of GALNT6-dependent
LGALS3BP O-glycosylation, which is required for LGALS3BP
secretion
We further attempted to narrow down the site(s)
required for additional O-glycosylation site(s) of secreted
LGALS3BP within the region encompassing amino acids 561-585, and
the first focus was on T571 (Fig.
5A). The results demonstrated that the double alanine
substitution of T556/T571 (T556A/T571A) markedly attenuated the
levels of O-glycosylation and extracellular release of
LGALS3BP compared with those of the full-length WT protein in
GALNT6-WT-transfected HeLa cells (Fig.
5B). Moreover, T556A/T571A resulted in substantial attenuation
compared with that resulting from the T571A mutation (Fig. S5A). These results indicate that
T556 and T571 are strong candidate GALNT6-dependent
O-glycosylation sites critical for extracellular release of
LGALS3BP.
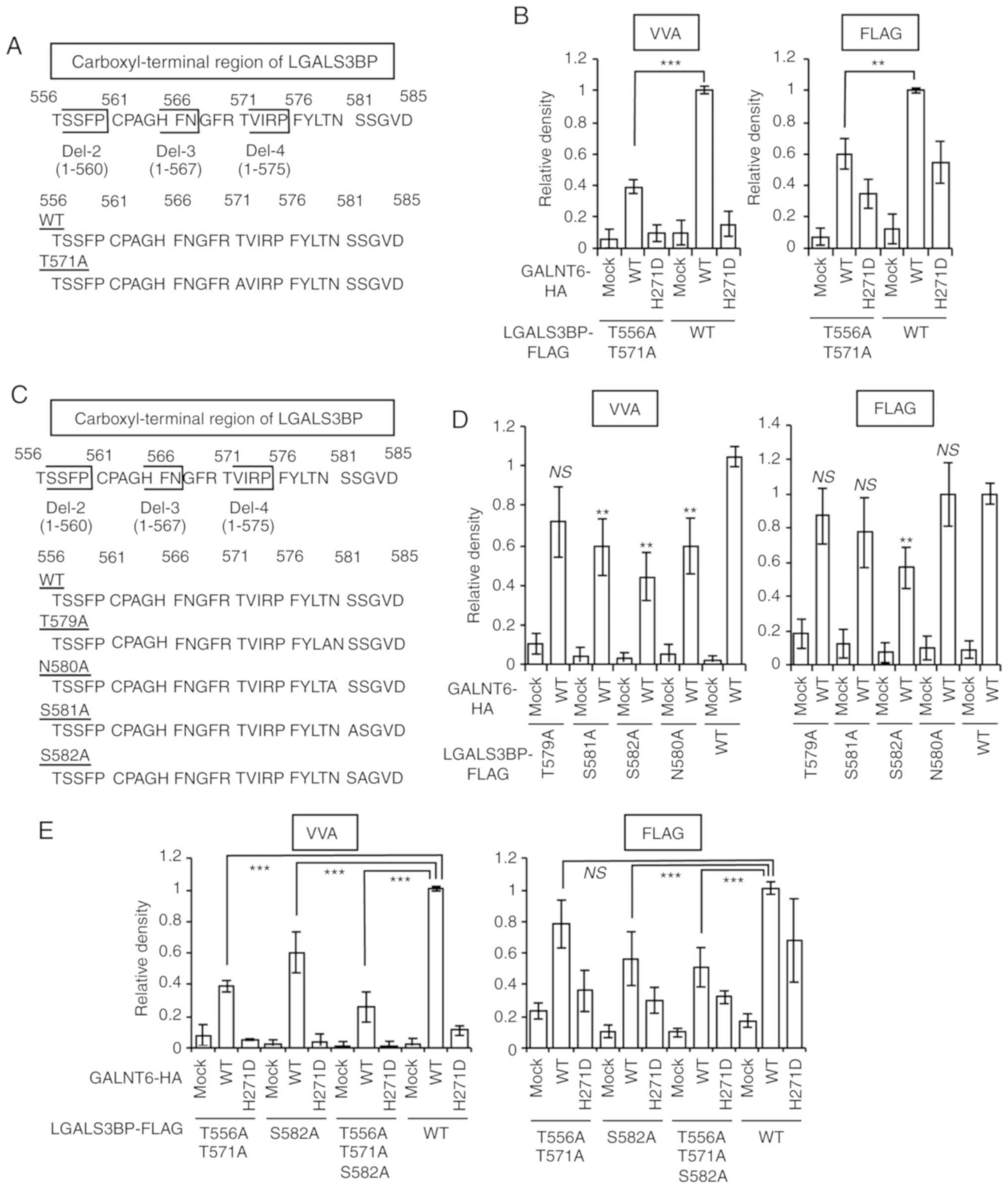 | Figure 5Identification of GALNT6-dependent
LGALS3BP O-glycosylation, which is required for LGALS3BP
secretion. (A) Schematic diagram of the LGALS3BP T571A point
mutant. (B) HeLa cells were cotransfected with either the LGALS3BP
T556A/T571A or WT-FLAG and the GALNT6-HA expression plasmid
vectors. After 72 h, cell supernatants and cell lysates were
harvested and immunoprecipitated with anti-FLAG affinity gel. The
band densities on the immunoblot are expressed relative to those of
cells cotransfected with the LGALS3BP WT-FLAG and GALNT6 WT-HA
expression plasmid vectors, which were defined as 1.0. The data are
presented as the means ± standard deviation of three independent
experiments (***P<0.001). (C) Schematic diagram of
the LGALS3BP T579A, S581A and S582A and N580A (a negative control
site for O-glycosylation; a potential N-glycosylation site)
point mutants. (D and E) HeLa cells were cotransfected with the
LGALS3BP point mutant-FLAG and GALNT6-HA expression plasmids. After
72 h, cell supernatants were harvested and immunoprecipitated with
anti-FLAG affinity gel. The band densities on the immunoblot are
expressed relative to those of cells cotransfected with the
LGALS3BP WT-FLAG and GALNT6 WT-HA expression plasmid vectors, which
were defined as 1.0. The data are presented at the means ± standard
deviation of three independent experiments
(**P<0.001, ***P<0.001; NS, not
significant) Identification of GALNT6-dependent LGALS3BP
O-glycosylation, which is required for LGALS3BP secretion.
(F) GALNT6 O-glycosylates the LGALS3BP protein in
vitro. In vitro O-glycosylation of the LGALS3BP-WT,
T571A, S582A and T571A+ S582A peptides (amino acids 568-585) by
WT-GALNT6 recombinant proteins after a 16 h reaction. The data are
presented as the means ± standard deviation of three independent
experiments (***P<0.001). (G) HeLa cells were
cotransfected with the LGALS3BP-FLAG and GALNT6-HA expression
plasmids. After 24 h, the medium was replaced with 1% FBS/RPMI
medium, and the CM was harvested at 72 h after the medium change.
Next, the medium of the HCC1806 cells was replaced with HeLa cell
CM, and cellular proliferation was determined using a WST-8 assay
at 96 h after the medium change. The data are presented as the
means ± standard deviation of three independent experiments
(*P<0.05, **P<0.01,
***P<0.001). NS, not significant; GALNT6, polypeptide
N-acetylgalactosaminyltransferase 6; LGALS3BP, lectin
galactoside-binding soluble 3 binding protein; WT, wild-type; CM,
conditioned medium; VVA, Vicia villosa agglutinin. |
Three candidate sites (T579, S581 and S582) in the
distal region of LGALS3BP (amino acids 572-585) were examined and
T579A, S581A and S582A mutants of LGALS3BP were generated (Fig. 5C). GALNT6-dependent
O-glycosylation of LGALS3BP S582A was most significantly
attenuated compared with that of LGALS3BP WT in
GALNT6-WT-transfected HeLa cells (Fig.
5D and Fig. S5B). More
importantly, among these mutants, only LGALS3BP S582A-FLAG
exhibited a significant decrease of GALNT6-dependent
O-glycosylation in the supernatant fraction compared with
that of LGALS3BP WT in HeLa cells (Fig. 5D and Fig. S5B). Accordingly, the results
mentioned above were validated using a triple Ala substitution
construct (T556A/T571A/T582A), and it was observed that
T556A/T571A/T582A resulted in the most significant decrease in
GALNT6-dependent O-glycosylation and secretion compared with
that of LGALS3BP WT in HeLa cells (Fig. 5E and Fig. S5C). These findings suggest the
possibility that T556, T571 and S582 are crucial for
GALNT6-mediated O-glycosylation and secretion of LGALS3BP,
although further analysis of the involvement of GALNT6-mediated
O-glycosylation in the secretion of LGALS3BP in breast
cancer cells is required.
Subsequently, to validate the
O-glycosylation of LGALS3BP by GALNT6, an in vitro
GalNAc transferase assay was performed. GALNT6
O-glycosylated LGALS3BP WT peptide (568-585 amino acids) in
a dose-dependent manner (Fig. 5F).
However, GALNT6-dependent O-glycosylation of LGALS3BP mutant
peptides (T571A and S582A) was significantly attenuated. Notably,
GALNT6-dependent O-glycosylation of the LGALS3BP
double-mutant peptide (T571A/S582A) was almost abolished. Taken
together, these findings strongly suggest that GALNT6-dependent
O-glycosylation of LGALS3BP at T556, T571 and S582 is
indispensable for the extracellular release of LGALS3BP in cancer
cells.
Autocrine growth-promoting effect of
GALNT6-O- glycosylated LGALS3BP
Finally, to investigate the significance of the
secretion of O-glycosylated LGALS3BP, the growth-promoting
effect of CM from LGALS3BP (T556A/T571A/S582A)- and GALNT6 (WT or
H271D)-overexpressing cells was evaluated. CM was transferred to
HCC1806 cells and then a cell proliferation assay was performed.
The promoting effect of CM derived from LGAL3SBP
T556A/T571A/S582A/GALNT6 WT+ HeLa cells was
significantly attenuated compared with that of CM from LGAL3SBP
WT/GALNT6 WT+ HeLa cells (Fig. 5G). These findings suggest that
O-glycosylation at T556, T571 and S582 of LGALS3BP is
crucial for the release of LGALS3BP and its autocrine
growth-promoting effects in breast cancer cells. Taken together,
these findings demonstrated that the GALNT6-LGALS3BP axis regulates
mammary carcinogenesis through GALNT6-dependent LGALS3BP
O-glycosylation and release.
Discussion
In the present study, LGALS3BP was identified as a
novel substrate of GALNT6 in breast cancer cells. Although
accumulating studies have revealed the significance of each protein
in breast cancer (10,17,19,21-23,24-27),
their interaction and regulation have yet to be elucidated. To the
best of our knowledge, the present study is the first to
demonstrate that overexpressed GALNT6 colocalizes with and
O-glycosylates LGALS3BP in the Golgi apparatus, thereby
enhancing the extracellular release of O-glycosylated
LGALS3BP from breast cancer cells, which then acts in an autocrine
manner to promote breast cancer cell growth. Previous studies have
demonstrated that LGALS3BP is overexpressed in and secreted by
several tumor tissues, including breast cancer tissues (23-32).
Several functional and structural studies indicate that the
LGALS3BP protein is potentially asparagine (N-) linked
glycosylated, but not O-linked glycosylated (36,37).
However, the recombinant LGALS3BP protein used in these studies was
generated from embryonic kidney cells that do not express GALNT6.
Therefore, the present findings were the first to demonstrate that
LGALS3BP is O-linked glycosylated by GALNT6, which is
overexpressed in breast cancer cells. More importantly, through
biochemical analysis using deletion mutant and alanine mutant
constructs, three GALNT6-mediated O-glycosylation sites
(T556, T571 and S582) were identified on the LGALS3BP protein, and
they are indispensable for its secretion and growth-promoting
autocrine activity. When GALNT6 expression was knocked down in
ZR-75-1, BT-20 and SK-BR-3 breast cancer cells, the
O-glycosylation and flow of endogenous LGALS3BP were
noticeably decreased, suggesting that GALNT6 is crucial for the
O-glycosylation and secretion of endogenous LGALS3BP in
breast cancer cells. Furthermore, it was demonstrated that
treatment with CM from LGAL3SBP+GALNT6+ HeLa
cells significantly promoted HCC1806 cell proliferation (Fig. 3C). On the other hand, the effect of
LGALS3BP in CM from 293T cells on cell proliferation was found to
be mildly suppressive for HCC1806 cells (Fig. 3D). As the endogenous LGAL3BP is not
GALNT6-mediated O-glycosylated in 293T cells (Fig. 4B), these findings suggest the
possibility that the dominant-negative effect of
non-O-glycosylated LGALS3BP derived from 293T cells may
result from interference with its cell proliferative function in
HCC1806 cells.
Furthermore, the ~75-kDa band of
O-glycosylated LGALS3BP by endogenous GALNT6 (Fig. 1A) was detected by VVA lectin blot
analysis, even when LGALS3BP-FLAG was overexpressed in any cells.
By contrast, the higher (~90 kDa) band of O-glycosylated
LGALS3BP was observed when GALNT6-FLAG or -HA was exogenously
overexpressed (Figs. 1B and
2A). As GALNT6 catalyzes the
transfer of GalNAc to serine or threonine residues during the first
step of O-linked protein glycosylation, the aberrant
structure of glycan chains that are covalently attached to LGALS3BP
may occur due to exogenous GALNT6 overexpression, thereby not
extending glycan chain in breast cancer cells, although further
analyses are require to clarify this issue.
Previous reports demonstrated that GALNT6
O-glycosylates several cancer-related proteins as
substrates, including MUC1, fibronectin, MUC4, GRP78 and ER, in
order to stabilize these proteins in cancer cells (10,17-19).
Notably, GALNT6 is also likely to be required for the stabilization
of the endogenous LGALS3BP protein via O-glycosylation, as
knockdown of GALNT6 reduced intracellular LGALS3BP
expression at the protein level (Fig.
2B) but not at the transcriptional level in breast cancer cells
(Fig. S3B). However, no reduction
was observed in the total amount of LGALS3BP protein in either
LGALS3BP (T556A/T571A/S582A)- or GALNT6 (WT)-overexpressing cells,
although we observed O-glycosylation of the LGALS3BP protein
in both LGALS3BP (T556A/T571A/S582A)- and GALNT6
(WT)-overexpressing cells (Fig.
S5C). Taken together, these findings suggest the possibility
that another region of LGALS3BP may contain at least one other
GALNT6-dependent O-glycosylation site for LGALS3BP
stability, whereas GALNT6-dependent O-glycosylation at
T556/T571/S582 is essential for the auto-crine growth-promoting
activity of LGALS3BP. Further analyses are required to identify the
O-glycosylation site(s) required for LGALS3BP stability in
breast cancer cells.
Finally, the importance of LGALS3BP
O-glycosylation at T556/T571/S582 to its secretion from
cancer cells was considered, as a decrease in O-glycosylated
LGALS3BP was observed in the supernatant from both LGALS3BP
(T556A/T571A/S582A)- and GALNT6 (WT)-overexpressing cells. LGALS3BP
released via GALNT6 promoted the proliferation of breast cancer
cells (Fig. 5G). It is predicted
that soluble LGALS3BP binds to adhesion molecules in the
extracellular matrix of tumor cells and regulates adhesion
molecule-dependent signaling pathways for oncogenic events.
Previous studies demonstrated that LGALS3BP oligomerizes and
interacts with galectin-3, the integrin β1 subunit, collagens IV, V
and VI, fibronectin and nidogen in the extracellular matrix
(38,39). In addition, LGALS3BP regulates
integrin-mediated cellular adhesion and promotes cell viability,
motility and migration during oncogenic cellular events through the
Akt, ERK1/2 and JNK1/2 signaling pathways (40). LGALS3BP O-glycosylated by
GALNT6 may interact with these adhesion molecules and enhance
downstream signaling during oncogenic events, although further
analyses are required to fully elucidate the connection between
GALNT6 and LGALS3BP in breast cancer cells. Furthermore, a recent
study demonstrated that LGALS3BP acts as a centriole- and basal
body-associated protein, and LGALS3BP leads to centrosome
hypertrophy in cancer cells (41).
Accordingly, it may be hypothesized that the intracellular
GALNT6-LGALS3BP axis plays a crucial role in regulating the
centrosome in breast cancer cells.
Clinically, the diagnostic markers available for
breast cancer are not sufficiently sensitive or specific to detect
early-stage tumors. Previous studies demonstrated that high serum
or tumor tissue levels of LGALS3BP were associated with shorter
survival in patients with several types of tumors, including breast
cancer (26-32). Moreover, a recent study
demonstrated that anti-LGALS3BP antibody blocks LGALS3BP-integrin
binding and LGALS3BP-integrin-mediated cellular adhesion, viability
and motility (40). These reports
and the results of the present study collectively demonstrate that
LGALS3BP is a potential prognostic biomarker and therapeutic target
for breast cancer. To the best of our knowledge, the present study
was the first to demonstrate that a cancer-specific
O-glycosidase, GALNT6, regulates LGALS3BP secretion to
induce its growth-promoting activity via O-glycosylation.
More importantly, GALNT6 glycosylates a number of cancer-related
proteins, including LGALS3BP, regulating the proliferation and
metastatic ability of cancer cells, suggesting that a
GALNT6-specific inhibitor may be a more attractive anticancer drug
for treating breast tumors, compared with LGALS3BP-targeted
drugs.
In conclusion, the GALNT6-LGALS3BP axis appears to
be crucial for mammary carcinogenesis and may hold promise as a
therapeutic target and biomarker for mammary tumors.
Supplementary Data
Funding
The present study was supported by a grant/research
support from the Tokushima Breast Care Clinic and Sagawa Cancer
Research Promotion Public Interest Foundation. The study was also
supported by the Project for Development of Innovative Research on
Cancer Therapeutics (P-DIRECT) (11104836) from the Japan Agency for
Medical Research and Development (AMED) and Grants-in-Aid for
Scientific Research on Innovative Areas (JP17H06419) and for Young
Scientists (B) (JSPS KAKENHI JP16K19052) from the MEXT of Japan.
The present study was also supported by the Research Clusters
program of Tokushima University.
Availability of data and materials
The data for GALNT6 mRNA expression (RNA Seq
V2 RSEM) in primary invasive breast carcinoma samples were
downloaded from TCGA network (http://cancergenome.nih. gov/) (42,43).
Kaplan-Meier analyses for breast cancer patients were downloaded
from the Kaplan-Meier plotter (KM plotter) database (http://kmplot.com/analysis/) (44). Breast cancer patients in the KM
plotter database were stratified into 'low’ and 'high’
GALNT6 mRNA expression groups (Affymetrix ID: 219956_at)
based on an autoselected best cutoff value. The datasets used
and/or analyzed during the current study are available from the
corresponding author on reasonable request.
Authors’ contributions
RK performed the GALNT6-LGALS3BP functional
analysis experiments, interpreted all data and prepared the draft
of the manuscript. TY performed the glycosylation assay,
interpreted all data, and the preparation of the draft of the
manuscript. YMa performed analyses for TCGA datasets, interpreted
all data, and the preparation of the draft of the manuscript. TM
performed the screening of GALNT6 binding protein and validation of
the GALNT6-LGALS3BP interaction. MO performed the 2DICAL analysis.
YMi and MS provided the interpretation of the clinical association
data. JHP and YN discussed the interpretation of identification of
GALNT6 binding proteins. TK was involved in the conception and
design of all studies, the interpretation of the data, and the
preparation of the draft and final version of the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments were conducted according to
protocols reviewed and approved by the Committee for Safe Handling
of Living Modified Organisms at Tokushima University (Permission
no. 30-67).
Patient consent for publication
Not applicable.
Competing interests
Jae-Hyun Park is an employee of Cancer Precision
Medicine, Inc. Yusuke Nakamura is a stockholder and scientific
advisor of OncoTherapy Science, Inc. Toyomasa Katagiri is an
external board member and stockholder of OncoTherapy Science, Inc.
The remaining authors declare no competing financial interests.
Acknowledgments
The authors would like to thank Ms. Hinako Koseki
and Ms. Hitomi Kawakami for providing excellent technical
support.
References
|
1
|
Tarp MA and Clausen H: Mucin-type
O-glycosylation and its potential use in drug and vaccine
development. Biochim Biophys Acta. 1780:546–563. 2008. View Article : Google Scholar
|
|
2
|
Gill DJ, Clausen H and Bard F: Location,
location, location: New insights into O-GalNAc protein
glycosylation. Trends Cell Biol. 21:149–158. 2011. View Article : Google Scholar
|
|
3
|
Schjoldager KT and Clausen H:
Site-specific protein O-glycosylation modulates proprotein
processing-deciphering specific functions of the large polypeptide
GalNAc-transferase gene family. Biochim Biophys Acta.
1820:2079–2094. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pinho SS and Reis CA: Glycosylation in
cancer: Mechanisms and clinical implications. Nat Rev Cancer.
15:540–555. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hakomori S: Glycosylation defining cancer
malignancy: New wine in an old bottle. Proc Natl Acad Sci USA.
99:10231–10233. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hussain MR, Hoessli DC and Fang M:
N-acetylgalact osaminyl-transferases in cancer. Oncotarget.
7:54067–54081. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Berois N, Mazal D, Ubillos L, Trajtenberg
F, Nicolas A, Sastre-Garau X, Magdelenat H and Osinaga E:
UDP-N-acetyl-D-galactosamine: Polypeptide
N-acetylgalactosaminyltransferase-6 as a new immunohistochemical
breast cancer marker. J Histochem Cytochem. 54:317–328. 2006.
View Article : Google Scholar
|
|
8
|
Freire T, Berois N, Sóñora C, Varangot M,
Barrios E and Osinaga E: UDP-N-acetyl-D-galactosamine:polypeptide
N-acetylgalactosaminyltransferase 6 (ppGalNAc-T6) mRNA as a
potential new marker for detection of bone marrow-disseminated
breast cancer cells. Int J Cancer. 119:1383–1388. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Patani N, Jiang W and Mokbel K: Prognostic
utility of glycosyltransferase expression in breast cancer. Cancer
Genomics Proteomics. 5:333–340. 2008.
|
|
10
|
Park JH, Nishidate T, Kijima K, Ohashi T,
Takegawa K, Fujikane T, Hirata K, Nakamura Y and Katagiri T:
Critical roles of mucin 1 glycosylation by transactivated
polypeptide N-acetylg alactosaminyltransferase 6 in mammary
carcinogenesis. Cancer Res. 70:2759–2769. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liesche F, Kölbl AC, Ilmer M, Hutter S,
Jeschke U and Andergassen U: Role of
N-acetylgalactosaminyltransferase 6 in early tumorigenesis and
formation of metastasis. Mol Med Rep. 13:4309–4314. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin TC, Chen ST, Huang MC, Huang J, Hsu
CL, Juan HF, Lin HH and Chen CH: GALNT6 expression enhances
aggressive phenotypes of ovarian cancer cells by regulating EGFR
activity. Oncotarget. 8:42588–42601. 2017.PubMed/NCBI
|
|
13
|
Sheta R, Bachvarova M, Plante M, Gregoire
J, Renaud MC, Sebastianelli A, Popa I and Bachvarov D: Altered
expression of different GalNAc-transferases is associated with
disease progression and poor prognosis in women with high-grade
serous ovarian cancer. Int J Oncol. 51:1887–1897. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kitada S, Yamada S, Kuma A, Ouchi S,
Tasaki T, Nabeshima A, Noguchi H, Wang KY, Shimajiri S, Nakano R,
et al: Polypeptide N-acetylgalactosaminyl transferase 3
independently predicts high-grade tumours and poor prognosis in
patients with renal cell carcinomas. Br J Cancer. 109:472–481.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Z, Yamada S, Wu Y, Wang KY, Liu YP,
Uramoto H, Kohno K and Sasaguri Y: Polypeptide
N-acetylgalactosaminyltransferase-6 expression independently
predicts poor overall survival in patients with lung adenocarcinoma
after curative resection. Oncotarget. 7:54463–54473.
2016.PubMed/NCBI
|
|
16
|
Li Z, Yamada S, Inenaga S, Imamura T, Wu
Y, Wang KY, Shimajiri S, Nakano R, Izumi H, Kohno K and Sasaguri Y:
Polypeptide N-acetylgalactosaminyltransferase 6 expression in
pancreatic cancer is an independent prognostic factor indicating
better overall survival. Br J Cancer. 104:1882–1889. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Park JH, Katagiri T, Chung S, Kijima K and
Nakamura Y: Polypeptide N-acetylgalactosaminyltransferase 6
disrupts mammary acinar morphogenesis through O-glycosylation of
fibronectin. Neoplasia. 13:320–326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tarhan YE, Kato T, Jang M, Haga Y, Ueda K,
Nakamura Y and Park JH: Morphological changes, cadherin switching,
and growth suppression in pancreatic cancer by GALNT6 knockdown.
Neoplasia. 18:265–272. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin J, Chung S, Ueda K, Matsuda K,
Nakamura Y and Park JH: GALNT6 stabilizes GRP78 protein by
O-glycosylation and enhances its activity to suppress apoptosis
under stress condition. Neoplasia. 19:43–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Deng B, Tarhan YE, Ueda K, Ren L, Katagiri
T, Park JH and Nakamura Y: Critical role of estrogen receptor alpha
O-glycosylation by N-acetylgalactosaminyltransferase 6 (GALNT6) in
its nuclear localization in breast cancer cells. Neoplasia.
20:1038–1044. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Koths K, Taylor E, Halenbeck R, Casipit C
and Wang A: Cloning and characterization of a human Mac-2-binding
protein, a new member of the superfamily defined by the macrophage
scavenger receptor cysteine-rich domain. J Biol Chem.
268:14245–14249. 1993.PubMed/NCBI
|
|
22
|
Ullrich A, Sures I, D’Egidio M, Jallal B,
Powell TJ, Herbst R, Dreps A, Azam M, Rubinstein M, Natoli C, et
al: The secreted tumor-associated antigen 90K is a potent immune
stimulator. J Biol Chem. 269:18401–18407. 1994.PubMed/NCBI
|
|
23
|
Iacobelli S, Arnò E, D’Orazio A and
Coletti G: Detection of antigens recognized by a novel monoclonal
antibody in tissue and serum from patients with breast cancer.
Cancer Res. 46:3005–3010. 1986.PubMed/NCBI
|
|
24
|
Linsley PS, Horn D, Marquardt H, Brown JP,
Hellström I, Hellström KE, Ochs V and Tolentino E: Identification
of a novel serum protein secreted by lung carcinoma cells.
Biochemistry. 25:2978–2986. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Iacobelli S, Bucci I, D’Egidio M, Giuliani
C, Natoli C, Tinari N, Rubistein M and Schlessinger J: Purification
and characterization of a 90 kDa protein released from human tumors
and tumor cell lines. FEBS Lett. 319:59–65. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Iacobelli S, Sismondi P, Giai M, D’Egidio
M, Tinari N, Amatetti C, Di Stefano P and Natoli C: Prognostic
value of a novel circulating serum 90K antigen in breast cancer. Br
J Cancer. 69:172–176. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tinari N, Lattanzio R, Querzoli P, Natoli
C, Grassadonia A, Alberti S, Hubalek M, Reimer D, Nenci I, Bruzzi
P, et al: High expression of 90K (Mac-2 BP) is associated with poor
survival in node-negative breast cancer patients not receiving
adjuvant systemic therapies. Int J Cancer. 124:333–338. 2009.
View Article : Google Scholar
|
|
28
|
Fornarini B, D’Ambrosio C, Natoli C,
Tinari N, Silingardi V and Iacobelli S: Adhesion to 90K (Mac-2 BP)
as a mechanism for lymphoma drug resistance in vivo. Blood.
96:3282–3285. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Strizzi L, Muraro R, Vianale G, Natoli C,
Talone L, Catalano A, Mutti L, Tassi G and Procopio A: Expression
of glycoprotein 90K in human malignant pleural mesothelioma:
Correlation with patient survival. J Pathol. 197:218–223. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Marchetti A, Tinari N, Buttitta F, Chella
A, Angeletti CA, Sacco R, Mucilli F, Ullrich A and Iacobelli S:
Expression of 90K (Mac-2 BP) correlates with distant metastasis and
predicts survival in stage I non-small cell lung cancer patients.
Cancer Res. 62:2535–2539. 2002.PubMed/NCBI
|
|
31
|
Grassadonia A, Tinari N, Iurisci I,
Piccolo E, Cumashi A, Innominato P, D’Egidio M, Natoli C, Piantelli
M and Iacobelli S: 90K (Mac-2 BP) and galectins in tumor
progression and metastasis. Glycoconj J. 19:551–556. 2002.
View Article : Google Scholar
|
|
32
|
Grassadonia A, Tinari N, Natoli C, Yahalom
G and Iacobelli S: Circulating autoantibodies to LGALS3BP: A novel
biomarker for cancer. Dis Markers. 35:747–752. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fukawa T, Ono M, Matsuo T, Uehara H, Miki
T, Nakamura Y, Kanayama HO and Katagiri T: DDX31 regulates the
p53-HDM2 pathway and rRNA gene transcription through its
interaction with NPM1 in renal cell carcinoma. Cancer Res.
72:5867–5877. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hang HC, Yu C, Ten Hagen KG, Tian E,
Winans KA, Tabak LA and Bertozzi CR: Small molecule inhibitors of
mucin-type O-linked glycosylation from a uridine-based library.
Chem Biol. 11:337–345. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ono M, Shitashige M, Honda K, Isobe T,
Kuwabara H, Matsuzuki H, Hirohashi S and Yamada T: Label-free
quantitative proteomics using large peptide data sets generated by
nanoflow liquid chromatography and mass spectrometry. Mol Cell
Proteomics. 5:1338–1347. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hohenester E, Sasaki T and Timpl R:
Crystal structure of a scavenger receptor cysteine-rich domain
sheds light on an ancient superfamily. Nat Struct Biol. 6:228–232.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hellstern S, Sasaki T, Fauser C, Lustig A,
Timpl R and Engel J: Functional studies on recombinant domains of
Mac-2-binding protein. J Biol Chem. 277:15690–15696. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sasaki T, Brakebusch C, Engel J and Timpl
R: Mac-2 binding protein is a cell-adhesive protein of the
extracellular matrix which self-assembles into ring-like structures
and binds beta1 integrins, collagens and fibronectin. EMBO J.
17:1606–1613. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Müller SA, Sasaki T, Bork P, Wolpensinger
B, Schulthess T, Timpl R, Engel A and Engel J: Domain organization
of Mac-2 binding protein and its oligomerization to linear and
ring-like structures. J Mol Biol. 291:801–813. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Stampolidis P, Ullrich A and Iacobelli S:
LGALS3BP, lectin galactoside-binding soluble 3 binding protein,
promotes oncogenic cellular events impeded by antibody
intervention. Oncogene. 34:39–52. 2015. View Article : Google Scholar
|
|
41
|
Fogeron ML, Müller H, Schade S, Dreher F,
Lehmann V, Kühnel A, Scholz AK, Kashofer K, Zerck A, Fauler B, et
al: LGALS3BP regulates centriole biogenesis and centrosome
hypertrophy in cancer cells. Nat Commun. 4:15312013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cancer Genome Atlas Network: Comprehensive
molecular portraits of human breast tumours. Nature. 490:61–70.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ciriello G, Gatza ML, Beck AH, Wilkerson
MD, Rhie SK, Pastore A, Zhang H, McLellan M, Yau C, Kandoth C, et
al: Comprehensive molecular portraits of invasive lobular breast
cancer. Cell. 163:506–519. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Györffy B, Lanczky A, Eklund AC, Denkert
C, Budczies J, Li Q and Szallasi Z: An online survival analysis
tool to rapidly assess the effect of 22,277 genes on breast cancer
prognosis using microarray data of 1,809 patients. Breast Cancer
Res Treat. 123:725–731. 2010. View Article : Google Scholar
|