Introduction
Intrahepatic cholangiocarcinoma (IHCC) is a rare and
highly aggressive primary epithelial cancer arising from the bile
duct within the liver (1). The
incidence and mortality rates associated with IHCC have been
continuously rising over the past decade (2). Patients are often diagnosed at an
advanced incurable stage with lymph node metastasis and
multicentric disease already present within the liver. Curative
intent surgery is available for only 30-40% of patients with IHCC
(1). Moreover, the 5-year survival
rate is 30% even for patients who have undergone surgical resection
(3) and is <5-10% for patients
with unresectable IHCC (4). The
roles of adjuvant chemo-radiotherapy and targeted therapy in IHCC
have not yet been well defined and these treatments may have only
modest therapeutic effects (3,5-7).
Therefore, further investi-gation into the mechanisms of IHCC
progression is critical in order to enable the detection of novel
diagnostic biomarkers and therapeutic targets.
Long non-coding RNAs (lncRNAs) are defined as
transcripts greater than 200 nucleotides in length that are not
translated into proteins (8). A
large number of lncRNAs have been identified in recent years.
lncRNAs have been reported to play critical roles in diverse
cellular processes, including transcriptional regulation in
cis or trans mode, the organization of nuclear
domains and the regulation of proteins or RNA molecules (9). Notably, research has suggested that
lncRNAs play pivotal roles in cancer biology (10) and can act as oncogenes or tumor
suppressors. Certain lncRNAs, such as MALAT1 (11), PANDA (12), H19 (13) and MEG3 (14), have been shown to be crucial
regulators in a wide range of cancer types. The lncRNA ZEB1
antisense 1 (ZEB1-AS1), an antisense transcript derived from the
promoter region of ZEB1, was discovered to be upregulated in
hepatocellular carcinoma by Li et al in 2015 (15). Since then, ZEB1-AS1 has been
demonstrated to be overexpressed in glioma (16), colorectal cancer (17), gastric cancer (18), prostate cancer (19) and cervical cancer (20). ZEB1-AS1 mainly functions as an
oncogene and promotes cancer progression (21). ZEB1-AS1 upregulation has been shown
to be associated with a poor prognosis in multiple types of cancer
(22). However, to the best of our
knowledge, the role of ZEB1-AS1 in IHCC has not yet been
revealed.
Epithelial-mesenchymal transition (EMT) is a pivotal
cellular process of epithelial cells acquiring mesenchymal
features. EMT contributes to the malignant progression, invasion
and metastasis of cancer cells (23). The EMT process can be influenced by
various factors (24). In total, 5
miR-200 family members (miR-200s) (miR-141, miR-200a/200b/200c and
miR-429) have been revealed as essential regulators of epithelial
characteristics in several types of cells and tissues (25,26).
However, the association between miR-200s and EMT in IHCC has not
yet been determined.
The present study aimed to determine the expression
and clinical significance of ZEB1-AS1 in IHCC. Additionally, the
present study investigated the functional roles of ZEB1-AS1 in IHCC
proliferation and metastasis in vitro and in vivo.
The mechanisms underlying the induction of ZEB1-AS1 in IHCC
progression were identified. The findings presented herein provide
novel insight into the molecular mechanisms of IHCC
progression.
Materials and methods
Tissue samples
A total of 118 IHCC tissues and 20 tumor-adjacent
tissues were collected at Shandong Provincial Qianfoshan Hospital,
the First Hospital Affiliated with Shandong First Medical
University and Qilu Hospital of Shandong University between May,
2008 and October, 2013. Tumor-adjacent tissues were obtained at a
distance of least 1 cm from the tumor tissues, and pathological
results confirmed that no cancer cells existed in the
tumor-adjacent tissues. R0 resection was performed in all recruited
patients. None of the patients had received any type of anticancer
therapy, such as chemotherapy, radiotherapy, interventional therapy
or targeted therapy. Patients diagnosed with 2 or more malignancies
or who succumbed to the disease within 1 month after surgery were
excluded. The final diagnosis of the patients was confirmed by
pathological results obtained by 2 pathologists independently. The
8th edition of the tumor-node-metastasis (TNM) classification
system of the International Union Against Cancer was utilized for
clinical staging (27).
Follow-up was performed by physical examination,
biochemical tests and ultrasonography at an outpatient clinic. The
determination of cancer progression was achieved by computed
tomography or magnetic resonance imaging. The overall survival (OS)
period was defined as the time interval between the date of surgery
and either the end of follow-up or the date of death.
Progression-free survival (PFS) was defined as the period between
the date of surgery and cancer progression. The research protocol
conformed to the principles outlined in the Declaration of
Helsinki. Each patient provided written and signed informed consent
and the protocol of the study was approved by the Ethics Committee
of Shandong Provincial Qianfoshan Hospital, the First Hospital
Affiliated with Shandong First Medical University.
Cells, cell culture and treatment
Human IHCC cell lines (HuH28, HuCCT1, RBE, CCLP-1
and HCCC-9810) and a normal human intrahepatic bile duct epithelial
cell line (HIBEC) were purchased from the Cell Bank of Chinese
Academy of Sciences, Shanghai Branch. The 293 Phoenix-Ampho
packaging cell line was purchased from the American Type Culture
Collection (ATCC). Cells were cultured in RPMI-1640 medium
(Invitrogen; Thermo Fisher Scientific, Inc.) (RBE and HCCC-9810) or
Dulbecco's modified Eagle's medium (DMEM, Invitrogen; Thermo Fisher
Scientific, Inc.) (HuH28, HuCCT1, CCLP-1, HIBEC and 293
Phoenix-Ampho packaging cells) supplemented with 10% fetal bovine
serum (FBS) (Gibco, Thermo Fisher Scientific, Inc.). The cells were
cultured in 5% CO2 and 90% humidity at 37°C.
Cell transfection
The full-length cDNA of ZEB1-AS1 and two short
hairpin RNAs (shRNAs) against human ZEB1-AS1 were synthesized by
GenePharma Co. Ltd. pBabe.puro retroviral vectors containing
ZEB1-AS1 cDNA or shRNA against human ZEB1-AS1 were constructed as
previously described (28). The
shRNA sequences were as follows: shZEB1-AS1-1, 5′-AAG UUC AAU CUC
AUU GAA GUC-3′ (antisense), 5′-CUU CAA UGA GAU UGA ACU UCA-3′
(sense); shZEB1-AS1-2, 5′-AAC UUC UAG CCU CUC UUU CAA-3′
(antisense), 5′-GAA AGA GAG GCU AGA AGU UCC-3′ (sense). The RBE
cells in which ZEB1-AS1 was stably overexpressed and the HuCCT1
cells in which ZEB1-AS1 was stably knocked down were generated
using retroviral vectors. Retrovirus was produced by transiently
transfecting the 293 Phoenix-Ampho packaging cells using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Briefly, the
virus-containing the supernatant was pooled, filtered and added to
RBE or HuCCT1 cells; infected cells were then treated with
puromycin (2 µg/ml) for 2 days and surviving cells were
maintained in complete medium with puromycin (0.5
µg/ml).
miR-200a mimics, miR-200a inhibitors and negative
controls (mimics NC and inhibitor NC) were purchased from
GenePharma Co. Ltd. When cell confluence reached 50%,
oligonucleotide transfection was performed using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Cells were subjected to
subsequent experimentation at 48 h following transfection.
Western blot analysis
Total protein was extracted from the cells using
radioimmunoprecipitation assay lysis buffer (Sigma-Aldrich, Merck
KGaA) containing protease inhibitors (Roche Diagnostics). An equal
amount of total protein (20 µg) was loaded per lane and
samples were separated by 10% SDS-PAGE and then transferred to a
polyvinylidene difluoride membrane (Roche Diagnostics). After being
blocked with 5% skimmed milk for 1 h at room temperature, the
membranes were probed with primary antibodies against ZEB1
(1:3,000, cat. no. ab180905, Abcam), E-cadherin (1:1,000, cat. no.
24E10, Cell Signaling Technology, Inc.), vimentin (1:1,000, cat.
no. 3932, Cell Signaling Technology, Inc.), fibronectin (1:1,000,
cat. no. ab2413, Abcam), α-catenin (1:1,000, cat. no. 2131, Cell
Signaling Technology, Inc.), N-cadherin (1:1,000, cat. no. 13116,
Cell Signaling Technology, Inc.) or β-actin (1:2,000, cat. no.
8457, Cell Signaling Technology, Inc.) overnight at 4°C.
Subsequently, the membranes were incubated with anti-mouse
(1:3,000, cat. no. 7076, Cell Signaling Technology, Inc.) or
anti-rabbit (1:3,000, cat. no. 7074, Cell Signaling Technology,
Inc.) horseradish peroxidase-conjugated secondary antibodies at
37°C for 1 h. Finally, the immunoreactive bands were visualized
using the ECL Western blot substrate (Promega Corpo.) and a
FluorChem E system (Protein Simple).
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol reagent (Thermo Fisher Scientific, Inc.) was
used to extract total RNA from the cells and tissues according to
the manufacturer's protocol. RNA purity was evaluated based on the
A260/A280 ratio. RNA was reverse transcribed into cDNA using a
TaqMan® MicroRNA reverse transcription kit (Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. The relative expression levels of the target genes
were determined by qPCR using SYBR Premix Ex Taq (Takara
Biotechnology Co., Ltd.). The thermocycling conditions were as
follows: Initial denaturation at 95°C for 30 sec; 40 cycles of 5
sec at 95°C; 1 min at 60°C and 72°C for 15 sec; with a final
extension cycle at 72°C for 5 min. The relative levels were
calculated using the 2−ΔΔCq method (29). The endogenous control gene was
glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The primer
sequences used are listed in Table
SI.
Immunohistochemistry (IHC)
IHC was conducted as described in a previous study
(27). Briefly, the sections were
fixed with 10% formalin for 24 h at room temperature. The
paraffin-embedded sections (5 µm in thickness) were
incubated with primary antibodies against ZEB1 (1:150, cat. no.
ab180905, Abcam) or E-cadherin (1:400, cat. no. 24E10, Cell
Signaling Technology, Inc.) overnight at 4°C. The samples were then
treated with a biotinylated secondary antibody (cat. no. SAP-9100,
ZSGB-BIO) for 40 min at 37°C according to the manufacturer's
protocol. The reactions were developed using a
SignalStain® DAB substrate kit (cat. no. 8059, Cell
Signaling Technology, Inc.). Immunostained sections were lightly
counterstained with hematoxylin for 2 min at room temperature,
dehydrated in ethanol and cleared in xylene. The scoring of
immunostaining was conducted based on the staining intensity and
the percentage of positively stained area. The percentage of
positively stained area was scored as follows: 0, no IHC signal at
all; 1, <10% of cells stained; 2, 10-50% of cells stained; and
3, >50% of cells stained. IHC intensity was scored as follows:
0, no IHC signal; 1, weak IHC signal; 2, moderate IHC signal; and
3, strong IHC signal. Five randomly selected high-power fields
(×400 magnification) were photographed for each IHC slide using a
light microscope (Olympus Corp.). The overall quantification of the
IHC score was obtained by multiplying the average intensity and
percentage scores of 5 different high-power fields with the maximum
possible score being 9.
Cell proliferation assays
The cell proliferative ability was evaluated using
the Cell Counting kit (CCK-8) assay and the colony formation assay.
For the CCK-8 assay, cells (3,000 per well) were seeded in 96-well
plates and incubated at different time points (0, 24, 48, 72 and 96
h)., CCK-8 reagent (10 µl) was added to each well.
Subsequently, the cells were incubated for an additional 4 h at
37°C. The absorbance was determined at 450 nm using a microplate
spectrophotometer (Thermo Fisher Scientific, Inc.).
For the colony formation assay, the cells were
seeded in 6-well plates (1,000 cells per well) and incubated at
37°C for 7 days. Neutral formalin (10%) was then used to fix the
cells and crystal violet (cat. no. C8470, Solarbio) was used for
staining (for 30 min) at room temperature. Finally, the cells were
photographed under a microscope (Olympus Corp.).
Wound healing assay
Cells were seeded in 6-well plates and cultured
until reaching 80% confluence. To create the wound, the layer of
cells was scratched with a sterile 20 µl pipette tip. The
cells were then incubated in fresh medium containing 0.5% FBS for
48 h. The scratch was imaged at 0 and 48 h under a fluorescence
microscope (Olympus Corp.).
Cell migration and invasion assays
For the Transwell migration assay, cells
(1×105) in 200 µl of serum-free medium were
seeded in the top compartment of a Boyden chamber (8-µm pore
size; Corning Inc.). The lower compartment was filled with 700
µl of medium supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) as a chemoattractant. Following 24 h of
incubation at 37°C, cells remaining on the upper side of the
membrane were removed with a cotton swab, and the cells that had
migrated to the lower side of the membrane were fixed with 70%
ethanol for 20 min and stained with 0.1% crystal violet for 20 min
at room temperature. The cells were then counted and imaged using a
light microscope (Olympus Corp.). The protocol of the invasion
assay was similar to that of the migration assay except that in the
invasion assay, the Boyden chamber was precoated with Matrigel (BD
Biosciences) before seeding the cells.
Luciferase reporter assay
Potential ZEB1-AS1-miRNA interactions were predicted
with starBase2.0 software. The QuikChange Site-directed Mutagenesis
kit (Stratagene, Agilent Technologies, Inc.) was used to generate
mutations in potential miR-200a-binding sites. The potential
binding sequence of miR-200a in the ZEB1-AS1 gene and the mutant
sequence were amplified by PCR and cloned into the pmirGLO
dual-luciferase vector (Promega Corp.); the constructs were named
ZEB1-AS1-WT and ZEB1-AS1-Mut, respectively. For co-transfection,
293 cells at a confluence of 60-80% were transfected with
ZEB1-AS1-WT, ZEB1-AS1-Mut (50 ng) and miR-200a mimics (150 ng)
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific). Luciferase assays were conducted at 48 h following
transfection using the Dual Luciferase Reporter Assay System
(Promega Corp.) and Renilla luciferase activity was
normalized to Firefly luciferase activity.
In vivo assay
Male nude mice (BALB/c nu/nu; aged 5-6 weeks;
weight, 18-22 g; 6 per group) were purchased from Shanghai
Experimental Animal Center (Shanghai, China) and housed under a
12-h light/12-h dark cycle and sterile conditions (temperature,
26-28°C; humidity, 40-60%) with ad libitum access to water
and food. For the tumor proliferation assay, xenograft tumors were
generated via the subcutaneous injection of 3.0×106
cells into the hind limbs of the nude mice. Tumor growth was
determined with a caliper every 7 days. After 42 days, the mice
were sacrificed and images of the tumors were captured. Tumor
volume (V) was calculated as follows: V=largest diameter x
(smallest diameter)2 ×0.5. For the tumor metastasis
assay, suspensions of the cells (3.0×106) in
phosphate-buffered saline were injected into the tail veins of host
mice. After 6 weeks, the animals were sacrificed, the lungs and
livers were dissected out, and metastasis was evaluated. The animal
experiment was approved by the Ethics Committee of Shandong
Provincial Qianfoshan Hospital.
Statistical analysis
Summarized data are presented as the means ±
standard error of the mean (SEM). Differences between groups were
evaluated using the χ2 test, Student's t-test, or
one-way analysis of variance with the Least-Significant Difference
correction. Linear regressions were evaluated using Spearman rank
correlation analysis. Survival curves were generated using the
Kaplan-Meier method and differences between these curves were
evaluated with the log-rank test. Univariate and multivariate Cox
proportional hazard regression models were conducted to identify
independent prognostic factors. In all cases, P<0.05 was
considered to indicate a statistically significant difference. All
statistical analyses were performed using GraphPad Prism 5.02
(GraphPad Software, Inc.) or SPSS 16.0 software (SPSS, Inc.).
Results
Upregulation of ZEB1-AS1 is associated
with cancer progression and predicts a poor prognosis of patients
with IHCC
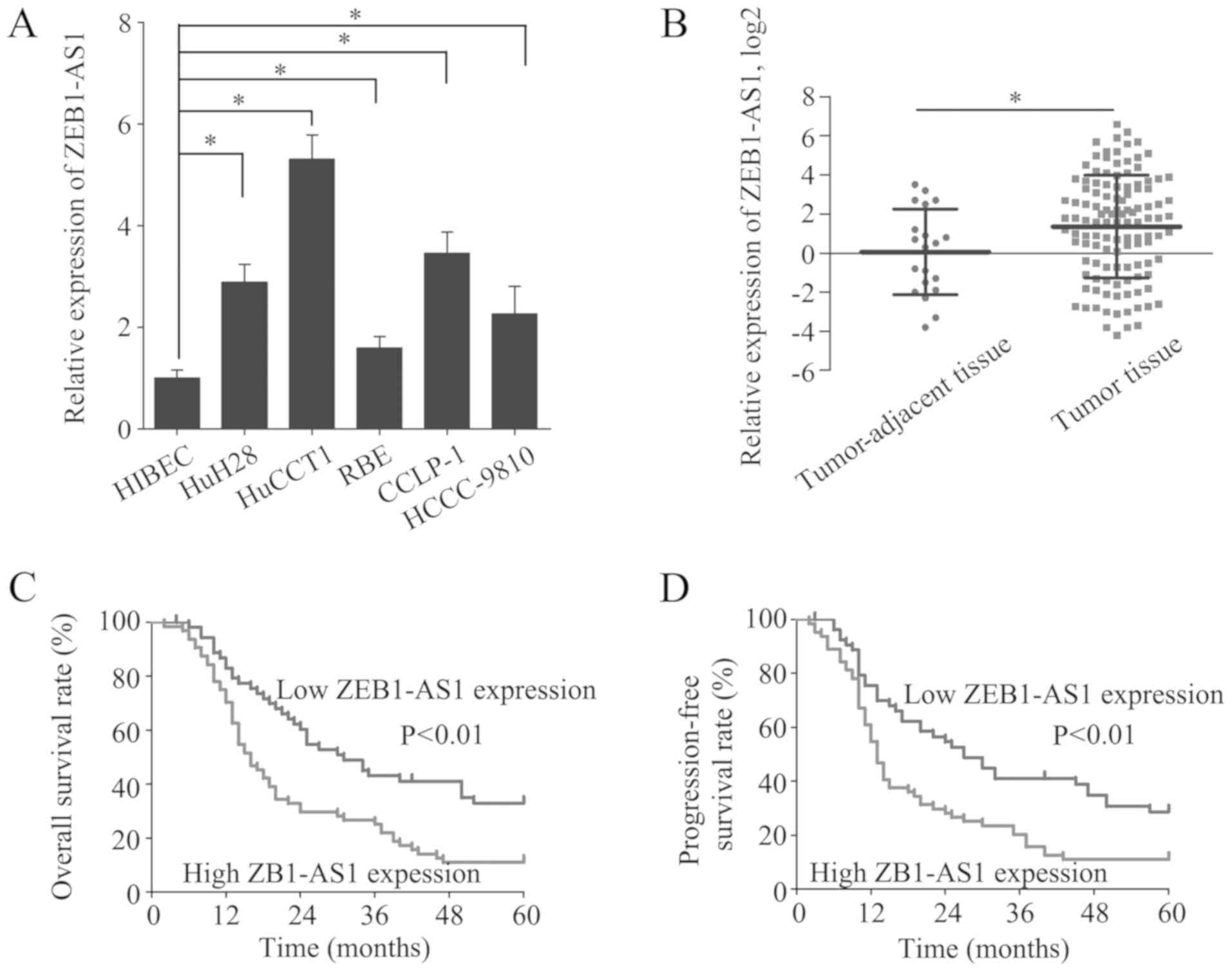
The expression of ZEB1-AS1 was measured in the
HIBECs and in 5 IHCC cell lines by RT-qPCR. The level of ZEB1-AS1
was higher in the 5 IHCC cell lines than in the HIBECs (Fig. 1A). Moreover, ZEB1-AS1 was
overexpressed in the IHCC tissues (n=118) relative to the
tumor-adjacent tissues (n=20) (Fig.
1B). To investigate the role of ZEB1-AS1 in IHCC, the patients
from whom the IHCC samples were collected were divided into a high
ZEB1-AS1 expression group (n=64) and a low ZEB1-AS1 expression
group (n=54), with the mean ZEB1-AS1 expression level serving as
the cut-off value. Of note, a high ZEB1-AS1 expression was
identified to be associated with microvascular invasion (P=0.020),
lymphatic metastasis (P=0.036) and an advanced TNM stage (P=0.037)
(Table I). These results suggest
that the overexpression of ZEB1-AS1 indicates the clinical
progression of IHCC.
 | Table IAssociation between the expression
level of ZEB1-AS1 and clinicopathological features of patients with
IHCC. |
Table I
Association between the expression
level of ZEB1-AS1 and clinicopathological features of patients with
IHCC.
| Parameters | No. of patients
(n=118) | ZEB1-AS1 expression
| P-value |
|---|
| High (n=64) | Low (n=54) |
|---|
| Age | | | | 0.611 |
| <55 years | 56 | 29 | 27 | |
| ≥55 years | 62 | 35 | 27 | |
| Sex | | | | 0.112 |
| Male | 64 | 39 | 25 | |
| Female | 54 | 25 | 29 | |
| CA19-9 | | | | 0.479 |
| <37 U/ml | 57 | 29 | 28 | |
| ≥37 U/ml | 61 | 35 | 26 | |
| No. of tumors | | | | 0.239 |
| Single | 95 | 49 | 46 | |
| Multiple | 23 | 15 | 8 | |
| Microvascular
invasion | | | | 0.020 |
| No | 65 | 29 | 36 | |
| Yes | 53 | 35 | 18 | |
| Encapsulation | | | | 0.612 |
| Absent | 43 | 22 | 21 | |
| Present | 75 | 42 | 33 | |
| Differentiation
grade | | | | 0.135 |
| Well | 46 | 21 | 25 | |
| Moderate/poor | 72 | 43 | 29 | |
| Tumor size | | | | 0.201 |
| <5 cm | 58 | 28 | 30 | |
| ≥5 cm | 60 | 36 | 24 | |
| Lymphatic
metastasis | | | | 0.036 |
| Yes | 33 | 23 | 10 | |
| No | 85 | 41 | 44 | |
| TNM stage | | | | 0.037 |
| I/II | 62 | 28 | 34 | |
| III/V | 56 | 36 | 20 | |
To further elucidate the clinical significance of
ZEB1-AS1 in IHCC, the prognosis of patients in the high and low
ZEB1-AS1 expression groups was compared. Patients with a high
ZEB1-AS1 expression had substantially lower OS and PFS rates than
patients with a low ZEB1-AS1 expression (Fig. 1C and D). Additionally, the risk
factors for poor OS and PFS were detected by univariate analysis
and the identified 6 risk factors were subjected to multivariate
analysis to discover independent risk factors for poor OS and PFS
(Table II). A high ZEB1-AS1
expression was revealed to be an independent risk factor for poor
OS (HR, 1.157; 95% CI, 1.059-1.265; P=0.001) and PFS (HR, 1.131;
95% CI, 1.038-1.232; P=0.005) (Table
III). These results indicate that high ZEB1-AS1 expression is a
potential biomarker for the prediction of a poor prognosis in
IHCC.
| Table IIUnivariate analysis of the risk
factors for poor overall survival and progression-free survival of
patients with IHCC. |
Table II
Univariate analysis of the risk
factors for poor overall survival and progression-free survival of
patients with IHCC.
| Parameters | Overall survival
| Progression-free
survival
|
|---|
| HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Age, ≥55 vs. <55
years | 1.000
(0.664-1.506) | 0.999 | 0.989
(0.660-1.482) | 0.956 |
| Sex, female vs.
male | 0.928
(0.614-1.402) | 0.722 | 0.948
(0.630-1.425) | 0.797 |
| CA19-9, ≥37 U/ml
vs. <37 U/ml | 1.069
(0.709-1.612) | 0.749 | 1.090
(0.726-1.637) | 0.676 |
| No. of tumors,
multiple vs. single | 2.091
(1.267-3.451) | 0.004 | 1.945
(1.181-3.203) | 0.009 |
| Microvascular
invasion, yes vs. no | 2.015
(1.327-3.060) | 0.001 | 1.998
(1.320-3.024) | 0.001 |
| Encapsulation,
present vs. absent | 1.194
(0.785-1.816) | 0.407 | 1.217
(0.804-1.842) | 0.353 |
| Differentiation,
poor/moderate vs. well | 1.681
(1.081-2.613) | 0.021 | 1.596
(1.035-2.462) | 0.034 |
| Tumor size, ≥5 vs.
<5 cm | 1.384
(0.918-2.087) | 0.121 | 1.439
(0.958-2.163) | 0.079 |
| Lymphatic
metastasis, yes vs. no | 3.130
(2.006-4.886) | <0.001 | 3.297
(2.114-5.142) | <0.001 |
| TNM stage, III/IV
vs. I/II | 2.662
(1.749-4.053) | <0.001 | 2.616
(1.725-3.969) | <0.001 |
| ZEB1-AS1
expression, high vs. low | 1.211
(1.114-1.316) | <0.001 | 1.185
(1.093-1.285) | <0.001 |
| Table IIIMultivariate analysis of the
independent risk factors for poor overall survival and
progression-free survivalof patients with IHCC. |
Table III
Multivariate analysis of the
independent risk factors for poor overall survival and
progression-free survivalof patients with IHCC.
| Parameters | Overall survival
| Progression-free
survival
|
|---|
| HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| No. of tumors,
multiple vs. single | 1.837
(1.090-3.095) | 0.022 | 1.556
(0.927-2.613) | 0.094 |
| Microvascular
invasion, yes vs. no | 1.095
(0.678-1.769) | 0.710 | 1.119
(0.694-1.806) | 0.644 |
| Differentiation,
poor/moderate vs. well | 0.977
(0.611-1.626) | 0.991 | 0.988
(0.614-1.588) | 0.959 |
| Lymphatic
metastasis, yes vs. no | 1.547
(0.863-2.771) | 0.143 | 1.715
(0.980-3.127) | 0.058 |
| TNM stage, III/IV
vs. I/II | 1.877
(1.099-3.205) | 0.021 | 1.777
(1.049-3.010) | 0.032 |
| ZEB1-AS1
expression, high vs. low | 1.157
(1.059-1.265) | 0.001 | 1.131
(1.038-1.232) | 0.005 |
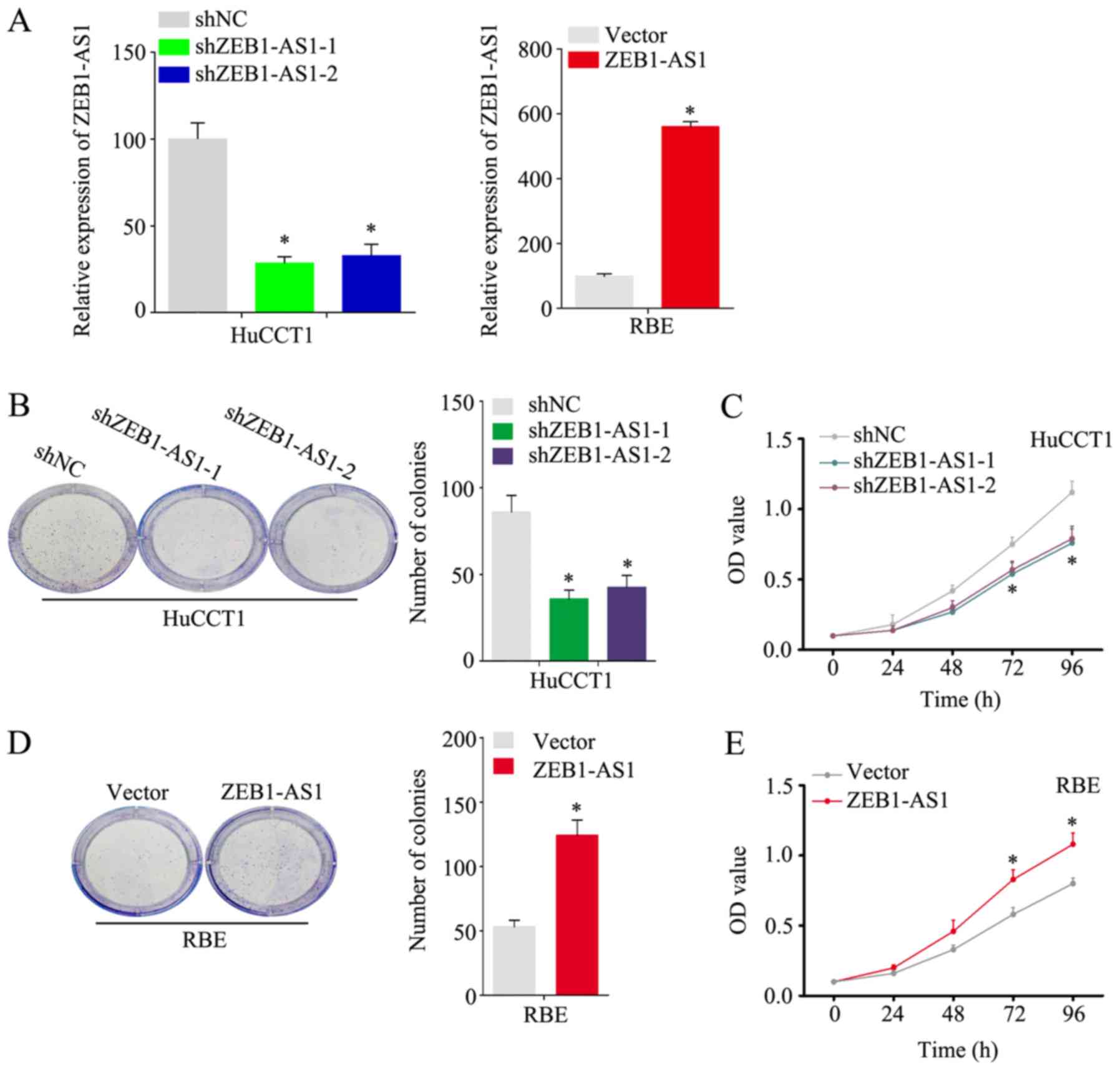
ZEB1-AS1 accelerates the proliferation,
migration and invasion of IHCC cells
As shown in Fig.
1A, among the 5 IHCC cell lines examined, the HuCCT1 line had
the highest expression level of ZEB1-AS1, and the RBE line had the
lowest. To better investigate the function of ZEB1-AS1 in IHCC, the
knockdown and stable overexpression of ZEB1-AS1 was performed in
the HuCCT1 and RBE cells, respectively. (Fig. 2A) Notably, the HuCCT1 cells with
ZEB1-AS1 deficiency exhibited a substantially reduced proliferative
ability in the colony formation and CCK-8 assays (Fig. 2B and C). Consistently, the ectopic
expression of ZEB1-AS1 significantly increased the proliferative
ability of the RBE cells (Fig. 2D and
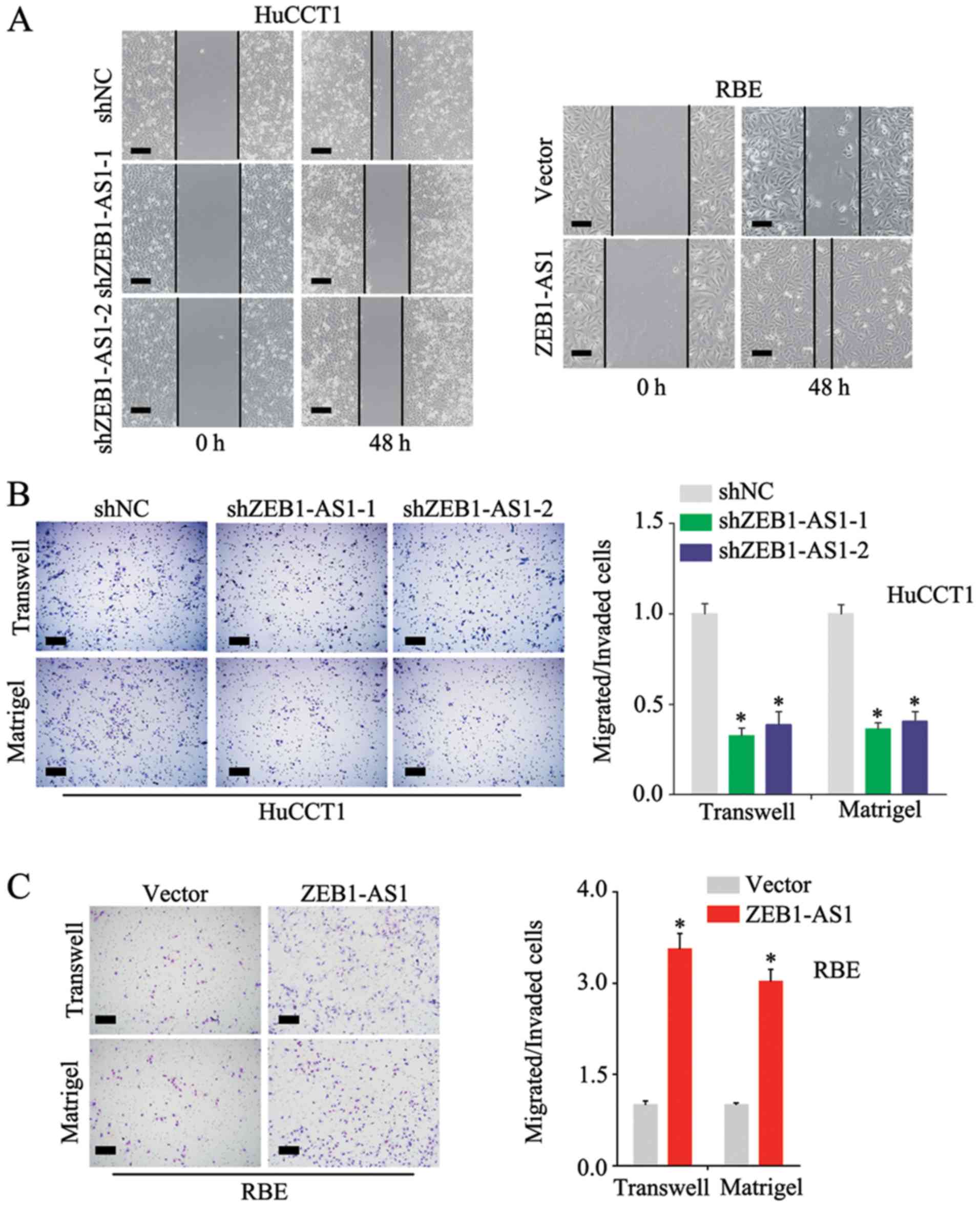
E). In the wound healing assay, the knockdown of ZEB1-AS1
inhibited the migration of the HuCCT1 cells and the overexpression
of ZEB1-AS1 promoted the migration of RBE cells (Fig. 3A); these results were confirmed by
the Transwell assay (Fig. 3B and
C, upper panels). Additional experiments demonstrated that
ZEB1-AS1 accelerated the invasion of IHCC cells (Fig. 3B and C, lower panels) These
findings demonstrate that ZEB1-AS1 promotes the proliferation,
migration and invasion of IHCC cells.
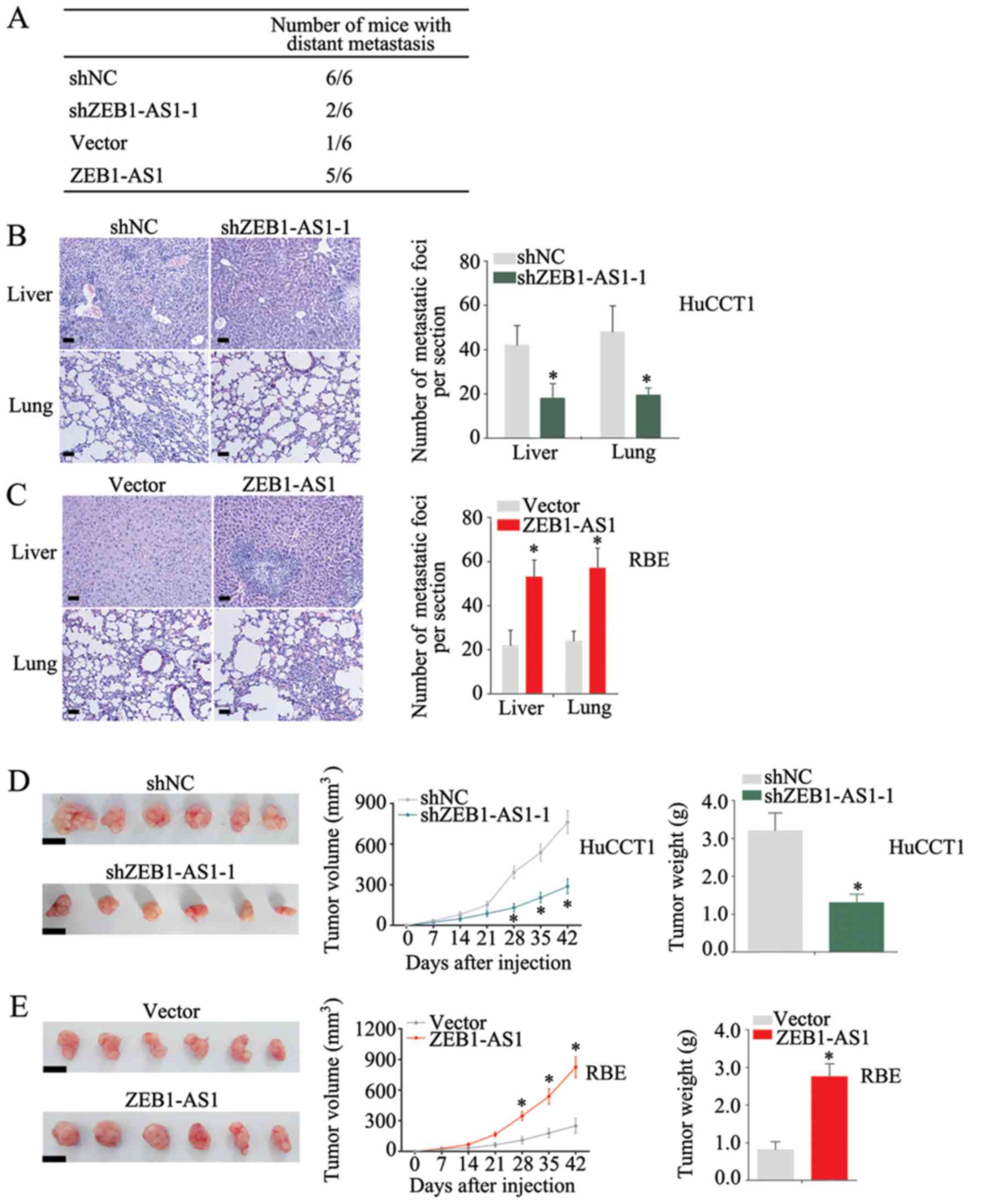
ZEB1-AS1 promotes IHCC metastasis and
growth in vivo
To investigate the function of ZEB1-AS1, in
vivo assays were performed. The knockdown of ZEB1-AS1 decreased
the number of mice with distant metastasis, whereas the
over-expression of ZEB1-AS1 increased this number (Fig. 4A). Moreover, the metastatic ability
of the HuCCT1 cells was significantly inhibited when ZEB1-AS1 was
knocked down (Fig. 4B), and
ectopic expression of ZEB1-AS1 increased the number of metastatic
foci in liver and lungs (Fig. 4C).
Additionally, a decreased tumor size and weight were observed when
ZEB1-AS1 was knocked down (Fig.
4D), whereas the overexpression of ZEB1-AS1 markedly increased
the tumorigenicity of IHCC cells (Fig.
4E). These results suggest that ZEB1-AS1 promotes the
metastasis and growth of IHCC.
ZEB1-AS1 induces EMT in IHCC
EMT is a fundamental process of cancer metastasis
(23). In the present study, to
explore the mechanisms underlying ZEB1-AS1 promotion of metastasis
in IHCC, the role of ZEB1-AS1 in EMT was investigated. The
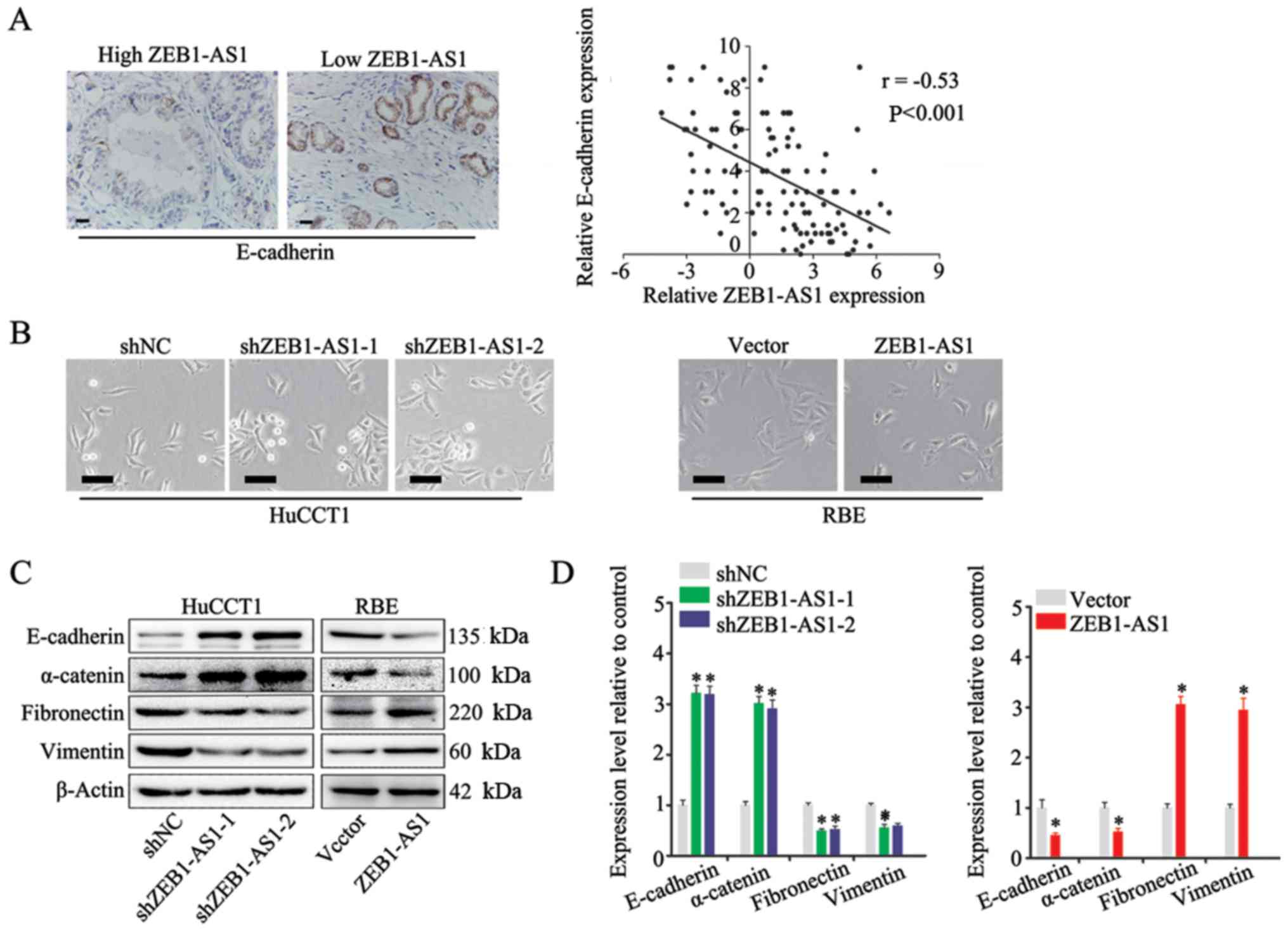
expression of E-cadherin, a marker of EMT (30), was measured by IHC in IHCC tissues,
and a negative association was found between the expression of
ZEB1-AS1 and E-cadherin (Fig. 5A,
left panel). In addition, Spearman's correlation analysis revealed
that there was a negative correlation between ZEB1-AS1 and
E-cadherin expression in IHCC (Fig.
5A, right panel). Moreover, the HuCCT1 cells in which ZEB1-AS1
was silenced displayed an epithelial morphology and formed colonies
with increased intercellular contacts (Fig. 5B, left panels). Consistently, the
overexpression of ZEB1-AS1 in the RBE cells induced a mesenchymal
phenotype and reduced intercellular contacts compared with that in
the control cells, which is a typical feature of cells undergoing
EMT (Fig. 5B, right panels)
Furthermore, the results of western blot analysis and RT-qPCR
revealed that the expression of ZEB1-AS1 was positively associated
with mesenchymal biomarkers (fibronectin, vimentin and N-cadherin)
and negatively associated with epithelial biomarkers (E-cadherin
and α-catenin) (Fig. 5C and D).
Based on these observations, it was concluded that ZEB1-AS1 induces
EMT in IHCC.
ZEB1-AS1 regulates EMT through the
miR-200a/ ZEB1 signaling pathway
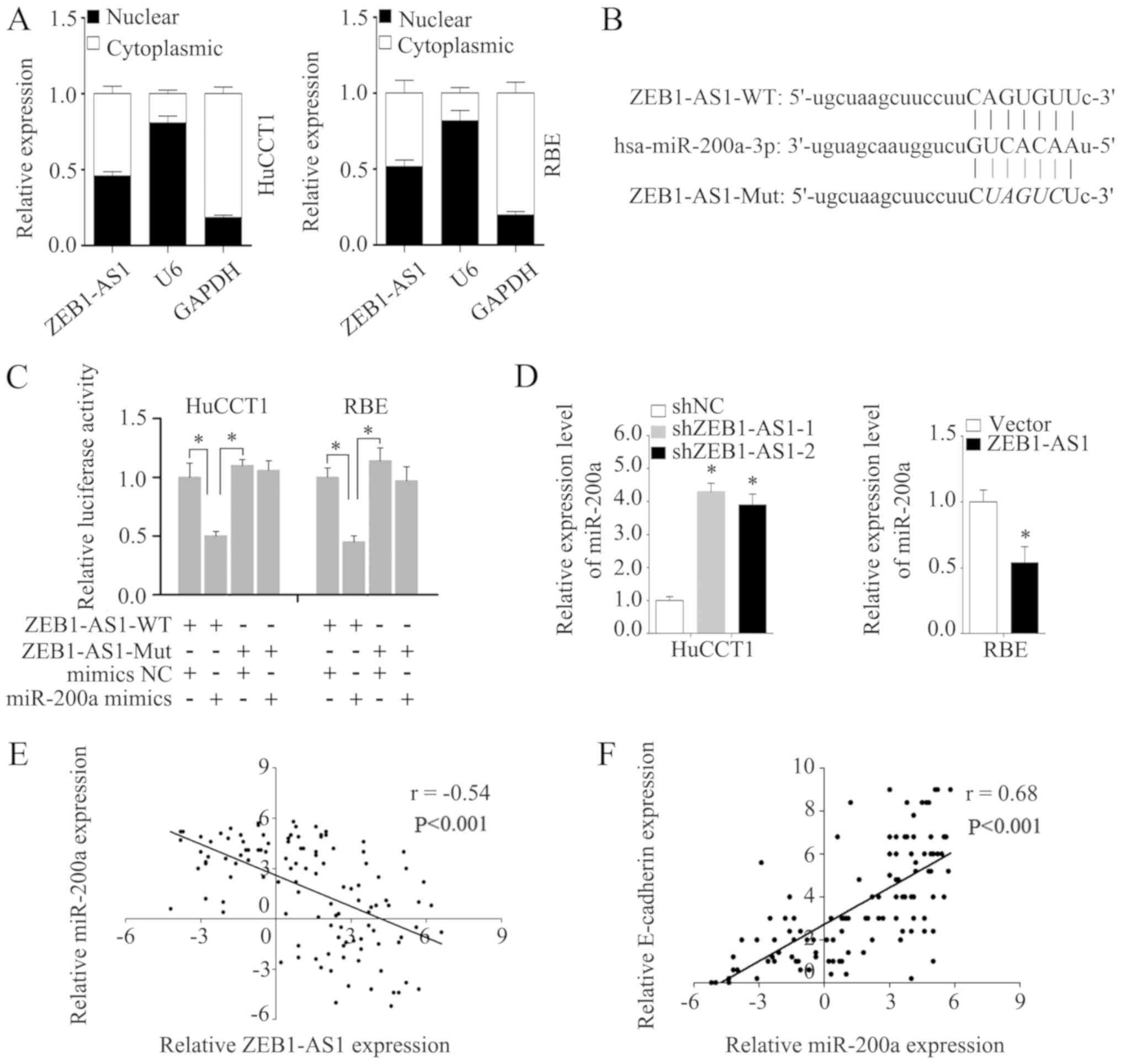
To enhance our understanding of ZEB1-AS1, the
subcellular localization of ZEB1-AS1 was detected via assays. The
analysis of the nuclear and cytoplasmic ZEB1-AS1 RNA levels by
RT-qPCR revealed that ZEB1-AS1 was located in the nucleus and
cytoplasm in HuCCT1 and RBE cells (Fig. 6A). Competing with miRNAs is a
primary mechanism of lncRNA functions (31). It has previously been demonstrated
that ZEB1-AS1 can sponge multiple miRNAs (32). The subcellular location of ZEB1-AS1
indicates that ZEB1-AS1 may function by competing with miRNAs.
Thus, in the present study, potential miRNA targets of ZEB1-AS1
were predicted using the bioinformatics tool starBase, and miR-200a
was predicted to be a potential target of ZEB1-AS1 (Fig. 6B). It was found that miR-200a
mimics markedly reduced the luciferase activity of ZEB1-AS1-WT, but
had no effect on ZEB1-AS1-Mut (Fig.
6C). Moreover, miR-200a expression was increased in the HuCCT1
cells in which ZEB1-AS1 was knocked down and decreased in the RBE
cells in which ZEB1-AS1 was overexpressed (Fig. 6D). Additionally, the level of
miR-200a negatively correlated with ZEB1-AS1 in the IHCC tissues
(Fig. 6E). Furthermore, a strong
positive correlation was found to exist between miR-200a and
E-cadherin expression in IHCC tissues (Fig. 6F), which indicates that miR-200a
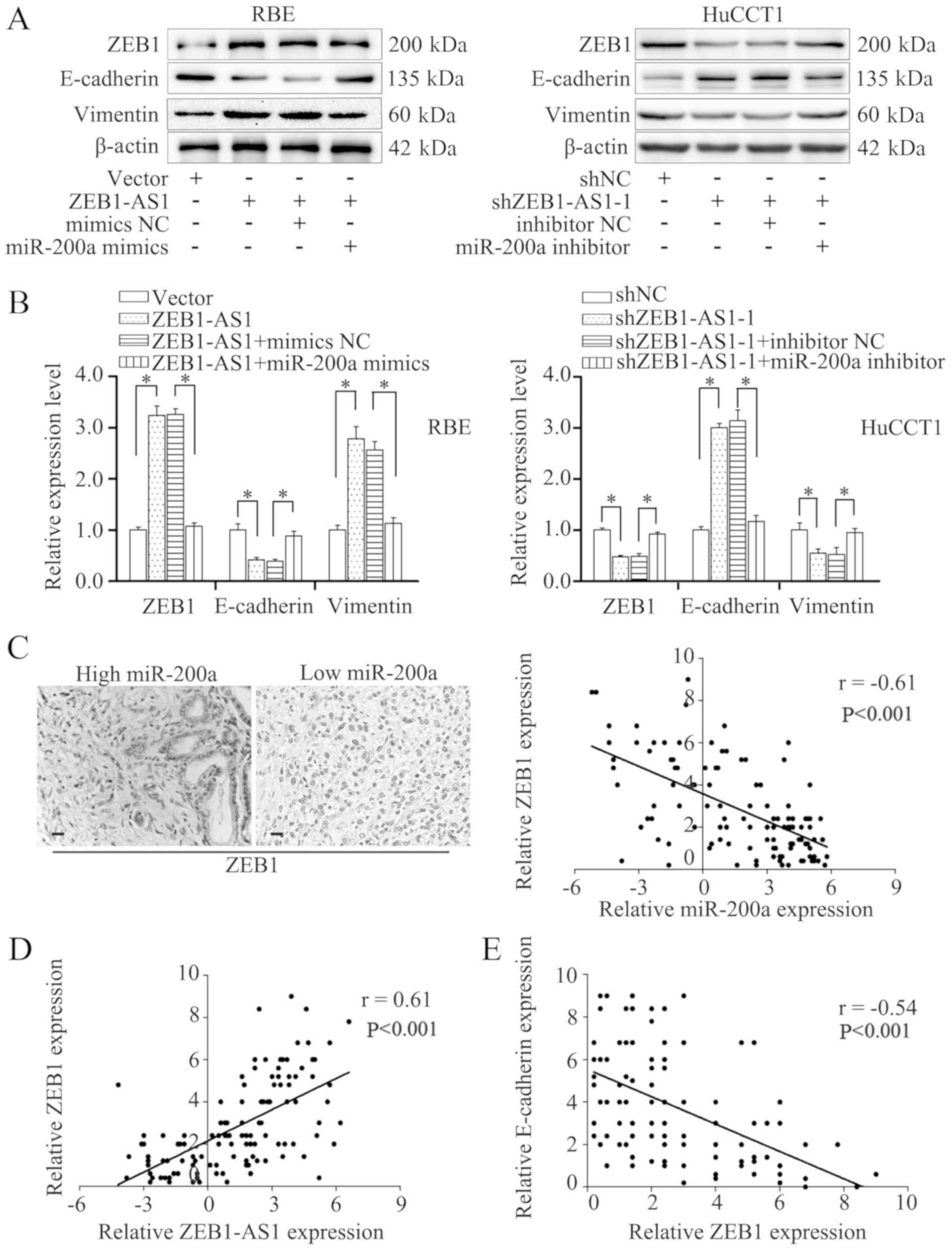
may influence EMT in IHCC. As expected, transfection with miR-200a
mimics reversed the downregulation of E-cadherin and the
upregulation of vimentin induced by the overexpression of ZEB1-AS1,
as evidenced by the results of western blot analysis and RT-qPCR
(Fig. 7A and B). Consistently,
transfection with miR-200a inhibitor blocked the increased
expression of E-cadherin and the decreased expression of vimentin
in the HuCCT1 cells in which ZEB1-AS1 was knocked down (Fig. 7A and B). These results demonstrate
that miR-200a blocks the EMT induced by ZEB1-AS1.
ZEB1, a master regulator of EMT (26), is a potential target of miR-200a
(33,34). Previous studies have reported that
ZEB1-AS1 can function through ZEB1 (32,35,36).
Thus, the present study evaluated the expression of ZEB1 by IHC;
the results revealed indicated a negative association between
miR-200a and ZEB1 (Fig. 7C, left
panels). In addition, Spearman's correlation analysis also revealed
a negative correlation between ZEB1 and miR-200a expression
(Fig. 7C, right panel).
Transfection with miR-200a mimics reversed the upregulated
expression of ZEB1 induced by ZEB1-AS1 overexpression, and
transfection with miR-200a inhibitor restored the reduced
expression of ZEB1 induced by ZEB1-AS1 silencing (Fig. 7A and B). Moreover, the expression
of ZEB1 positively correlated with that of ZEB1-AS1, but negatively
correlated with E-cadherin expression in the IHCC tissues (Fig. 7D and E). These findings demonstrate
that ZEB1-AS1 induces EMT through the miR-200a/ZEB1 signaling
pathway.
Discussion
The findings of the present study demonstrated that
ZEB1-AS1 was overexpressed in IHCC and promoted IHCC proliferation
and metastasis in vitro and in vivo. Moreover, a high
ZEB1-AS1 expression was associated with microvascular invasion,
lymphatic metastasis and an advanced TNM stage; all these factors
are pivotal in influencing the survival of patients with IHCC. The
log-rank test revealed that patients with a high ZEB1-AS1
expression had considerably lower OS and PFS rates than patients
with a low ZEB1-AS1 expression. Additionally, a high ZEB1-AS1
expression was identified as an independent risk factor for low OS
and PFS. A previous meta-analysis aimed at discovering the
prognostic value of ZEB1-AS1 in cancer, which included 11 studies
representing 891 cancer patients, indicated that a high ZEB1-AS1
expression was an unfavorable predictor of cancer prognosis in
terms of OS, disease-free survival and recurrence-free survival
(37). Taken together, these
findings indicate that ZEB1-AS1 functions as an oncogene in cancer
and may be a promising prognostic biomarker for cancer patients
including patients with IHCC.
In cholangiocarcinoma, EMT plays a prominent role in
cancer progression and may lead to highly metastatic and
desmoplastic stroma (30). EMT
biomarkers have prognostic values in cholangiocarcinoma (30). Additionally, EMT pathway-targeted
therapy is regarded as a potential IHCC treatment (38,39).
The present study revealed that the level of ZEB1-AS1 expression
was associated with morphological changes in IHCC cells. Moreover,
the level of ZEB1-AS1 was found to be positively associated with
epithelial biomarkers and negatively associated with mesenchymal
biomarkers at the protein and mRNA level, suggesting that ZEB1-AS1
may regulate EMT at the transcriptional level. Further experiments
were performed to confirm this hypothesis. As presented in the
current study, no statistically significant differences were
observed between the expression of ZEB1-AS1 and the number of
tumors and tumor size in the clinical specimens; however, ZEB-AS1
accelerated cell proliferation in colony formation and CCK-8
assays. This may be caused by the differences between the in
vitro assays and the human body.
miR-200a is a member of the miR-200 family (40). Several studies have demonstrated
that miR-200 family members are downregulated during tumor
progression and act as key inhibitors of EMT (25,26,41,42).
In addition, they can target the transcriptional factors of EMT
(ZEB1 and ZEB2) and thereby modulate the expression of E-cadherin
(33,34). In cholangiocarcinoma, miR-200b/c
has been reported to mediate the migration and invasion by directly
targeting rho-kinase 2 (43).
Moreover, DLC1 has been demonstrated to be a potential target of
miR-200a and miR-429 may target FBXW7 and CDH6 in
cholangiocarcinoma (44).
Additionally, miR-200a has been shown to suppress the invasive
ability of cholangiocarcinoma cells (45). However, the mechanisms underlying
the regulatory functions of miR-200 family members functions in
IHCC require further investigation. The present study found that
ZEB1-AS1 mediated the expression of ZEB1 by competing with
miR-200a; this finding was confirmed by luciferase reporter assay.
Moreover, a negative correlation was observed between ZEB1-AS1 and
miR-200a expression in IHCC cells and tissues. Furthermore,
miR-200a reversed the effects of an enhanced or reduced ZEB1-AS1
expression. These results confirm the role of miR-200a in mediating
the function of ZEB1-AS1.
In conclusion, the findings of the present study
demonstrate that the upregulation of ZEB1-AS1 promotes IHCC
progres-sion by accelerating proliferation and metastasis.
Moreover, high ZEB1-AS1 expression indicates poor OS and PFS rates,
and serves as an independent risk factor for poor OS and PFS.
ZEB1-AS1 acts as a molecular sponge for miR-200a and regulates the
expression of its target gene ZEB1. These findings contribute to
the better understanding of the mechanisms underlying IHCC
progression. ZEBA-AS1 may be a promising biomarker for the
prediction of prognosis and a potential target for IHCC
treatment.
Supplementary Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Sciences Foundation of China (grant nos. 81903044 and
31701013), and the Natural Sciences Foundation of Shandong Province
(grant no. ZR2017BC032).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MiJ, SN, JC and HY were involved in the
conceptualization of the study. ZC was involved in data curation.
SN and HY were responsible for funding acquisition. JC, LC, MeJ, LG
and WM were involved in the study methodology. MiJ, SN, JC and LG
were involved in project administration. ZC was responsible for
obtaining resources. MiJ, LC and WM were involved in data
validation. MeJ was invovled in the acquisition of the software
used. HY was involved in the writing of the original draft, and in
the writing, reviewing and editing of the manuscript. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
The research protocol conformed to the principles
outlined in the Declaration of Helsinki. Each patient provided
written and signed informed consent and the protocol of the study
was approved by the Ethics Committee of Shandong Provincial
Qianfoshan Hospital, the First Hospital Affiliated with Shandong
First Medical University. The animal experiment was approved by the
Ethics Committee of Shandong Provincial Qianfoshan Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sirica AE, Gores GJ, Groopman JD, Selaru
FM, Strazzabosco M, Wei Wang X and Zhu AX: Intrahepatic
cholangiocarcinoma: Continuing challenges and translational
advances. Hepatology. 69:1803–1815. 2019. View Article : Google Scholar :
|
|
2
|
Squires MH, Cloyd JM, Dillhoff M, Schmidt
C and Pawlik TM: Challenges of surgical management of intrahepatic
cholangiocarcinoma. Expert Rev Gastroenterol Hepatol. 12:671–681.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mavros MN, Economopoulos KP, Alexiou VG
and Pawlik TM: Treatment and prognosis for patients with
intrahepatic cholangiocarcinoma: Systematic review and
meta-analysis. JAMA Surg. 149:565–574. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shaib Y and El-Serag HB: The epidemiology
of cholangiocarcinoma. Semin Liver Dis. 24:115–125. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Farges O, Fuks D, Boleslawski E, Le Treut
YP, Castaing D, Laurent A, Ducerf C, Rivoire M, Bachellier P,
Chiche L, et al: Influence of surgical margins on outcome in
patients with intrahepatic cholangiocarcinoma: A multicenter study
by the AFC-IHCC-2009 study group. Ann Surg. 254:824–830. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
de Jong MC, Nathan H, Sotiropoulos GC,
Paul A, Alexandrescu S, Marques H, Pulitano C, Barroso E, Clary BM,
Aldrighetti L, et al: Intrahepatic cholangiocarcinoma: An
international multi-institutional analysis of prognostic factors
and lymph node assessment. J Clin Oncol. 29:3140–3145. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moeini A, Sia D, Bardeesy N, Mazzaferro V
and Llovet JM: Molecular pathogenesis and targeted therapies for
intrahepatic cholangiocarcinoma. Clin Cancer Res. 22:291–300. 2016.
View Article : Google Scholar
|
|
8
|
Gutschner T and Diederichs S: The
hallmarks of cancer: A long non-coding RNA point of view. RNA Biol.
9:703–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ulitsky I and Bartel DP: LincRNAs:
Genomics, evolution, and mechanisms. Cell. 154:26–46. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang Y, Cheung BB, Atmadibrata B, Marshall
GM, Dinger ME, Liu PY and Liu T: The regulatory role of long
noncoding RNAs in cancer. Cancer Lett. 391:12–19. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gutschner T, Hammerle M and Diederichs S:
MALAT1-a paradigm for long noncoding RNA function in cancer. J Mol
Med (Berl). 91:791–801. 2013. View Article : Google Scholar
|
|
12
|
Zou Y, Zhong Y, Wu J, Xiao H, Zhang X,
Liao X, Li J, Mao X, Liu Y and Zhang F: Long non-coding PANDAR as a
novel biomarker in human cancer: A systematic review. Cell Prolif.
51:e124222018. View Article : Google Scholar
|
|
13
|
Chen JS, Wang YF, Zhang XQ, Lv JM, Li Y,
Liu XX and Xu TP: H19 serves as a diagnostic biomarker and
up-regulation of H19 expression contributes to poor prognosis in
patients with gastric cancer. Neoplasma. 63:223–230. 2016.
|
|
14
|
Zhang L, Yang Z, Trottier J, Barbier O and
Wang L: Long noncoding RNA MEG3 induces cholestatic liver injury by
interaction with PTBP1 to facilitate shp mRNA decay. Hepatology.
65:604–615. 2017. View Article : Google Scholar
|
|
15
|
Li T, Xie J, Shen C, Cheng D, Shi Y, Wu Z,
Deng X, Chen H, Shen B, Peng C, et al: Upregulation of long
noncoding RNA ZEB1-AS1 promotes tumor metastasis and predicts poor
prognosis in hepatocellular carcinoma. Oncogene. 35:1575–1584.
2016. View Article : Google Scholar
|
|
16
|
Lv QL, Hu L, Chen SH, Sun B, Fu ML, Qin
CZ, Qu Q, Wang GH, He CJ and Zhou HH: A long noncoding RNA ZEB1-AS1
promotes tumorigenesis and predicts poor prognosis in glioma. Int J
Mol Sci. 17:pii: E1431. 2016. View Article : Google Scholar
|
|
17
|
Fu J and Cui Y: Long noncoding RNA
ZEB1-AS1 expression predicts progression and poor prognosis of
colorectal cancer. Int J Biol Markers. 32:e428–e433. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Y, Wen X, Wang L, Sun X, Ma H, Fu Z and
Li L: LncRNA ZEB1-AS1 predicts unfavorable prognosis in gastric
cancer. Surg Oncol. 26:527–534. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Su W, Xu M, Chen X, Chen N, Gong J, Nie L,
Li L, Li X, Zhang M and Zhou Q: Long noncoding RNA ZEB1-AS1
epigenetically regulates the expressions of ZEB1 and downstream
molecules in prostate cancer. Mol Cancer. 16:1422017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cheng R, Li N, Yang S, Liu L and Han S:
Long non-coding RNA ZEB1-AS1 promotes cell invasion and epithelial
to mesenchymal transition through inducing ZEB1 expression in
cervical cancer. Onco Targets Ther. 11:7245–7253. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ni Y, Fang J, Zhu L, Jiang H, Liu Y, Miao
R, Shao C and Shao S: The significant prognostic value of ZEB1-AS1
up-regulation in patients with cancer. J Cancer. 9:2502–2509. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu Y, Ding M, Wei S, Wu T, Xu R, Zhu X and
Liu H: The prognostic value of long noncoding RNA ZEB1-AS1 on
clinical outcomes in human cancer. J Cancer. 9:3690–3698. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pastushenko I and Blanpain C: EMT
transition states during tumor progression and metastasis. Trends
Cell Biol. 29:212–226. 2019. View Article : Google Scholar
|
|
24
|
Dongre A and Weinberg RA: New insights
into the mechanisms of epithelial-mesenchymal transition and
implications for cancer. Nat Rev Mol Cell Biol. 20:69–84. 2019.
View Article : Google Scholar
|
|
25
|
Brabletz S and Brabletz T: The ZEB/miR-200
feedback loop-a motor of cellular plasticity in development and
cancer? EMBO Rep. 11:670–677. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hill L, Browne G and Tulchinsky E:
ZEB/miR-200 feedback loop: At the crossroads of signal transduction
in cancer. Int J Cancer. 132:745–754. 2013. View Article : Google Scholar
|
|
27
|
Ueno M, Morizane C, Ikeda M, Okusaka T,
Ishii H and Furuse J: A review of changes to and clinical
implications of the eighth TNM classification of hepatobiliary and
pancreatic cancers. Jpn J Clin Oncol. 49:1073–1082. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang H, Lu X, Liu Z, Chen L, Xu Y, Wang Y,
Wei G and Chen Y: FBXW7 suppresses epithelial-mesenchymal
transition, stemness and metastatic potential of cholangiocarcinoma
cells. Oncotarget. 6:6310–6325. 2015.PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expres-sion data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
30
|
Vaquero J, Guedj N, Claperon A, Nguyen
Ho-Bouldoires TH, Paradis V and Fouassier L: Epithelial-mesenchymal
transition in cholangiocarcinoma: From clinical evidence to
regulatory networks. J Hepatol. 66:424–441. 2017. View Article : Google Scholar
|
|
31
|
Bartonicek N, Maag JL and Dinger ME: Long
noncoding RNAs in cancer: Mechanisms of action and technological
advancements. Mol Cancer. 15:432016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li J, Li Z, Leng K, Xu Y, Ji D, Huang L,
Cui Y and Jiang X: ZEB1-AS1: A crucial cancer-related long
non-coding RNA. Cell Prolif. 51:e124232018. View Article : Google Scholar
|
|
33
|
Park SM, Gaur AB, Lengyel E and Peter ME:
The miR-200 family determines the epithelial phenotype of cancer
cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes
Dev. 22:894–907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qian W, Cai X, Qian Q, Peng W, Yu J, Zhang
X, Tian L and Wang C: LncRNA ZEB1-AS1 promotes pulmonary fibrosis
through ZEB1-mediated epithelial-mesenchymal transition by
competitively binding miR-141-3p. Cell Death Dis. 10:1292019.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qu R, Chen X and Zhang C: LncRNA
ZEB1-AS1/miR-409-3p/ZEB1 feedback loop is involved in the
progression of non-small cell lung cancer. Biochem Biophys Res
Commun. 507:450–456. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen C, Feng Y and Wang X: LncRNA ZEB1-AS1
expression in cancer prognosis: Review and meta-analysis. Clin Chim
Acta. 484:265–271. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hirose A, Tajima H, Ohta T, Tsukada T,
Okamoto K, Nakanuma S, Sakai S, Kinoshita J, Makino I, Furukawa H,
et al: Low-dose paclitaxel inhibits the induction of
epidermal-mesenchymal transition in the human cholangiocarcinoma
CCKS-1 cell line. Oncol Lett. 6:915–920. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu Z, Wang J, Zheng T, Liang Y, Yin D,
Song R, Pei T, Pan S, Jiang H and Liu L: FTY720 inhibits
proliferation and epithelial-mesenchymal transition in
cholangiocarcinoma by inactivating STAT3 signaling. BMC Cancer.
14:7832014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Senfter D, Madlener S, Krupitza G and
Mader RM: The microRNA-200 family: Still much to discover. Biomol
Concepts. 7:311–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cano A and Nieto MA: Non-coding RNAs take
centre stage in epithelial-to-mesenchymal transition. Trends Cell
Biol. 18:357–359. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gregory PA, Bracken CP, Bert AG and
Goodall GJ: MicroRNAs as regulators of epithelial-mesenchymal
transition. Cell Cycle. 7:3112–3118. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Peng F, Jiang J, Yu Y, Tian R, Guo X, Li
X, Shen M, Xu M, Zhu F, Shi C, et al: Direct targeting of
SUZ12/ROCK2 by miR-200b/c inhibits cholangiocarcinoma
tumourigenesis and metastasis. Br J Cancer. 109:3092–3104. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Goeppert B, Ernst C, Baer C, Roessler S,
Renner M, Mehrabi A, Hafezi M, Pathil A, Warth A, Stenzinger A, et
al: Cadherin-6 is a putative tumor suppressor and target of
epigenetically dysregulated miR-429 in cholangiocarcinoma.
Epigenetics. 11:780–790. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen C, Yang D, Wang Q and Wang X:
Expression and clinical pathological significance of miR-200a in
concurrent cholangio-carcinoma associated with hepatolithiasis. Med
Sci Monit. 21:3585–3590. 2015. View Article : Google Scholar : PubMed/NCBI
|