Introduction
Prostate cancer (PCa) is one of the most common
malignant tumors of the male genitourinary system and the second
leading cause of death among men worldwide (1). There were 1,276,106 new cases and
358,989 deaths (3.8% of all deaths caused by cancer in men)
associated with PCa in 2018 (2).
Risk factors for PCa include age, ethnicity, family history and
diet; among those, age is the most consistent factor (3). The incidence of PCa is highest among
older men, with millions of people suffering from this disease
(4,5). Lifestyle modifications such as
smoking cessation, exercise and weight control may help to decrease
the risk of PCa (6). Primary PCa
is curable; in addition to the combination of surgery and
multiagent chemotherapy, androgen deprivation therapy (ADT) has
been used as the first-line treatment for PCa (7,8).
Although ADT is usually initially effective, the improvement in the
prognosis of PCa is still far from satis fying (9). Since therapeutic agents act by
natural selection to promote the acquisition of new tumor traits,
PCa becomes resistant to ADT, resulting in a fatal outcome known as
castration resistant PCa (CRPC) (10). In addition, recurrent and
metastatic PCa is typically lethal (11). Along with the development of
molecular medicine, various potential key factors in the
development of PCa have been identified. For example, GALNT7 and
ST6GalNAc1 have been reported to be significantly upregulated in
clinical PCa tissues (9). Despite
such advances, more accurate and specific targets still need to be
explored to benefit patients with PCa.
Germ cell-specific gene 2 (GSG2), also termed
haploid germ cell specific nuclear protein kinase, is an atypical
serine/threonine protein kinase that is present in all major
eukaryotic phyla, including yeasts, microsporidia, plants,
nematodes, flies, fish, amphibians and mammals (12). GSG2 has long been considered an
inactive pseudokinase due to its low structural homology with
classical protein kinases (13).
However, GSG2 is responsible for the phosphorylation of histones,
particularly histone H3 at Thr3 (H3T3) during mitosis, which serves
a role in chromosome segregation and thus has a great potential as
an anticancer therapeutic target (14). In addition, phosphorylation of H3T3
by GSG2 creates a recognition motif for docking of the chromosomal
passenger complex, which is essential for the progression of cell
division (15). GSG2
overexpression has been demonstrated to serve an important role in
cancer cells, where it interrupts the normal dissociation of
centromeric cohesion (16), which
leads to a delay before metaphase and ultimately results in
defective mitosis (17). Han et
al (18) have reported that
GSG2 may be considered as a viable anti melanoma target, and the
concomitant inhibition of GSG2 may represent a novel therapeutic
strategy with improved efficacy for the treatment of melanoma.
Nevertheless, the role of GSG2 in PCa has not been reported to the
best of our knowledge and remains largely unclear. Therefore, the
present aimed to demonstrate for the first time the expression of
GSG2 in PCa and its role in the development and progression of PCa,
and to determine the role that GSG2 may serve in the treatment and
prognosis of PCa as a potential therapeutic target.
Materials and methods
Materials
DU 145 (HTB 81) and PC 3 (CRL 1435) cell lines were
purchased from the Cell Bank of the Chinese Academy of Sciences and
cultured with 90% RPMI 1640 (Sigma Aldrich; Merck KGaA) and 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) medium at 37°C in a
humidified incubator containing 5% CO2. TOP10
Escherichia coli competent cells (CB104 03) purchased from
Tiangen Biotech Co., Ltd. were cultured with Luria-Bertani (LB)
liquid medium (1% tryptone, 0.5% yeast extract and 1% NaCl) at 37°C
with gentle agitation. Anti-GSG2 (cat. no. ab21686; Abcam), anti
GAPDH (cat. no. AP0063; Bioworld Technology, Inc.) and anti Ki67
(cat. no. ab16667; Abcam) primary antibodies, HRP conjugated goat
anti rabbit IgG (cat. no. A0208; Beyotime Institute of
Biotechnology) for western blotting, HRP conjugated goat anti
rabbit IgG (cat. no. ab6721; Abcam) for immunohistochemical
staining and western blotting.
Female BALB/c nude mice (4 week old) were purchased
from Shanghai SLAC Laboratory Animal Co. Ltd. and divided into two
groups randomly (n=6 mice/group). All mice were housed under
standard housing conditions as previously described (19).
PCa and normal prostate tissues (>5 cm away from
the PCa tissue) were collected from patients (mean age, 59 years)
who had been diagnosed with PCa and underwent surgical resec tion
in the First Affiliated Hospital of Nanchang University (Nanchang,
China) between May 2015 and May 2017, and the age range of patients
was between 20 and 97 years. Ethical approval was obtained from the
Ethics Committee of the First Affiliated Hospital of Nanchang
University, and written informed consent was obtained from all
patients.
Immunohistochemical staining
PCa and normal prostate tissues from 159 patients
were collected, and the expression of GSG2 in PCa and normal
prostate tissues was detected by immuno histochemistry. Following
dewaxing of the paraffin-embedded sections, antigen retrieval was
performed with citrate buffer, followed by incubation with 3%
H2O2 at room temperature for 10 min. The
paraffin sections were incubated with the primary antibody against
GSG2 (1:200) at 4°C overnight and subse quently incubated with the
secondary antibody (1:400) at room temperature for 30 min. Paraffin
sections were stained with 3-3' diaminobenzidine (DAB) at room
temperature for 10 min and counterstained with hematoxylin at room
temperature for 2 min. For each section, 10 fields (x100
magnification) were selected to be captured using an Olympus
optical microscope (Olympus Corporation) and analyzed. The scoring
standard for GSG2 staining intensity was graded as 0 (negative), 1
(weak), 2 (posi tive ++) and 3 (positive +++). The staining extent
was graded as 0 (0%), 1 (1 25%), 2 (26 50%), 3 (51 75%) or 4 (76
100%). The staining intensity varied from weak to strong. The
sections were classified into negative (0 points), positive (1-4
points), ++ posi tive (5 8 points) or +++ positive (9 12 points)
based on the sum of the staining intensity and staining extent
scores. If the score of the sections was above the median, the
expression of GSG2 was high; if the score was below the median, the
expression of GSG2 was low.
Target gene RNA interference lentiviral
vector preparation
The GSG2 gene was used as a template to design an
RNA interference target sequence (target sequence 5'-CCA CAG GAC
AAT GCT GAA CTT-3') to synthesize a single stranded DNA oligo,
which was then paired to generate double stranded DNA. The negative
control interference target sequence was 5'-TTC TCC GAA CGT GTC ACG
T-3'. Subsequently, the double-stranded DNA was directly ligated
into the double-digested linearized BR-V-108 lentivirus vector
(Shanghai Yibeirui Biomedical Science and Technology Co., Ltd)
overnight. The ligation product was transferred to TOP10 E.
coli competent cells (TIANGEN) with antibiotic free LB liquid
medium and incubated at 37°C for 1 h. The correctly cloned
bacterial solution was screened by bacterial liquid PCR
amplification and sequencing for plasmid extraction. Bacterial
liquid PCR amplification was performed using a Taq Plus Master Mix
(Vazyme Biotech Co., ltd.), forward and reverse primers (GeneRay
Biotech Co., Ltd.), a single TOP10 E. coli cell colony and
ddH2O. The reaction conditions were as follows: 94°C for
3 min, 42 cycles of 94°C for 30 sec, 55°C for 30 sec and 72°C for
30 sec, and a final extension at 72°C for 5 min. The
quality-eligible plasmids were used for lentiviral packaging. The
primer sequences were as follows: Negative control interference
target forward, 5' CCA TGA TTC CTT CAT ATT TGC 3' and reverse, 5'
GTA ATA CGG TTA TCC ACG CG 3'; GSG2 RNA interference target
forward, 5' CCT ATT TCC CAT GAT TCC TTC ATA 3' and reverse, 5' GTA
ATA CGG TTA TCC ACG CG 3'.
Cell infection and fluorescence
immunoassay
DU 145 and PC 3 cells were infected with lentivirus
containing GSG2 interference target sequences (1x107
TU/ml) or negative control interference target sequences
(1x107 TU/ml), and the cells were then cultured at 37°C
for 72 h. A fluorescent micro scope (EMD Millipore) was used to
observe the expression of green fluorescent protein (GFP, carried
by the lentivirus vector) to assess infection efficiency. The
infection efficiency was evaluated by the ratio of fluorescent
cells to total cells (observed under white light).
Reverse transcription-quantitative PCR
(RT-qPCR)
DU 145 and PC 3 cells were collected and centrifuged
at 800 x g for 5 min at room temperature. The supernatant was
removed, and total RNA was isolated using the TRIzol®
reagent (Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The M-MLV Reverse Transcriptase kit
(Promega Corporation) was used to synthesize cDNAs. The reverse
transcription primers, total RNA, M-MLV-RTase and Rnasin were mixed
at 42°C for 1 h and incubated in water at 70°C for 10 min to
inactivate the RT enzyme. The reaction system for qPCR was prepared
by real-time quanti tative PCR instrument (Applied Biosystems;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instruc tions. The reaction system included: SYBR®
Premix Ex Taq (Takara Biotechnology Co., Ltd.), upstream and
downstream primers (GeneRay Biotech Co., Ltd.), reverse
transcription products and RNase Free H2O. The reaction
conditions were as follows: 95°C for 30 sec (predenaturation), 95°C
for 15 sec (denaturation) and 60°C for 10 sec (annealing) for a
total of 42 cycles, and 72°C for 5 min (elongation). GAPDH was used
as the internal reference. The relative expression of genes was
calculated using the 2ΔΔCq method (20). Each experiment was repeated three
times. The primer sequences were as follows: GAPDH forward, 5' TGA
CTT CAA CAG CGA CAC CCA 3' and reverse, 5' CAC CCT GTT GCT GTA GCC
AAA 3'; GSG2 forward, 5' GGA AGG GGT GTT TGG CGA AGT 3' and
reverse, 5' TGA GGA GCA AGG GAG GGT AAG 3'.
Western blot assay
The expression levels of GSG2 in PCa cell lines and
tumor tissues from mice were detected by western blotting. DU 145
and PC 3 cells were collected and lysed with RIPA lysis buffer
(Beyotime Institute of Biotechnology) containing protease
inhibitors on ice according to the manu facturer's instructions.
BCA Protein Assay kit (HyClone; GE Healthcare Life Sciences) was
used to detect the concentration of the extracted protein. SDS PAGE
(10%) was performed to isolate the total cellular proteins with 20
µg protein/lane, which were then transferred to PVDF membranes. The
membranes were blocked with TBS + 0.1% Tween 20 (TBST) solution
containing 5% skimmed milk for 60 min at room tempera ture and
incubated with the primary anti GSG2 (1:1,000) and anti-GAPDH
(1:3,000) antibodies overnight at 4°C. After washing with TBST, the
membranes were incubated with the HRP conjugated goat anti rabbit
IgG polyclonal antibody (1:3,000) for 2 h at room temperature.
Color development for signal detection was performed using the ECL™
Prime Western Blotting Detection Reagent kit (Amersham; GE
Healthcare Life Sciences). The experiment was performed in
triplicate.
Celigo cell count assay
Short hairpin (sh)RNA lenti virus infected DU 145
and PC 3 cells at the logarithmic growth phase were trypsinized,
resuspended in complete medium (RPMI 1640 + 10% FBS), counted and
seeded in 96 well plates (2,000 cells/well). The next day, the
plate was detected by Celigo Imaging Cytometry System (Nexcelom
Bioscience) to count the cells, which was repeated once a day for 5
consecutive days. This experiment was performed independently three
times.
Fluorescence-activated cell sorting
(FACS) assay
Apoptosis was determined by FACS assay. Following
infection with shRNA lentivirus, DU 145 and PC 3 cells were
cultured in 6 well plates until the cell density reached 85%. The
cells were trypsinized and centrifuged to collect the cell pellet,
which was washed with D-Hank's solution precooled at 4°C. Cell
suspensions (100 µl; 1x105 1x106 cells) were
stained with 5 µl Annexin V-APC (cat. no. 10010-09; Southern
Biotech) in the dark at room temperature for 15 min, centrifuged at
1,000 x g at room temperature for 3 min and resuspended.
Subsequently, 5 µl propidium iodide (PI) was added to the cell
suspension, which was replenished to 300 µl with the cell dye
buffer from the apoptosis kit. The cells were detected with a Guava
easy Cyte HT flow cytometer (EMD Millipore) and analyzed by FlowJo
VX10 (FlowJo LLC). In the graph of apoptosis results, the lower
left quadrant represented living cells and the two right quadrants
represented apoptotic cells, of which the upper right quadrant
represented late apoptotic cells and the lower right quadrant
represented early apoptotic cells. The experi ment was performed in
triplicate.
Colony formation assay
Three days after shRNA lentivirus infection, DU 145
and PC 3 cells in the logarithmic growth phase were digested with
trypsin, resuspended in complete medium and inoculated into 6 well
plates (1,000 cells/well). Cells were cultured for additional 8
days, and the medium was changed every 3 days. Subsequently, the
cells were fixed with 4% paraformaldehyde at room temperature for 1
h after washing with PBS. Giemsa was used for staining the cells at
room temperature for 20 min, and double distilled H2O
was used to wash the cells three times. The cells were dried and
images were captured using a digital camera to count the number of
colonies containing >50 cells. The assay was performed in
triplicate.
Tumor-bearing animal model
All animal experiments were approved and performed
under the supervision of the Institutional Animal Care and Use
Ethics Committee of the First Affiliated Hospital of Nanchang
University. PC-3 cells infected with the shRNA lentivirus at the
logarithmic growth phase were digested with trypsin, resuspended
and subcu taneously injected into the right forelimb armpit of each
nude mouse (200 µl suspension containing 4x106
cells/ml). All mice were maintained for 35 days, during which the
volume of tumors in the mice was measured nine times with a Vernier
caliper. On day 35, the mice were intraperitoneally injected with
D-Luciferin (10 µg) for 15 min, anesthetized by intraperitoneal
injection of 0.7% pentobarbital sodium (70 mg/kg) and placed under
a small animal multispectral living imaging system for imaging
(Lumina LT; PerkinElmer, Inc.). Subsequently, the mice were
sacrificed by cervical dislocation, and tumors were dissected from
the mice to measure the tumor volume and weight. The formula for
the calculation of the tumor volume in mm3 was as
follows: V=π/6 x L x W2, where V is the tumor volume, L
is the tumor length and W is the tumor width. In addition, the
expression of GSG2 in tumor tissues from mice in the control
(shCtrl) and GSG2 knockdown (shGSG2) groups was detected by western
blotting as described above, and the isolated tumors were preserved
in liquid nitrogen at -80°C.
Ki-67 staining
Paraffin sections of tumor tissues removed from mice
were dewaxed, rehydrated in a descending ethanol gradient (100, 95,
80 and 70%) and incubated with an anti Ki 67 antibody (1:200) at
4°C overnight. After washing with PBS, the paraffin sections were
incubated with a secondary antibody (1:400) for 2 h at room
temperature, and then counterstained with hematoxylin and observed
under an Olympus optical microscope (x100 or x200 magnification) to
analyze Ki-67 expression. A total of 10 fields were selected for
each section to be captured and analyzed. The experiment was
performed in triplicate.
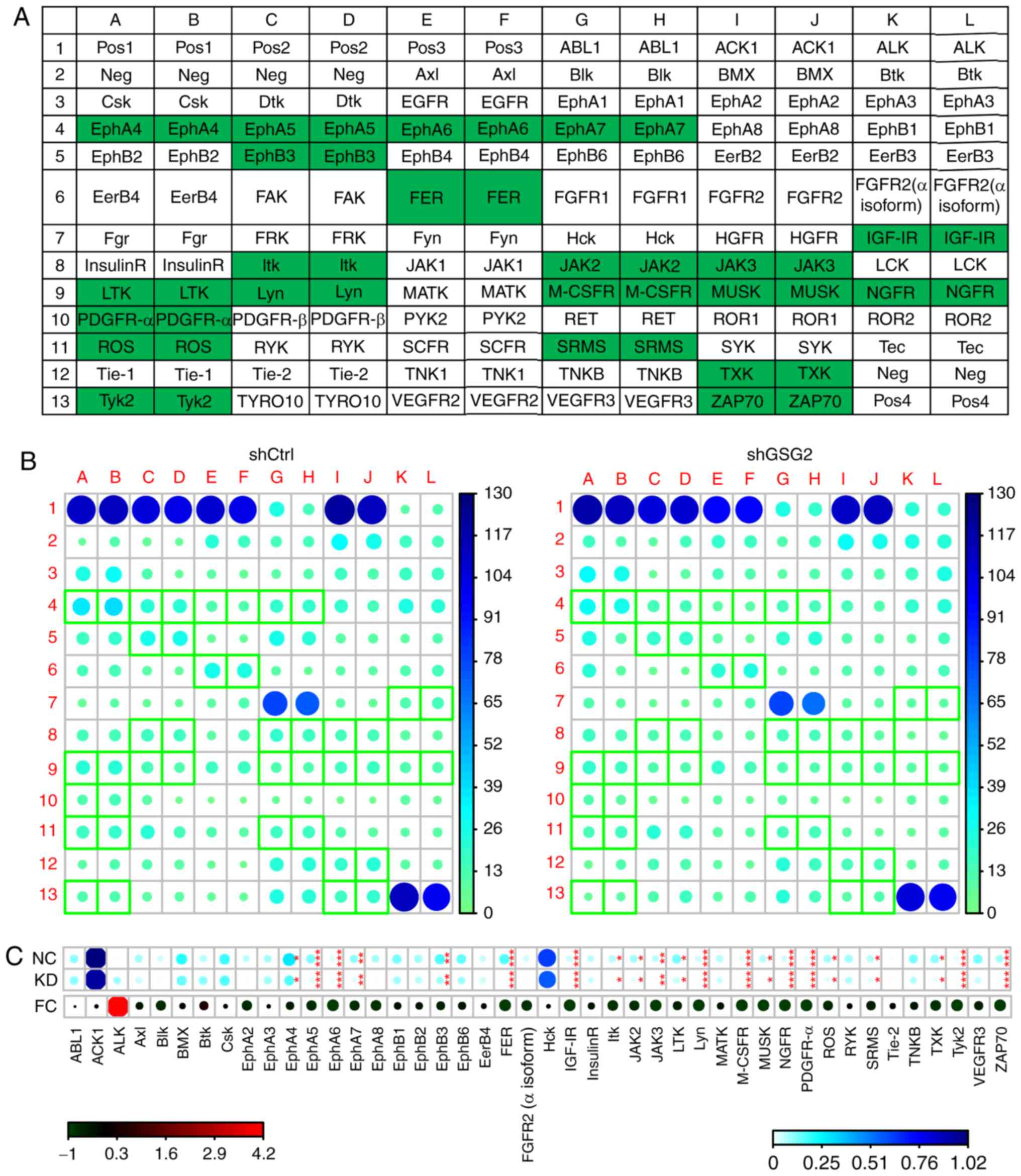
Human RTK Phosphorylation Antibody
Array
To investigate the potential downstream signaling
pathway and functional targets of GSG2 in PC 3 cells, Human RTK
Phosphorylation Antibody Array (cat. no. ab193662; Abcam) was used.
PC 3 cells infected with shCtrl or shGSG2 were resuspended and
lysed with 1X lysis buffer for 30 min. Simultaneously, the
membranes were blocked with 2 ml 1X Blocking Buffer for 30 min at
room temperature. Subsequently, the samples were pipetted into
wells and incubated overnight at 4°C. Following washing, 1 ml 1X
biotinylated anti phosphotyrosine antibody was added to each well
and incubated overnight at 4°C. Next, 2 ml 1X HRP conjugated
streptavidin was added into each well and incubated for 2 h at room
temperature. After the membranes were washed, any excess washing
buffer was removed by blotting the membrane edges, and 500 µl detec
tion buffer mixture (equal volumes of Detection Buffer C and
Detection Buffer D) was added to each membrane for 2 min at room
temperature. The signal density was detected using a
chemiluminescence imaging system and analyzed by ImageJ software
version 1.8.0 (National Institutes of Health). The experiment was
performed in duplicate.
Statistical analysis
Data were analyzed using GraphPad Prism 6 software
(GraphPad Software, Inc.) and presented as the mean ± standard
deviation. Unpaired Student's t test was used to compare the
differences between two groups. The differences in GSG2 expression
between patients with PCa with different clinicopathological
characteristics were compared using the chi-square test. The
correlation between GSG2 expression and clinicopathological
characteristics was analyzed by Spearman rank correlation analysis.
The plot histogram of GSG2 related signaling molecules in cancer
cells was produced using SignaLink 2.0 analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
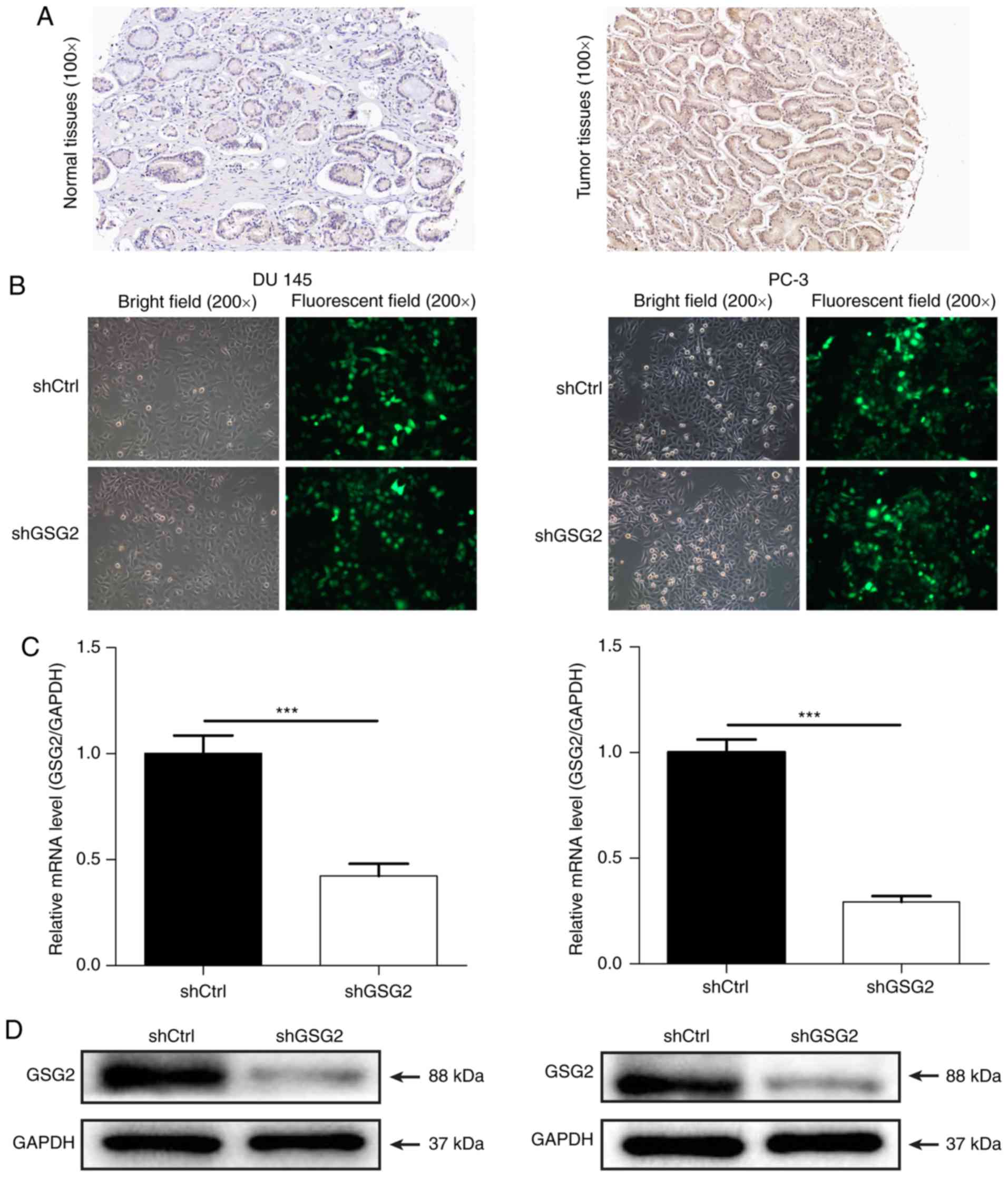
Expression of GSG2 in clinical PCa
tissues
To investigate the role of GSG2 in the development
and progression of PCa, the expression of GSG2 was detected in
clinical PCa and para carcinoma tissues by immunohistochemical
staining. As presented in Fig. 1A,
the results indicated a cytoplasmic localization of GSG2 and
demonstrated that the expres sion of GSG2 in PCa tissues was
significantly upregulated compared with that in para carcinoma
tissues (Table I). In addition,
the association between the expression of GSG2 and the
clinicopathological characteristics of patients with PCa was
evaluated by statistical analysis. The results indicated
significant differences in GSG2 expression levels among patients
with different Gleason scores (grade) and pathological grades
(P<0.05; Table II). Spearman
rank correlation analysis revealed that the Gleason score (grade)
and pathological grade were weakly positively correlated with GSG2
expression levels (Table III).
These results demonstrated that the gene expression of GSG2 was
significantly associated with the development and progression of
PCa.
 | Table IGSG2 expression patterns in prostate
cancer tissues and para carcinoma tissues determined by
immunohistochem istry analysis. |
Table I
GSG2 expression patterns in prostate
cancer tissues and para carcinoma tissues determined by
immunohistochem istry analysis.
| GSG2
expression | Tumor tissue
| Para carcinoma
tissue
| P value |
|---|
| N | % | N | % |
|---|
| Low | 93 | 58.5 | 80 | 100 | 0.000a |
| High | 66 | 41.5 | 0 | 0 | |
| Table IIRelationship between GSG2 expression
levels and clinicopathological characteristics of patients with
prostate cancer. |
Table II
Relationship between GSG2 expression
levels and clinicopathological characteristics of patients with
prostate cancer.
| Characteristic | | GSG2 expression
| P-value |
|---|
No. of patients
| Low
| High
|
|---|
| N | % | N | % | N | % |
|---|
| All patients | 159 | | 93 | 58.5 | 66 | 41.5 | |
| Age, years | | | | | | | 0.184 |
| ≤69 | 84 | 52.8 | 45 | 28.3 | 39 | 24.5 | |
| >69 | 75 | 47.2 | 48 | 30.2 | 27 | 17.0 | |
| Gleason Score | | | | | | | 0.043a |
| <8 | 60 | 37.7 | 40 | 25.2 | 20 | 12.6 | |
| ≥8 | 92 | 57.9 | 46 | 28.9 | 46 | 28.9 | |
| Grade | | | | | | | 0.005b |
| 1 | 12 | 7.5 | 10 | 6.3 | 2 | 1.3 | |
| 2 | 38 | 23.9 | 26 | 16.4 | 12 | 7.5 | |
| 3 | 102 | 64.2 | 50 | 31.4 | 52 | 32.7 | |
| T Infiltrate | | | | | | | 0.553 |
| T1 | 2 | 1.3 | 2 | 1.3 | 0 | 0.0 | |
| T2 | 68 | 42.8 | 49 | 30.8 | 19 | 11.9 | |
| T3 | 38 | 23.9 | 28 | 17.6 | 10 | 6.3 | |
| T4 | 6 | 3.85 | 6 | 3.8 | 0 | 0.0 | |
| Lymphatic
metastasis | | | | | | | 0.088 |
| N0 | 106 | 66.7 | 77 | 48.4 | 29 | 18.2 | |
| N1 | 8 | 5.0 | 8 | 5.0 | 0 | 0.0 | |
| Stage | | | | | | | 0.833 |
| 1 | 14 | 8.8 | 13 | 8.2 | 1 | 0.6 | |
| 2 | 56 | 35.2 | 38 | 23.9 | 18 | 11.3 | |
| 3 | 32 | 20.1 | 22 | 13.8 | 10 | 6.3 | |
| 4 | 12 | 7.5 | 12 | 7.5 | 0 | 0.0 | |
| Gleason grade | | | | | | | 0.026a |
| 2 | 8 | 5.0 | 6 | 3.8 | 2 | 1.3 | |
| 3 | 38 | 23.9 | 28 | 17.6 | 10 | 6.3 | |
| 4 | 54 | 34.0 | 26 | 16.4 | 28 | 17.6 | |
| 5 | 51 | 32.1 | 25 | 15.7 | 26 | 16.4 | |
| 6 | 1 | 0.6 | 1 | 0.6 | 0 | 0.0 | |
| Table IIICorrelation between GSG2 expression
and tumor characteristics in patients with prostate cancer. |
Table III
Correlation between GSG2 expression
and tumor characteristics in patients with prostate cancer.
| Characteristic | N | Spearman's r | P value |
|---|
| Gleason score | 152 | 0.164 | 0.043a |
| Gleason grade | 152 | 0.181 | 0.025a |
| Grade | 152 | 0.227 | 0.005b |
Knockdown of GSG2 in PCa cells
To investigate the role of GSG2 in PCa, shRNA
targeting GSG2 was cloned into lenti viral vectors carrying GFP.
Subsequently, shGSG2 or shCtrl lentivirus was infected into human
PCa cells PC 3 and DU 145. As presented in Fig. 1B, the fluorescence intensity in
cells infected with shGSG2 or shCtrl revealed >80% transfection
efficiency in the two cell lines. The relative expression levels of
GSG2 in PC-3 and DU 145 cells were detected by RT-qPCR; the results
demonstrated that the GSG2 mRNA level was reduced by 70.8% in PC 3
cells and by 57.7% in DU 145 cells in the shGSG2 groups compared
with the shCtrl groups (P<0.05; Fig. 1C). In addition, western blotting
also demonstrated that GSG2 protein expression was significantly
downregulated post infection compared with that in cells infective
with shCtrl (Fig. 1D). These
results indicated the successful construction of a GSG2 knockdown
cell model.
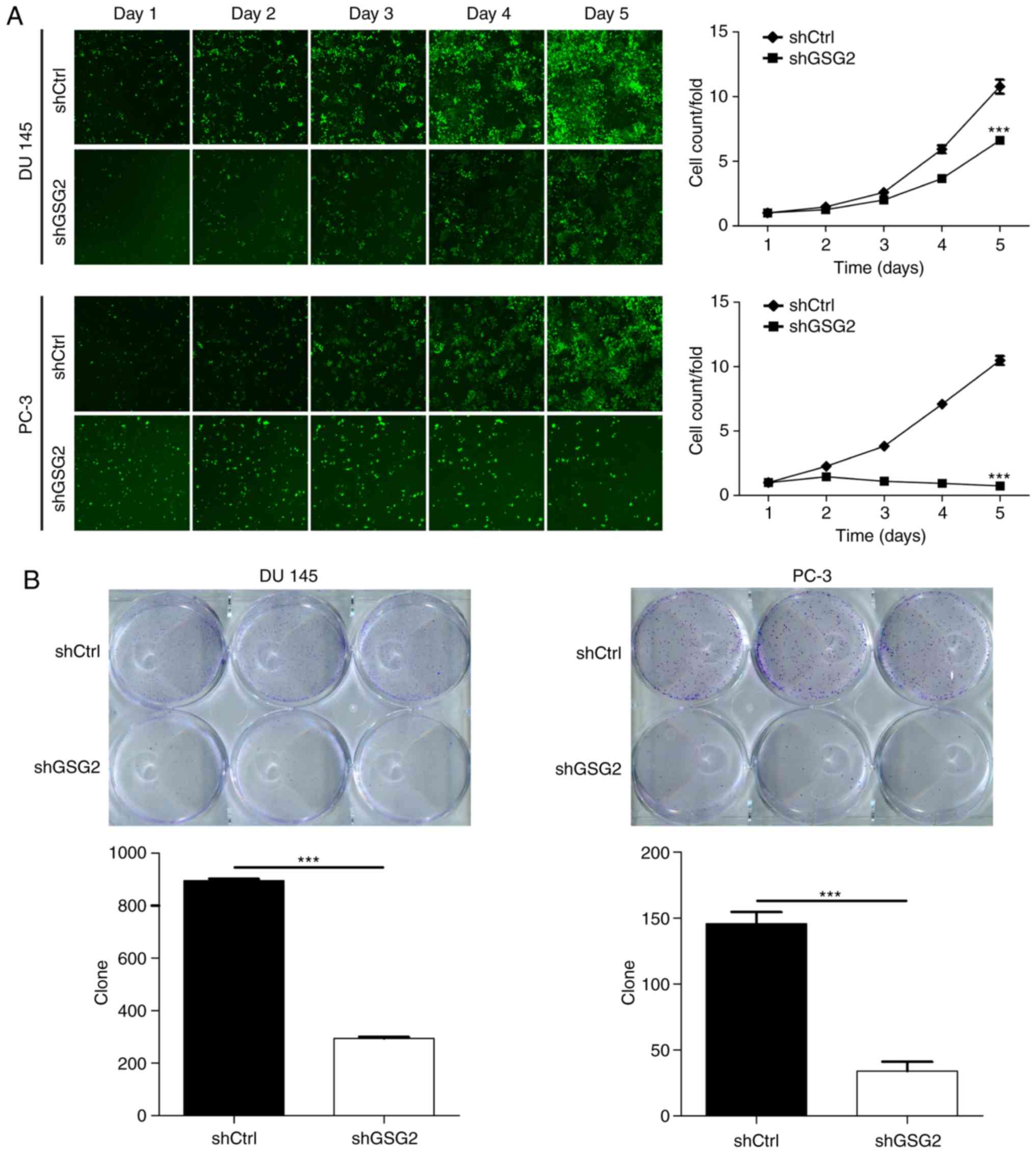
Knockdown of GSG2 inhibits PCa cell
proliferation and promotes apoptosis
In order to examine the effects of GSG2 on cell
proliferation, Celigo Imaging Cytometry System was used for
analysis of the cell growth curve. As presented in Fig. 2A, cell proliferation was
significantly inhibited in the shGSG2 group compared with that in
the shCtrl group in PC 3 and DU 145 cells. In addition, colony
formation assay was performed to assess the colony formation
ability of PCa cells, which is an important feature of malignant
tumors. The number of cell colo nies was significantly decreased by
80.0 and 66.7%, respectively, in the shGSG2 group of PC 3 and DU
145 cells compared with that of the shCtrl group (P<0.05;
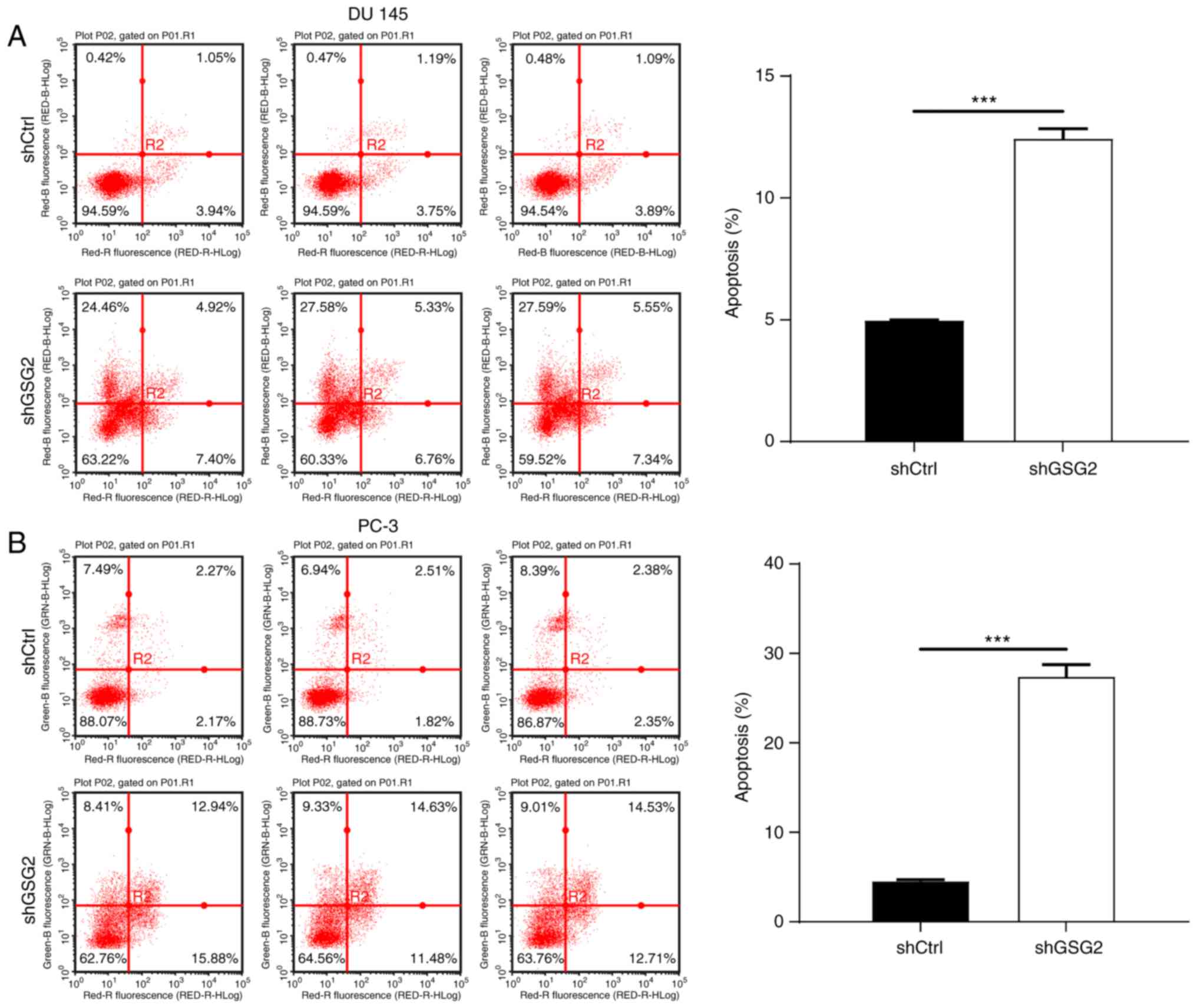
Fig. 2B). To detect the effects of
GSG2 knockdown on the apoptosis of PCa cells, FACS was used.
Compared with that of the shCtrl group, the percentage of apoptotic
cells in the shGSG2 group was increased 2.5 fold in PC 3 cells and
6 fold in DU 145 cells (P<0.05; Fig. 3), suggesting that GSG2 knockdown
promoted the apoptosis of PCa cells.
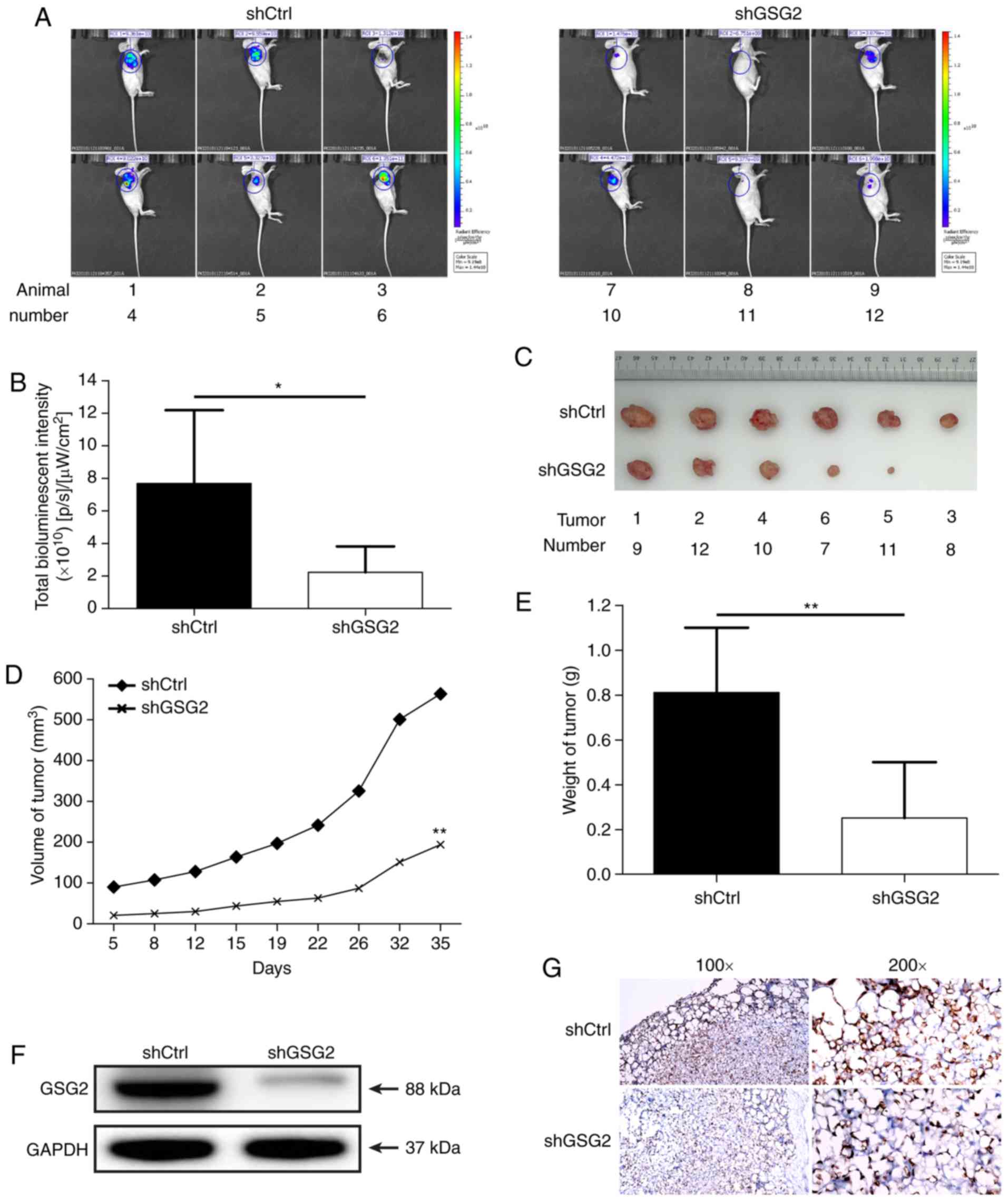
Effects of GSG2 knockdown on tumor
progression in vivo
To determine the potential of shGSG2 as a
therapeutic target for PCa therapy, a nude mouse xenograft model
was constructed using GSG2 knockdown PC 3 cells. At 72 h post
infection of PC 3 cells with shGSG2 or shCtrl lentivirus, the
infection efficiency was determined to be 90-100% (Fig. S1), and the mice were
subcutaneously injected with these cells. The levels of
bioluminescence intensity (µW/cm2) were measured by
in vivo imaging of anaesthetized mice following an injection
of D luciferin. The bioluminescence intensity in the shGSG2 group
was ~71% lower compared with that in the shCtrl group (P<0.05;
Fig. 4A and B). The results
suggested that tumor growth was slower in the shGSG2 group compared
with the shCtrl group (P<0.05). In addition, according to the
tumor volume and weight, tumors from mice in the shGSG2 group were
66% smaller in diameter, and the tumor weight was 69% lower
compared with those in the shCtrl group (P<0.05; Fig. 4C E). The expression of GSG2 in
tumor tissues from mice was inhibited in the shGSG2 group compared
with the shCtrl group (Fig. 4F).
Ki 67 staining demonstrated that the proliferation of PCa cells was
significantly inhibited in the shGSG2 group compared with that in
the shCtrl group (Fig. 4G). These
results confirmed that GSG2 knockdown suppressed tumor development
in vivo. Taken together, these results indicated that
targeting GSG2 with shGSG2 may have an inhibitory effect on PCa
in vivo.
Mechanism of GSG2 knockdown in PC-3
cells
To investigate the regulatory mechanism of GSG2 in
the tumor develop ment of PCa, Human RTK Phosphorylation Antibody
Array was used in the shGSG2 and shCtrl groups of PC 3 cells for
exploring potential downstream signaling pathways and func tional
targets that mediate the effect of GSG2 knockdown on PCa. Among
them, the levels of ephrin receptor A4 (EphA4), EphA5, EphA6,
EphA7, EphB3, FER tyrosine kinase (FER), insulin like growth factor
(IGF) IR, interleukin 2 inducible T cell kinase (Itk), Janus kinase
2 (JAK2), JAK3, leukocyte receptor tyrosine kinase (LTK), Lyn proto
oncogene Src family tyrosine kinase (Lyn), macrophage colony
stimulating factor 1 receptor (M CSFR), muscle associated receptor
tyro sine kinase (MUSK), nerve growth factor receptor (NGFR),
platelet derived growth factor receptor α (PDGFR α), ROS proto
oncogene 1 receptor tyrosine kinase (ROS), tyrosine protein kinase
Srms (SRMS), TXK tyrosine kinase (TXK), tyrosine kinase 2 (Tyk2)
and zeta chain of T cell receptor associated protein kinase 70
(ZAP70) were determined to be significantly downregulated in PC-3
cells following GSG2 knockdown compared with cells infected with
shCtrl (P<0.05; Fig. 5A D),
indicating the potential func tional targets in the regulatory
effect of GSG2 on PCa and guiding the direction of our future
work.
Discussion
The treatment of PCa has attracted increasing
attention in recent years (12).
Previous studies have reported numerous gene targets that were able
to inhibit cell proliferation and promote apoptosis in PCa
(21 23). A recent study has suggested that
certain natural products, such as green tea, can effectively induce
apoptosis and inhibit invasion of PCa cells via the PI3K/Akt
signaling pathway (24). However,
more efficient and accurate targets still need to be explored.
The GSG2 kinase is a member of the eukaryotic
protein kinase (ePK) family that structurally diverges from the
majority of ePKs (25). A number
of studies have tried to use small molecule inhibitors to determine
the functions of GSG2 in mitosis (26,27);
to the best of our knowledge, histone H3 phosphorylated by GSG2 at
threonine 3 (H3T3ph) is the only currently known product of GSG2
activity; thus, GSG2 inhibitors strongly reduce the levels of
H3T3ph in cells (28). However,
research on GSG2 in prostate tumors is rare. The present study
initially detected GSG2 expression in PCa tissues by
immunohistochemical staining, and the results revealed that GSG2
expression levels in PCa tissues were significantly higher compared
with those in para-carcinoma tissues. Therefore, to study the role
of GSG2 in PCa cells, PC 3 and DU 145 cells were selected for the
construction of GSG2 knockdown cell models.
Kim et al (16,29)
demonstrated that the inhibition of GSG2 activity by coumestrol
inhibited the proliferation of cancer cells. In our study, Celigo
cell counting assay was performed to demonstrate the effect of GSG2
knockdown on PCa cell proliferation, and the results demonstrated
that cell prolifera tion was significantly inhibited in the shGSG2
group compared with that in the shCtrl group, which was consistent
with the results of the colony formation assay. The results of the
present study also revealed that knockdown of GSG2 increased the
apoptotic rate of PC 3 and DU 145 cells. In addition, it was
observed that the growth of PCa tumors in vivo was signifi
cantly inhibited in the shGSG2 group compared with that in the
shCtrl group, which was also confirmed by the relatively lower Ki
67 expression in the tumors of the shGSG2 group. The levels of
bioluminescence intensity also revealed marked inhibition of tumor
growth in the shGSG2 group.
Zabala Letona et al (30) have reported that the production of
polyamine is a hallmark of highly proliferating cells, and an
essential metabolic route for oncogenicity is mechanistic target of
rapamycin complex 1, which regulates polyamine dynamics. The
present study preliminarily screened the regulatory mechanism of
GSG2 in tumor development in PCa. The results revealed that
knockdown of GSG2 inhibited the phosphorylation of EphA4, EphA5,
EphA6, EphA7, EphB3, FER, IGF IR, Itk, JAK2, JAK3, LTK, Lyn, M
CSFR, MUSK, NGFR, PDGFR α, ROS, SRMS, TXK, Tyk2 and ZAP70 in PCa
cells compared with those infected with shCtrl. According to
previous studies, EphA4, EphA5, EphA6 and EphA7 promote
angiogenesis and PCa metastasis, and are associated with human PCa
progres sion (29,31 33).
FER kinase serves a central role in breast cancer metastasis and
PCa cell proliferation (34). IGF
serves key roles in the progression of different types of cancer,
such as breast, prostate, lung, ovarian and skin cancer (35). The JAK2 signaling pathway is
involved in cytokine signaling, which regulates hematopoietic cell
development, cell metabolism and immune responses (36). In addition, LTK shares a high
degree of similarity with anaplastic lymphoma kinase, which is
mutated in human cancers including adenocarcinomas of the lung,
neuroblastomas, breast and esophageal cancers (37). TXK is a member of the Tec family of
tyrosine kinases, which acts as Th1 cell-specific transcription
factor (38). These results
suggested that GSG2 may serve a crucial role in the establish ment
and progression of PCa.
The present study had certain limitations, including
the low number of clinical specimens. In addition, the specific
downstream genes and regulatory mechanisms must be inves tigated
and verified in the future. Further research is needed to discover
the prognostic significance of GSG2 in PCa and the role of GSG2 in
different PCa cell lines.
In conclusion, the present study is the first to
link GSG2 to PCa progression. The results of the present study
suggested that GSG2 knockdown inhibited the development and progres
sion of PCa. Thus, GSG2 may be a potential therapeutic target for
PCa treatment.
Supplementary Data
Funding
This work was financially supported by National
Natural Science Foundation (grant nos. 81001144 and 81360379),
Jiangxi Natural Science Foundation (grantno. 20181BAB205056),
Jiangxi Provincial Department of Education Project (grant no.
170056) and Zhejiang Province Natural Science Foundation (grant no.
LY16H280001). The funding bodies had no role in the design of the
study, data collection, analysis, and the writing of the
manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
AX designed the study. FY and YL conceived and
coordi nated the study, performed experiments, analyzed the data
and wrote the manuscript. XX, WL, DT and YZ collected and analyzed
the data and revised the manuscript. XZ and GW supervised the study
and participated in the writing of the manuscript. All authors
reviewed the results and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Ethical approval was obtained from the Ethics
Committee of the First Affiliated Hospital of Nanchang University
and informed written consent was obtained from all patients. All
animal experiments were approved and performed under the supervi
sion of the Institutional Animal Care and Use Ethics Committee of
the First Affiliated Hospital of Nanchang University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
PCa
|
prostate cancer
|
|
GSG2
|
germ cell specific gene 2 protein
|
|
ePK
|
eukaryotic protein kinase
|
|
H3T3ph
|
histone H3 phosphorylated by GSG2 at
threonine 3
|
|
ADT
|
androgen deprivation therapy
|
|
FACS
|
fluorescence activated cell
sorting
|
|
shRNA
|
short hairpin RNA
|
|
shGSG2
|
cells transfected with GSG2-targeting
shRNA
|
|
shCtrl
|
cells transfected with control
shRNA
|
References
|
1
|
Weng CC, Ding PY, Liu YH, Hawse JR,
Subramaniam M, Wu CC, Lin YC, Chen CY, Hung WC and Cheng KH: Mutant
Kras induced upregulation of CD24 enhances prostate cancer stemness
and bone metastasis. Oncogene. 38:2005–2019. 2019. View Article : Google Scholar
|
|
2
|
Rawla P: Epidemiology of prostate cancer.
World J Oncol. 10:63–89. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fong LY, Jing R, Smalley KJ, Wang ZX,
Taccioli C, Fan S, Chen H, Alder H, Huebner K, Farber JL, et al:
Human like hyper plastic prostate with low ZIP1 induced solely by
Zn deficiency in rats. Proc Natl Acad Sci USA. 115:E11091–E11100.
2018. View Article : Google Scholar
|
|
4
|
Barry MJ and Simmons LH: Prevention of
prostate cancer morbidity and mortality: Primary prevention and
early detection. Med Clin North Am. 101:787–806. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Daniyal M, Siddiqui ZA, Akram M, Asif HM,
Sultana S and Khan A: Epidemiology, etiology, diagnosis and
treatment of prostate cancer. Asian Pac J Cancer Prev.
15:9575–9578. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cuzick J, Thorat MA, Andriole G, Brawley
OW, Brown PH, Culig Z, Eeles RA, Ford LG, Hamdy FC, Holmberg L, et
al: Prevention and early detection of prostate cancer. Lancet
Oncol. 15:e484–e492. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pezaro C, Woo HH and Davis ID: Prostate
cancer: Measuring PSA. Intern Med J. 44:433–440. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mohamad NV, Soelaiman IN and Chin KY: A
review on the effects of androgen deprivation therapy (ADT) on bone
health status in men with prostate cancer. Endocr Metab Immune
Disord Drug Targets. 17:276–284. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Munkley J, Vodak D, Livermore KE, James K,
Wilson BT, Knight B, Mccullagh P, Mcgrath J, Crundwell M, Harries
LW, et al: Glycosylation is an androgen regulated process essential
for prostate cancer cell viability. EBioMedicine. 8:103–116. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Reina Campos M, Linares JF, Duran A,
Cordes T, L'Hermitte A, Badur MG, Bhangoo MS, Thorson PK, Richards
A, Rooslid T, et al: Increased serine and one carbon pathway
metabolism by PKClambda/iota deficiency promotes neuroendo crine
prostate cancer. Cancer Cell. 35:385–400.e9. 2019. View Article : Google Scholar
|
|
11
|
Xu K, Ganapathy K, Andl T, Wang Z, Copland
JA, Chakrabarti R and Florczyk SJ: 3D porous chitosan-alginate
scaffold stiffness promotes differential responses in prostate
cancer cell lines. Biomaterials. 217:1193112019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dai J, Sultan S, Taylor SS and Higgins JM:
The kinase haspin is required for mitotic histone H3 Thr 3
phosphorylation and normal metaphase chromosome alignment. Genes
Dev. 19:472–488. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kestav K, Uri A and Lavogina D: Structure,
Roles and inhibi tors of a mitotic protein kinase haspin. Curr Med
Chem. 24:2276–2293. 2017. View Article : Google Scholar
|
|
14
|
Opoku Temeng C, Dayal N, Aflaki
Sooreshjani M and Sintim HO: 3H-pyrazolo[4,3-f]quinoline haspin
kinase inhibi tors and anticancer properties. Bioorg Chem.
78:418–426. 2018. View Article : Google Scholar
|
|
15
|
Lavogina D, Kestav K, Chaikuad A, Heroven
C, Knapp S and Uri A: Co crystal structures of the protein kinase
haspin with bisubstrate inhibitors. Acta Crystallogr F Struct Biol
Commun. 72:339–345. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim JE, Lee SY, Jang M, Choi HK, Kim JH,
Chen H, Lim TG, Dong Z and Lee KW: Coumestrol epigenetically
suppresses cancer cell proliferation: Coumestrol is a natural
haspin kinase inhibitor. Int J Mol Sci. 18:pii: E2228. 2017.
View Article : Google Scholar
|
|
17
|
Amoussou NG, Bigot A, Roussakis C and
Robert JH: Haspin: A promising target for the design of inhibitors
as potent anticancer drugs. Drug Discov Today. 23:409–415. 2018.
View Article : Google Scholar
|
|
18
|
Han L, Wang P, Sun Y, Liu S and Dai J:
Anti melanoma activi ties of haspin inhibitor CHR 6494 deployed as
a single agent or in a synergistic combination with MEK inhibitor.
J Cancer. 8:2933–2943. 2017. View Article : Google Scholar :
|
|
19
|
Scheff NN, Alemu RG, Klares R III, Wall
IM, Yang SC, Dolan JC and Schmidt BL: Granulocyte colony
stimulating factor induced neutrophil recruitment provides opioid
mediated endogenous anti-nociception in female mice with oral
squamous cell carci noma. Front Mol Neurosci. 12:2172019.
View Article : Google Scholar
|
|
20
|
Pfaffl MW: Pfaffl A new mathematical model
for relative quan tification in real-time RT-PCR. Nucleic Acids
Res. 29:e452001. View Article : Google Scholar
|
|
21
|
Metzger E, Wang S, Urban S, Willmann D,
Schmidt A, Offermann A, Allen A, Sum M, Obier N, Cottard F, et al:
KMT9 monomethylates histone H4 lysine 12 and controls proliferation
of prostate cancer cells. Nat Struct Mol Biol. 26:361–371. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fong KW, Zhao JC, Song B, Zheng B and Yu
J: TRIM28 protects TRIM24 from SPOP mediated degradation and
promotes pros tate cancer progression. Nat Commun. 9:50072018.
View Article : Google Scholar
|
|
23
|
Canesin G, Evans Axelsson S, Hellsten R,
Sterner O, Krzyzanowska A, Andersson T and Bjartell A: The STAT3
inhibitor galiellalactone effectively reduces tumor growth and
metastatic spread in an orthotopic xenograft mouse model of
prostate cancer. Eur Urol. 69:400–404. 2016. View Article : Google Scholar
|
|
24
|
Wang Z, Wang Y, Zhu S, Liu Y, Peng X,
Zhang S, Zhang Z, Qiu Y, Jin M, Wang R, et al: DT 13 inhibits
proliferation and metastasis of human prostate cancer cells through
blocking PI3K/Akt pathway. Front Pharmacol. 9:14502018. View Article : Google Scholar
|
|
25
|
Maiolica A, de Medina Redondo M, Schoof
EM, Chaikuad A, Villa F, Gatti M, Jeganathan S, Lou HJ, Novy K,
Hauri S, et al: Modulation of the chromatin phosphoproteome by the
Haspin protein kinase. Mol Cell Proteomics. 13:1724–1740. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Patnaik D, Jun X, Glicksman MA, Cuny GD,
Stein RL and Higgins JM: Identification of small molecule
inhibitors of the mitotic kinase haspin by high throughput
screening using a homogeneous time resolved fluorescence resonance
energy transfer assay. J Biomol Screen. 13:1025–1034. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Balzano D, Santaguida S, Musacchio A and
Villa F: A general framework for inhibitor resistance in protein
kinases. Chem Biol. 18:966–975. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang F, Ulyanova NP, Daum JR, Patnaik D,
Kateneva AV, Gorbsky GJ and Higgins JM: Haspin inhibitors reveal
centro meric functions of Aurora B in chromosome segregation. J
Cell Biol. 199:251–268. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li S, Wu Z, Ma P, Xu Y, Chen Y, Wang H, He
P, Kang Z, Yin L, Zhao Y, et al: Ligand dependent EphA7 signaling
inhibits pros tate tumor growth and progression. Cell Death Dis.
8:e31222017. View Article : Google Scholar
|
|
30
|
Zabala Letona A, Arruabarrena Aristorena
A, Martín Martín N, Fernandez-Ruiz S, Sutherland JD, Clasquin M,
Tomas-Cortazar J, Jimenez J, Torres I, Quang P, et al: mTORC1
dependent AMD1 regulation sustains polyamine metabolism in prostate
cancer. Nature. 547:109–113. 2017. View Article : Google Scholar
|
|
31
|
Jing X, Sonoki T, Miyajima M, Sawada T,
Terada N, Takemura S and Sakaguchi K: EphA4 deleted
microenvironment regulates cancer development and leukemoid
reaction of the isografted 4T1 murine breast cancer via reduction
of an IGF1 signal. Cancer Med. 5:1214–1227. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li S, Zhu Y, Ma C, Qiu Z, Zhang X, Kang Z,
Wu Z, Wang H, Xu X, Zhang H, et al: Downregulation of EphA5 by
promoter methylation in human prostate cancer. BMC Cancer.
15:182015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li S, Ma Y, Xie C, Wu Z, Kang Z, Fang Z,
Su B and Guan M: EphA6 promotes angiogenesis and prostate cancer
metastasis and is associated with human prostate cancer
progression. Oncotarget. 6:22587–22597. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ivanova IA, Vermeulen JF, Ercan C,
Houthuijzen JM, Saig FA, Vlug EJ, van der Wall E, van Diest PJ,
Vooijs M and Derksen PW: FER kinase promotes breast cancer
metastasis by regulating α6 and β1 integrin dependent cell adhesion
and anoikis resistance. Oncogene. 32:5582–5592. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vishwamitra D, George SK, Shi P, Kaseb AO
and Amin HM: Type I insulin like growth factor receptor signaling
in hemato logical malignancies. Oncotarget. 8:1814–1844. 2017.
View Article : Google Scholar
|
|
36
|
Jay J, Hammer A, Nestor Kalinoski A and
Diakonova M: JAK2 tyrosine kinase phosphorylates and is negatively
regulated by centrosomal protein Ninein. Mol Cell Biol. 35:111–131.
2015. View Article : Google Scholar :
|
|
37
|
Roll JD and Reuther GW: ALK activating
homologous mutations in LTK induce cellular transformation. PLoS
One. 7:e317332012. View Article : Google Scholar
|
|
38
|
Prchal-Murphy M, Witalisz-Siepracka A,
Bednarik KT, Putz EM, Gotthardt D, Meissl K, Sexl V, Müller M and
Strobl B: In vivo tumor surveillance by NK cells requires TYK2 but
not TYK2 kinase activity. Oncoimmunology. 4:e10475792015.
View Article : Google Scholar :
|