Introduction
Chondrosarcoma is a malignant bone tumor
characterized by the production of a modified cartilage-type
extracellular matrix (ECM). It is a heterogeneous, mesenchymal
origin tumor that exhibits different histopathology and clinical
behavior. Chondrosarcoma is the second most common bone tumor after
osteosarcoma (1). The primary
treatment for localized chondrosarcomas is surgical resection
(2), as these tumors of
mesenchymal origin exhibit resistance to classical chemotherapy and
radiotherapy. The possible mechanisms of resistance to chemotherapy
are a low mitotic rate and attenuated penetration into the tumor
microenvironment resulting from low vascularity and the specific
structure of the tumor-derived ECM (3). However, in some rare subtypes, such
as mesenchymal chondrosarcomas, chemotherapy may be useful. Thus, a
retrospective study, suggests that the combination of surgery with
chemotherapy results in a better outcome for undifferentiated
chondrosarcoma treatment in comparison to surgery alone (4). The generation of efficient
conjunctive therapy for chondrosarcoma is an unmet medical
need.
As cancer progresses, significant changes occur in
the structural and mechanical properties of ECM constituents
(5). The ECM provides a scaffold
on which cancer cells adhere and migrate. However, by regulating a
myriad of signaling pathways, the ECM components likewise affect
critical cellular events, such as cellular motility, adhesion,
differentiation, invasion and metastasis (6,7). The
tumor ECM is extensively remodeled by enzymatic digestion,
releasing active mediators that facilitate tumor cell growth and
spreading (8,9).
Small leucine-rich proteoglycans (SLRPs) are diverse
and multifaceted matrix constituents contributing to matrix
organization and crucial mediators of ECM-cell signal transduction
(10,11). SLRPs are composed of a core protein
undergoing post-translational modifications, including substitution
with glycosaminoglycan (GAG) side chains of various types (10,12).
The GAG chains bind covalently into the protein core through serine
or threonine residues (10). The
protein core of these proteoglycans (PGs) is in the molecular
weight range between 36 and 77 kDa, characterized by a variable
number of central leucine-rich repeat (LRR) domains; whereas, the
total PG molecular weight depends on the level of its glycosylation
(13). The SLRP family consists of
17 members distributed into 5 classes based on characteristics,
such as conserved leucine-rich repeats (LRR), N-terminal
cysteine-rich clusters and unique chromosomal organization
(14). Upon synthesis, SLRPs are
secreted into the pericellular space, where they are sequestered
through binding to cell membrane receptors or diffuse and
incorporate into the tissue ECM by tethering to collagen fibers
(15). Thus, in the cellular
milieu, the SLRPs are distributed among the pericellular matrix,
bound into 'proper' ECM, and also present as a pool of free
molecules (12). SLRPs are an
essential constituent of mesenchymal origin tissues, including bone
and cartilage, as well as intimately involved in these tissue
growth processes (16-19). Importantly, it is well determined
that an abnormal SLRP expression, as well as structure, conclude in
abnormal function of the ECMs and disease (13). Indeed, the SLRPs are implicated in
the carcinogenesis of various solid tumors (20). Furthermore, the SLRPs contribute to
cartilage pathologies (19),
including degenerative cartilage disease (21,22).
The putative participation of SLRPs in the processes
of chondrosarcoma tumorigenesis is unknown, with only a few reports
addressing the issue (23,24). In the present study, the expression
of the SLRPs members, decorin, biglycan and lumican, in an in vitro
model of chondrosarcoma was examined and the main focus was paid to
the putative effects of these mediators on chondrosarcoma cell
biological functions.
Materials and methods
Materials
Recombinant human insulin-like growth factor I
(IGF-I; 291-G1; 10 ng/ml) and transforming growth factor-β2
(TGF-β2; 302-B2-010; 10 ng/ml) were obtained from R&D Systems.
A selective inhibitor of ERK1/2 (U0126; 10 μM; Cell Signaling
Technology, Inc.) and allosteric inhibitor of IGF-IR (AG1024; 1
µM; Sigma-Aldrich; Merck KGaA) were used in the present
study for 1 h. Primary antibodies from Santa Cruz Biotechnology,
Inc. were used, and these included anti-lumican (sc166871; mouse
monoclonal; 1/100 dilution for western blot analysis or 1/50 for
immunofluorescence), anti-β-catenin (sc7963; mouse monoclonal;
1/300 dilution), anti-ERK1/2 (sc514302; mouse monoclonal; 1/200
dilution), anti-IGF-IR (sc81464; mouse monoclonal; 1/100 dilution),
anti-pERK1/2 (sc136521; mouse monoclonal; 1/100 dilution),
anti-Smad2 (sc6200; goat polyclonal; 1/200 dilution) and
anti-pSmad2 (sc101801; rabbit polyclonal; 1/200 dilution). In
addition, anti-actin (MAB1501; mouse monoclonal; 1/5,000 dilution;
EMD Millipore), anti-p-IGF-IR (PA5-37602; polyclonal rabbit; 1/500
dilution; Thermo Fisher Scientific, Inc.), keratan sulfate (KS;
270427; mouse monoclonal; 1/1,000 dilution; Seikagaku Corporation)
and keratanase II (100812; 0,005 µ/ml; Seikagaku
Corporation) were utilized. Secondary-HRP antibodies anti-rabbit
(AP182PR) and anti-mouse (AP192PM) were used at a 1/5,000 dilution
and obtained from Millipore.
Cells and cell culture
In the present study, the HTB94 (ATCC®
HTB-94™) human chondrosarcoma cell line was utilized. Cells were
grown in DMEM (Biosera AG; LM-D1111) supplemented with 10% fetal
bovine serum (FBS; Invitrogen; Thermo Fisher Scientific, Inc.;
10500-064; heat-inactivated), gentamycin (Invitrogen; Thermo Fisher
Scientific, Inc.; 15710-049) and penicillin/streptomycin
(100units/ml; Biosera LMA4118). Prior to the addition of
treatments, cells were cultured in serum-free medium for 24 h at
37°C and 5% CO2. Inhibitors, when used (ERK inhibitor or
IGF-IR inhibitor), were added 1h prior to growth factor
treatment.
Cell adherence assay
Following the treatments, the cells were harvested,
and 10,000 cells/well were seeded onto fibronectin (FN)-coated
96-well plates for 1 h. According to the manufacturer's
instructions, the number of adherent cells was measured using the
CyQUANT fluorometric assay (Molecular Probes; Thermo Fisher
Scientific, Inc.). Fluorescence was measured on a Fluorometer
(BioTek Instruments, Inc.) using the proposed excitation (485 nm)
and emission filters (528 nm), as previously described (25). A separate standard curve was used
to convert fluorescence units to cell numbers. All adhesion
experiments were repeated at least 3 times and performed in
triplicate.
Cell migration assay
HB94 cells were seeded in 24-well culture plates at
a concentration of 10×104 cells per well. The optimal
concentration for plating was selected so that the cells would be
confluent at almost 100% after 72 h of culture at 37°C and 5%
CO2. RNA interference was performed according to the
protocol described in the section below entitled 'Transfection with
siRNA' using siRNAs specific for lumican, or decorin, or biglycan.
Serum-free medium was utilized for culture. The cell layer was
scratched with a sterile 10 µl pipette tip. Detached cells
were removed by washing the cell layer twice with serum-free
medium. The wound closure was monitored at 6 and 12 h with a
digital image processor connected to a microscope (Leica DMIL), at
5 different positions across the wound. Cell motility was
quantified by ImageJ analysis (ImageJ 1.4.3.67 launcher; Symmetry
Software).
Proliferation assay
Growing cells from confluent cultures were harvested
and seeded in black 96-well plates (3603; Corning, Inc.) at a
density of 5,000 cells per well in 200 µl of DMEM (10% FBS).
The cell density number was selected from optimization experiments
(data not shown). The cells were allowed to rest overnight. If
necessary, transfection with short interfering RNAs (siRNAs) was
performed in a serum-free medium without antibiotics for 24 h. This
was then replaced with fresh medium (0% FBS) with antibiotics. All
treatments (inhibitors and/or growth factors) were added for the
next 24 h at 37°C and 5% CO2 in 0% FBS. The cells were
then lysed, and their number was calculated using the CyQUANT
fluorometric assay (C7026; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Fluorescence was
measured in a Fluorometer (BioTek Instruments, Inc.) using the
proposed excitation (485 nm) and emission filters (528 nm). A
separate standard curve was used to convert fluorescence units to
cell numbers. All experiments were performed in triplicate.
MTT assay
Growing cells from confluent cultures were harvested
and seeded in 96-well plates at a density of 12,000 cells per well
in 200 µl of DMEM (10% FBS). The cells were allowed to rest
overnight. Transfection with siRNAs was performed in a serum-free
medium without antibiotics for 24 h (siLum) or 12 h (siDec, siBig).
This medium was then replaced with fresh medium (0% FBS) and cells
were incubated at 37°C and 5% CO2 in 0% FBS for the
following 24 h. The Vybrant MTT Cell Proliferation Assay (cat. no.
11465007001; Roche Diagnostics) was performed according to the
manufacturer's instructions. In brief, the medium was replaced with
100 μl of fresh medium (0% FBS) and 10 µl of MTT stock
solution (12 Mm) were added to each well. Following 4-h incubation
at 37°C, 50 µl of DMSO was added to cells for the next 10
min. The absorbance was measured at 540 nm using a Synergy HTX
multimode microplate reader (BioTek). A separate standard curve was
used to convert absorbance to cell number. All experiments were
performed in triplicate.
Transfection with siRNA
For transfection experiments, the cells were plated
in serum- and antibiotic-free medium in either 96-well plates
(5,000 cells/well) or T25 flasks (1:8 dilution of a 90% confluent
T75 flask). siRNA specific for lumican (siLum; stealth siRNAs
HSS106200; Invitrogen; Thermo Fisher Scientific, Inc.), decorin
(siDec; (stealth siRNAs HSS102673; Invitrogen; Thermo Fisher
Scientific, Inc.) and biglycan (siBig; stealth siRNAs HSS184531;
Invitrogen; Thermo Fisher Scientific, Inc.) and RNAi negative
control (siScr; medium GC content negative control; Invitrogen;
Thermo Fisher Scientific, Inc.). For transfection, siRNA, and
Lipofectamine 2000 (11668-027; Invitrogen; Thermo Fisher
Scientific, Inc.) were diluted in Opti-MEM I Reduced Serum Medium
(31985-070; Invitrogen; Thermo Fisher Scientific, Inc.). Following
5 min of incubation at room temperature, diluted Lipofectamine 2000
was mixed with diluted siRNA (100 nm) for 20 min at room
temperature to allow siRNA-liposome complexes to form and added to
cell layers. Transfection was allowed to take place during 24 h for
siLum or 12 h for siDec and siBig, when the medium was replaced
with fresh (0% FBS) containing antibiotics and the incubation
period continued for 24 or 36 h, respectively. Cells were then
harvested, and mRNA or protein was extracted. When necessary,
treatments were performed at the 24 h point after the initial
transfection period. All transfection experiments were repeated at
least 3 times and performed in triplicate.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
According to the manufacturer's instructions, total
ribonucleic acid isolation was performed using TRIzol (15596026;
Invitrogen; Thermo Fisher Scientific, Inc.). Total RNA (1
µg) was added for cDNA synthesis using the Takara (RR037A)
RT cDNA synthesis kit. For semi-quantification of the genes of
interest, qPCR reactions were performed on a Mx300P cycler using
the Universal qPCR kit (KK4602; KAPA Biosystems) in a total volume
of 20 µl. The thermocycling conditions were as follows: 94°C
for 15 min, 40 cycles at 94°C for 20 sec, 55°C for 30 sec, 72°C for
30 sec, 72°C for 10 min. The PCR primer sequences were as follows:
GAPDH forward, 5′-GGAAGGTGAAGGTCGGAGTCA-3′ and reverse,
5′-GTCATTGATGGCAACAATATCCACT-3′; lumican forward,
5′-CTTCAATCAGATAGCCAGACTGC-3′ and reverse,
5′-AGCCAGTTCGTTGTGAGATAAAC-3′; decorin forward,
5′-TCAATGGACTGAACCAGATGA-3′ and reverse, 5′-CCTTGA
GGAATGCTGGTGAT-3′; biglycan forward, 5′-TCTGAA GTCTGTGCCCAA-3′ and
reverse, 5′-TCTGAGATGCGCAG GTA-3′; p53 forward,
5′-CGTCGTGGCTTCTTGCAT TC-3′ and reverse, 5′-AAGACCTGCCCTGTGCAGC-3′;
p21WAF1/ CIP1 forward, TGGAGACTCTCAAGGTCGAAA-3′ and reverse,
5′-AAGATCAGCCGGCGTTTG-3′. Standard curves were run in each
optimized assay, which produced a linear plot of the threshold
cycle Ct (dRn) against initial quantity (copies). The amount of
each target was semi-quantified based on the concentration of the
standard curve and was presented as arbitrary units. GAPDH was
utilized as a housekeeping gene.
Western blot analysis
The total protein secreted into the serum-free
culture medium was concentrated using Amicon Ultra 15ml (UFC901024;
10 kDa cut-off) centrifugal concentrator tubes. The initial volume
of 3ml serum-free medium collected from culture, to isolate
secreted proteins, was concentrated to a final volume of 500
µl whereas, harvested cells were lysed with RIPA solution
(50 mM Tris-HCl, 1% NP-40, 0.25% Na-Deoxycholate, 150 mM NaCl, 1 mM
EDTA with protease and phosphatase inhibitors). Equal amounts of
protein, either cell extracts or secreted, were subjected to
SDS-PAGE using 10% polyacrylamide gels under reducing conditions.
Separated protein bands were transferred to nitrocellulose
membranes in 10 mM CAPS (pH 11), containing 10% methanol. Membranes
were blocked overnight at 4°C with PBS containing 0.1% Tween-20
(PBS-Tween) and 5% (w/v) low-fat milk powder. The membranes were
incubated for 1 h at room temperature with the primary antibodies
in PBS containing 0.1% Tween-20 (PBS-Tween) and 1% (w/v) low-fat
milk powder. The immune complexes were detected following
incubation with the appropriate peroxidase-conjugated secondary
antibody diluted (1:5,000) in PBS-Tween, 2% low-fat milk for 1 h at
room temperature, using the LumiSensor Chemiluminescent HRP
substrate kit (Genscript; L00221V500), according to the
manufacturer's instructions. The protein expression of actin was
used to correct for the amount of each sample analyzed using ImageJ
Analysis Software.
Immunofluorescence
HTB94 cells were seeded on round coverslips placed
in 24-well plates, at a concentration of 70,000 cells/well, and
incubated in complete medium for 24 h. Subsequently, the cells were
incubated for 48 h at 37°C and 5% CO2 in 0% FBS medium.
The cells were fixed with 5% formaldehyde and 2% sucrose in PBS for
10 min at room temperature. Following 3 washes with PBS
supplemented with 1% FBS, the permeabilizing agent Triton X-100 was
added for 10 min and then washed prior to the addition of the
primary antibody for 1h at room temperature. Coverslips not
incubated with the primary antibody were utilized as negative
controls. The coverslips were rewashed and incubated for 1 h, in
the dark at room temperature, with anti-mouse Alexa Fluor 488
(A21206; 1/200 dilution; Molecular Probes). TO-PRO-3 iodide
(Molecular Probes; T3605) diluted 1:300 in de-ionized
H2O was applied for 40 min to stain the nuclei. The
coverslips were then placed onto slides using glycerol as a
mountant and visualized using confocal microscopy.
Statistical analysis
Statistical significance was evaluated using a
Student's t-test, or one-way ANOVA analysis of variance with
Tukey's post-test, using GraphPad Prism (version 4.0) software.
Results
Expression of decorin, biglycan and
lumican in HTB94 chondrosarcoma cells
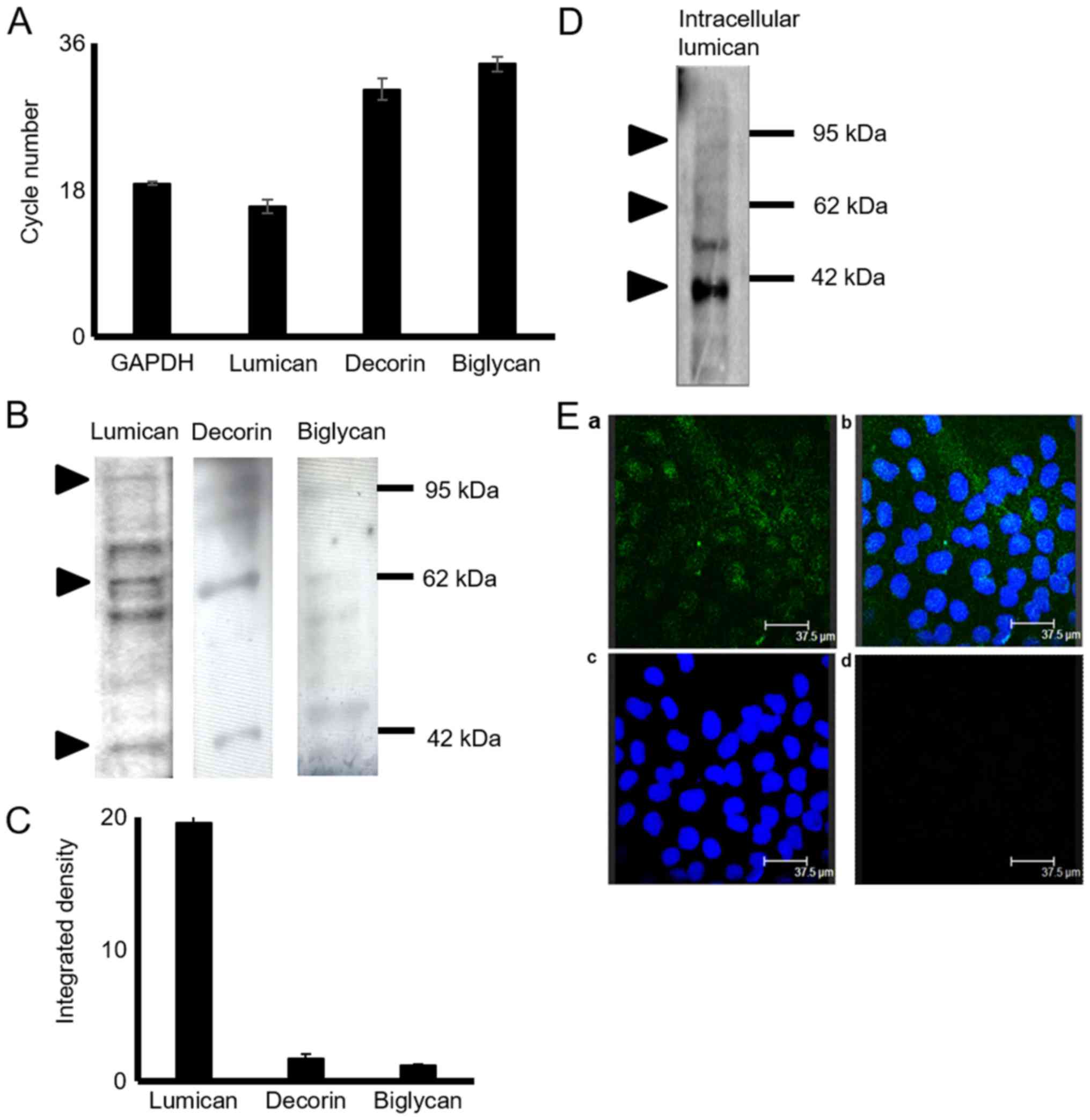
The expression of decorin, biglycan and lumican in
HTB94 chondrosarcoma cells was estimated at the mRNA and protein
level. The results of RT-qPCRdemonstrated that the HTB94
chondrosarcoma cells expressed lumican, decorin and biglycan at the
mRNA level (Fig. 1A). The lumican
transcripts were several fold higher than the low expression levels
of decorin and biglycan (Fig. 1A).
GAPDH was utilized as a housekeeping gene. To determine SLRP
expression in HTB94 cells at the protein level, total protein was
extracted from the cell culture medium, as well as from harvested
cells. Western blot analysis of the proteins secreted by the HTB94
cells using specific antibodies revealed that the 3 SLRPs were
secreted at discrete levels (Fig.
1B). The most abundant SLRP secreted by the HTB94 cells was
lumican, with decorin and biglycan exhibiting low levels of
expression, as demonstrated by densitometric analysis (Fig. 1C) and by transcript data. In all
cases, in addition to the approximately 40 kDa band representative
of the protein core, bands of higher molecular weight were present,
indicating that the 3 SLRPs are also secreted as variously
glycosylated protein products (Fig.
1B). In continuation, the present study focused on lumican, the
main member of the SLRPs, secreted by the HTB94 chondrosarcoma
cells. As shown in Fig. 1C, the
bands representing glycosylated lumican were mostly localized in
the 55 to 80 kDa molecular weight range. The monoclonal antibody
(5D4) specific for KS chains was utilized to characterize the type
of lumican substitution. To further evaluate the production of
lumican by the HTB94 cells, respective cell extracts were probed
with an anti-lumican antibody. Western blot analysis (Fig. 1D) revealed lumican specific bands
of various molecular weights indicative of different stages of
protein glycosylation. The utilization of fluorescence likewise
demonstrated the deposition, of under synthesis, lumican to the
cytoplasm of HTB94 cells (Fig.
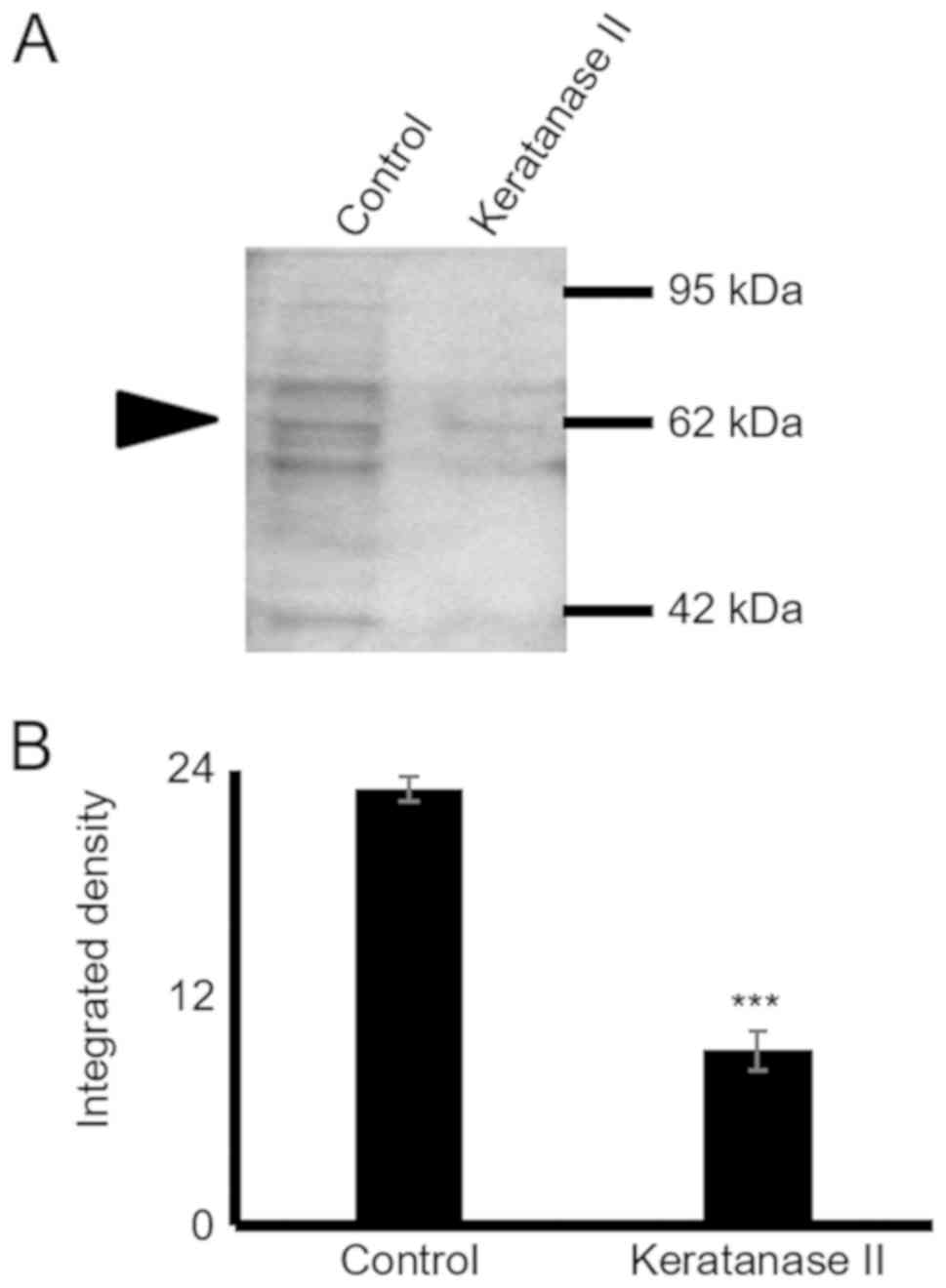
1E). As shown in Fig. 2, the
use of the 5D4 antibody gives the specific 55-80 kDa band pattern,
identical to that obtained upon probing with the anti-lumican
antibody. In continuation, secreted proteins were subjected to
enzymic treatments with keratanase II, which cleaves within
sulfated lactosamine residues, and subjected to SDS-PAGE and
western blot analysis with anti-KS antibody to confirm the nature
of the carbohydrate component. The specific KS-reaction was
markedly attenuated in the samples treated with keratanase II,
demonstrating that lumican secreted by HTB94 was partially
substituted with KS (Fig. 2).
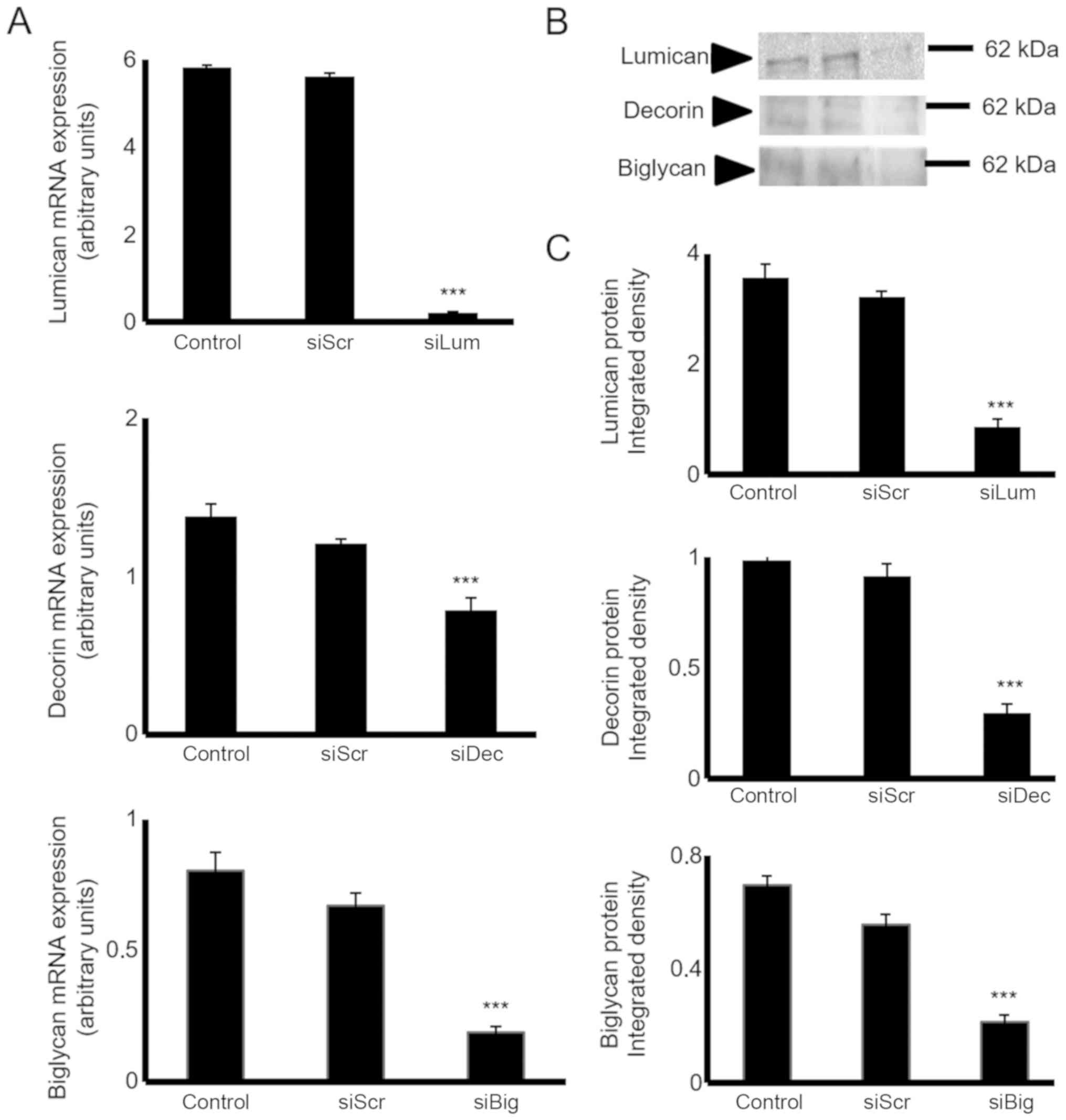
SLRP expression levels are efficiently
downregulated following transfection with specific siRNAs
The HTB94 cells were transfected with lumican-,
decorin- and biglycan-specific siRNAs to estimate their putative
biological roles. The downregulation of SLRP mRNA expression was
verified by RT-qPCR. Transfection of the HTB94 cells with siLum,
siDec, and siBig for 24h resulted in the potent inhibition of mRNA
expression (P≤0.001), respectively (Fig. 3A). The downregulation of lumican,
decorin and biglycan transcripts was followed by a significant
decrease in lumican protein secretion, as demonstrated by western
blot analysis (Fig. 3B and C).
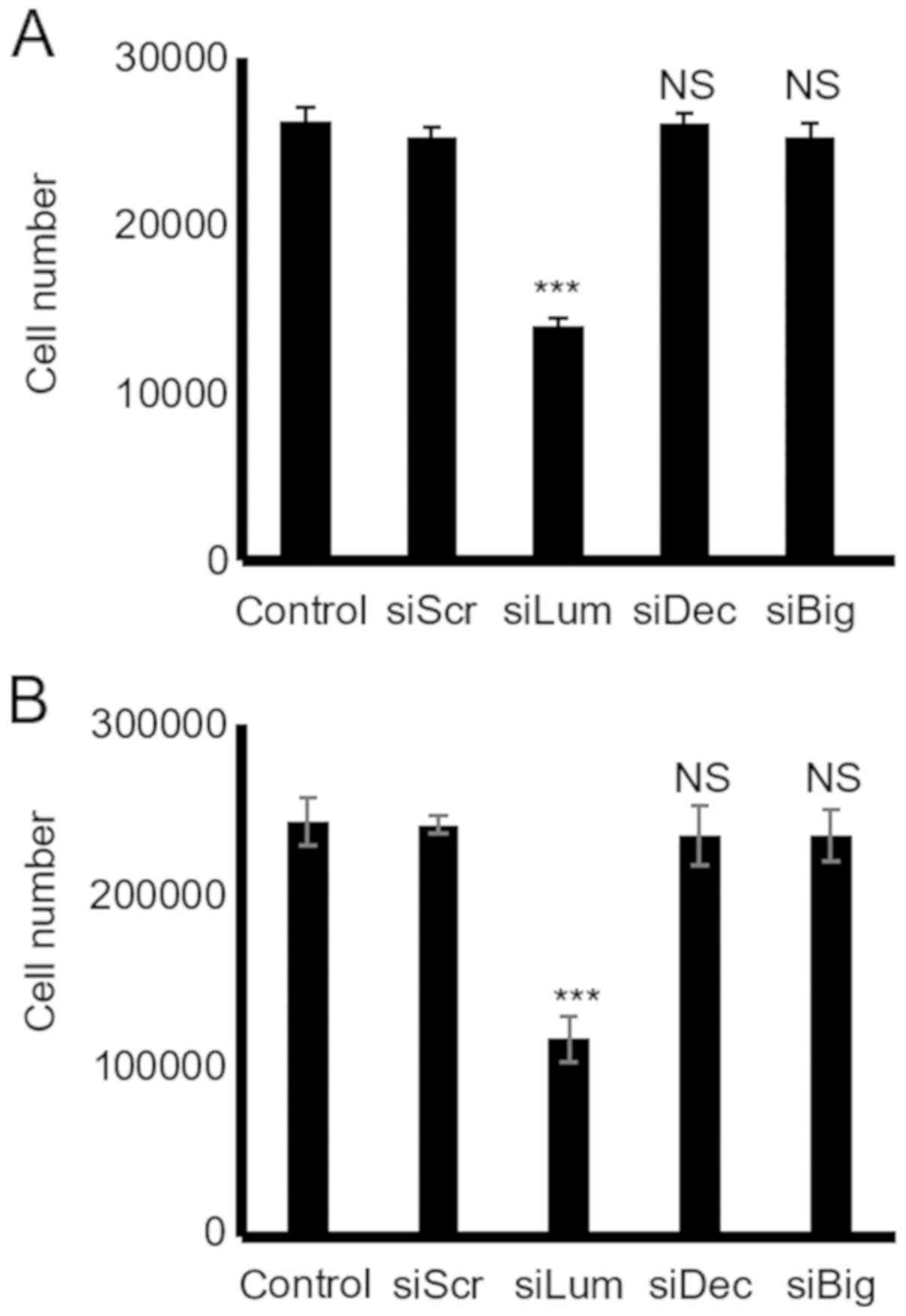
Effect of endogenous lumican, decorin and
biglycan on HTB94 chondrosarcoma cell growth
The downregulation of lumican secretion following
transfection of the HTB94 cells with lumican siRNA resulted in the
potent inhibition of growth as compared to the cells transfected
with scramble siRNA (P≤0.001; Fig.
4). However, the downregulation of the low endogenous
expression of decorin and biglycan did not affect the growth of the
HTB94 cells, as shown by the CyQUANT fluorometric and MTT assays
(P=NS) (Fig. 4).
Effect of endogenous lumican, decorin and
biglycan on HTB94 chondrosarcoma cell migration and adhesion
SLRPs have previously been shown to affect cell
motility and adhesion by modulating the cell-matrix interaction of
cells (26). The present study
thus examined the effect of endogenous decorin, biglycan and
lumican production on the motility of HTB94 cells using a 'wound
healing' assay and transfection with siRNA, as previously
demonstrated (26). In the present
study, the HTB94 cells, however, migrated with equal efficiency in
the presence or absence of decorin, biglycan and lumican siRNA,
indicating that these SLRP members do not affect their migratory
capabilities (Fig. 5A and B). In
continuation, the adhesion ability of these cells was examined
utilizing a 96-well plate adhesion assay, as previously described
(27). The results demonstrated
that transfection with decorin, biglycan and lumican siRNA did not
affect the ability of the cells to attach to the fibronectin
substrate (Fig. 5C).
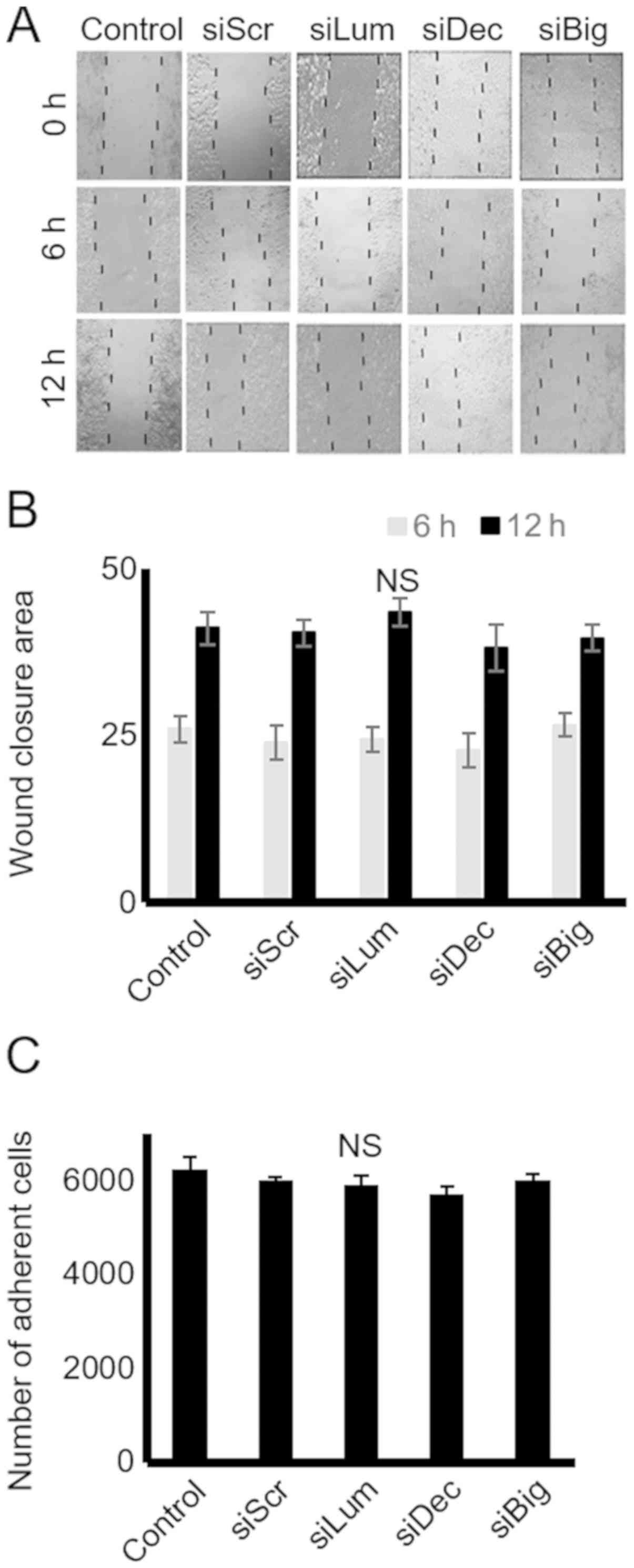 | Figure 5Effect of SLRP downregulation on
chondrosarcoma cell migration and adhesion. (A) Cells were
harvested and seeded (100,000 cells/well) in 24-well plates, to be
treated with the siRNAs specific for 3 SLRPs, lumican, decorin and
biglycan for 48 h (at which point they were confluent). Following
treatment, the cell layer was 'scratched' with a 10 µl
sterile pipette tip, and the wound surface area was measured at the
0, 6 and 12 h points. (B) The cell substratum surface area was
measured utilizing the ImageJ program and expressed as arbitrary
units. (C) HTB94 cells were treated with siRNAs specific for 3
SLRPs, lumican, decorin and biglycan for 48 h. A non-specific RNA
sequence was used as a control (siScr). Following treatment, cells
were harvested and seeded (10,000 cells/well) for 1 h at 37°C in
96-well plates coated with fibronectin. The number of attached
cells was determined using fluorometric CyQUANT Assay kit Results
represent the average of 3 separate experiments. Data are the means
± SEM. NS, not significant; SLRPs, small leucine-rich
proteoglycans. |
Mechanisms of action of lumican
TGF-β2 has previously been shown to negatively
mediate the growth of physiological chondrocytes (28,29)
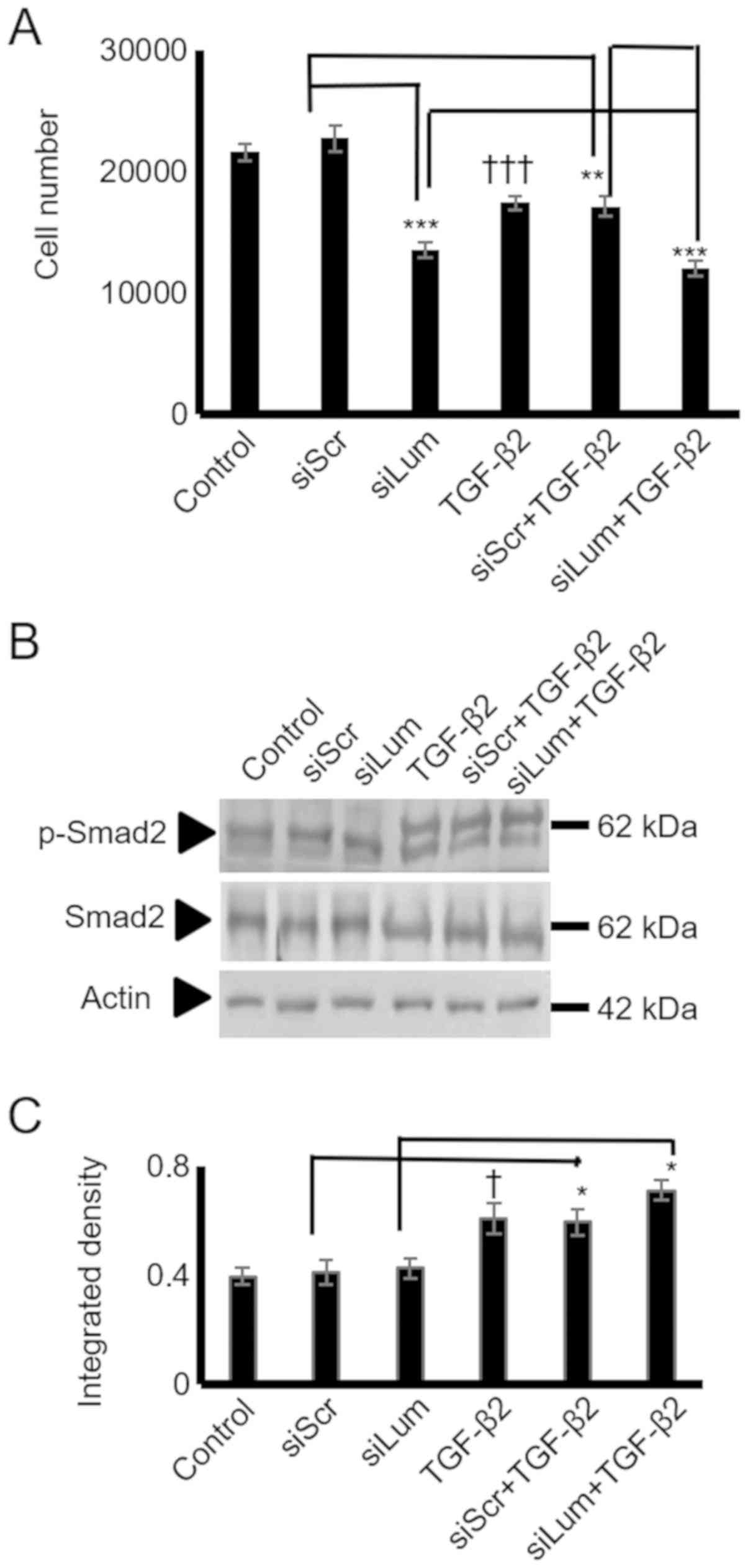
and human chondrosarcoma cells (30). In the present study, treatment of
the HTB94 cells with TGF-β2 resulted in a potent decrease in growth
(P≤0.001; Fig. 6A). As SLRP
members may affect cell proliferation by modulating TGF-β2
signaling, both lumican and scramble siRNA-transfected cells were
treated with TGF-β2, and cell proliferation was measured. The
lumican siRNA-transfected HTB94 cells treated with TGF-β2 exhibited
lower growth rates than the siScr-transfected cells treated with
TGF-β2, suggesting an additive effect of lumican on the
TGF-β2-dependent decrease in cell growth. Therefore, the difference
in growth between the lumican- and scramble siRNA-transfected cells
treated with TGF-β was attributed to the effect of lumican. These
results suggest that the regulation of HTB94 cell proliferation by
lumican is TGF-β2-independent.
To further examine the putative interaction between
lumican and TGF-β2 in HTB94 cells growth, the TGF-β2 activation of
Smad2, an established downstream mediator of TGF-β2 action
(31), was assessed. To
characterize Smad2 expression and phosphorylation levels, specific
anti-Smad2 and anti-phospho-Smad2antibodies were used. As was
expected, treatment ofthe HTB94 cells with TGF-β2 enhanced the
Smad2 phosphorylation levels (Fig. 6B
and C). In continuation, we examined the possible effects of
lumican downregulation on Smad2 activation. Western blot analysis
showed that neither Smad2 protein expression (specific 65 kDa band)
nor the expression of phospho-Smad2 (specific 60 kDa band) was
affected by lumican downregulation (Fig. 6B and C). Exogenously added TGF-β2
increased Smad 2 phosphorylation to a similar extent in both
scramble and lumican-deficient cells (Fig. 6B and C). Densitometric analysis of
the respective bands is presented in Fig. 6C. Protein amounts were normalized
against actin. These results revealed that lumican affected neither
the TGF-β2 receptor-restricted, Smad2 signaling, or consecutively
the TGF-β2-dependent growth of HTB94 cells.
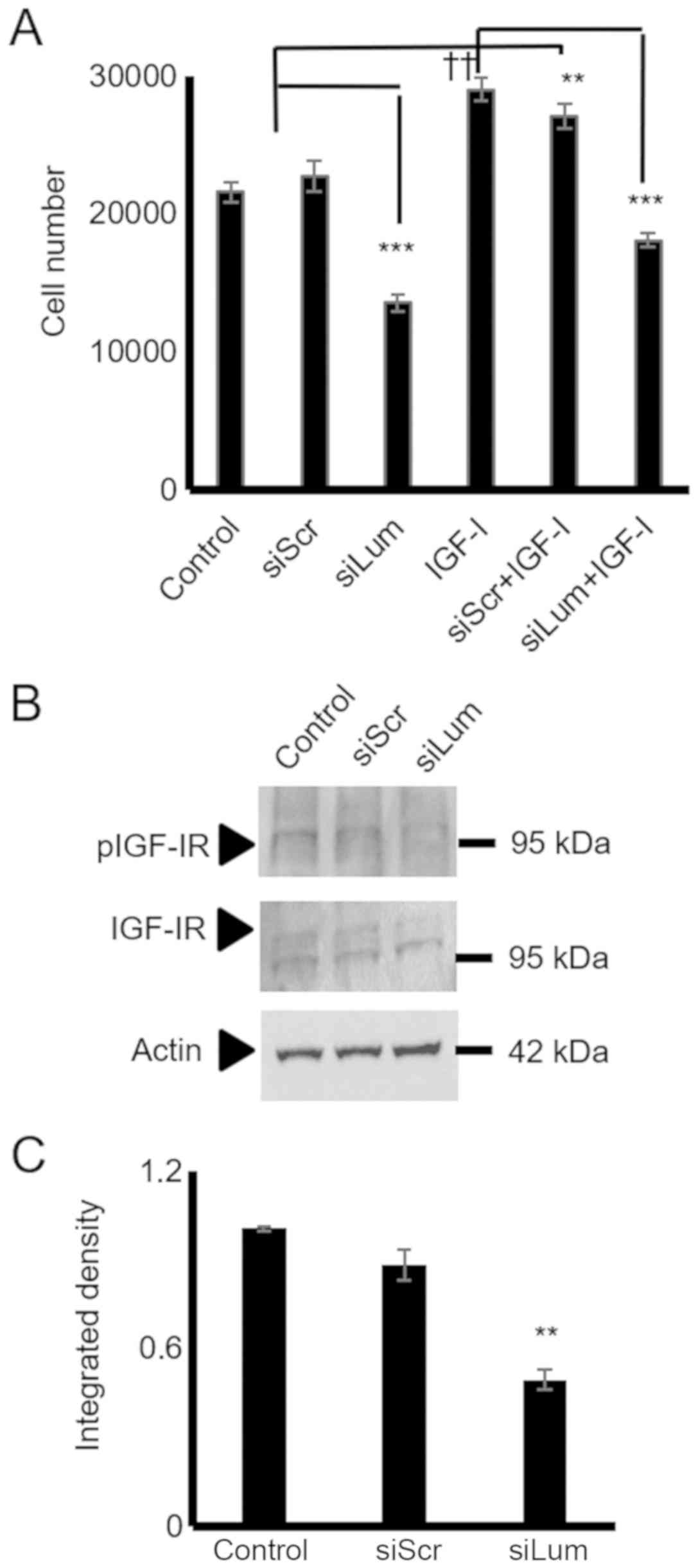
IGF-I is a well-established stimulator of
chondroblast growth (32) and a
moderate stimulator of chondrosarcoma proliferation (33). In initial experiments, it was
verified, utilizing western blot analysis, that the HTB94 cells
expressed a functional IGF-IR receptor with the ability to respond
to IGF-I stimulation (Fig. 7A and
B). Subsequently, the HTB94 cells were treated with IGF-I and
IGF-IR inhibitor or their combination. IGF-I treatment
significantly enhanced cell growth (P≤0.01), whereas the blockage
of IGF-IR induced a marked attenuation of HTB94 cell basal and
IGF-I-dependent growth (P≤0.01; Fig.
7C). When the lumican siRNA-transfected HTB94 cells were
exposed to an IGF-IR inhibitor, no further attenuation of cell
growth was detected (Fig. 7D). As
SLRP members have earlier been shown to affect IGF-IR-dependent
growth processes (12), both
lumican and scramble siRNA-transfected cells were treated with
IGF-I, and cell proliferation was measured. The lumican
siRNA-transfected HTB94 cells treated with IGF-I exhibited complete
abolishment of IGF-I-dependent growth stimulation (Fig. 8A), suggesting that the effects of
lumican are partially mediated through IGF-IR-dependent signaling.
The lumican-deficient cells were probed with IGF-IR and pIGF-IR
antibodies to verify the contribution of lumican to IGF-IR
signaling in the regulation of HTB94 cell growth. This approach
revealed that IGF-IR activation was significantly downregulated in
the lumican-deficient cells (P≤0.01; Fig. 8B and C).
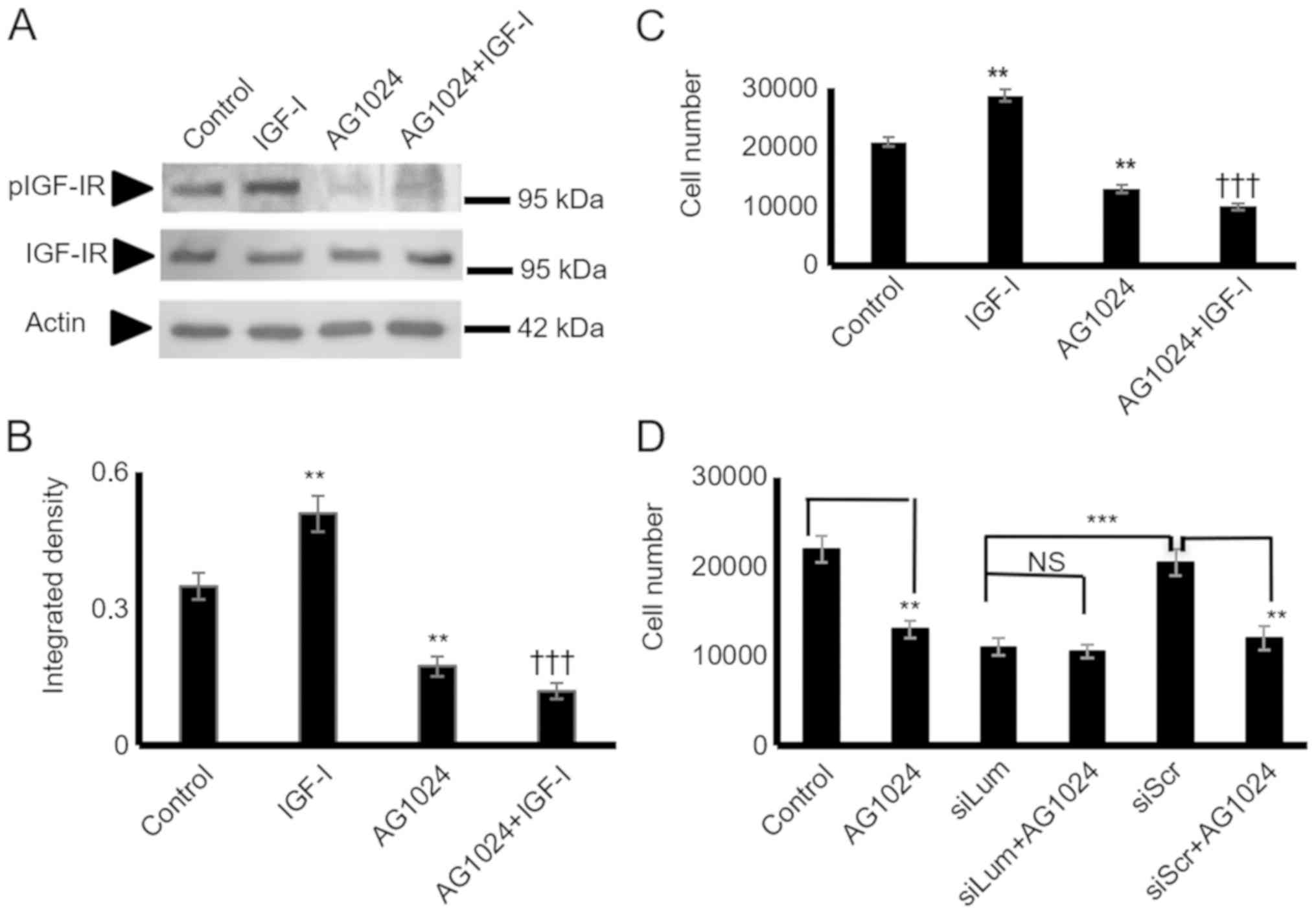 | Figure 7HTB94 chondrosarcoma cell IGF-IR
expression and effect on cell growth. Following 24 h of starvation,
the cells in each well were incubated with serum-free medium
(control), IGF-I (10 ng/ml) in 0% FBS DMEM, 1 µM AG1024
(IGF-IR inhibitor) and 10 ng/ml IGF-I + 1 µM AG1024 for 24
h. (A) Representative western blots showing the expression of
IGF-IR total protein (IGF-IR) (95 kDa) and phosphorylated IGF-IR
protein (pIGF-IR) (95 kDa). (B) Protein bands were
densitometrically analyzed and adjusted against actin (42 kDa), and
the ratio of pIGF-IR/IGF-IR was measured and presented. The
position of the nearest respective protein marker band is depicted
to the right. Results represent the average of 3 separate
experiments. Data are the means ± SEM; **P≤0.01,
statistically significant difference between control and IGF-I and
AG1024 treatments. †††P≤0.01, statistically significant
difference between IGF-I and AG1024+IGF-I treatment groups. (C)
HTB94 cells were harvested and seeded (5,000 cells/well) on 96-well
plates. Following 24 h of starvation, cells in each well were
incubated with serum-free medium (control), IGF-I (10 ng/ml) in 0%
FBS DMEM, 1 µM AG1024 (IGF-IR inhibitor) and 10 ng/ml IGF-I
+ 1 µM AG1024 for 24 h. Cell number was determined using a
fluorometric CyQUANT Assay kit. Results represent the average of 3
separate experiments. Data are the means ± SEM;
**P≤0.01, statistically significant difference between
control and IGF-I, AG1024 treatments. †††P≤0.01,
statistically significant difference between IGF-I and AG1024+IGF-I
treatment groups. (D) HTB94 cells were treated for 48 h with siRNA
specific for lumican. A non-specific RNA sequence was used as a
control (siScr). Following 24 h of transfection with siLum, cells
were treated with 1 µM AG1024 for 24 h in 0% FBS DMEM. Cell
number was determined using a fluorometric CyQUANT Assay kit.
Results represent the average of 3 separate experiments. Data are
the means ± SEM; **P≤0.01, statistically significant
difference between siScr and AG1024 treatments.
***P≤0.001, statistically significant difference between
siLum and siScr; NS, no significance between the siLum and siLum +
AG1024 + treatment groups. |
Erk1/2 is a downstream mediator of
lumican/IGF-IR growth regulation
Subsequently, the present study examined critical
IGF-I downstream mediators focusing on ERK1/2, a well-established
IGF-I downstream conduit in the regulation of tumor cell growth,
migration, and adhesion (27,34).
The utilization of a specific ERK1/2 inhibitor led to the
significant suppression of the HTB94 basal level of cell
proliferation (P<0.001), and the complete abolishment of
IGF-I-dependent growth (P<0.01; Fig. 9A). To determine the effects of
lumican on IGF-I-dependent ERK1/2 activation in HTB94 cells, the
ERK1/2 phosphorylation levels in both the control and IGFI-treated
lumican-deficient cellswere examined. As shown in Fig. 9B and C, lumican participation was
found to be crucial for ERK1/2 phosphorylation, as the
lumican-deficient cells exhibited an attenuated ERK1/2 activation
(P<0.05). Furthermore, when the lumican-deficient cells were
treated with the ERK1/2 inhibitor, no further downregulation of
cell growth was evident, demonstrating that ERK1/2 participation is
necessary for the lumican effect (Fig.
9D). Upon activation of the upstream effectors the degradation
of β-catenin was inhibited, resulting in its increased expression
in the cytoplasm and enhanced translocation to the nucleus. No
changes in the total β-catenin expression in the lumican-deficient
cells were detected by western blot analysis and immunofluorescence
(data not shown). On the other hand, β-catenin signaling did not
participate in the lumican/IGF-IR-mediated growth effects (Fig. S1).
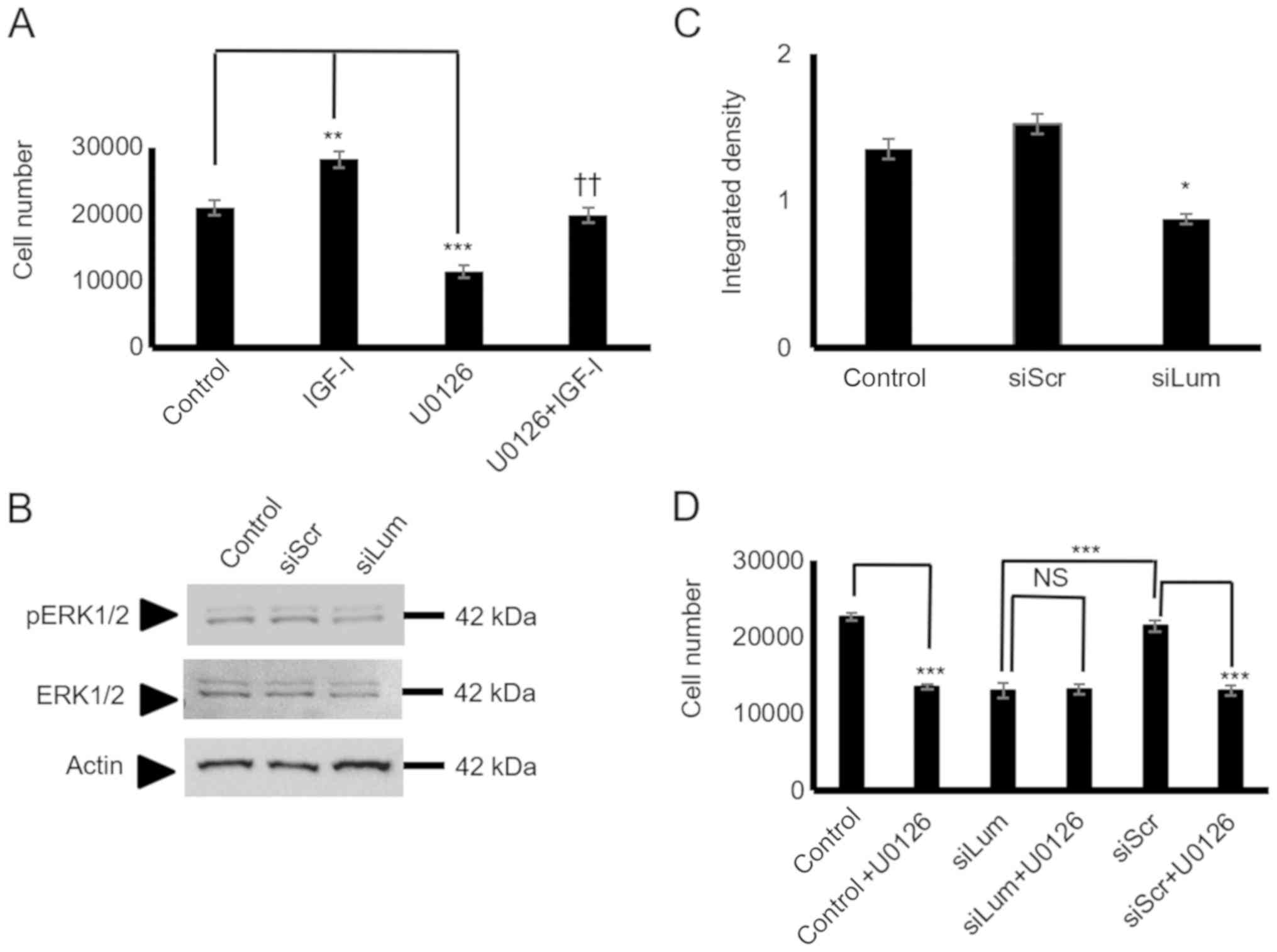 | Figure 9Effect of lumican on ERK1/2
activation. (A) HTB94 cells were harvested and seeded (5,000
cells/well) in 96-well plates. Cells in each well were incubated
with serum-free medium (control), IGF-I (10 ng/ml) in 0% FBS DMEM,
10 µM U0126 (ERK1/2 inhibitor) and 10 µM U0126 + 10
ng/ml IGF-I for 24 h. Cell number was determined using a
fluorometric CyQUANT Assay kit. (B) Expression of ERK 1/2 total
protein (ERK1/2) (42-44 kDa) and phos-phorylated ERK1/2 protein
(pERK1/2) (42-44 kDa) of cells transfected for 48 h with specific
siRNA against lumican (siLum) were determined by western blot
analysis. A non-specific RNA sequence was used as a control
(siScr). (C) ERK1/2 and pERK1/2 protein bands were
densitometrically analyzed and adjusted against actin, and the
ratio of pERK1/2/ERK1/1 was measured and presented. The position of
the nearest respective protein marker band is depicted to the
right. Representative plots are presented. Results represent the
average of 3 separate experiments. Data are the means ± SEM;
*P≤0.05, **P≤0.01 and ***P≤0.01,
statistically significant difference between control and IGF-I and
U0126 treatments. ††P≤0.01, statistically significant
difference between IGF-I and U0126 + IGF-I treatment groups. (D)
HTB94 cells were treated for 48 h with siRNA specific for lumican.
A non-specific RNA sequence was used as a control (siScr).
Following 24 h of transfection with siLum, cells were treated with
10 µM UO126 for 24 h in 0% FBS DMEM. Cell number was
determined using a fluorometric CyQUANT Assay kit. (D) Results
represent the average of 3 separate experiments. Data are the means
± SEM; ***P≤0.01, statistically significant difference
between siScr and UO126 treatments or between siLum and siScr; NS,
no significance between the siLum and siLum + U0126 treatment
groups. |
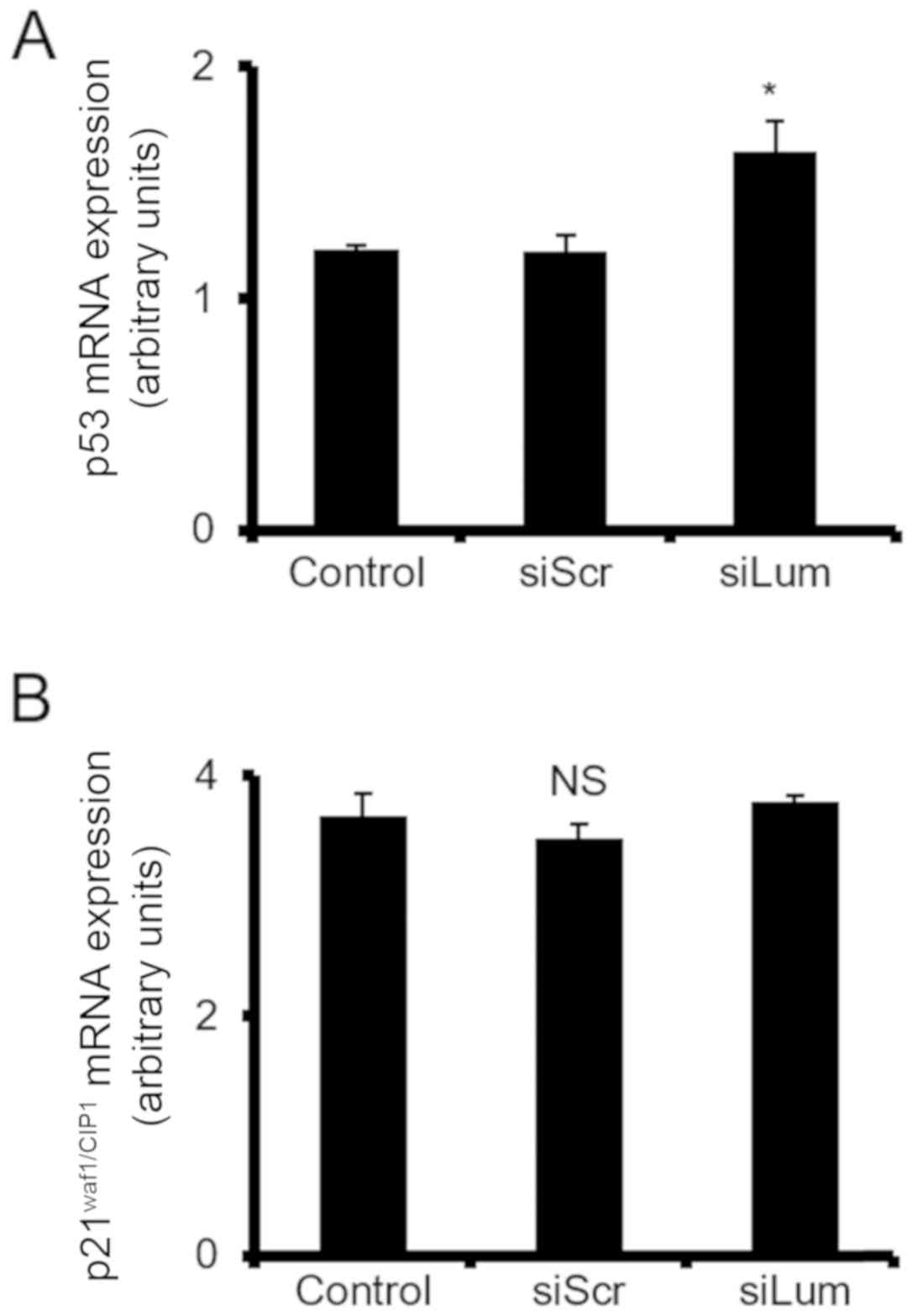
Lumican affects p53 cell cycle regulator
expression
The expression of 3 cell cycle-related genes was
then analyzed to characterize the intracellular molecular
mechanisms involving the Erk1/2 pathway on lumican-induced
chondrosarcoma cell growth. The mRNA levels of p21WAF1/CIP1 and p53
were estimated in lumican-deficient cells to investigate the
possible effects of lumican. RT-qPCR demonstrated that the levels
of p53 tumor suppressor were significantly upregulated in the
siLum-treated cells as compared to the siScr cells (P≤0.05). On the
other hand, no change was demonstrated in the expression of
p21WAF1/CIP1. These results collectively suggest that lumican is
involved in transcriptional control of cell cycle-related genes
(Fig. 10).
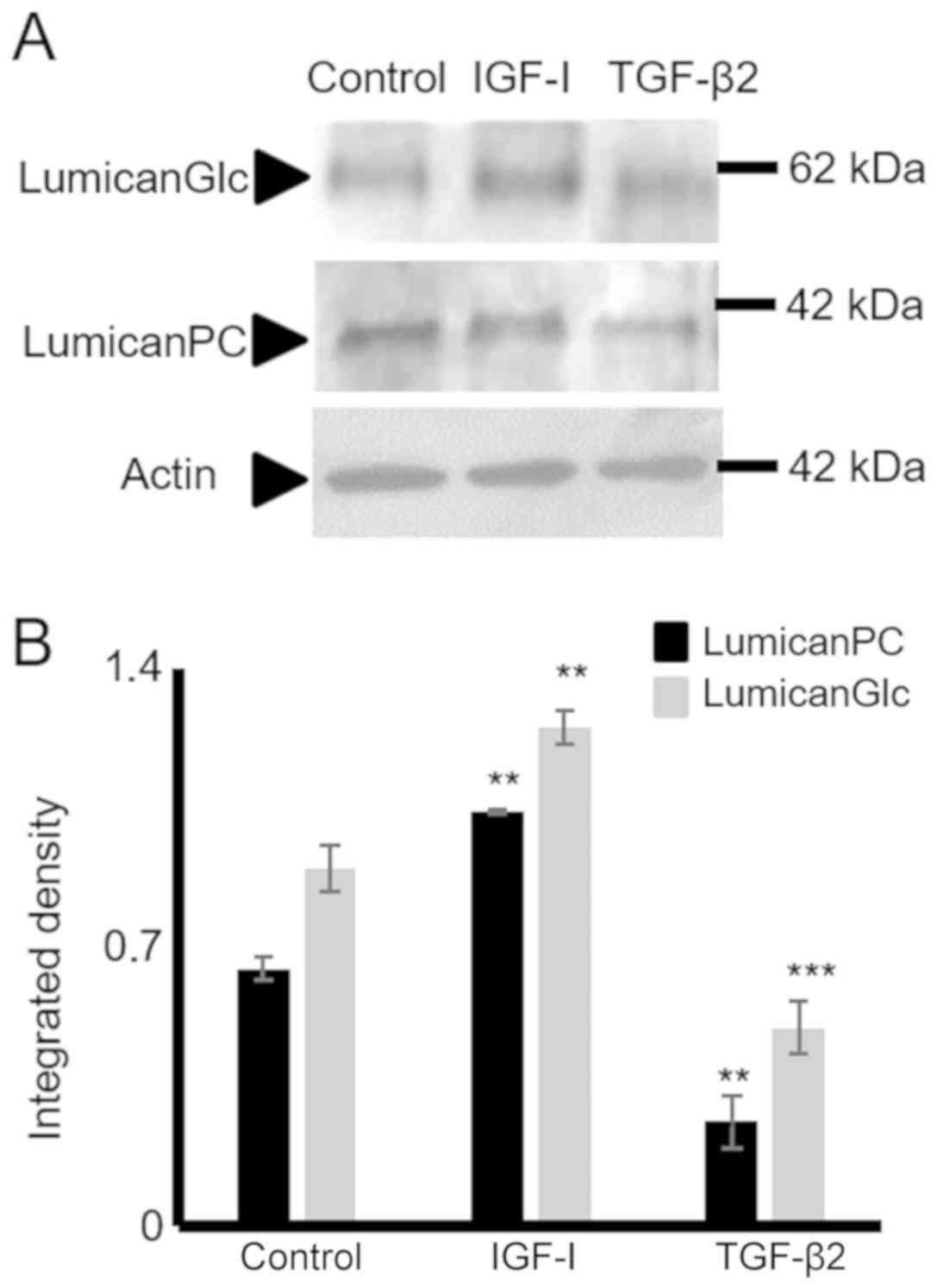
TGF-β2 and IGF-I mediate lumican
expression in HTB94 chondrosarcoma cells
In separate experiments, the HTB94 cells were
treated with TGF-β2 (10 ng/ml) and IGF-I (10 ng/ml), and the
expression of lumican in HTB94 cells was estimated. The data
obtained by western blot analysis revealed that TGF-β2 decreased
(P≤0.001) and IGF-I enhanced (P≤0.01) lumican protein expression
(Fig. 11).
Discussion
Alterations in the expression of ECM molecules
affect tumor cell functions, but also modify the behavior of
stromal cells, exerting tumor-dependent pro-angiogenesis and
pro-inflammation effects thus, facilitating the creation of a
tumor-derived microenvironment (8). Notably, chondrosarcoma cells exhibit
an extensive remodeling of ECM constituents, including hyalectan
PGs, collagen fibers, fibronectin and laminin (16). In the present study, the HTB94
human chondrosarcoma cells with a high metastatic capacity were
analyzed for decorin, biglycan and lumican expression and possible
regulation of cellular function(s) by the respective ECM
constituents. The present study demonstrated, as a novel finding,
that the chon-drosarcoma cells synthesize and efficiently secrete
the class II SLRP, lumican. This SLRP was found to be the most
abundantly expressed SLRP, as compared to the low expression levels
of decorin and biglycan, in the HTB94 chondrosarcoma model. Both
the lumican protein core and its glycosylated forms were found to
be secreted. Furthermore, the utilization of a specific anti-KS
antibody and digestion with keratanase demonstrated that the
secretion oflumican by chondrosarcoma cells was partially
glycosylated with KS chains. Indeed, this finding is similar to the
glycosylation pattern of lumican secreted by osteosarcoma cells
(35).
The putative contribution of lumican has been
examined in the growth and metastasis of several types of cancer
(6,20,36-38),
and tumor-enhancing, as well as tumor-inhibitory functions have
been indicated and are dependent on tumor type, and abundance and
the stage of the disease (20,39).
Lumican-contingent effects have been found to be
partly dependent on the type and extent of lumican glycosylation
(6). In the present study, siRNA
was utilized to examine the putative role of lumican in
chondrosarcoma growth, migration and adhesion. This approach
resulted in the efficient downregulation of lumican secretion. The
attenuated lumican secretion was associated with the decreased
growth of HTB94 cells, whereas their migratory and adhesion ability
were not affected. Previous studies have demonstrated that lumican
enhances the growth potential of lung cancer (40), bladder cancer (37) and gastric cancer (41), whereas it was found to have
inhibitive effects on breast cancer (42) and melanoma growth (43). In continuation, the present study
examined the possible mechanisms of lumican action in the
regulation of chondrosarcoma growth. The regulation of the TGF-β2
signaling pathway is one previously established mechanism of action
of lumican (6,25). In the present study, and in
agreement with previous findings, exogenous TGF-β2 was found to
decrease chondrosarcoma cell growth (44). Treatment of lumican-deficient cells
with TGF-β2 increased HTB94 cell growth inhibition. The pathway
restricted Smad2 activation (45)
was assessed in lumican and scramble siRNA-transfected cells to
examine the potential contribution of lumican to the TGF-B2
signaling pathway. Neither the basal nor the TGF-β2-dependent
levels of Smad2 phosphorylation were affected, as exogenously added
TGF-β2 enhanced Smad2 phosphorylation to a similar extent in both
scramble control-transfected and lumican-deficient cells. These
results demonstrate that the effect of lumican on HTB94 cell growth
was independent of TGF signaling.
Decorin and biglycan have previously been implicated
in the regulation of IGF/IGF-IR signaling (46), which is a crucial player in the
regulation of both physiological growth (47,48)
and malignant transformation (46). Recently, a 'Phase I trial of the
IGF-1R antibody ganitumab (AMG 479) in combination with everolimus
(RAD001) and panitumumab in patients with advanced cancer'
exhibited benefits in the case of chondrosar-coma patients
(49). On the other hand, in a
separate study, 'no preclinical rationale for IGF1R directed
therapy in chondro-sarcoma of bone' was indicated (50).
Importantly, decorin has been shown to bind IGF-IR
and attenuate its signaling (51),
whereas we have recently demonstrated that biglycan modulates the
growth of MG63 osteosarcoma cells through an LPR6/β-catenin/IGFR-IR
signaling axis (52). Upon
activation of the upstream effectors, the degradation of β-catenin
is inhibited, resulting in the increased expression in the
cytoplasm and the enhanced trans-location to the nucleus (52). No changes in the total β-catenin
expression in lumican-deficient HTB94 cells were detected in the
present study, suggesting that the effects of lumican did not
involve β-catenin signaling. The present study demonstrated that
the IGF-I/IGF-IR axis positively modulated HTB94 cell growth,
similar to previous findings (53). Notably, lumican deficiency induced
an abolishment of IGF-I-dependent HTB94 cell growth. It should be
noted that the lumican-deficient cells exhibited an attenuation of
IGF-IR basal level activation, whereas no effect on the expression
levels of the receptor was evident. Addressing the putative
contribution of β-catenin signaling on lumican/IGF-IR-mediated
growth effects revealed that β-catenin was not a downstream
regulator. ERK1/2 is an established downstream effector of IGF-IR
signaling (54). Indeed, the
IGF-IR/ERK1/2 signaling axis has been implicated in the propagation
of fibrosarcoma migration (34)
and breast cancer adhesion (27),
as well as adrenocortical cancer cell proliferation (55). In the present study, ERK1/2 was
verified as an IGF-IR downstream mediator, as well as a necessary
component in IGF-IR-dependent facilitation of HTB94 cell growth.
The downregulation of lumican induced a substantial attenuation of
ERK1/2 activation, indicating that ERK1/2 is a necessary component
of lumican/IGF-IR-mediated HTB94 cell proliferation.
Lumican-deficient cells did not exhibit the further downregulation
of cell growth when treated with the ERK1/2 inhibitor, indicating
that ERK1/2 participation was necessary for the lumican effect.
Previously it has been suggested that ERK1/2 exerts pro-oncogenic
effects in chon-drosarcoma cells (56,57).
Moreover, ERK1/2, along with PI3K and p38 signaling
pathways, has been shown to promote chondrosarcoma cell motility,
invasion and lung colonization (58). The ERK1/2 pathway affects cell
cycle progression and apoptosis through the modulation of specific
cell cycle regulators expression, including p53 and p21 in OVCAR-3
human ovarian and MCF-7 breast cancer cells (59,60).
In the present study, p53 expression was elevated in the HTB94
cells, in a manner dependent on active ERK1/2 signaling. The fold
change in p53 expression was not marked, yet it was significant,
and it is suggested that p53 is one of the downstream targets of
the determined signaling pathway. Importantly, p53 has been
implicated in the progression of chondrosarcoma to a higher grade
(61) and was suggested, among
others, as a novel therapeutic target (62).
At this point, the mechanisms through which lumican
enhances IGF-IR activation remain obscure. Taking into account that
lumican is a secreted protein, it is likely to engage
extracellularly either with IGF-IR or other components of the IGF
system (63). Previously, another
SLRP member, decorin, was shown to bind and to either enhance
(64) or attenuate (46) IGF-IR activation in different
models. The results of the present study demonstrate for the first
time, to the best of our knowledge, that lumican can affect IGF-IR
activation. Of note, IGF-I was shown to enhance lumican secretion,
suggesting a feedback loop supporting chondrosarcoma growth.
Therefore, further studies on the mechanisms through which lumican
enhances IGF-IR activation are required. Furthermore, the
utilization of a single cell line is a limitation of the present
study, justified by the scarcity of data on chondrosarcomas. Thus,
an assessment of the proposed mechanism in other model systems,
including extrapolation to in vivo experimentation, will be the
focus of a future study.
In the present study, the class I SLRPs decorin and
biglycan were expressed at low levels at both the mRNA and protein
level. In two previously available studies on decorin and biglycan
expression in chondrosarcoma, decorin mRNA was isolated in some
chondrosarcoma samples (23),
whereas no decorin protein and low biglycan protein was detected in
a human chondrosarcoma cell line (24). In the present study, the low levels
of HTB94 cell decorin and biglycan expression were inadequate to
affect cell growth, migration and adhesion. To the archetypal SLRP
member decorin, a tumor-suppressive effect has been designated, and
this SLRP is not expressed by the majority of tumors, which is in
accordance with the findings of the present study (65,66).
The role of biglycan in carcinogenesis is not clear,
with the majority of studies suggesting that biglycan
overexpression in the tumor microenvironment facilitates cancer
cell growth, migration and angiogenesis by regulating downstream
intracellular signaling and biglycan-dependent modifications of the
ECM milieu (50,67-70).
In conclusion, in the present study, human
chondrosarcoma cells were shown to express the SLRPs lumican,
decorin and biglycan. Lumican was identified as the major secreted
SLRP, whereas decorin and biglycan exhibited low levels of
expression. The secreted lumican was shown to be partly substituted
with KS glycosaminoglycans. Importantly, HTB94 cell growth was
enhanced, whereas migration and adhesion were not affected by
lumican. Basal IGF-IR and IGF-IR-dependent cell growth and
phosphorylation levels were positively associated with lumican
expression, suggesting that this SLRP may affect the activation of
IGF-IR. The downregulation of lumican induced a substantial
attenuation of the IGF-IR downstream mediator, ERK1/2, activation,
indicating that ERK1/2 is a necessary component of
lumican/IGF-IR-mediated HTB94 cell growth. Moreover,
lumican-deficient cells specifically exhibited increased mRNA
levels of p53, suggesting that lumican facilitated HTB94 cell
growth through an IGF-IR/ERK1/2/p53 signaling cascade. Further
studies on the mechanistic aspects of lumican/ IGF-IR interactions
in chondrosarcoma and the association between lumican expression
and chondrosarcoma progression are essential.
Supplementary Data
Abbreviations:
|
SLRPs
|
small leucine-rich proteoglycans
|
|
ECM
|
extracellular matrix
|
|
IGF-I
|
insulin-like growth factor I
|
|
IGF-IR
|
insulin-like growth factor I
receptor
|
|
ERK1/2
|
extracellular regulated kinase 1/2
|
|
TGF-β2
|
transforming growth factor-β2
|
|
siRNA
|
short interfering RNA
|
|
PG
|
proteoglycan
|
|
KS
|
keratan sulfate
|
Acknowledgments
Not applicable.
Funding
The present study was supported by a Special Fund
for Research Grants (ELKE) with grant no. KA: 10028 of the
University of Crete to DN.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
AP and DN designed the outline of the study. AP,
EMG, AB and IS conducted the experiments, and contributed to data
interpretation and manuscript preparation. AP wrote the manuscript.
DAS, AT and GNT contributed to data interpretation, and manuscript
preparation and revision. DN supervised the study, contributed to
data interpretation and manuscript preparation/revision. All
authors have read and approved the final version of this
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
DAS is the Editor-in-Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
The other authors declare that they have no competing
interests.
References
|
1
|
International Agency for Research on
Cancer (IARC): WHO Classification of Tumours of Soft Tissue and
Bone. Fletcher CDM, Bridge JA, Hogendoorn PCW and Mertens F: 5. 4th
edition. IARC; Lyon: 2013
|
|
2
|
Heck RK Jr, Peabody TD and Simon MA:
Staging of primary malignancies of bone. CA Cancer J Clin.
56:366–375. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chow WA: Update on chondrosarcomas. Curr
Opin Oncol. 19:371–376. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tsuda Y, Ogura K, Hakozaki M, Kikuta K, Ae
K, Tsuchiya H, Iwata S, Ueda T, Kawano H and Kawai A: Mesenchymal
chon-drosarcoma: A Japanese Musculoskeletal Oncology Group (JMOG)
study on 57 patients. J Surg Oncol. 115:760–767. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Theocharis AD, Manou D and Karamanos NK:
The extracellular matrix as a multitasking player in disease. FEBS
J. 286:2830–2869. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nikitovic D, Papoutsidakis A, Karamanos NK
and Tzanakakis GN: Lumican affects tumor cell functions, tumor-ECM
interactions, angiogenesis and inflammatory response. Matrix Biol.
35:206–214. 2014. View Article : Google Scholar
|
|
7
|
Varol C and Sagi I:
Phagocyte-extracellular matrix crosstalk empowers tumor development
and dissemination. FEBS J. 285:734–751. 2018. View Article : Google Scholar
|
|
8
|
Yuzhalin AE, Lim SY, Kutikhin AG and
Gordon-Weeks AN: Dynamic matrisome: ECM remodeling factors
licensing cancer progression and metastasis. Biochim Biophys Acta
Rev Cancer. 1870:207–228. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: Regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Neill T, Schaefer L and Iozzo RV: Decoding
the Matrix: Instructive Roles of Proteoglycan Receptors.
Biochemistry. 54:4583–4598. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Karamanou K, Perrot G, Maquart FX and
Brézillon S: Lumican as a multivalent effector in wound healing.
Adv Drug Deliv Rev. 129:344–351. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nikitovic D, Aggelidakis J, Young MF,
Iozzo RV, Karamanos NK and Tzanakakis GN: The biology of small
leucine-rich proteoglycans in bone pathophysiology. J Biol Chem.
287:33926–33933. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen S and Birk DE: The regulatory roles
of small leucine-rich proteoglycans in extracellular matrix
assembly. FEBS J. 280:2120–2137. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schaefer L and Iozzo RV: Biological
functions of the small leucine-rich proteoglycans: From genetics to
signal transduction. J Biol Chem. 283:21305–21309. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Merline R, Schaefer RM and Schaefer L: The
matricellular functions of small leucine-rich proteoglycans
(SLRPs). J Cell Commun Signal. 3:323–335. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Grogan SP, Chen X, Sovani S, Taniguchi N,
Colwell CW Jr, Lotz MK and D'Lima DD: Influence of cartilage
extracellular matrix molecules on cell phenotype and neocartilage
formation. Tissue Eng Part A. 20:264–274. 2014. View Article : Google Scholar :
|
|
17
|
Lewis JL, Krawczak DA, Oegema TR Jr and
Westendorf JJ: Effect of decorin and dermatan sulfate on the
mechanical properties of a neocartilage. Connect Tissue Res.
51:159–170. 2010. View Article : Google Scholar
|
|
18
|
Burdan F, Szumiło J, Korobowicz A,
Farooquee R, Patel S, Patel A, Dave A, Szumiło M, Solecki M,
Klepacz R, et al: Morphology and physiology of the epiphyseal
growth plate. Folia Histochem Cytobiol. 47:5–16. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Melrose J, Shu C, Whitelock JM and Lord
MS: The cartilage extracellular matrix as a transient developmental
scaffold for growth plate maturation. Matrix Biol. 52-54:363–383.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Appunni S, Anand V, Khandelwal M, Gupta N,
Rubens M and Sharma A: Small Leucine Rich Proteoglycans (decorin,
biglycan and lumican) in cancer. Clin Chim Acta. 491:1–7. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu L, Li Z, Liu SY, Xu SY and Ni GX:
Asporin and osteoarthritis. Osteoarthritis Cartilage. 23:933–939.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ni GX, Li Z and Zhou YZ: The role of small
leucine-rich proteoglycans in osteoarthritis pathogenesis.
Osteoarthritis Cartilage. 22:896–903. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Söderström M, Böhling T, Ekfors T,
Nelimarkka L, Aro HT and Vuorio E: Molecular profiling of human
chondrosarcomas for matrix production and cancer markers. Int J
Cancer. 100:144–151. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chansky H, Robbins JR, Cha S, Raskind WH,
Conrad EU and Sandell LJ: Expression of cartilage extracellular
matrix and potential regulatory genes in a new human chondrosarcoma
cell line. J Orthop Res. 16:521–530. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nikitovic D, Chalkiadaki G, Berdiaki A,
Aggelidakis J, Katonis P, Karamanos NK and Tzanakakis GN: Lumican
regulates osteosarcoma cell adhesion by modulating TGFβ2 activity.
Int J Biochem Cell Biol. 43:928–935. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Berdiaki A, Datsis G, Nikitovic D,
Tsatsakis A, Katonis P, Karamanos N and Tzanakakis G: Parathyroid
hormone (PTH) peptides through the regulation of hyaluronan
metabolism affect osteosarcoma cell migration. IUBMB Life.
62:377–386. 2010.PubMed/NCBI
|
|
27
|
Voudouri K, Nikitovic D, Berdiaki A,
Kletsas D, Karamanos NK and Tzanakakis GN: IGF-I/EGF and E2
signaling crosstalk through IGF-IR conduit point affects breast
cancer cell adhesion. Matrix Biol. 56:95–113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Im GI, Jung NH and Tae SK: Chondrogenic
differentiation of mesenchymal stem cells isolated from patients in
late adulthood: The optimal conditions of growth factors. Tissue
Eng. 12:527–536. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Khaghani SAB, Akbarova G, Soon CF and
Dilbazi G: Effect of transforming growth factor-β2 on biological
regulation of multilayer primary chondrocyte culture. Cell Tissue
Bank. 19:763–775. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Boumédiene K, Takigawa M and Pujol JP:
Cell density-dependent proliferative effects of transforming growth
factor (TGF)-β 1, β 2, and β 3 in human chondrosarcoma cells
HCS-2/8 are associated with changes in the expression of TGF-β
receptor type I. Cancer Invest. 19:475–486. 2001. View Article : Google Scholar
|
|
31
|
Boeuf S, Bovée JV, Lehner B, van den Akker
B, van Ruler M, Cleton-Jansen AM and Richter W: BMP and TGFbeta
pathways in human central chondrosarcoma: Enhanced endoglin and
Smad 1 signaling in high grade tumors. BMC Cancer. 12:4882012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
De Luca F: Regulatory Role for Growth
Hormone in Statural Growth: IGF-Dependent and IGF-Independent
Effects on Growth Plate Chondrogenesis and Longitudinal Bone
Growth. Pediatr Endocrinol Rev. 16(Suppl 1): 33–38. 2018.PubMed/NCBI
|
|
33
|
Hiraoka K, Zenmyo M, Komiya S, Kawabata R,
Yokouchi M, Suzuki R, Hamada T, Kato S and Nagata K: Relationship
of p21 (waf1/cip1) and differentiation in chondrosarcoma cells.
Virchows Arch. 440:285–290. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mytilinaiou M, Nikitovic D, Berdiaki A,
Papoutsidakis A, Papachristou DJ, Tsatsakis A and Tzanakakis GN:
IGF-I regulates HT1080 fibrosarcoma cell migration through a
syndecan-2/Erk/ezrin signaling axis. Exp Cell Res. 361:9–18. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nikitovic D, Berdiaki A, Zafiropoulos A,
Katonis P, Tsatsakis A, Karamanos NK and Tzanakakis GN: Lumican
expression is positively correlated with the differentiation and
negatively with the growth of human osteosarcoma cells. FEBS J.
275:350–361. 2008. View Article : Google Scholar
|
|
36
|
Coulson-Thomas VJ, Coulson-Thomas YM,
Gesteira TF, Andrade de Paula CA, Carneiro CR, Ortiz V, Toma L, Kao
WW and Nader HB: Lumican expression, localization and antitumor
activity in prostate cancer. Exp Cell Res. 319:967–981. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mao W, Luo M, Huang X, Wang Q, Fan J, Gao
L, Zhang Y and Geng J: Knockdown of lumican inhibits proliferation
and migration of bladder cancer. Transl Oncol. 12:1072–1078. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen L, Zhang Y, Zuo Y, Ma F and Song H:
Lumican expression in gastric cancer and its association with
biological behavior and prognosis. Oncol Lett. 14:5235–5240.
2017.PubMed/NCBI
|
|
39
|
Hsiao KC, Chu PY, Chang GC and Liu KJ:
Elevated Expression of Lumican in Lung Cancer Cells Promotes Bone
Metastasis through an Autocrine Regulatory Mechanism. Cancers
(Basel). 12:2332020. View Article : Google Scholar
|
|
40
|
Yang CT, Hsu PC and Chow SE:
Downregulation of lumican enhanced mitotic defects and aneuploidy
in lung cancer cells. Cell Cycle. 19:97–108. 2020. View Article : Google Scholar
|
|
41
|
Chen X, Li X, Hu X, Jiang F, Shen Y, Xu R,
Wu L, Wei P and Shen X: LUM Expression and Its Prognostic
Significance in Gastric Cancer. Front Oncol. 10:6052020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Karamanou K, Franchi M, Piperigkou Z,
Perreau C, Maquart FX, Vynios DH and Brézillon S: Lumican
effectively regulates the estrogen receptors-associated functional
properties of breast cancer cells, expression of matrix effectors
and epithelial-to-mesenchymal transition. Sci Rep. 7:451382017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jeanne A, Untereiner V, Perreau C, Proult
I, Gobinet C, Boulagnon-Rombi C, Terryn C, Martiny L, Brézillon S
and Dedieu S: Lumican delays melanoma growth in mice and drives
tumor molecular assembly as well as response to matrix-targeted
TAX2 therapeutic peptide. Sci Rep. 7:77002017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rahimi RA and Leof EB: TGF-β signaling: A
tale of two responses. J Cell Biochem. 102:593–608. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Herpin A and Cunningham C: Cross-talk
between the bone morphogenetic protein pathway and other major
signaling pathways results in tightly regulated cell-specific
outcomes. FEBS J. 274:2977–2985. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Iozzo RV, Buraschi S, Genua M, Xu SQ,
Solomides CC, Peiper SC, Gomella LG, Owens RC and Morrione A:
Decorin antagonizes IGF receptor I (IGF-IR) function by interfering
with IGF-IR activity and attenuating downstream signaling. J Biol
Chem. 286:34712–34721. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Baker J, Liu JP, Robertson EJ and
Efstratiadis A: Role of insulin-like growth factors in embryonic
and postnatal growth. Cell. 75:73–82. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu JP, Baker J, Perkins AS, Robertson EJ
and Efstratiadis A: Mice carrying null mutations of the genes
encoding insulin-like growth factor I (Igf-1) and type 1 IGF
receptor (Igf1r). Cell. 75:59–72. 1993.PubMed/NCBI
|
|
49
|
Vlahovic G, Meadows KL, Hatch AJ, Jia J,
Nixon AB, Uronis HE, Morse MA, Selim MA, Crawford J, Riedel RF, et
al: A Phase I Trial of the IGF-1R Antibody Ganitumab (AMG 479) in
Combination with Everolimus (RAD001) and Panitumumab in Patients
with Advanced Cancer. Oncologist. 23:782–790. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Peterse EF, Cleven AH, De Jong Y,
Briaire-de Bruijn I, Fletcher JA, Danen EH, Cleton-Jansen AM and
Bovée JV: No preclinical rationale for IGF1R directed therapy in
chondro-sarcoma of bone. BMC Cancer. 16:4752016. View Article : Google Scholar
|
|
51
|
Schönherr E, Sunderkötter C, Iozzo RV and
Schaefer L: Decorin, a novel player in the insulin-like growth
factor system. J Biol Chem. 280:15767–15772. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Aggelidakis J, Berdiaki A, Nikitovic D,
Papoutsidakis A, Papachristou DJ, Tsatsakis AM and Tzanakakis GN:
Biglycan regulates MG63 osteosarcoma cell growth through a LPR6/
β-catenin/IGFR-IR signaling axis. Front Oncol. 8:4702018.
View Article : Google Scholar
|
|
53
|
Matsumura T, Whelan MC, Li XQ and Trippel
SB: Regulation by IGF-I and TGF-beta1 of Swarm-rat chondrosarcoma
chon-drocytes. J Orthop Res. 18:351–355. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Metalli D, Lovat F, Tripodi F, Genua M, Xu
SQ, Spinelli M, Alberghina L, Vanoni M, Baffa R, Gomella LG, et al:
The insulin-like growth factor receptor I promotes motility and
invasion of bladder cancer cells through Akt- and mitogen-activated
protein kinase-dependent activation of paxillin. Am J Pathol.
176:2997–3006. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Cantini G, Lombardi A, Piscitelli E, Poli
G, Ceni E, Marchiani S, Ercolino T, Galli A, Serio M, Mannelli M,
et al: Rosiglitazone inhibits adrenocortical cancer cell
proliferation by interfering with the IGF-IR intracellular
signaling. PPAR Res. 2008:9040412008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Qin J, Shaukat I, Mainard D, Netter P,
Barré L and Ouzzine M: Constitutive activation of EGFR is
associated with tumor progression and plays a prominent role in
malignant phenotype of chondrosarcoma. Oncotarget. 10:3166–3182.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kamemura N, Murakami S, Komatsu H, Sawanoi
M, Miyamoto K, Ishidoh K, Kishimoto K, Tsuji A and Yuasa K: Type II
cGMP-dependent protein kinase negatively regulates fibroblast
growth factor signaling by phosphorylating Raf-1 at serine 43 in
rat chondrosarcoma cells. Biochem Biophys Res Commun. 483:82–87.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Guan PP, Yu X, Guo JJ, Wang Y, Wang T, Li
JY, Konstantopoulos K, Wang ZY and Wang P: By activating matrix
metalloproteinase-7, shear stress promotes chondrosarcoma cell
motility, invasion and lung colonization. Oncotarget. 6:9140–9159.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lee KS, Kim SW and Lee HS: Orostachys
japonicus induce p53-dependent cell cycle arrest through the MAPK
signaling pathway in OVCAR-3 human ovarian cancer cells. Food Sci
Nutr. 6:2395–2401. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Boroumand Moghaddam A, Moniri M, Azizi S,
Abdul Rahim R, Bin Ariff A, Navaderi M and Mohamad R: Eco-Friendly
Formulated Zinc Oxide Nanoparticles: Induction of Cell Cycle Arrest
and Apoptosis in the MCF-7 Cancer Cell Line. Genes (Basel).
8:2812017. View Article : Google Scholar
|
|
61
|
Dai X, Ma W, He X and Jha RK: Review of
therapeutic strategies for osteosarcoma, chondrosarcoma, and
Ewing's sarcoma. Med Sci Monit. 17:RA177–RA190. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Mery B, Espenel S, Guy JB, Rancoule C,
Vallard A, Aloy MT, Rodriguez-Lafrasse C and Magné N: Biological
aspects of chon-drosarcoma: Leaps and hurdles. Crit Rev Oncol
Hematol. 126:32–36. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Philippou A, Christopoulos PF and
Koutsilieris DM: Clinical studies in humans targeting the various
components of the IGF system show lack of efficacy in the treatment
of cancer. Mutat Res Rev Mutat Res. 772:105–122. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Fiedler LR, Schönherr E, Waddington R,
Niland S, Seidler DG, Aeschlimann D and Eble JA: Decorin regulates
endothelial cell motility on collagen I through activation of
insulin-like growth factor I receptor and modulation of α2β1
integrin activity. J Biol Chem. 283:17406–17415. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Santra M, Eichstetter I and Iozzo RV: An
anti-oncogenic role for decorin. Down-regulation of ErbB2 leads to
growth suppression and cytodifferentiation of mammary carcinoma
cells. J Biol Chem. 275:35153–35161. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhang W, Ge Y, Cheng Q, Zhang Q, Fang L
and Zheng J: Decorin is a pivotal effector in the extracellular
matrix and tumour micro-environment. Oncotarget. 9:5480–5491. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Xing X, Gu X and Ma T: Knockdown of
biglycan expression by RNA interference inhibits the proliferation
and invasion of, and induces apoptosis in, the HCT116 colon cancer
cell line. Mol Med Rep. 12:7538–7544. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Schaefer L, Tredup C, Gubbiotti MA and
Iozzo RV: Proteoglycan neofunctions: Regulation of inflammation and
autophagy in cancer biology. FEBS J. 284:10–26. 2017. View Article : Google Scholar :
|
|
69
|
Liu B, Xu T, Xu X, Cui Y and Xing X:
Biglycan promotes the chemotherapy resistance of colon cancer by
activating NF-κB signal transduction. Mol Cell Biochem.
449:285–294. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Schulz GB, Grimm T, Sers C, Riemer P,
Elmasry M, Kirchner T, Stief CG, Karl A and Horst D: Prognostic
value and association with epithelial-mesenchymal transition and
molecular subtypes of the proteoglycan biglycan in advanced bladder
cancer. Urol Oncol. 37:530.e9–530.e18. 2019. View Article : Google Scholar
|