Introduction
Colorectal cancer (CRC) is a tumor type
characterized by high patient morbidity and mortality (1). At present, surgery, radiotherapy and
chemotherapy are the primary strategies for CRC treatment (2). Although some progress has been made
in regards to CRC treatment on account of improvement in the
medical level, the strong metastatic tendency of CRC still
increases the risk of patient death (3). Notably, drug resistance and the
generation of side effects are the primary obstacles for CRC
treatment (4). Therefore,
investigating the pathological progression of CRC is essential and
beneficial to investigate new diagnostic and treatment strategies
for CRC.
Long non-coding RNAs (lncRNAs), working as pivotal
intracellular regulatory molecules, have functional bioactivity in
various physiological processes (5). Crucially, lncRNAs have been proven to
be pivotal regulators in tumor progression. In gastric cancer,
lncRNA MAGI2-AS3 was found to be primarily distributed in the
cytoplasm and was found to accelerate tumor progression by
interacting with miR-141/200a (6).
In cervical cancer, lncRNA SOX21-AS1 overexpression was found to
inhibit cervical cancer progression, and its methylation was found
to have clinical prognostic value (7). In prostate cancer, lncRNA SNHG7 was
demonstrated to regulate tumor progression via the miR-324-3p/WNT2B
axis (8). Consistently, reports
indicate that multiple lncRNAs participate in the regulation of CRC
processes. For example, lncRNA LEF1-AS1 was found to accelerate CRC
progression by targeting the miR-489/DIAPH1 axis (9). lncRNA MIR503HG alleviated CRC
progression by downregulating TGF-β2 expression (10). In addition, lncRNA TTN-AS1
accelerated CRC progression by sponging miR-376a-3p (11). LUNAR1 is a 491-nucleotide
transcript located at 15q26.3, with 4 exons and a poly(A) tail
(12). Increasing evidence
indicates that LUNAR1 participates in tumor progression. In diffuse
large B-cell lymphoma, LUNAR1 was found to serve as a candidate
prognostic biomarker via growth regulation (13). Moreover, the novel Notch-induced
LUNAR1 promoted the proliferation of cancer cells, and may serve as
a prognostic marker for CRC (14).
These findings showed that LUNAR1 may act as an oncogene and plays
an important role in cancer progression. However, to date, the
roles of LUNAR1 in CRC progression have not been fully
elucidated.
A microRNA (miRNA/mRNA) is defined as a series of
non-coding RNA sequences of approximately 18-22 bp (15). Reports indicate that miRNAs play
essential roles in gene expression via regulating
post-transcriptional translation, which may be involved in the
regulation of the progression of various diseases (16). Research has demonstrated that
miR-495-3p participates in the regulation of the progression of
multiple diseases. For example, miR-495-3p was found to inhibit
rifampicin-induced SULT2A1 expression via accel-erating the
degradation of mRNAs, which in turn reduces hepatocellular toxicity
(17). Increasing evidence
indicates that miRNAs might be sponged by lncRNAs to regulate
disease-related gene expression, resulting in the regulation of
disease progression. Hence, lncRNAs may serve as competitive
endogenous RNAs (ceRNAs) (18).
For example, miR-495-3p is sponged by lncRNA NEAT1 to regulate
STAT3 expression, resulting in alleviation of septicemia-related
inflammatory response (19).
Moreover, it is worth noting that miR-495-3p serves as an
inhibitory regulator in CRC (20).
However, the relationship between LUNAR1 and miR-495-3p, and the
specific mechanisms involved remain unclear.
C-myc binding protein (MYCBP) plays a vital
role in disease progression. MYCBP binds to the N-terminal region
of MYC corresponding to the transactivation domain via its
C-terminal region and stimulates the activation of E box-dependent
transcription by c-MYC (21). In
esophageal cancer, miR-26a and miR-26b inhibit tumor cell
proliferation by inhibition of MYCBP expression (22). Overexpression of MYCBP binding
protein was found to promote the invasion and migration of gastric
cancer (23). These findings
indicate that MYCBP plays a carcinogenic role in most cancers. In
the present study, we further investigated the specific mechanism
of MYCBP in CRC.
In this research, we aimed to explore the role of
LUNAR1 in CRC progression and the underlying mechanisms by
evaluating the proliferation, migration, invasion, and apoptosis of
CRC cell lines, including SW480 and LoVo cells. Our findings
suggest novel prognostic biomarkers for predicting the progression
and prognosis of CRC.
Materials and methods
Patients
Fifteen CRC patients (age range, 25-60 years,
average age, 42; 7 males and 8 females) at The First Affiliated
Hospital, College of Medicine, Zhejiang University (Hangzhou,
Zhejiang, China) between March 2018 and March 2019 were surveyed.
These patients did not receive chemotherapy and radiotherapy before
the operation; and did not present with diseases such as infectious
diseases and multiple cancers. The clinical staging was based on
the TNM analysis system of Union for International Cancer Control,
UICC (version 8). All patients were informed before their
inclusion; written consent of the patients was obtained.
Multivariate analysis was performed to identify factors associated
with overall survival using the Cox proportional hazards model.
Tissue specimens
Tumor tissues or corresponding paracancerous tissues
were obtained by surgical extraction from 15 CRC patients (age
range, 25-60 years, average age, 42; 7 males and 8 females) at The
First Affiliated Hospital, College of Medicine, Zhejiang University
(Hangzhou, Zhejiang, China) between March 2018 and March 2019. All
experimental protocols were approved by the Ethics Committee of The
First Affiliated Hospital, College of Medicine, Zhejiang University
(Zhejiang, China; ethical approval no. PRO20180916-R1) and
experimental procedures were conducted according to the Declaration
of Helsinki Principles.
Cell culture
CRC cells lines, including HT29, LoVo, SW480, SW620
cells and normal HIEC cells which served as the control were
obtained from Kunming Medical University (Kunming, Yunnan, China).
Dulbecco's modified Eagle's medium (DMEM; Roche) supplemented with
10% fetal bovine serum (FBS) (Roche) and 1% penicillin-streptomycin
solution (Solarbio) was applied to the cultured cells in a humid
incubator containing 5% CO2 at 37°C.
Cell transfection
The transfection doses for pLKO.1 plasmid shRNAs
targeting lncRNA LUNAR1, MYCBP and its negative control sh-NC
(synthesized by Sangon Biotech) were 500 ng for cells in each well
of 6-well plates. The transfection doses of miR-495-3p mimics or
inhibitors (synthesized by Sangon Biotech), as well as their
corresponding controls were 100 nM for cells in each well of 6-well
plates. The transfection was performed using Lipofectamine™ 3,000
Transfection Reagent (Takara). Following a 48-h transfection, the
SW480 and LoVo cells were applied to subsequent experiments.
Detailed sequences for these shRNAs, mimics and inhibitors are
presented in Table I.
 | Table IDetailed information regarding the
sequences of the miRNA mimics, inhibitors and shRNAs. |
Table I
Detailed information regarding the
sequences of the miRNA mimics, inhibitors and shRNAs.
| Sequence name | Sequences
(5′-3′) |
|---|
| miR-495-3p
mimics |
5′-AAACAAACATGGTGCACTTCTT-3′ |
| NC mimics |
5′-ATCGTGCTAGTCGATGCTAGCT-3′ |
| miR-495-3p
inhibitors |
5′-GCTTTATATGTGACGAAACAA-3′ |
| NC inhibitors |
5′-CGATCGCAGCGGTGCAGTGCG-3′ |
| sh-LncRNA
LUNAR1 |
5′-GCCTGTTGAGTCACAGTTTCC-3′ |
| sh-MYCBP |
5′-GCCCATTACAAAGCCGCCGAC-3′ |
| sh-NC |
5′-CGATGTCGTAGCTGACTGACG-3′ |
RT-qPCR
Trizol reagent (Takara) was applied to extracted
total RNA from CRC cell lines or tissues. M-MLV Reverse
Transcriptase (RNase H) kit (Takara) was performed to synthesize
cDNA. RT-qPCR was performed as previously described (24). Primers applied to this research are
shown in Table II.
| Table IIPrimer sequences. |
Table II
Primer sequences.
| Primer name | Primer sequences
(5′-3′) |
|---|
| F-LUNAR1 |
5′-CTCAGTAGCTCTCTCTCTCTCTCTCTCT-3′ |
| R-LUNAR1 |
5′-TTGTCTCCCTAGATATCA-3′ |
| F-MYCBP | 5′-ATGGCCCATTA
CAAGCCGC-3′ |
| R-MYCBP | 5′-CTATTCAGCG
CTCTCTCTCTCTCT-3′ |
| F-GAPDH |
5′-GAGTCAACGGATTTGGTCGT-3′ |
| R-GAPDH |
5′-TTGATTTTGGAGGGATCTCG-3′ |
| F-miR-495-3p |
5′-AAACAAACAUGGUGCACUUCUU-3′ |
| R-miR-495-3p |
5′-GAAGUGCACCAUGUUUGUUUUU-3′ |
| F-U6 |
5′-CTCGCTTCGGCAGCACA-3′ |
| R-U6 |
5′-AACGCTTCACGAATTTGCGT-3′ |
Subcellular fractionation analysis
PARIS™ kit (Invitrogen; Thermo Fisher Scientific,
Inc.) was applied for subcellular fractionation analysis, according
to the manufacturer's instructions. Nuclear and cytoplasmic
extraction reagents (Beyotime Institute of Biotechnology) were
applied to separate cytoplasm and nuclear grade from SW480 or LoVo
cells. RT-qPCR was conducted to analyze cytoplasmic and nuclear RNA
extracts; GAPDH and U6 served as normalizing controls,
respectively.
Western blot analysis
Total proteins were isolated from SW480 and LoVo
cells which were transfected with corresponding plasmids by using
cell lysis buffer (Beyotime Institute of Biotechnology). Western
blot analysis was conducted as previously described (25). All antibodies, including cyclin D1
(ab16663), p21 (ab109520), Bax (ab32503), Bcl-2 (ab32124),
cleaved-caspase 3 (ab2302), cleaved-caspase 9 (ab2324), Cox-2
(ab179800), MMP-2 (ab92536), MMP-9 (ab76003), MYCBP (ab86078) and
GAPDH (ab9485) used in this research were obtained from Abcam
(dilution, 1:1,000). The optical density of protein bands was
quantified by Image J software1.8.0 (National Institutes of Health,
Bethesda, MD, USA).
CCK-8 assay
SW480 or LoVo cell proliferation was analyzed by
Cell Counting Kit-8 (Roche) according to the manufacturer's
instructions.
Colony formation assay
Long-term cell proliferation was detected by colony
formation assay. In detail, SW480 and LoVo cells were added into
12-well plates, at a dose of 50 cells per well. Dishes were taken
out when the cell colonies in each well were more than 25. The
numbers of cell clones were counted following staining with 1%
crystal violet for 5 min at room temperature, and images were
captured by an optical microscope (Olympus, CX23).
FACs analysis
A total of 1×105 SW480 or LoVo cells were
cultured in 12-well plate by using serum-free DMEM for 24 h,
centrifuged at 150 × g for 5 min, and washed twice with pre-cooled
1X PBS. Subsequently, the cells were fixed with 70% ethanol
(Solarbio), maintained at 20°C for 15 min, centrifuged at 150 × g
for 5 min, and washed twice with pre-chilled 1X PBS. Next, the
cells were incubated with 10 µg/µl DNase-free RNaseA
(Sigma-Aldrich; Merck KGaA) for 45 min at 37°C to eliminate RNA,
and washed twice with pre-chilled 1X PBS. Finally, following
centrifuging at 150 × g for 5 min, the cells were incubated with 1
mg/ml iodide (Sigma-Aldrich; Merck KGaA) in the dark at 4°C for 12
min. The cell distribution at each phase of the cell cycle was
quantified in a flow cytometer and using ModFit software 3.3
(Verity Software House). In addition, according to the
manufacturer's instructions, Annexin-FITC/PI Apoptosis Detection
Kit (Beyotime Institute of Biotechnology) was combined with flow
cytometry to detect cell apoptosis (19). ModFit software 3.3 was performed to
analyze the data.
Wound-healing migration assay
SW480 and LoVo cells, which were transfected with
corresponding plasmids, were applied to the wound-healing assay.
The detailed procedure was described previously (26).
Transwell chamber assay
A Transwell chamber with a membrane with pores of 8
µm in diameter was applied to assess cell migration.
Specific operation protocol has been previously described (27).
RIP (RNA immunoprecipitation) assay
In detail, RNA immunoprecipitation was performed
using the EZ-Magna RIP kit (Millipore) following the manufacturer's
protocol. SW480 and LoVo cells at 80-90% confluency was scraped
off, and then were lysed in complete RIP lysis buffer, after which
100 µl of whole cell extract was incubated with RIP buffer
containing magnetic beads conjugated with human anti-Ago2 antibody
(Sigma-Aldrich; Merck KGaA, 04-642) and negative control normal
mouse IgG (Sigma-Aldrich; Merck KGaA, 12-371). Anti-SNRNP70
(Sigma-Aldrich; Merck KGaA, SAB1305762) was used as a positive
control for the RIP procedure. Samples were incubated with
Proteinase K with shaking to digest the protein and then
immunoprecipitated RNA was isolated. The RNA concentration was
measured using a NanoDrop (Thermo Fisher Scientific, Inc.) and the
RNA quality was assessed using a bioanalyzer (Agilent Technologies,
Inc.). Furthermore, purified RNA was subjected to RT-qPCR analysis
to demonstrate the presence of the binding targets using respective
primers.
Bioinformatics and luciferase reporter
assay
TargetScan (www.targetscan.org) was conducted to predict the
underlying target genes. The dual-luciferase reporter assay was
performed as previously described (28).
Statistical analysis
The data are expressed as mean ± SD as represented
from three independent experiments, and GraphPad Prism version 5.0
software (GraphPad Software, Inc.) was used for statistical
analysis of all data. Pearson's correlation was performed to
evaluate potential correlations between groups (Pearson's rho).
Student's t test or one-way ANOVA was used for comparison between
two groups, and Tukey post-test was used for comparison between
multiple groups. P-value <0.05 was indicative of a statistically
significant difference.
Results
LUNAR1 is highly expressed and associated
with overall survival of the CRC patients
It has been reported that LUNAR1 participates in the
progression of various tumor types, such as diffuse large B-cell
lymphoma (13). To explore the
roles of LUNAR1 in CRC, RT-qPCR was performed to assess LUNAR1
expression in CRC tissues and cell lines. The findings revealed
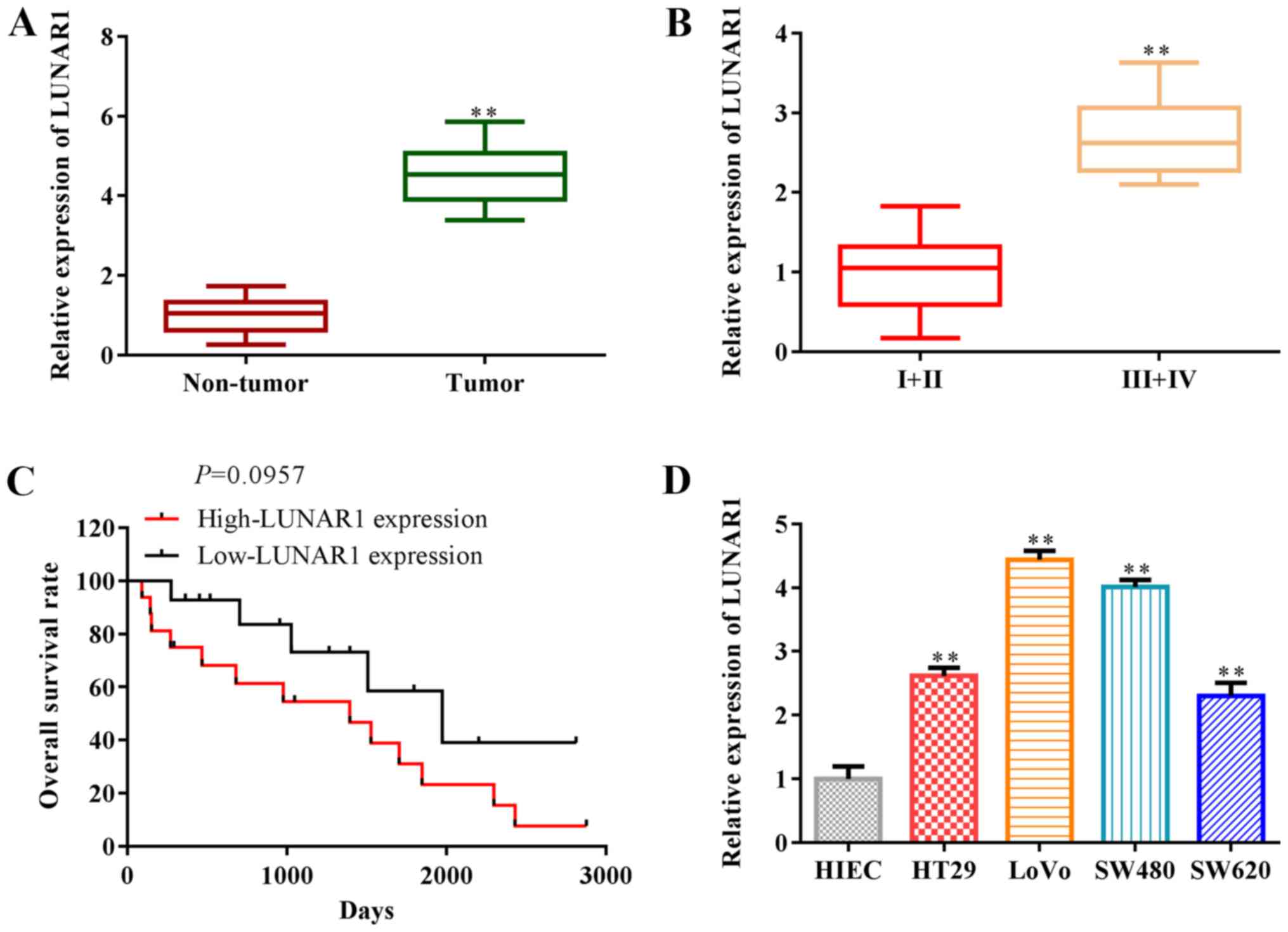
that LUNAR1 expression was significantly upregulated in the tumor
tissues, compared with that in non-tumor tissues (P<0.01;
Fig. 1A), and LUNAR1 expression in
low-grade tumor tissues (n=7; 4 male, 3 female) was lower than that
in high-grade tumor tissues (n=8; 4 male, 4 female) (P<0.01;
Fig. 1B). Overall survival rate
analysis indicated that LUNAR1 expression was negatively associated
with overall survival in the CRC patients (P=0.0957; Fig. 1C). LUNAR1 was upregulated in CRC
cell lines, including HT29, LoVo, SW480, SW620, compared with the
control HIEC cells (P<0.01; Fig.
1D). Note that we selected two types of CRC cells, including
SW480 and LoVo cells in subsequent research, since LUNAR1 was
significantly upregulated in these cells. Collectively, LUNAR1 was
found to be highly expressed and associated with the overall
survival of the CRC patients, indicating that LUNAR1 serves as a
prognostic biomarker for CRC patients.
LUNAR1 knockdown inhibits CRC cell
proliferation and accelerates cell apoptosis
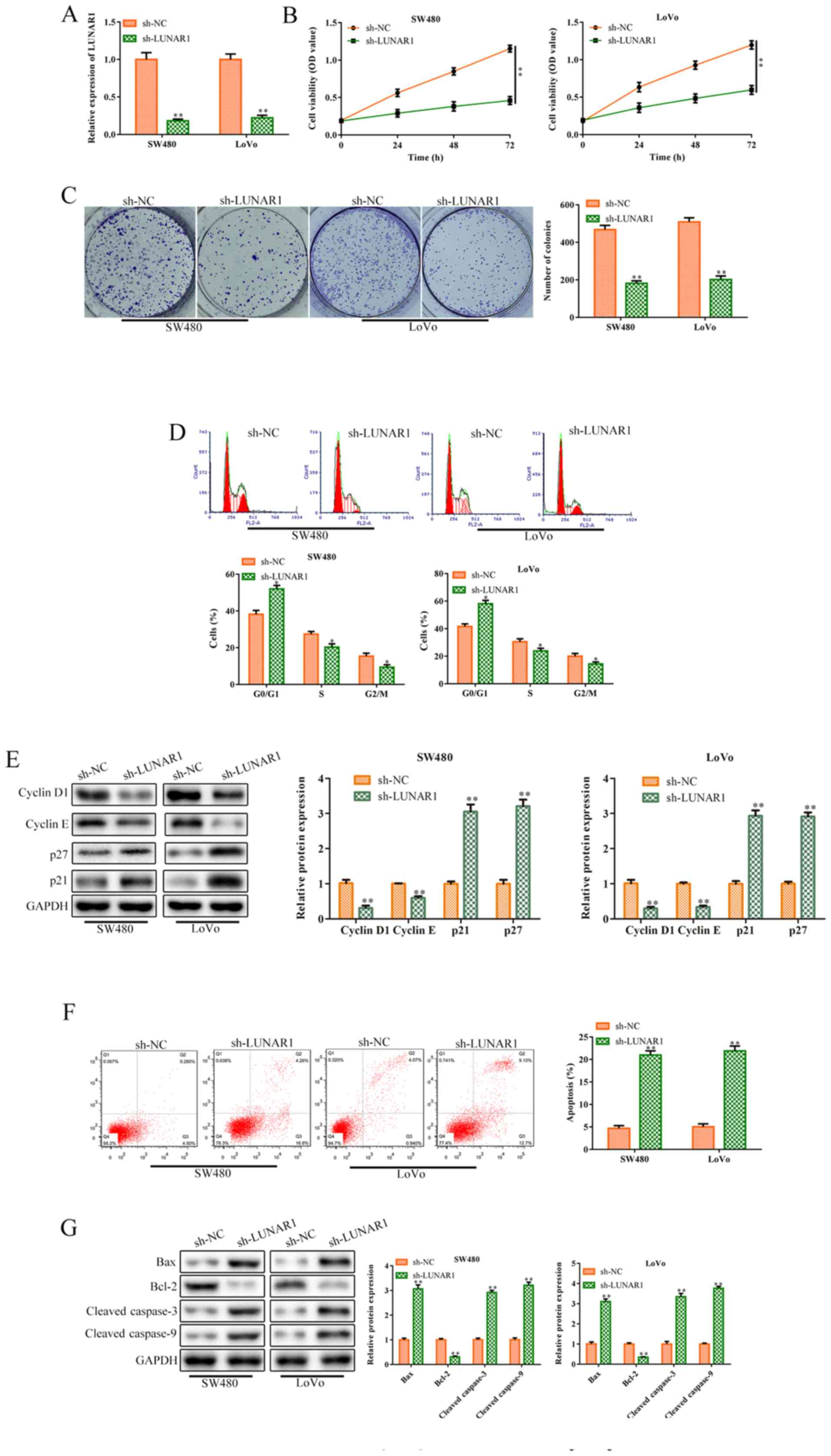
To further explore the roles of LUNAR1 in CRC cell
proliferation, sh-LUNAR1 was transfected into SW480 and LoVo cells
for LUNAR1 knockdown. RT-qPCR analysis indicated that LUNAR1 was
significantly knocked down following sh-LUNAR1 transfection
(P<0.01; Fig. 2A). CCK-8 assay
analysis indicated that LUNAR1 knockdown significantly inhibited
cell viability in the SW480 and LoVo cells, compared to the control
cells (P<0.01; Fig. 2B).
Consistently, colony formation assay revealed that LUNAR1 knockdown
inhibited cell proliferation, compared to the control in both cell
lines (P<0.01; Fig. 2C). Cell
cycle analysis was performed to verify whether the decrease in cell
viability was caused by cell cycle arrest or apoptosis. FACS
analysis showed that LUNAR1 knockdown arrested the cell cycle in
the G1 phase (P<0.05; Fig. 2D),
which was further verified by western blot analysis. In brief,
LUNAR1 knockdown significantly inhibited the levels of cyclin D1
and cyclin E proteins, while increasing p21 and p27 protein
expression (P<0.01; Fig. 2E).
Furthermore, FACs analysis indicated that LUNAR1 knockdown
accelerated cell apoptosis, compared to the control (P<0.01;
Fig. 2F). Consistently, LUNAR1
knock-down upregulated the levels of Bax, cleaved-caspase 3 and
cleaved-caspase 9 proteins, and inhibited Bcl-2 protein expression
(P<0.01; Fig. 2G). Taken
together, LUNAR1 knockdown inhibited CRC cell proliferation and
accelerated apoptosis.
LUNAR1 knockdown inhibits CRC cell
migration and invasion
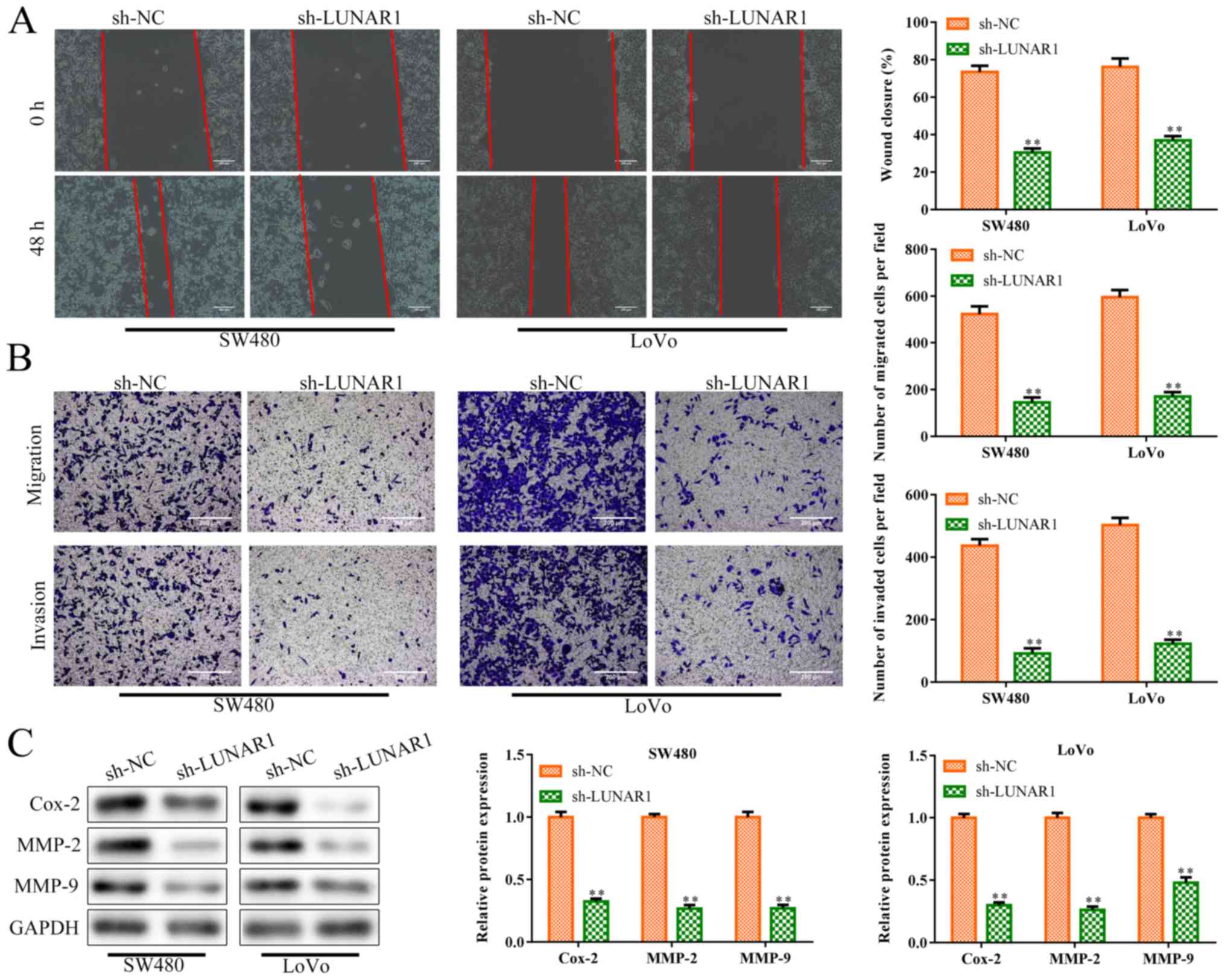
Excessive migration and invasion of tumor cells is a
hallmark of cancer (29).
Therefore, to investigate the roles of LUNAR1 in CRC cell migration
and invasion, sh-LUNAR1 was transfected into SW480 and LoVo cells
in order to knock down LUNAR1. Wound-healing migration assay showed
that LUNAR1 knockdown inhibited cell migration of the SW480 and
LoVo cells, compared to the controls (P<0.01; Fig. 3A). Similarly, as shown in Fig. 3B, LUNAR1 knockdown inhibited cell
migration and invasion abilities (P<0.01). Simultaneously,
western blot analysis (Fig. 3C)
showed that the levels of invasion-related proteins were inhibited
in the SW480 and LoVo cells by LUNAR1 knockdown (P<0.01),
including cyclooxygenase 2 (Cox-2), matrix metalloprotein (MMP)-2
and MMP-9, which play pivotal roles in angiogenesis and tumor
metastasis (30). Collectively,
LUNAR1 knockdown inhibited CRC cell migration and invasion,
indicating that LUNAR1 plays an active role in CRC progression.
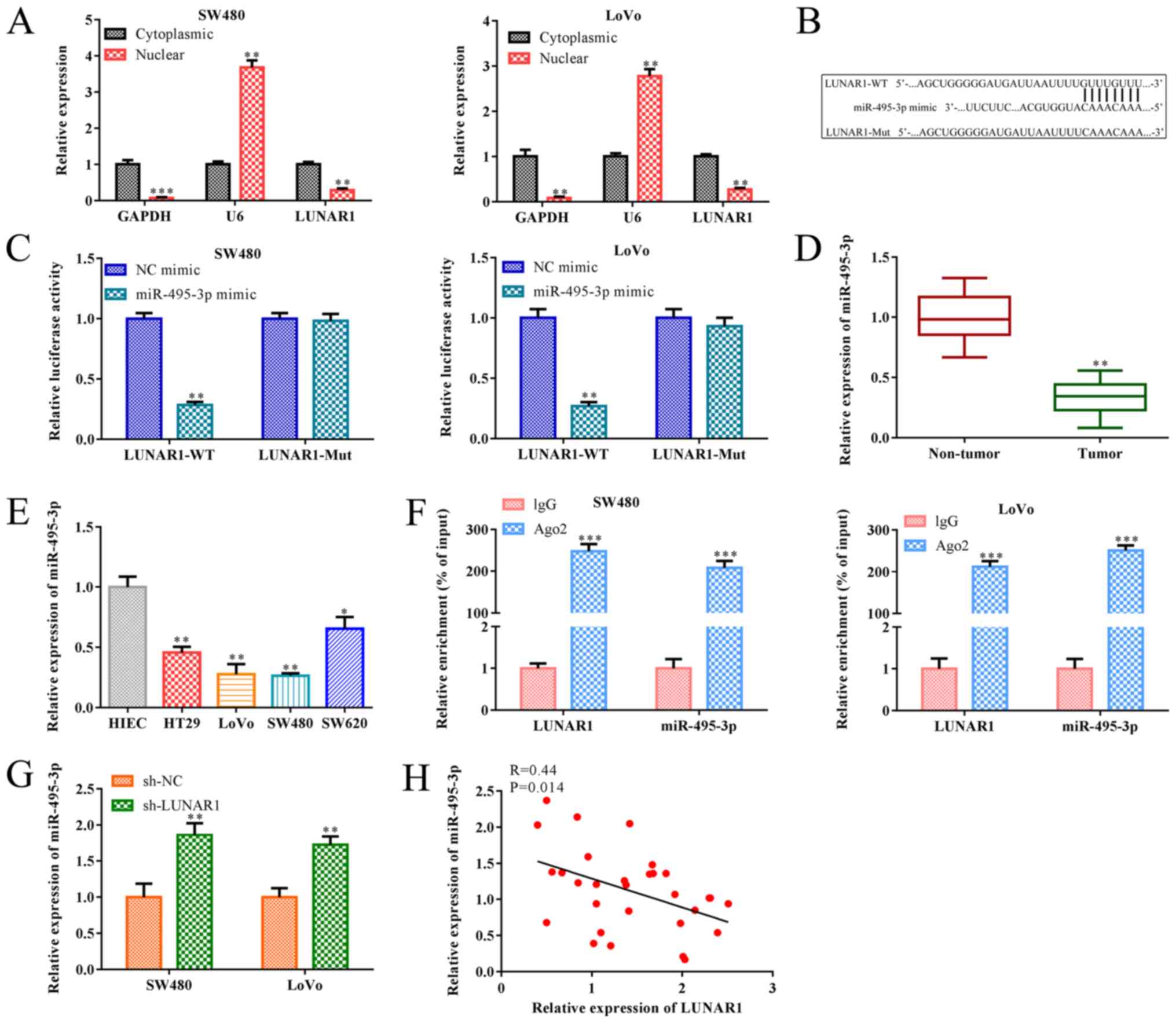
LUNAR1 functions as a sponge of
miR-495-3p in CRC cells
As is known, lncRNAs may function as a ceRNA of
miRNA, and their subcellular localization is closely related to the
biological effects (31).
Therefore, subcellular fractionation analysis was performed to
detect LUNAR1 subcellular distribution in the SW480 and LoVo cells.
RT-qPCR analysis indicated that LUNAR1 was highly expressed in the
cytoplasm and slightly expressed in the nucleus (P<0.01;
Fig. 4A), indicating that LUNAR1
primarily exerted its biological functions in the cytoplasm of
SW480 and LoVo cells. In addition, research has shown that
miR-495-3p serves as an inhibitory regulator in the progression of
various tumors (19). In the
present paper, bioinformatics analysis indicated a putative
interaction between LUNAR1 and miR-495-3p, and the luciferase
reporter assay confirmed that miR-495-3p mimics decreased the
luciferase activity of the wild-type (WT) 3'UTR of LUNAR1 in both
SW480 and LoVo cell lines, whereas this inhibition was blocked when
the putative binding sites were mutated (Mut) (P<0.01; Fig. 4B and C). Furthermore, RT-qPCR
analysis indicated that miR-495-3p was downregulated in the CRC
tissues and cell lines, including HT29, LoVo, SW480, SW620 cells,
when compared to the non-tumor tissues and control NIEC cells
(P<0.05; Fig. 4D and E). As
shown in Fig. 4F, the interaction
between LUNAR1 and miR-495-3p was further verified by RIP assay
where both LUNAR1 and miR-495-3p were enriched in AGO2
immunoprecipitants compared with that of IgG group (P<0.001). In
addition, RT-qPCR analysis showed that LUNAR1 knockdown
significantly accelerated miR-495-3p expression in SW480 and LoVo
cells (P<0.01), and Pearson's correlation analysis showed that
the level of LUNAR1 was negatively correlated with miR-495-3p in
SW480 and LoVo cells (R=0.44; P=0.014; Fig. 4G and H). Taken together, LUNAR1
affects the dysregulation of miR-495-3p via the sponging in CRC
cells.
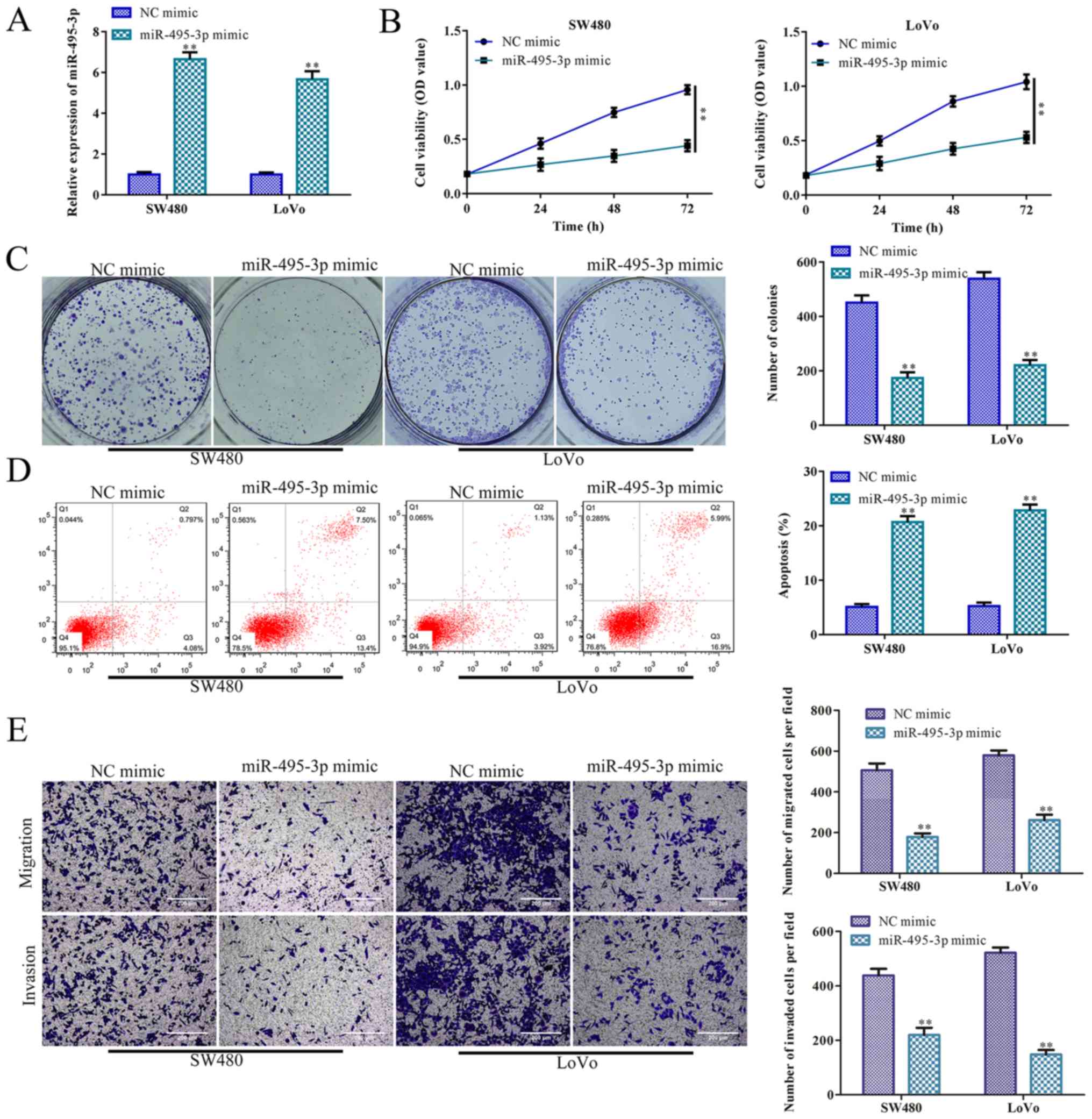
miR-495-3p overexpression inhibits CRC
cell proliferation, migration and invasion, and promotes
apoptosis
It has been reported that miR-495-3p participates in
the regulation of tumor cell bioactivity. For example, miR-495-3p
regulates osteosarcoma cell apoptosis by targeting C1q/TNF-related
protein 3 (32). To investigate
the roles of miR-495-3p in CRC cell proliferation, miR-495-3p was
effectively overexpressed in the SW480 and LoVo cells (P<0.01;
Fig. 5A). CCK-8 and colony
formation assays indicated that SW480 and LoVo cell proliferation
was inhibited by miR-495-3p overexpression, compared to the control
(NC mimic group) (P<0.01; Fig. 5B
and C). In accordance with these results, miR-495-3p
overexpression accelerated SW480 and LoVo cell apoptosis, compared
with the control (P<0.01; Fig.
5D). Furthermore, Transwell chamber assays indicated that SW480
and LoVo cell migration and invasion were significantly inhibited
by miR-495-3p overexpression, compared with the NC mimic groups
(P<0.01; Fig. 5E).
Collectively, miR-495-3p overexpression inhibited CRC cell
proliferation, migration and invasion, and promoted apoptosis,
further indicating that miR-495-3p suppresses CRC progression.
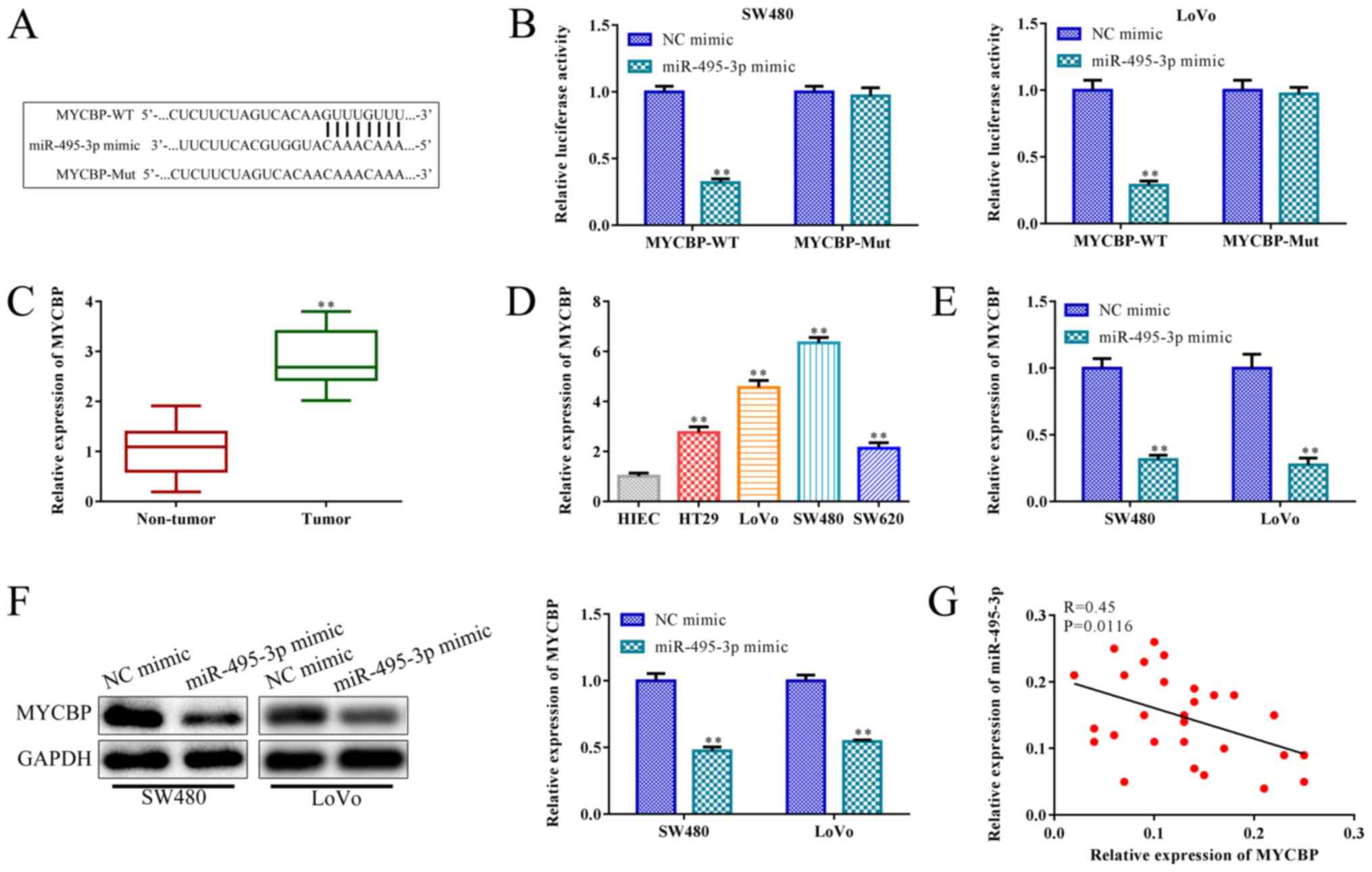
miR-495-3p negatively targets MYCBP
miRNAs participate in the regulation of tumor
progression by regulating the post-transcriptional translation of
target genes (33). To verify the
downstream regulator of miR-495-3p in CRC, bioinformatics analysis
was applied to predict the putative target gene of miR-495-3p. The
findings indicated that a binding site of miR-495-3p was observed
in 3' UTR (untranslated region) of MYCBP (Fig. 6A), which resides on chromosome 1p
and its tumor-promoting effect was confirmed. Subsequently, the
luciferase reporter assay confirmed this prediction. Reduced
luciferase activity was observed in the SW480 and LoVo cells
co-transfected with wild-type (WT) plasmids of MYCBP and miR-495-3p
mimics, whereas this trend was blocked when the putative binding
sites were mutated (Mut) (P<0.01; Fig. 6B). RT-qPCR analysis indicated that
MYCBP expression was upregulated in the CRC tissues and cell lines,
including HT29, LoVo, SW480, SW620 cells when compared with the
non-tumor tissue and HIEC cell line (P<0.01; Fig. 6C and D). Meanwhile, miR-495-3p
overexpression inhibited the MYCBP levels in the SW480 and LoVo
cells, compared to the NC mimic group (P<0.01; Fig. 6E). Consistently, western blot
analysis indicated that miR-495-3p overexpression inhibited MYCBP
expression in the SW480 and LoVo cells, compared to the control
(P<0.01; Fig. 6F). Furthermore,
Pearson's correlation analysis showed that MYCBP was negatively
associated with miR-495-3p in the SW480 and LoVo cells (R=0.45;
P=0.0116; Fig. 6G). Taken
together, miR-495-3p negatively targets MYCBP in SW480 and LoVo
cells.
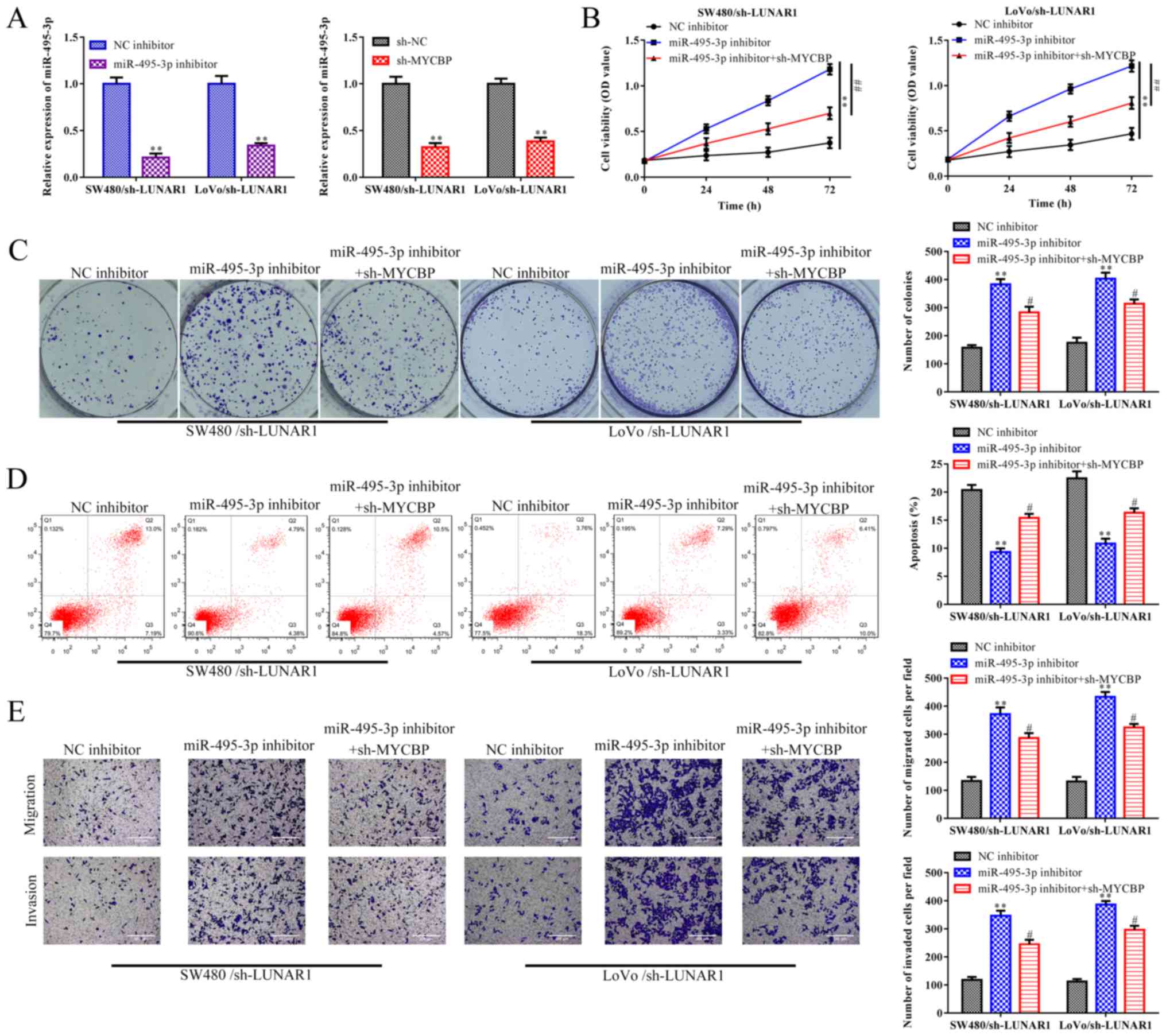
LUNA R1 accelerates CRC progression via
the miR-495-3p/MYCBP axis
Based on the above results, rescue assays were
performed to demonstrate whether LUNAR1 exerts its function in CRC
via the miR-495-3p/MYCBP axis. RT-qPCR analysis indicated that the
increased expression of miR-495-3p promoted by sh-LUNAR1 was
reversed by the miR-495-3p inhibitor in the SW480 and LoVo cells.
In addition, the increased expression of miR-495-3p promoted by
sh-LUNAR1 was also reversed by sh-MYCBP in the SW480 and LoVo cells
(P<0.01; Fig. 7A).
CCK-8 and colony formation assays depicted that
LUNAR1 knockdown-induced reduction in cell proliferation ability of
SW480 and LoVo cells was promoted by miR-495-3p inhibitors, whereas
such an impact of miR-459-3p inhibitors was subsequently recovered
by MYCBP depletion (P<0.05; Fig. 7B
and C). Furthermore, FACs analysis indicated that LUNAR1
knockdown-induced cell apoptosis was abolished by miR-495-3p
inhibitors, while MYCBP knockdown antagonized the effect of
miR-495-3p inhibitors in the LUNAR1-knockdown SW480 and LoVo cells
(P<0.05; Fig. 7D). Similar
results were observed in the migration and invasion abilities of
the SW480 and LoVo cells (P<0.05; Fig. 7E). Collectively, LUNAR1 accelerates
CRC progression via targeting the miR-495-3p/MYCBP axis.
Discussion
Currently, various diagnostic and therapeutic
strategies have been applied to the treatment of CRC. However, the
overall survival rate of CRC patients remains unsatisfactory
(34). Thus, the search for novel
molecular biomarkers for CRC may contribute to the improvement of
clinical treatment (2). In the
present study, we elucidated the specific mechanisms through which
LUNAR1 accelerates CRC progression. Our findings demonstrated that
LUNAR1 accelerated CRC progression via the miR-495-3p/MYCBP axis,
indicating that LUNAR1 may serve as a prognostic biomarker for CRC
patients.
Long non-coding (lnc.) RNAs are defined as a novel
class of non-protein coding RNA with a length of over 200 nt, which
are involved in the regulation of multiple molecular functions
(35). Increasing evidence reveals
that lncRNAs regulate the post-transcriptional translation of genes
by sponging miRNAs, thus playing a vital role in the pathological
process of various diseases (13).
LUNAR1 is a transcription of 491 nucleotides at 15q26.3 with 4
exons and a poly(A) tail (14).
Research has found that LUNAR1 regulates disease progression via
sponging miRNAs, such as acute leukemia and diffuse large B-cell
lymphoma (13). In the present
study, our findings indicated that LUNAR1 is highly expressed and
negatively associated with the overall survival of CRC patients,
indicating that LUNAR1 serves as a prognostic biomarker for CRC
patients. In addition, LUNAR1 knockdown inhibited CRC cell
proliferation, migration and invasion, and accelerated cell
apoptosis. Our findings indicated that LUNAR1 plays an active role
in CRC progression. Mechanistically, LUNAR1 functioned as a sponge
of miR-495-3p, and miR-495-3p over-expression inhibited CRC cell
progression. Hence, LUNAR1 accelerated CRC progression by sponging
miR-945-3p.
Myc binding protein (MYCBP), belonging to the Myc
family is located on chromosome 1p (36). It has been reported that MYCBP, as
a co-activator of c-Myc, accelerates tumor progression due to
carcinogenesis. In glioma, partial loss of MYCBP was found to
significantly improve the overall survival rate of the patients
(37). In ESCC, miR-26a depressed
tumor cell proliferation by inhibiting MYCBP expression (22). In addition, in hepatocellular
carcinoma, EYA4 alleviated tumor progression by inhibiting MYCBP
(36). In the present study, our
results indicated that MYCBP was upregulated in CRC tissues and
cells. The carcinogenic effects of MYCBP were further verified.
Bioinformatics and luciferase reporter analyses showed that MYCBP
was negatively targeted by miR-495-3p, and further functional
verification indicated that LUNAR1 accelerated CRC cell
proliferation, migration and invasion, and inhibited cell apoptosis
via the miR-495-3p/MYCBP axis.
In summary, our findings demonstrated that LUNAR1
was highly expressed in CRC tissues and cell lines and was
negatively associated with overall survival of CRC patients. LUNAR1
knockdown accelerated cell apoptosis and inhibited CRC cell
proliferation, migration and invasion. Moreover, LUNAR1 functioned
as a sponge of miR-495-3p; miR-495-3p overexpression inhibited CRC
cell proliferation, migration and invasion, and depressed cell
apoptosis by negatively targeting MYCBP. Collectively, LUNAR1
accelerates CRC progression through the miR-495-3p/MYCBP axis. Our
findings indicate that LUNAR1 acts as a prognostic biomarker for
CRC patients.
Funding
Not applicable.
Availability of data and materials
The raw data supporting the conclusions of this
manuscript will be made available by the authors, without undue
reservation, to any qualified researcher.
Authors' contributions
JQ and QS conceived and designed the study. AG, FL
and KX performed the experiments. JQ and QS wrote the paper. AG, FL
and KX reviewed and edited the manuscript. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
All patients were informed before their inclusion;
written consent of the patients was obtained. All experimental
protocols were approved by the Ethics Committee of The First
Affiliated Hospital, College of Medicine, Zhejiang University
(Zhejiang, China; ethical approval no. PRO20180916-R1) and
experimental procedures were conducted according to the Declaration
of Helsinki Principles.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing of
interests.
Abbreviations:
|
CRC
|
colorectal cancer
|
|
lncRNA
|
long non-coding RNA
|
|
LUNAR1
|
lncRNA LUNAR1
|
|
ceRNA
|
competitive endogenous RNA
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
FBS
|
fetal bovine serum
|
|
miRNA
|
microRNA
|
|
Cox-2
|
cyclooxygenase 2
|
|
MMP-2
|
matrix metalloprotein 2
|
|
MMP-9
|
matrix metalloprotein 9
|
|
MYCBP
|
Myc binding protein
|
Acknowledgments
Not applicable.
References
|
1
|
Wang Y, Lina L, Xu L, Yang Z, Qian Z, Zhou
J and Suoni L: Arctigenin enhances the sensitivity of cisplatin
resistant colorectal cancer cell by activating autophagy. Biochem
Biophys Res Commun. 520:20–26. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lech G, Słotwiński R, Słodkowski M and
Krasnodębski IW: Colorectal cancer tumour markers and biomarkers:
Recent therapeutic advances. World J Gastroenterol. 22:1745–1755.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tauriello DV, Calon A, Lonardo E and
Batlle E: Determinants of metastatic competency in colorectal
cancer. Mol Oncol. 11:97–119. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zeng A, Hua H, Liu L and Zhao J: Betulinic
acid induces apoptosis and inhibits metastasis of human colorectal
cancer cells in vitro and in vivo. Bioorg Med Chem. 27:2546–2552.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu Y, Xu L, Lu B, Zhao M, Li L, Sun W,
Qiu Z and Zhang B: LncRNA H19/microRNA-675/PPARα axis regulates
liver cell injury and energy metabolism remodelling induced by
hepatitis B X protein via Akt/mTOR signalling. Mol Immunol.
116:18–28. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li D, Wang J, Zhang M, Hu X, She J, Qiu X,
Zhang X, Xu L, Liu Y and Qin S: LncRNA MAGI2-AS3 is regulated by
brd4 and promotes gastric cancer progression via maintaining ZEB1
overexpression by sponging miR-141/200a. Mol Ther Nucleic Acids.
19:109–123. 2020. View Article : Google Scholar
|
|
7
|
Wang R, Li Y, Du P, Zhang X, Li X and
Cheng G: Hypomethylation of the lncRNA SOX21-AS1 has clinical
prognostic value in cervical cancer. Life Sci. 233:1167082019.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Han Y, Hu H and Zhou J: Knockdown of
LncRNA SNHG7 inhibited epithelial-mesenchymal transition in
prostate cancer though miR-324-3p/WNT2B axis in vitro. Pathol Res
Pract. 215:1525372019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cheng Y, Wu J, Qin B, Zou BC, Wang YH and
Li Y: CREB1-induced lncRNA LEF1-AS1 contributes to colorectal
cancer progression via the miR-489/DIAPH1 axis. Biochem Biophys Res
Commun. 526:678–684. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chuo D, Liu F, Chen Y and Yin M: LncRNA
MIR503HG is downregulated in Han Chinese with colorectal cancer and
inhibits cell migration and invasion mediated by TGF-β2. Gene.
713:1439602019. View Article : Google Scholar
|
|
11
|
Wang Y, Jiang F, Xiong Y, Cheng X, Qiu Z
and Song R: LncRNA TTN-AS1 sponges miR-376a-3p to promote
colorectal cancer progression via upregulating KLF15. Life Sci.
244:1169362020. View Article : Google Scholar
|
|
12
|
Trimarchi T, Bilal E, Ntziachristos P,
Fabbri G, Dalla-Favera R, Tsirigos A and Aifantis I: Genome-wide
mapping and characterization of Notch-regulated long noncoding RNAs
in acute leukemia. Cell. 158:593–606. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Peng W and Feng J: Long noncoding RNA
LUNAR1 associates with cell proliferation and predicts a poor
prognosis in diffuse large B-cell lymphoma. Biomed Pharmacother.
77:65–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Z, Li G, Qiu H, Yang J, Bu X, Zhu S,
Zheng J, Dang C, Wang W and Chu D: The novel notch-induced long
noncoding RNA LUNAR1 determines the proliferation and prognosis of
colorectal cancer. Sci Rep. 9:199152019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hawkins LJ and Storey KB: MicroRNA
expression in the heart of Xenopus laevis facilitates metabolic
adaptation to dehydration. Genomics. 112:3525–3536. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tu Y, Ma T, Wen T, Yang T, Xue L, Cai M,
Wang F, Guan M and Xue H: MicroRNA-377-3p alleviates IL-1β-caused
chondrocyte apoptosis and cartilage degradation in osteoarthritis
in part by downregulating ITGA6. Biochem Biophys Res Commun.
523:46–53. 2020. View Article : Google Scholar
|
|
17
|
Li D, Knox B, Chen S, Wu L, Tolleson WH,
Liu Z, Yu D, Guo L, Tong W and Ning B: MicroRNAs hsa-miR-495-3p and
hsa-miR-486-5p suppress basal and rifampicin-induced expression of
human sulfotransferase 2A1 (SULT2A1) by facilitating mRNA
degradation. Biochem Pharmacol. 169:1136172019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou T, Wu L, Ma N, Tang F, Zong Z and
Chen S: LncRNA PART1 regulates colorectal cancer via targeting
miR-150-5p/miR-520h/CTNNB1 and activating Wnt/β-catenin pathway.
Int J Biochem Cell Biol. 118:1056372020. View Article : Google Scholar
|
|
19
|
Xia D, Yao R, Zhou P, Wang C, Xia Y and Xu
S: LncRNA NEAT1 reversed the hindering effects of miR-495-3p/STAT3
axis and miR-211/PI3K/AKT axis on sepsis-relevant inflammation. Mol
Immunol. 117:168–179. 2020. View Article : Google Scholar
|
|
20
|
Lin L, Tong G, Li M, Liu A and Wang S:
MiR-495-3p facilitates colon cancer cell proliferation via
Wnt/β-catenin signaling pathway by restraining Wnt inhibitory
factor. Trop J Pharm Res. 16:21132017. View Article : Google Scholar
|
|
21
|
Jung HC and Kim K: Identification of MYCBP
as a beta-catenin/LEF-1 target using DNA microarray analysis. Life
Sci. 77:1249–1262. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li J, Liang Y, Lv H, Meng H, Xiong G, Guan
X, Chen X, Bai Y and Wang K: miR-26a and miR-26b inhibit esophageal
squamous cancer cell proliferation through suppression of c-MYC
pathway. Gene. 625:1–9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gong L, Xia Y, Qian Z, Shi J, Luo J, Song
G, Xu J and Ye Z: Overexpression of MYC binding protein promotes
invasion and migration in gastric cancer. Oncol Lett. 15:5243–5249.
2018.PubMed/NCBI
|
|
24
|
Dong SM, Cui JH, Zhang W, Zhang XW, Kou
TC, Cai QC, Xu S, You S, Yu DS, Ding L, et al: Inhibition of
translation initiation factor eIF4A is required for apoptosis
mediated by Microplitis bicoloratus bracovirus. Arch Insect Biochem
Physiol. 3:962017.
|
|
25
|
Cui JH, Dong SM, Chen CX, Xiao W, Cai QC,
Zhang LD, He HJ, Zhang W, Zhang XW, Liu T, et al: Microplitis
bicoloratus bracovirus modulates innate immune suppression through
the eIF4E-eIF4A axis in the insect Spodoptera litura. Dev Comp
Immunol. 95:101–107. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pulito C, Mori F, Sacconi A, Goeman F,
Ferraiuolo M, Pasanisi P, Campagnoli C, Berrino F, Fanciulli M,
Ford RJ, et al: Metformin-induced ablation of microRNA 21-5p
releases Sestrin-1 and CAB39L antitumoral activities. Cell Discov.
3:170222017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen L, Zhu M, Yu S, Hai L, Zhang L, Zhang
C, Zhao P, Zhou H, Wang S and Yang X: Arg kinase mediates
CXCL12/CXCR4-induced invadopodia formation and invasion of glioma
cells. Exp Cell Res. 389:1118932020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pei G, Xu L, Huang W and Yin J: The
protective role of microRNA-133b in restricting hippocampal neurons
apoptosis and inflammatory injury in rats with depression by
suppressing CTGF. Int Immunopharmacol. 78:1060762020. View Article : Google Scholar
|
|
29
|
Li Z, Cao Y, Jie Z, Liu Y, Li Y, Li J, Zhu
G, Liu Z, Tu Y, Peng G, Lee DW and Park SS: miR-495 and miR-551a
inhibit the migration and invasion of human gastric cancer cells by
directly interacting with PRL-3. Cancer Lett. 323:41–47. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ahmed EM, Hassan MSA, El-Malah AA and
Kassab AE: New pyridazine derivatives as selective COX-2 inhibitors
and potential anti-inflammatory agents; design, synthesis and
biological evaluation. Bioorg Chem. 95:1034972020. View Article : Google Scholar
|
|
31
|
Zhou Z, Zhu Y, Gao G and Zhang Y: Long
noncoding RNA SNHG16 targets miR-146a-5p/CCL5 to regulate
LPS-induced WI-38 cell apoptosis and inflammation in acute
pneumonia. Life Sci. 228:189–197. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao G, Zhang L, Qian D, Sun Y and Liu W:
miR-495-3p inhibits the cell proliferation, invasion and migration
of osteosarcoma by targeting C1q/TNF-related protein 3. Onco
Targets Ther. 12:6133–6143. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao Y, He J, Gao P, Niu Y, Zhang J, Wang
L, Liu M, Wei X, Liu C, Zhang C, et al: miR-769-5p suppressed cell
proliferation, migration and invasion by targeting TGFBR1 in
non-small cell lung carcinoma. Oncotarget. 8:113558–113570. 2017.
View Article : Google Scholar
|
|
34
|
Kuipers EJ, Grady WM, Lieberman D,
Seufferlein T, Sung JJ, Boelens PG, van de Velde CJ and Watanabe T:
Colorectal cancer. Nat Rev Dis Primers. 5:150652015. View Article : Google Scholar
|
|
35
|
Liu L, Wang HJ, Meng T, Lei C, Yang XH,
Wang QS, Jin B and Zhu JF: lncRNA GAS5 inhibits cell migration and
invasion and promotes autophagy by targeting miR-222-3p via the
GAS5/PTEN-signaling pathway in CRC. Mol Ther Nucleic Acids.
17:644–656. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ma D: 153P-Exosomal LINC00174 facilitates
epithelial-mesenchymal transition in residual hepatocellular
carcinoma after insufficient radiofrequency ablation by regulating
c-JUN/MYCBP/c-Myc axis. Ann Oncol. 30(Suppl 9): ix53–ix54. 2019.
View Article : Google Scholar
|
|
37
|
Lehrer S, Rheinstein PH and Rosenzweig KE:
Loss of MycBP may be associated with the improved survival in 1P
co-deletion of lower grade glioma patients. Clin Neurol Neurosurg.
172:112–115. 2018. View Article : Google Scholar : PubMed/NCBI
|