Introduction
Breast cancer is one of the most commonly diagnosed
malignant cancers and is the second leading cause of cancer-related
deaths among women worldwide (1).
An estimated 36.1% incidence and 8.8% mortality occurred in China
in 2018 (2,3). Although there are treatments for
breast cancer, including individualized treatment, the pathogenesis
of breast cancer has not yet been fully elucidated, leading to poor
outcomes of patients. The 5-year survival rate of patients with
breast cancer is usually <30%, which requires improvement
(4). Therefore, there is an urgent
need to further investigate the carcinogenesis and development of
breast cancer.
Long non-coding RNAs (lncRNAs) are a large group of
transcripts >200 nucleotides with limited protein-coding
potential (5,6). Studies have demonstrated that lncRNAs
serve a range of specific functions in various pathophysiological
processes (6,7), especially in cancer (8,9). For
example, HOX transcript antisense RNA, an oncogenic lncRNA, is
highly expressed in breast cancer, and its expression is associated
with tumor invasiveness and migration (10,11).
Upregulation of lncRNA P21-associated non-coding RNA DNA
damage-activated is associated with poor prognosis and accelerates
cell proliferation in cervical cancer (12). The expression of lncRNA paternally
expressed 10 is upregulated in esophageal cancer tissues compared
with that in adjacent non-neoplastic tissues, and its depletion
suppresses proliferation and invasion and facilitates apoptosis in
esophageal cancer cells (13).
lncRNAs have been recommended for use as indicators for tumor
detection due to their key role in tumors and their stability in
the serum and plasma (14,15). lncRNA MAF BZIP transcription factor
G antisense RNA 1 (MAFG-AS1) is abnormally expressed in various
types of tumors and is involved in the regulation of cancer cell
proliferation and migration (16-18).
However, the potential mechanisms of MAFG-AS1 in breast cancer have
not been elucidated.
MicroRNAs (miRNAs) are a group of endogenous
non-coding RNAs 21-23 nucleotides long that are present in
eukaryotes (19). Numerous studies
have revealed that miRNAs participate in cancer progression. For
example, miR-195 expression is low in head and neck squamous cell
carcinoma tissues compared with that in normal tissues and is
sponged by LINC00355 to affect cell viability, invasion, migration,
the epithelial-mesenchymal transition (EMT) process and apoptosis
of cancer stem cells (20).
miR-7-5p inhibits the proliferation, migration, invasion and EMT
process of non-small cell lung cancer by suppressing
neuro-oncological ventral antigen 2 (21). miR-182 is sponged by prostate
cancer gene expression marker 1 and is involved in the progression
of cervical cancer (22).
miR-150-5p has been reported to serve an essential role in several
types of cancer (23-25). However, whether miR-150-5p exerts a
role in breast cancer remains unknown.
The present study aimed to explore the role of
MAFG-AS1 and its regulatory mechanisms in breast cancer, which
might provide new targets for breast cancer treatment.
Materials and methods
GEPIA analysis
The GEPIA database (http://gepia.cancer-pku.cn/) was used to analyze the
expression of MAFG-AS1 in breast cancer tissues. The breast cancer
dataset was selected, and the levels of MAFG-AS1 in breast cancer
tissues (n=1,085) and non-tumor tissues (n=291) were plotted.
Cell culture and clinical specimens
Breast cancer cells (MDA-MB-231, MCF-7 and
MDA-MB-468) and the normal epithelial breast cell line MCF-10A were
obtained from ATCC. The cells were cultured in RPMI-1640 (HyClone;
Cytiva) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.) at 37°C with 5% CO2.
A total of 20 samples of breast cancer tissues
(median age, 48 years; range, 37-65 years) and 15 samples of
adjacent non-tumor tissues (3-5 cm distal to the edge of tumor)
were obtained after resection and immediately frozen at −80°C at
the First Hospital of Jilin University (Changchun, China) between
April 2018 and June 2019. Based on the Tumor-Node-Metastasis
staging system (26), five cases
were stage I, 11 cases were stage II, and four cases were stage
III. The clinical study was approved by the Ethics Committee of the
First Hospital of Jilin University. All patients participating in
the study provided written informed consent. All procedures were
performed according to the guidelines revised by the First Hospital
of Jilin University and in accordance with the 1964 Declaration of
Helsinki and its later amendments. The diagnoses were based on
pathology reports. Patients with other tumors, tumor history or
those who received chemotherapy and radiotherapy prior to the
surgery were excluded.
Cell transfection
The MAFG-AS1 sequence and negative control were
constructed by Shanghai GenePharma Co., Ltd. and cloned into the
pcDNA3.1 vector (Shanghai GenePharma Co., Ltd.). The day before
transfection, MCF-7 cells were seeded in 6-well plates at the
confluency of 25-30%. miRNA mimics, inhibitor, short hairpin
(sh)RNA plasmids and the corresponding negative controls (NC
mimics, NC inhibitor and sh-NC) were purchased from Guangzhou
RiboBio Co., Ltd. and were transfected at a final concentration of
50 nM using Lipofectamine® RNAiMax (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. The sequences were as follows: miR-150-5p mimics,
5′-UCU CCC AAC CCU UGU ACC AGU G-3′, NC mimics, 5′-UUC UCC GAA CGU
GUC ACG UTT-3′; miR-150-5p inhibitor, 5′-CAC UGG UCA UCC UUC GGA
GA-3′; NC inhibitor, 5′-UUG UAC UAC ACA AAA GUA CUG-3′;
sh-MAFG-AS1, 5′-TTA TCT TCC TCC CGA GTC C-3′; and sh-NC, 5′-AAT TCT
CCG AAC GTG TCA CGT-3′. The breast cancer cells were transfected
with the above plasmids for 72 h for subsequent functional
experiments.
RNA isolation, reverse
transcription-quantitative (RT-q)PCR
Total RNA was extracted from breast cancer tissues
and cells using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). RNA (500 ng) was reverse-transcribed into
cDNA using a Reverse Transcription kit (Takara Bio, Inc.) for 5 min
at 65°C. The reverse transcription products were diluted 1:5 with
sterile water and stored at -20°C. The subsequent PCR amplification
was performed on an Mx3000P instrument (Agilent Technologies, Inc.)
with SYBR® Premix Ex Taq II (Takara Biotechnology Co.,
Ltd.). The thermocycling conditions were as follows:
Pre-denaturation at 95°C for 10 sec, followed by 40 cycles of
denaturation at 95°C for 10 sec and elongation at 60°C for 30 sec.
GAPDH and U6 were used as internal controls, and the fold-changes
were calculated using the 2−ΔΔCq method (27). The primer sequences used were as
follows: MAFG-AS1 forward, 5′-CGT TCT TAG TTG GTG GAG CG-3′ and
reverse, 5′-CCG GAC ATC TAA GGG CAT CA-3′; GAPDH forward, 5′-ATG
GGA CGA TGC TGG TAC TGA-3′ and reverse, 5′-TGC TGA CAA CCT TGA GTG
AAA T-3′; miR-150-5p stem-loop, 5′-GTC GTA TCC AGT GCA GGG TCC GAG
GTA TTC GCA CTG GAT ACG ACC CAC TGG-3′, forward, 5′-TCTC CCA ACC
CTT GTA-3′ and reverse, 5′-GTG CAG GGT CCG AGG T-3′; U6 forward,
5′-CTC GCT TCG GCA GCA CA-3′ and reverse, 5′-AAC GCT TCA CGA ATT
TGC GT-3′; and MYB forward, 5′-ACA GAT GGG CAG AAA TCG CA-3′ and
reverse, 5′-GCT GGC TGG CTT TTG AAG AC-3′.
Western blotting analysis
Total cellular proteins from MCF-7 cells were
extracted using RIPA lysis buffer (Beyotime Institute of
Biotechnology). Protein concentration was determined using a
bicinchoninic acid assay (Beyotime Institute of Biotechnology), and
10 µg protein/lane was separated by 10% SDS-PAGE followed by
transferring to polyvinylidene fluoride membranes (EMD Millipore).
The membranes were blocked with 5% non-fat milk for 1 h and
incubated at 4°C with the following primary antibodies overnight:
Anti-MYB (cat. no. ab191064; 1:1,000; BD Biosciences),
anti-E-cadherin (cat. no. ab269767; 1:1,000; BD Biosciences),
anti-N-cadherin (cat. no. ab18203; 1:1,000; BD Biosciences),
anti-Bcl-2 (cat. no. ab194583; 1:1,000; BD Biosciences), anti-Bax
(cat. no. ab104156; 1:1,000; BD Biosciences), anti-cyclin A1 (cat.
no. ab53699; 1:1,000; BD Biosciences), anti-CDK2 (cat. no. ab64669;
1:1,000; BD Biosciences) and anti-GAPDH (cat. no. ab9485; 1:1,000;
Affinity Biosciences). The membranes were washed three times using
PBS, incubated with goat anti-rabbit IgG secondary antibody
conjugated to horseradish peroxidase (cat. no. ab7090; 1:5,000;
Abcam) for 1 h at room temperature and visualized using a Pierce
ECL Western Blotting kit (Pierce; Thermo Fisher Scientific, Inc.).
The protein expression was quantified using Image-Pro®
Plus software (version 6.0; Media Cybernetics, Inc.).
Subcellular fractionation
The cytoplasmic and nuclear RNA was isolated from
MCF-7 cells using a PARIS kit (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
cells were lysed in the cell fractionation buffer, followed by
centrifugation for 5 min at 500 × g at 4°C and lysis in cell
disruption buffer. RT-qPCR was used to determine the expression of
MAFG-AS1 in both fractions as aforementioned, with GAPDH and U6 as
cytoplasmic and nuclear controls, respectively.
Luciferase reporter assay
The binding site between miR-150-5p and MAFG-AS1 was
predicted using the starBase website (28). The fragment of MYB or MAFG-AS1
containing the specific binding sites of miR-150-5p was transfected
into the pmirGLO Dual-luciferase vector (Promega Corporation) to
construct the reporter vectors containing wild-type MYB (WT-MYB) or
wild-type MAFG-AS1 (WT-MAFG-AS1) 3′ untranslated region (3′UTR).
Three online algorithms miRDB (http://mirdb.org/), TargetScan (http://www.targetscan.org/vert_72/) and miRanda
(http://www.miranda.org/) were used to predict the
potential target genes of miR-150-5p. The mutated MYB (Mut-MYB) or
mutated MAFG-AS1 (Mut-MAFG-AS1) vectors contained mutated binding
sites. MCF-7 cells were co-transfected with WT-MAFG-AS1/MYB or
Mut-MAFG-AS1/MYB and miR-150-5p mimics using
Lipofectamine® RNAiMax for 48 h at 22°C. The luciferase
activity was detected using the Dual-Luciferase Reporter Assay
System (Promega Corporation). Renilla luciferase activity
was used to normalize the firefly luciferase activity.
Cell viability detection assay
After transfection with the indicated plasmids,
MCF-7 cells (2×103 cells/well) were re-seeded on 96-well
plates. The viability of breast cancer cells was assessed by the
MTS method (Promega Corporation) at daily intervals for 3 days
post-transfection. At each time point, MTS was added to the
corresponding plates and then incubated for 2 h at 37°C according
to the manufacturer's instructions. The optical density values were
measured at 490 nm using a Multiskan EX microplate reader
(Labsystems Diagnostics Oy).
Wound healing assay
MCF-7 were reseeded in a new 6-well plate
(~5×105 cells/well) after transfection for 24 h and
cultured to 80-90% confluence. Subsequently, a 200-µl
sterile pipette tip was used to generate a straight scratch. The
cells were cultured in serum-free medium in a humidified incubator
at 37°C with 5% CO2. Finally, the migration distance was
observed, and images were captured at 0 h and 24 h post-scratch.
The scratch area was recorded by a light microscope (Olympus
Corporation) and analyzed with ImageJ version 1.47 software
(National Institutes of Health). The distance of cell migration
into the wound was measured to calculate the wound healing rate as
follows: Wound healing rate (%)=(wound distance at 0 h-wound
distance at 24 h)/wound distance at 0 h × 100%.
Cell cycle and apoptosis analysis
MCF-7 cells (1×106 cells/well) were fixed
with 70% pre-cooled ethanol overnight at 4°C, treated with RNase A
(Sigma-Aldrich; Merck KGaA) at 37°C for 30 min and stained with
propidium iodide (PI) (Nanjing Keygen Biotech Co., Ltd.) staining
solution at 4°C for 30 min following transfection with miRNA
mimics. Subsequently, each group of cells was tested using a BD
FACSCalibur flow cytometer (BD Biosciences), and the results were
analyzed using FlowJo7.6 software (FlowJo Software, LLC). For
apoptosis, an Annexin V-FITC/propidium iodide (PI) staining kit (BD
Biosciences) was used. Briefly, MCF-7 cells suspended in 100
µl binding buffer (1×105 cells/ml) were incubated
at room temperature for 15 min in the dark with 5 µl Annexin
V-FITC and 5 µl PI solution. The apoptotic cells were
quantified using a BD FACSCalibur flow cytometer, and the data were
analyzed with BD Accuri™ C6 software (BD Biosciences). The
apoptotic rate was calculated using the following formula:
Apoptotic rate=(early apoptotic cells + late apoptotic cells)/total
cell number ×100%.
Xenograft study
A total of 50 (n=6-8 mice/group) female BALB/C nude
mice at age of 6-8 weeks were obtained from Beijing Vital River
Laboratory Animal Technology Co., Ltd. The animal experiments were
approved by the Ethics Committee of Animal Experiments of the First
Hospital of Jilin University (approval no. DWLL-2019-0021). All
mice were housed in a 12-h light/dark cycle at 22°C with 40-70%
humidity for at least 7 days before the experiments. Mozart K448
sonata was used to alleviate fear and stress, and the mice had free
access to water and food. Animal health and behavior was monitored
every day. The mice were randomly divided into five groups: pcDNA +
NC mimics; MAFG-AS1 + NC mimics; MAFG-AS1 + miR-150-5p mimics;
pcDNA + miR-150-5p mimics; and miR-150-5p mimics + MYB. MCF-7 cells
(5×106) suspended in 150 µl PBS were
subcutaneously injected into the right armpit of mice. At 21 days
post-inoculation, the mice were sacrificed by cervical dislocation.
The xenograft tumors were carefully excised from the sacrificed
mice and weighted, and the tumor volume was calculated using the
following formula: Volume=0.5 × length × width2.
Statistical analysis
Data are presented as the mean ± standard deviation.
All experimental data were processed and analyzed using SPSS 22.0
statistical software (IBM Corp.). The Kaplan-Meier plotter database
(http://kmplot.com/analysis/) was used to
analyze the association between miR-150-5p expression and overall
survival (OS) of patients with breast cancer. The total number of
patients was 1,262, and based on the median expression of miR-150,
the patients were divided into high and low expression groups. The
Kaplan-Meier method was used for survival analysis, and the
log-rank test was performed compare the two groups. Differences
between two groups were evaluated by two-tailed Student's t-test.
Statistical significance among three or more groups was assessed by
one- or two-way ANOVA followed by Dunnett's (for comparisons with
one control) or Tukey's (for comparisons among various groups) post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
MAFG-AS1 is upregulated in breast
carcinoma
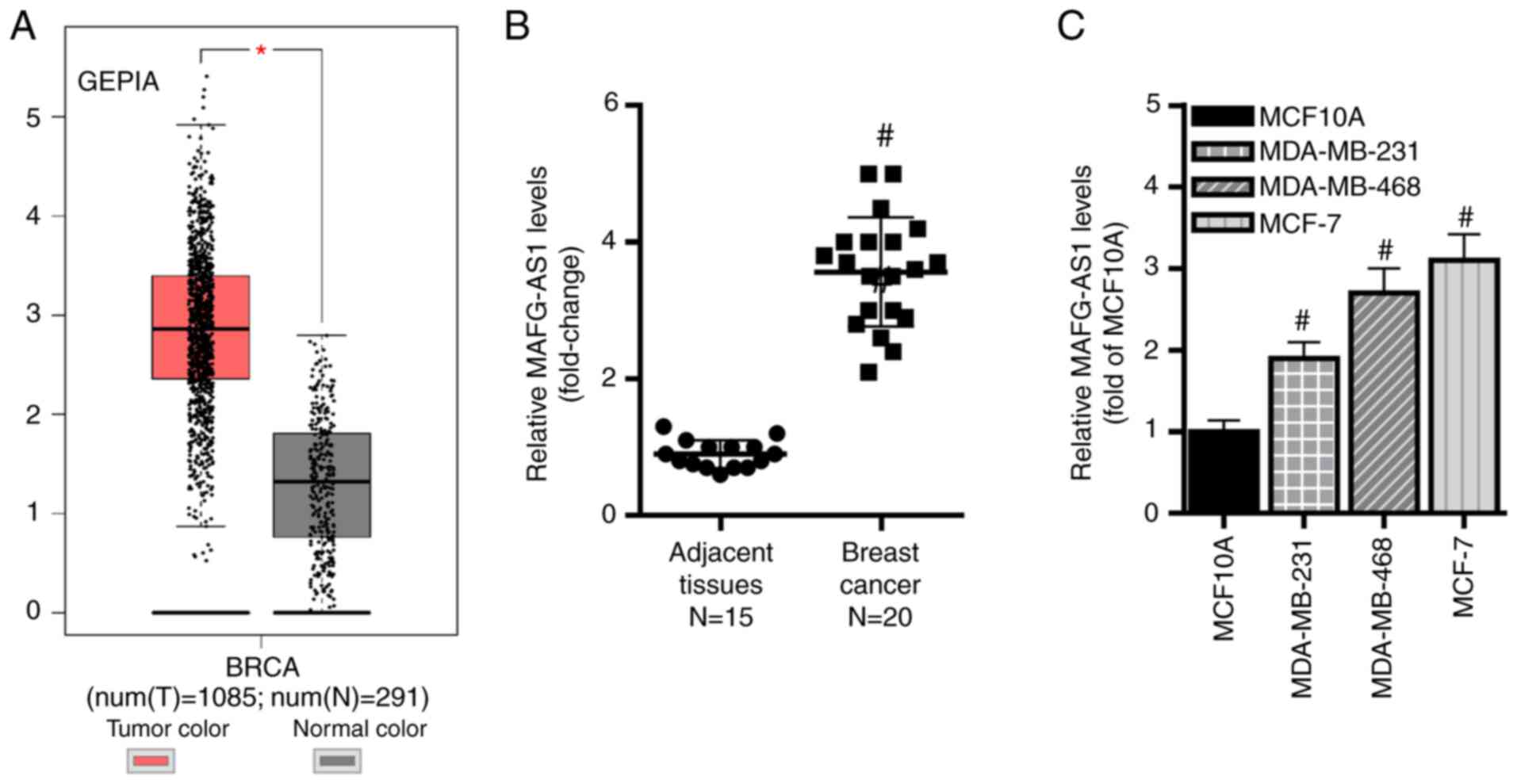
In the present study, the expression levels of
MAFG-AS1 were compared between breast cancer and adjacent non-tumor
tissues from patients using original published data available from
the GEPIA database. The results demonstrated that the expression
MAFG-AS1 was significantly upregulated in breast cancer samples
compared with that in non-tumor samples (Fig. 1A). In addition, RT-qPCR revealed
that MAFG-AS1 expression levels were higher in cancer tissues
compared with those in adjacent non-tumor tissues (Fig. 1B). Similarly, MAFG-AS1 levels were
significantly upregulated in three human breast cancer cell lines
(MDA-MB-468, MDA-MB-231 and MCF-7) compared with those in the
normal breast epithelial cell line MCF-10A (Fig. 1C). These results suggested that
MAFG-AS1 may function as a tumor-promoting lncRNA in breast
cancer.
MAFG-AS1 sponges miR-150-5p
Considering that the competing endogenous RNA
(ceRNA) hypothesis, which indicates that lncRNAs can sponge miRNAs
to upregulate the expression of specific mRNAs and further regulate
the progression of diseases (29),
is currently widely accepted, we hypothesized that MAFG-AS1 may
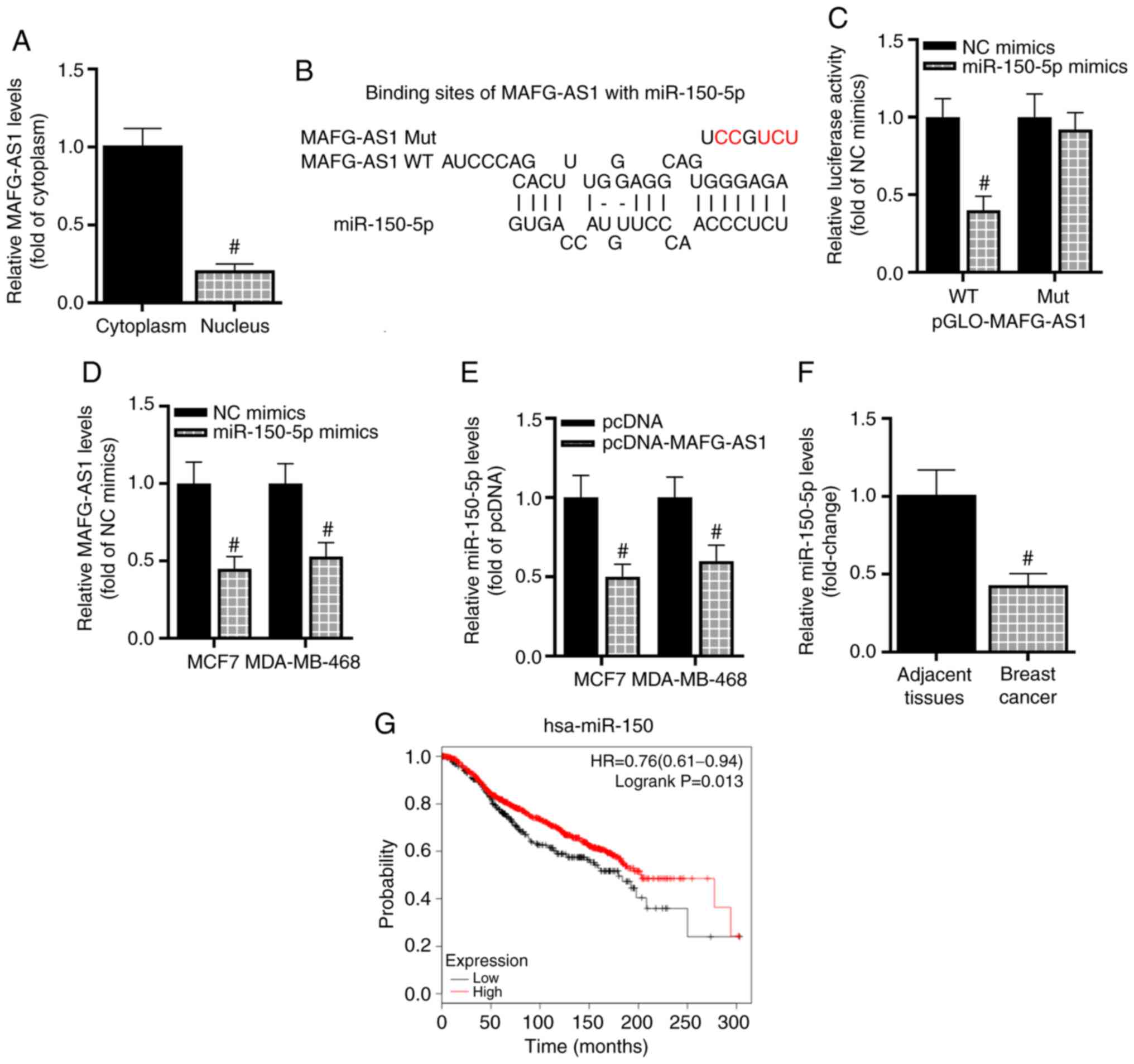
also function in this manner in breast cancer. Cytoplasmic lncRNAs
have been reported to present with a high probability of acting as
ceRNAs (30); thus, cytoplasmic
and nuclear MAFG-AS1 were extracted separately, and RT-qPCR was
used to analyze the expression of MAFG-AS1 in the nucleus and
cytoplasm of breast cancer cells. The results demonstrated that
MAFG-AS1 was mainly concentrated in the cytoplasm of breast cancer
cells (Fig. 2A), suggesting that
MAFG-AS1 may be involved in a ceRNA network in breast cancer. To
identify the potential mechanism of MAFG-AS1 in the modulation of
the development of breast cancer, the miRNAs sponged by MAFG-AS1
were predicted using the online bioinformatics analysis tool
starBase (28). The results
revealed that MAFG-AS1 bound miR-150-5p (Fig. 2B). To overexpress miR-150-5p,
transfection of the miR-150-5p mimics into breast cancer cells was
used (Fig. S1A). The subsequent
luciferase reporter assay results demonstrated that the luciferase
activity of MCF-7 cells was significantly decreased by miR-150-5p
mimics compared with that in the NC mimics group following
transfection with WT-MAFG-AS1, whereas miR-150-5p mimics exerted no
inhibitory effects on the luciferase activity of MCF-7 cells
transfected with Mut-MAFG-AS1 (Fig.
2C). Taken together, these results indicated that MAFG-AS1
sponged miR-150-5p. The relationship between MAFG-AS1 and
miR-150-5p was then further examined; as presented in Fig. 2D, transfection with the miR-150-5p
mimics inhibited the expression of MAFG-AS1 in MCF-7 and MDA-MB-468
cells compared with the NC mimics group. Next, overexpression
efficiency of MAFG-AS1 was verified by RT-qPCR analysis (Fig. S1B). Similarly, the expression of
miR-150-5p was downregulated following overexpression of MAFG-AS1
in breast cancer cells compared with that in cells transfected with
the empty vector (Fig. 2E). In
addition, the mean expression levels of miR-150-5p were
significantly reduced in cancer tissues compared with those in the
adjacent non-tumor tissues (Fig.
2F). The association between miR-150-5p expression and OS of
patients with breast cancer was analyzed using the Kaplan-Meier
plotter database. Based on the median miR-150 expression, the
patients were divided into high and low expression groups. The OS
time of patients with high miR-150-5p expression was significantly
longer compared with that of patients with low miR-150-5p
expression (Fig. 2G). In
conclusion, these results demonstrated that MAFG-AS1 interacted
with miR-150-5p in breast cancer, and that high expression levels
of miR-150-5p was associated with a favorable prognosis in patients
with breast cancer.
MAFG-AS1/miR-150-5p regulates the
viability and migration of breast cancer cells
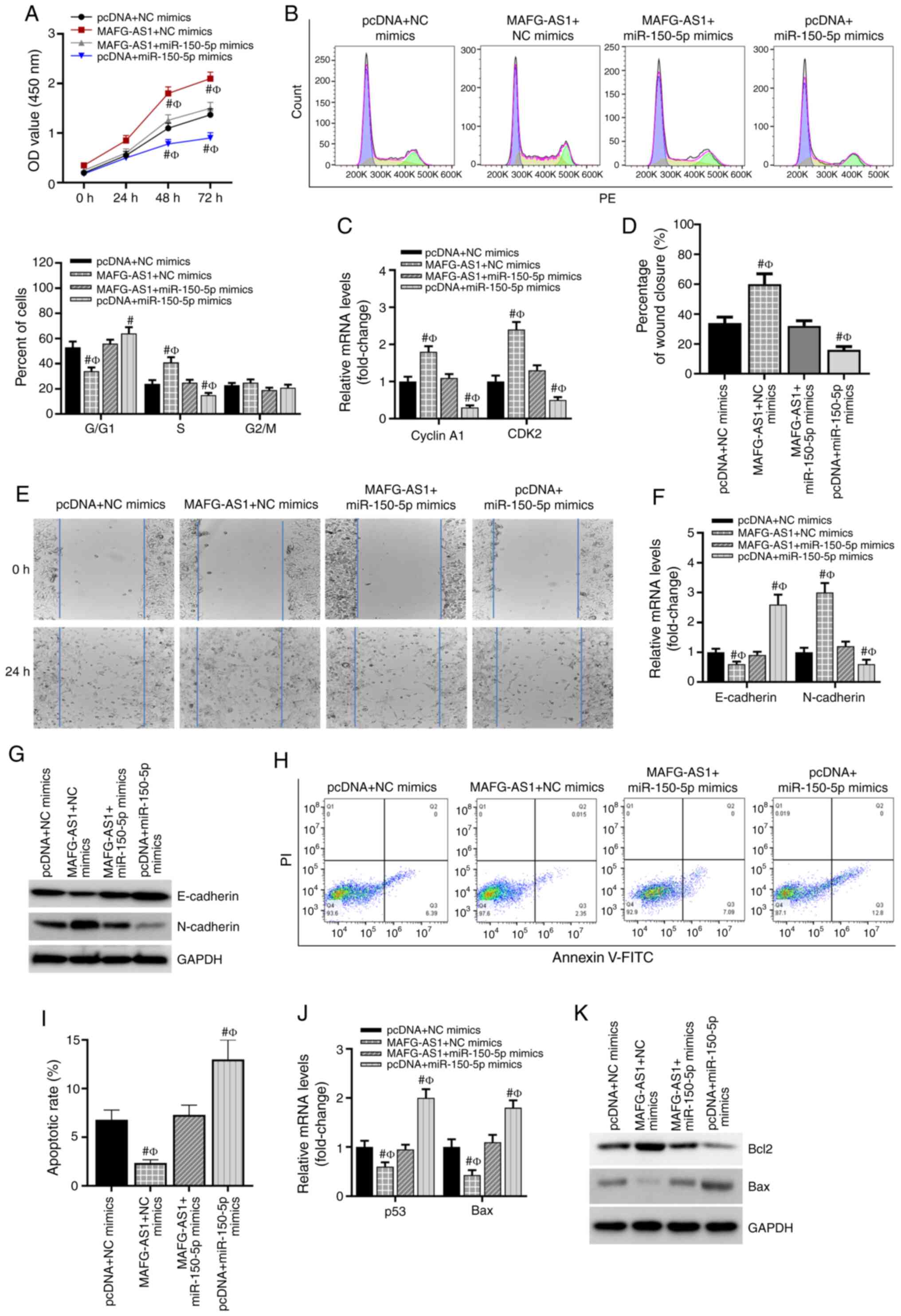
The biological role of the MAFG-AS1/miR-150-5p axis
in the development of breast cancer was further investigated. As
presented in Fig. 3A, compared
with that in the pcDNA + NC mimics group, overex-pression of
MAFG-AS1 significantly promoted the viability of breast cancer
cells, whereas the miR-150-5p mimics reduced cell viability after
48 h. In addition, the results demonstrated that co-transfection
with the miR-150-5p mimics significantly abolished the increase in
cell viability induced by MAFG-AS1 overexpression. The cell cycle
analysis demonstrated that MCF-7 cells accumulated in the S phase
and were decreased in the G1/G0 phase after pcDNA-MAFG-AS1
transfection, whereas the miR-150-5p mimics induced cell cycle
arrest at the G1/G0 phase. However, the miR-150-5p mimics failed to
induce G1/G0 arrest in pcDNA-MAFG-AS1 co-transfected MCF-7 cells
(Fig. 3B). RT-qPCR was conducted
to determine the expression of the cell cycle-associated genes
cyclin A1 and CDK2. These results demonstrated that the expression
levels of cyclin A1 and CDK2 were significantly upregulated in
MCF-7 cells transfected with the pcDNA-MAFG-AS1 vector, but
downregulated by the miR-150-5p mimics compared with the pcDNA + NC
mimics group (Fig. 3C). A wound
healing assay was conducted to assess the migratory ability of
breast cancer cells. The results revealed that transfection with
the miR-150-5p mimics inhibited, whereas overexpression of MAFG-AS1
promoted cell migration compared with that in the control group; in
addition, the promotive effect of MAFG-AS1 overexpression on cell
migration was attenuated by the miR-150-5p mimics (Fig. 3D and E). EMT-related gene
expression was subsequently assessed. The mRNA and protein
expression levels of E-cadherin were significantly downregulated,
and those of N-cadherin were upregulated in breast cancer cells
transfected with pcDNA-MAFG-AS1 vectors compared with those in the
control cells, whereas co-transfection with the miR-150-5p mimics
reversed the effects of MAFG-AS1 on the expression of these markers
(Fig. 3F and G). Additionally,
overexpression of MAFG-AS1 decreased the apoptotic rates of breast
cancer cells as measured by flow cytometry with PI and Annexin V
staining, whereas the numbers of apoptotic cells were markedly
increased by the miR-150-5p mimics (Fig. 3 H and I). The inhibitory role of
MAFG-AS1 overexpression on apoptosis was reversed by the miR-150-5p
mimics (Fig. 3H and I). In
addition, the mRNA and protein levels of the apoptosis-related gene
Bcl-2 was significantly upregulated, whereas those of Bax was
significantly downregulated in breast cancer cells transfected with
pcDNA-MAFG-AS1 vectors compared with those in the control cells
(Fig. 3J and K). By contrast, the
miR-150-5p mimics decreased the mRNA and protein levels of Bcl-2,
but increased those of Bax; however, the miR-150-5p mimics failed
to induce Bcl-2 downregulation and Bax upregulation in MAFG-AS1
vector co-transfected MCF-7 cells (Fig. 3J and K). These results suggested
that the MAFG-AS1/miR-150-5p axis may regulate the viability,
apoptosis and migration of breast cancer cells.
MYB is a functional target gene of
miR-150-5p
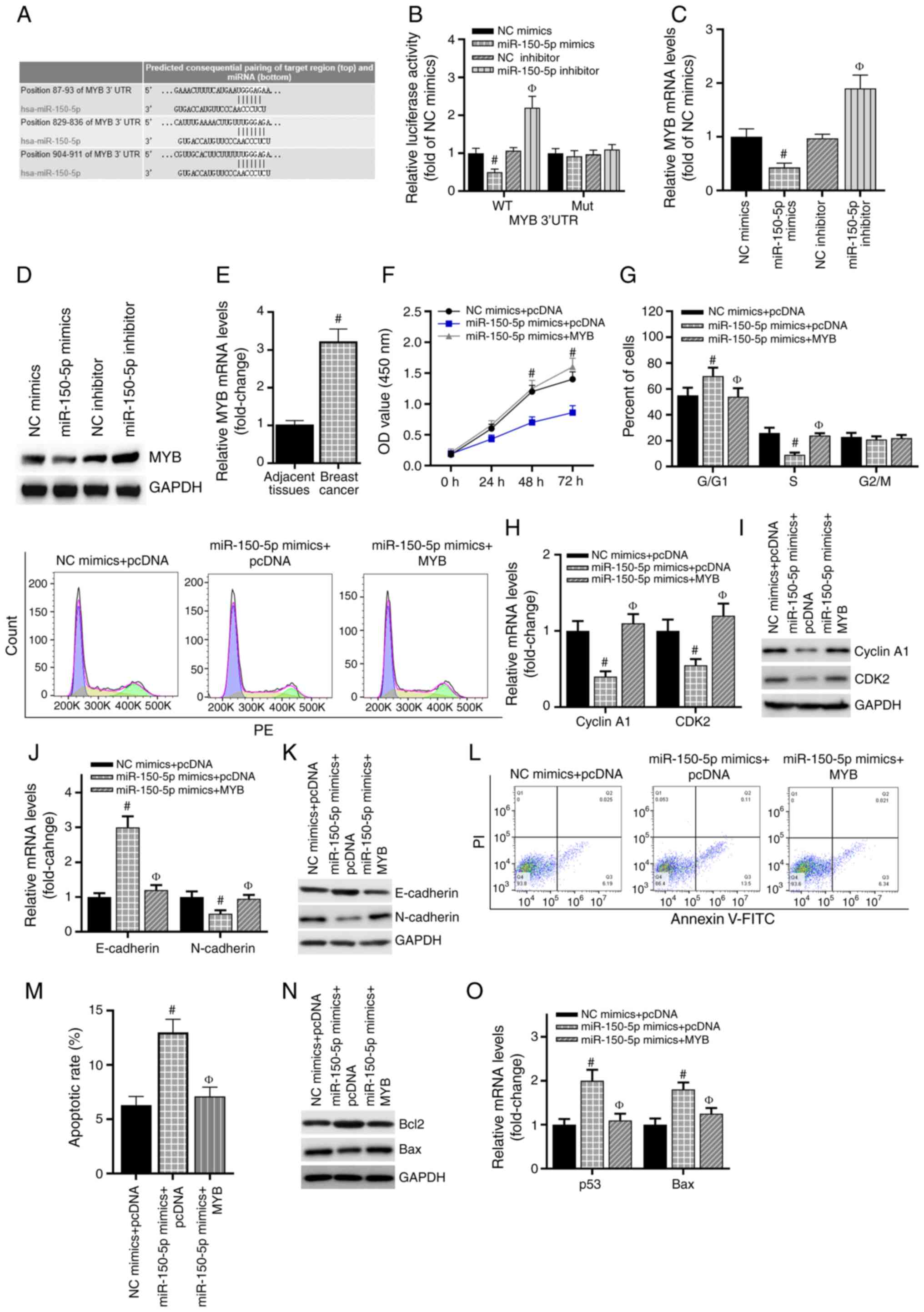
Subsequently, the downstream target of miR-150-5p
was explored. Three online algorithms (miRDB, TargetScan and
miRanda) were used to predict the potential target genes of
miR-150-5p. The results revealed that MYB was a candidate target
gene of miR-150-5p (Fig. 4A). The
miR-150-5p inhibitor was used to knock down miR-150-5p expression,
and the knockdown efficiency was validated by RT-qPCR (Fig. S1C). As presented in Fig. 4B, the luciferase activity was
significantly weakened by the miR-150-5p mimics compared with the
NC mimics, but exhibited no changes when the 3′UTR binding sites
were mutated. Co-transfection of the miR-150-5p inhibitor and the
WT-MYB 3′UTR plasmid resulted in a significant increase in the
luciferase signal. Additionally, MYB mRNA levels were downregulated
following transfection with the miR-150-5p mimics, but upregulated
when miR-150-5p expression was inhibited (Fig. 4C). Similar results were obtained
for MYB protein expression by western blotting (Fig. 4D). In addition, compared with
adjacent normal breast tissues, MYB mRNA expression was upregulated
in breast cancer tissues (Fig.
4E). For further experiments, a MYB overexpression vector was
used, and the transfection efficiency of MYB was verified by
RT-qPCR (Fig. S1D). The MTS assay
demonstrated that MYB overexpression reversed the suppressive
effect of the miR-150-5p mimics on MCF-7 cell viability (Fig. 4F). In addition, overexpression of
MYB inhibited the miR-150-5p-induced cell cycle arrest (Fig. 4G). The miR-150-5p mimics
significantly inhibited cyclin A1, CDK2 and N-cadherin mRNA and
protein expression levels and increased the expression levels of
E-cadherin compared with those in the control cells; following
co-transfection with the MYB overexpression vector, these results
were reversed (Fig. 4H-K).
Additionally, the miR-150-5p mimics promoted apoptosis in breast
cancer cells, whereas the number of apoptotic cells was recovered
following MYB overexpression (Fig. 4L
and M). The decreases in Bcl-2 and increases in Bax expression
levels induced by the miR-150-5p mimics were reversed by MYB
overexpression (Fig. 4N and O).
Overall, these results demonstrated that MYB was a functional
target gene of miR-150-5p in breast cancer.
MAFG-AS1 positively regulates the
expression of the MYB gene by sponging miR-150-5p
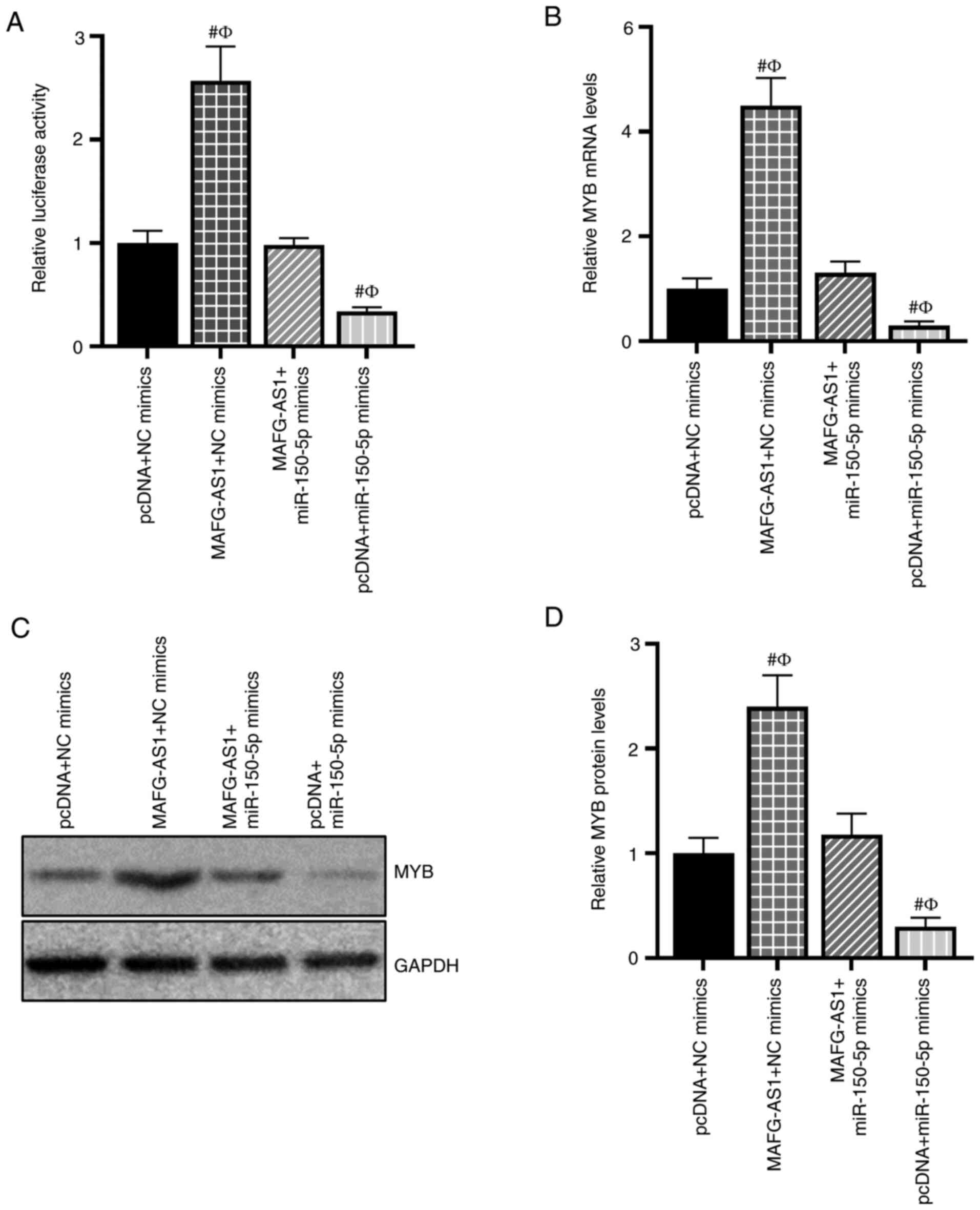
To confirm the relationship between
MAFG-AS1/miR-150-5p and MYB expression, a luciferase reporter assay
was performed. As presented in Fig.
5A, the MYB-WT luciferase activity was significantly increased
by MAFG-AS1 overexpression and decreased by the miR-150-5p mimics.
However, induction of luciferase activity of MYB-WT was abrogated
following co-transfection with the MAFG-AS1 overexpression vector
and the miR-150-5p mimics. The effects of MAFG-AS1/miR150-5p on MYB
mRNA and protein expression were also assessed; RT-qPCR and western
blotting results demonstrated that MAFG-AS1 overexpression
upregulated the levels of MYB mRNA and protein expression, whereas
transfection with the miR-150-5p mimics downregulated MYB mRNA and
protein expression levels. Furthermore, MAFG-AS1 overexpression
attenuated the inhibitory effects of the miR-150-5p mimics on the
mRNA and protein expression of MYB (Fig. 5B-D). These results indicated that
MAFG-AS1 competitively bound miR-150-5p to upregulate the
expression of MYB.
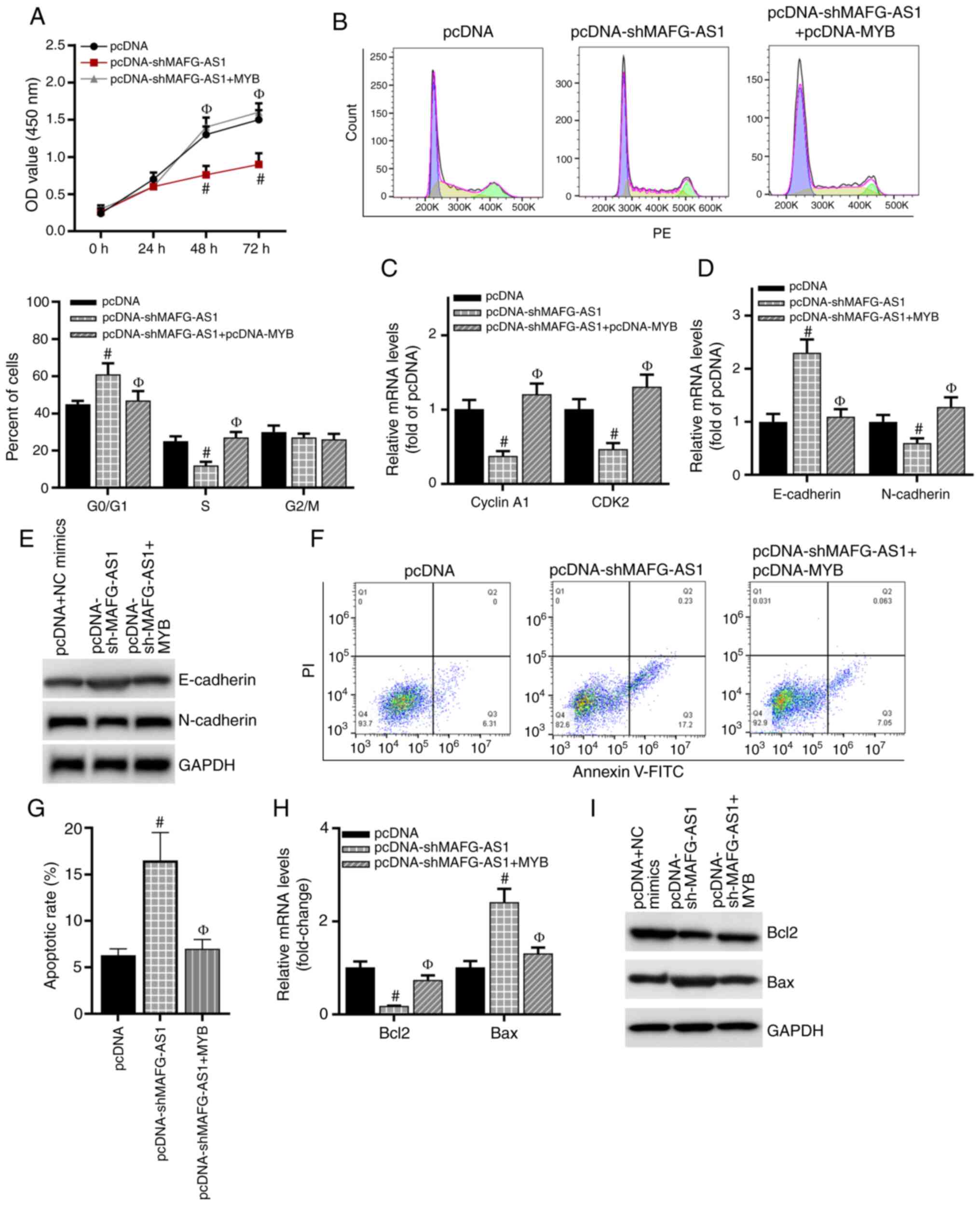
MYB is a downstream molecule of MAFG-AS1
in the regulation of cell biological functions
To further validate the effects of MYB on the role
of MAFG-AS1 in breast cancer cells, MAFG-AS1 expression was knocked
down by transfection of shMAFG-AS1 into MCF-7 cells, and the
knockdown efficiency of MAFG-AS1 was validated by RT-qPCR analysis
(Fig. S1E). The MTS assay results
demonstrated that compared with that in the pcDNA group, cell
proliferation was significantly inhibited after silencing MAFG-AS1
expression but restored following co-transfection with the MYB
overexpression vector (Fig. 6A).
In addition, knockdown of MAFG-AS1 induced breast cancer cell cycle
arrest in the G0/G1 phase and reduced the number of cells in the S
phase compared with that in the cells transfected with the control
vector. The effects of shMAFG-AS1 were reversed following
co-transfection with the MYB over-expression vector (Fig. 6B). The expression levels of cyclin
A1 and CDK2 were downregulated after silencing MAFG-AS1 compared
with those in the control cells, whereas overexpression of MYB
restored their expression (Fig.
6C). Similarly, the expression levels of the EMT-related
protein E-cadherin were upregulated and those of N-cadherin were
downregulated in the shMAFG-AS1 group compared with those in the
control group, and overexpression of MYB restored their expression
(Fig. 6D and E). As presented in
Fig. 6F and G, compared with that
in the control group, the apoptotic rate was increased in the
shMAFG-AS1 group, but decreased following co-transfection with the
MYB overexpression vector. The mRNA and protein expression levels
of Bcl-2 were downregulated, and those of Bax were upregulated in
breast cancer cells transfected with shMAFG-AS1 compared with those
in cells transfected with the control vector, whereas MYB
overexpression increased the levels of Bcl-2 and inhibited the
levels of Bax compared with those in the shMAFG-AS1 group (Fig. 6H and I). Overall, these results
demonstrated that MAFG-AS1 may aggravate the progression of breast
cancer by upregulating MYB.
MAFG-AS1 regulates the progression of
breast cancer in vivo by targeting the miR-150-5p/MYB axis
The present study further investigated whether
MAFG-AS1 regulated breast cancer tumor growth in vivo by
sponging miR-150-5p. As presented in Fig. S2A, MAFG-AS1 overexpression
increased the xenograft tumor size, whereas the miR-150-5p mimics
reduced the tumor size in vivo; the miR-150-5p mimics
abolished the MAFG-AS1 overexpression-mediated increase in tumor
size. In addition, the elevated tumor volume and tumor weight
induced by MAFG-AS1 overexpression were abrogated by the miR-150-5p
mimics (Fig. 2B and C). By
contrast, the tumor size, volume and weight suppressed by the
miR-150-5p mimics were recovered by MYB overexpression (Fig. S2D-F). These results suggested that
MAFG-AS1 regulated the progression of breast cancer in vivo
by targeting the miR-150-5p/MYB axis.
Discussion
Breast cancer is a major health threat to women
worldwide (1,2). Although a number of studies have
investigated the cellular and molecular mechanisms underlying the
occurrence and development of breast cancer (31-33),
early detection is hindered by the lack of effective diagnostic
biomarkers. Numerous studies have indicated that lncRNAs may be
used as potential therapeutic targets or prognostic markers of
breast cancer (34). For instance,
lncRNA SNHG14 induces breast cancer cell chemoresistance to
trastuzumab by regulating poly(A)-binding protein cytoplasmic 1
through trimethylation of lysine 27 acetylation (35). In addition, lncRNA integrin subunit
β2 (ITGB2)-AS1 promotes the migration and invasion of breast cancer
cells through upregulating ITGB2 (36). However, the role of MAFG-AS1 in
breast cancer has not been explored. The results of the present
study demonstrated that the expression of MAFG-AS1 in breast cancer
tissues and cells was increased compared with that in non-cancerous
tissues and cells. These results indicated that MAFG-AS1 had the
potential to become a biomarker for breast cancer.
The canonical theory for the mechanism of lncRNAs is
that lncRNA serve as miRNA 'sponges' to antagonize the inhibition
of miRNAs on mRNAs, thus enabling the expression of downstream
target genes (15). For example,
the lncRNA growth arrest-specific 5 regulates autophagy in patients
with breast cancer by sponging miR-23a to upregulate autophagy
related 3 homolog (37). lncRNA
bladder cancer-associated transcript 1 induces breast cancer cell
proliferative, migratory and invasive abilities by regulating C-C
chemokine receptor type 2 expression by binding to the specific
sequence of miR-150-5p (15). The
androgen receptor negatively induced lncRNA binds to miR-204 to
increase the migration and invasion of triple-negative breast
cancer cells (38). miRNAs serve
important regulatory roles in physiological activities and
pathological processes of breast cancer. miR-1287-5p suppresses the
growth of triple negative breast cancer growth via interaction with
phosphoinositide 3-kinase CB (39). miR-132-3p inhibits the migration
and invasion of breast cells by targeting lysosomal-associated
protein transmembrane 4 beta (40). The results of the present study
confirmed that miR-150-5p was sponged by MAFG-AS1 in breast cancer
cells. In addition, miR-150-5p was downregulated in breast cancer
cells and tissues compared with normal adjacent tissues and normal
breast epithelial cells. The miR-150-5p mimics blocked the
progression of breast cancer and abolished the promotive role of
MAFG-AS1 overexpression on the progression of breast cancer.
Additionally, the in vivo experiment results demonstrated
that the tumor size, volume and weight increased by MAFG-AS1
overexpression were abrogated by the miR-150-5p mimics. These
results demonstrated that MAFG-AS1 promoted the progression of
breast cancer by sponging miR-150-5p in vitro and in
vivo.
Bioinformatics analysis in the present study
demonstrated that the oncogene MYB was a candidate target gene for
miR-150-5p. Previous studies have demonstrated that MYB exerts a
cancer-promoting effect in a variety of tumors, including breast
cancer (41-44). However, the molecular mechanism of
how the oncogenic driver MYB affects breast cancer progression has
not yet been fully elucidated. In the present study, the luciferase
reporter assay confirmed that miR-150-5p directly targeted the
3′UTR of MYB. In addition, the miR-150-5p mimics significantly
inhibited the expression of MYB in MCF-7 cells. The miR-150-5p
mimics also inhibited breast cancer tumor growth in vivo by
suppressing MYB expression. In addition, MYB overexpression
abolished the miR-150-5p mimic-mediated inhibition of tumor size,
volume and weight in vivo. Further experiments verified that
MYB may be an important molecule in the MAFG-AS1-mediated
regulation of breast cancer cell biological function.
In conclusion, the results of the present study
revealed a crucial role of lncRNA MAFG-AS1 in the proliferation and
migration of breast cancer cells. MAFG-AS1 may promote human breast
cancer tumorigenesis by targeting the miR-150-5p/MYB axis,
suggesting a novel molecular mechanism of breast cancer.
Supplementary Data
Acknowledgments
Not applicable.
Funding
This work was supported by the Jilin Provincial
Department of Science and Technology (grant no. 20190201209JC).
Availability of data and materials
All data generated or analyzed in this study are
available from the corresponding author on reasonable request.
Authors' contributions
HJ and SL conceived and designed the study. HJ, DW
and ZZ performed the experiments. HJ, DW and SL analyzed the data.
HJ and SL wrote the manuscript. SL edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The clinical study was approved by the Ethics
Committee of the First Hospital of Jilin University (approval no.
DWLL-2019-0021), and written informed consent was obtained from all
patients. All relevant methods were performed according to the
relevant guidelines and regulations. The animal study was approved
with the ethical approval from the Ethics Committee of Animal
Experiments of the First Hospital of Jilin University (approval no.
DWLL-2019-0021).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Colombet M, Soerjomataram I,
Mathers C, Parkin DM, Piñeros M, Znaor A and Bray F: Estimating the
global cancer incidence and mortality in 2018: GLOBOCAN sources and
methods. Int J Cancer. 144:1941–1953. 2019. View Article : Google Scholar
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Feng RM, Zong YN, Cao SM and Xu RH:
Current cancer situation in China: Good or bad news from the 2018
global cancer statistics? Cancer Commun (Lond). 39:222019.
View Article : Google Scholar
|
|
4
|
de la Mare JA, Contu L, Hunter MC, Moyo B,
Sterrenberg JN, Dhanani KC, Mutsvunguma LZ and Edkins AL: Breast
cancer: Current developments in molecular approaches to diagnosis
and treatment. Recent Pat Anticancer Drug Discov. 9:153–175. 2014.
View Article : Google Scholar
|
|
5
|
Shi X, Sun M, Liu H, Yao Y and Song Y:
Long non-coding RNAs: A new frontier in the study of human
diseases. Cancer Lett. 339:159–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tsai MC, Spitale RC and Chang HY: Long
intergenic noncoding RNAs: New links in cancer progression. Cancer
Res. 71:3–7. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen X, Dai M, Zhu H, Li J, Huang Z, Liu
X, Huang Y, Chen J and Dai S: Evaluation on the diagnostic and
prognostic values of long non-coding RNA BLACAT1 in common types of
human cancer. Mol Cancer. 16:1602017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dai M, Chen X, Mo S, Li J, Huang Z, Huang
S, Xu J, He B, Zou Y, Chen J and Dai S: Meta-signature LncRNAs
serve as novel biomarkers for colorectal cancer: Integrated
bioinformatics analysis, experimental validation and diagnostic
evaluation. Sci Rep. 7:465722017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ye JR, Liu L and Zheng N: Long noncoding
RNA bladder cancer associated transcript 1 promotes the
proliferation, migration, and invasion of nonsmall cell lung cancer
through sponging miR-144. DNA Cell Biol. 36:845–852. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rinn JL, Kertesz M, Wang JK, Squazzo SL,
Xu X, Brugmann SA, Goodnough LH, Helms JA, Farnham PJ, Segal E and
Chang HY: Functional demarcation of active and silent chromatin
domains in human HOX loci by noncoding RNAs. Cell. 129:1311–1323.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang HW, Xie H, Ma X, Zhao F and Gao Y:
Upregulation of LncRNA PANDAR predicts poor prognosis and promotes
cell proliferation in cervical cancer. Eur Rev Med Pharmacol Sci.
21:4529–4535. 2017.PubMed/NCBI
|
|
13
|
Zang W, Wang T, Huang J, Li M, Wang Y, Du
Y, Chen X and Zhao G: Long noncoding RNA PEG10 regulates
proliferation and invasion of esophageal cancer cells. Cancer Gene
Ther. 22:138–144. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ding X, Zhang S, Li X, Feng C, Huang Q,
Wang S, Wang S, Xia W, Yang F, Yin R, et al: Profiling expression
of coding genes, long noncoding RNA, and circular RNA in lung
adenocarcinoma by ribosomal RNA-depleted RNA sequencing. FEBS Open
Bio. 8:544–555. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu X, Liu Y, Du Y, Cheng T and Xia W: Long
non-coding RNA BLACAT1 promotes breast cancer cell proliferation
and metastasis by miR-150-5p/CCR2. Cell Biosci. 9:142019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jia YC, Wang JY, Liu YY, Li B, Guo H and
Zang AM: LncRNA MAFG-AS1 facilitates the migration and invasion of
NSCLC cell via sponging miR-339-5p from MMP15. Cell Biol Int.
43:384–393. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cui S, Yang X, Zhang L, Zhao Y and Yan W:
LncRNA MAFG-AS1 promotes the progression of colorectal cancer by
sponging miR-147b and activation of NDUFA4. Biochem Biophys Res
Commun. 506:251–258. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li H, Zhang GY, Pan CH, Zhang XY and Su
XY: LncRNA MAFG-AS1 promotes the aggressiveness of breast carcinoma
through regulating miR-339-5p/MMP15. Eur Rev Med Pharmacol Sci.
23:2838–2846. 2019.PubMed/NCBI
|
|
19
|
Wang JS, Liu QH, Cheng XH, Zhang WY and
Jin YC: The long noncoding RNA ZFAS1 facilitates bladder cancer
tumorigenesis by sponging miR-329. Biomed Pharmacother.
103:174–181. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu S, Sun Z, Tang L and Chen L: LINC00355
promotes tumor progression in HNSCC by hindering
MicroRNA-195-mediated suppression of HOXA10 expression. Mol Ther
Nucleic Acids. 19:61–71. 2020. View Article : Google Scholar
|
|
21
|
Xiao H: MiR-7-5p suppresses tumor
metastasis of non-small cell lung cancer by targeting NOVA2. Cell
Mol Biol Lett. 24:602019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Q, Zheng J and Liu L: The long
noncoding RNA PCGEM1 promotes cell proliferation, migration and
invasion via targeting the miR-182/FBXW11 axis in cervical cancer.
Cancer Cell Int. 19:3042019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen X, Xu X, Pan B, Zeng K, Xu M, Liu X,
He B, Pan Y, Sun H and Wang S: miR-150-5p suppresses tumor
progression by targeting VEGFA in colorectal cancer. Aging (Albany
NY). 10:3421–3437. 2018. View Article : Google Scholar
|
|
24
|
Lu W, Zhang H, Niu Y, Wu Y, Sun W, Li H,
Kong J, Ding K, Shen HM, Wu H, et al: Long non-coding RNA linc00673
regulated non-small cell lung cancer proliferation, migration,
invasion and epithelial mesenchymal transition by sponging
miR-150-5p. Mol Cancer. 16:1182017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sakr M, Takino T, Sabit H, Nakada M, Li Z
and Sato H: miR-150-5p and miR-133a suppress glioma cell
proliferation and migration through targeting membrane-type-1
matrix metallo-proteinase. Gene. 587:155–162. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014. View Article : Google Scholar
|
|
29
|
Sen R, Ghosal S, Das S, Balti S and
Chakrabarti J: Competing endogenous RNA: The key to
posttranscriptional regulation. ScientificWorldJournal.
2014:8962062014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Khatib A, Solaimuthu B, Yosef MB, Rmaileh
AA, Tanna M, Oren G, Frisch MS, Axelrod JH, Lichtenstein M and
Shaul YD: The glutathione peroxidase 8 (GPX8)/IL-6/STAT3 axis is
essential in maintaining an aggressive breast cancer phenotype.
Proc Natl Acad Sci USA. 117:21420–21431. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cataldo A, Romero-Cordoba S, Plantamura I,
Cosentino G, Hidalgo-Miranda A, Tagliabue E and Iorio MV: MiR-302b
as a combinatorial therapeutic approach to improve cisplatin
chemotherapy efficacy in human triple-negative breast cancer.
Cancers (Basel). 12:22612020. View Article : Google Scholar
|
|
33
|
Naik SK, Lam EWF, Parija M, Prakash S,
Jiramongkol Y, Adhya AK, Parida DK and Mishra SK: NEDDylation
negatively regulates ERRβ expression to promote breast cancer
tumorigenesis and progression. Cell Death Dis. 11:7032020.
View Article : Google Scholar
|
|
34
|
Rodríguez Bautista R, Ortega Gómez A,
Hidalgo Miranda A, Zentella Dehesa A, Villarreal-Garza C,
Ávila-Moreno F and Arrieta O: Long non-coding RNAs: Implications in
targeted diagnoses, prognosis, and improved therapeutic strategies
in human non- and triple-negative breast cancer. Clin Epigenetics.
10:882018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dong H, Wang W, Mo S, Liu Q, Chen X, Chen
R, Zhang Y, Zou K, Ye M, He X, et al: Long non-coding RNA SNHG14
induces trastuzumab resistance of breast cancer via regulating
PABPC1 expression through H3K27 acetylation. J Cell Mol Med.
22:4935–4947. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu M, Gou L, Xia J, Wan Q, Jiang Y, Sun
S, Tang M, He T and Zhang Y: LncRNA ITGB2-AS1 could promote the
migration and invasion of breast cancer cells through up-regulating
ITGB2. Int J Mol Sci. 19:18662018. View Article : Google Scholar :
|
|
37
|
Gu J, Wang Y, Wang X, Zhou D, Wang X, Zhou
M and He Z: Effect of the LncRNA GAS5-MiR-23a-ATG3 axis in
regulating autophagy in patients with breast cancer. Cell Physiol
Biochem. 48:194–207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang F, Shen Y, Zhang W, Jin J, Huang D,
Fang H, Ji W, Shi Y, Tang L, Chen W, et al: An androgen receptor
negatively induced long non-coding RNA ARNILA binding to miR-204
promotes the invasion and metastasis of triple-negative breast
cancer. Cell Death Differ. 25:2209–2220. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schwarzenbacher D, Klec C, Pasculli B,
Cerk S, Rinner B, Karbiener M, Ivan C, Barbano R, Ling H,
Wulf-Goldenberg A, et al: MiR-1287-5p inhibits triple negative
breast cancer growth by interaction with phosphoinositide 3-kinase
CB, thereby sensitizing cells for PI3Kinase inhibitors. Breast
Cancer Res. 21:202019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li S, Xu JJ and Zhang QY: MicroRNA-132-3p
inhibits tumor malignant progression by regulating
lysosomal-associated protein transmembrane 4 beta in breast cancer.
Cancer Sci. 110:3098–3109. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rettig EM, Tan M, Ling S, Yonescu R,
Bishop JA, Fakhry C and Ha PK: MYB rearrangement and
clinicopathologic characteristics in head and neck adenoid cystic
carcinoma. Laryngoscope. 125:E292–E299. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
West RB, Kong C, Clarke N, Gilks T,
Lipsick JS, Cao H, Kwok S, Montgomery KD, Varma S and Le QT: MYB
expression and translocation in adenoid cystic carcinomas and other
salivary gland tumors with clinicopathologic correlation. Am J Surg
Pathol. 35:92–99. 2011. View Article : Google Scholar :
|
|
43
|
Andersson MK, Afshari MK, Andrén Y, Wick
MJ and Stenman G: Targeting the oncogenic transcriptional regulator
MYB in adenoid cystic carcinoma by inhibition of IGF1R/AKT
signaling. J Natl Cancer Inst. 109:2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mitra P: Transcription regulation of MYB:
A potential and novel therapeutic target in cancer. Ann Transl Med.
6:4432018. View Article : Google Scholar :
|