In 1967, Peter Wolf first discovered that platelets
can release numerous vesicles. At that time, researchers believed
these vesicles were simply cell fragments with no relevant
biological function (1). In 1983,
exosomes were first observed in sheep reticulocytes (2). Johnstone named these vesicles
'exosomes' in 1987 and defined them as vesicles with a diameter of
between 40 and 100 nm (3). In
2013, the Nobel Prize in physiology or medicine was awarded to
James E. Rothman, Randy W. Schekman and Thomas C. Südhof for their
study of extracellular vesicles (EVs) (4), which resulted in this evolving into
a highly studied field.
Exosomes are EVs that originate from the endosome
system and are important carriers of cell-to-cell communication in
the microenvironment (5). Tumor
cells can interact with immune cells, mesenchymal cells and
endothelial cells in the microenvironment to promote tumor
progression (6). Tumor exosomes
can carry biological information in the form of proteins, lipids
and nucleic acids. These molecules can domesticate recipient cells
and may become tumor-specific markers (7). Therefore, exosomes and cells have
significant interactions.
In clinical applications, circulating EVs derived
from cancer patients are associated with tumor metastasis or
recurrence; therefore, they can be used as important diagnostic and
prognostic indicators, and therapeutic targets (8). In addition, exosomes, as natural
nanoparticles, have great advantages over artificial materials in
encapsulating chemotherapy drugs, such as paclitaxel (9). However, we do not have a complete
understanding of EVs, which hinders the application of them as
clinical treatments or diagnostic methods (10). Kalluri and LeBleu (11) stated that animal models with which
biogenesis, trafficking and cellular entry of exosomes can be
studied should be investigated rapidly. The study also summarized
the challenges affecting the exosome field, such as improving the
exosome isolation method, the influence of the administration route
for exosome uptake on potential therapeutic strategies, and
combining exosomal RNA, protein and metabolites to enhance the
specificity and sensitivity of exosome-based diagnostics.
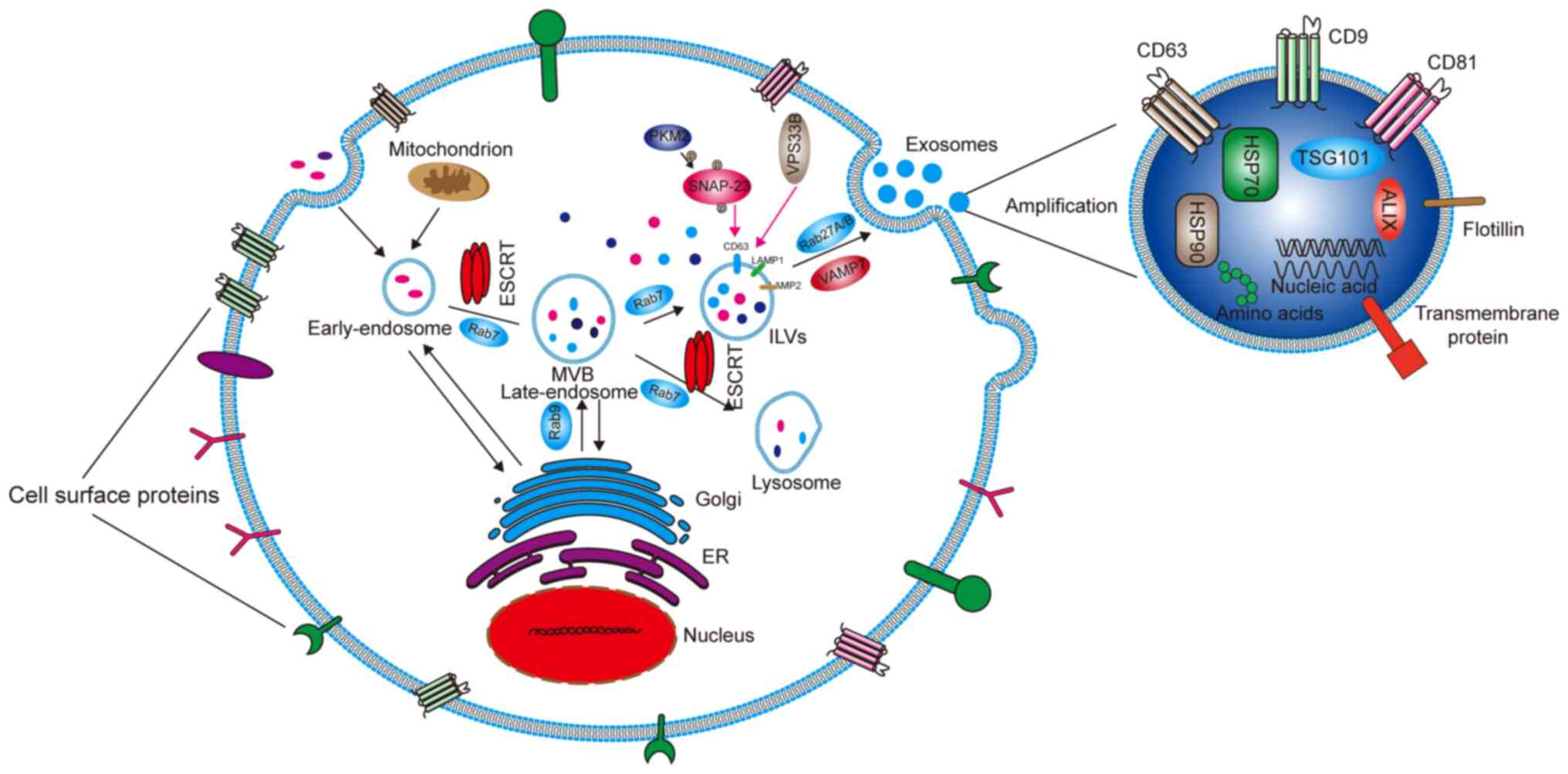
Cells capture extracellular materials via
endocytosis and then form early endosomes (Fig. 1). During the process of transition
to a late endosome, the endocytic body sprouts inside to envelop
specific proteins and nucleic acids to form intraluminal vesicles
(ILVs). Advanced endosomes containing multiple ILVs are known as
multivesicular bodies (MVBs) (12). Most MVBs will be digested by
lysosomes in the cells. Only a few vesicles containing cluster of
differentiation (CD)63, recombinant lysosomal-associated membrane
protein (LAMP)1 and LAMP2 on the surface can fuse with the cell
membrane and mediate the secretion of exosomes (13). ILVs and MVBs act as pre-exosomes
and are formed primarily with the assistance of endosomal sorting
complexes required for transport (ESCRT) protein complexes, which
are mainly composed of ESCRT-0, I, II and III, as well as auxiliary
proteins vacuolar protein sorting-associated protein 4 (VPS4),
vacuolar protein sorting-associated protein 1 (VTA1) and
apoptosis-linked gene 2 interacting protein X (ALIX). The main
function of ESCRT is to sort specific substances into ILVs. ESCRT-0
contains a hepatocyte growth factor-regulated tyrosine kinase
substrate (HRS) that recognizes ubiquitinated proteins and
interacts with signal-transducing adaptor molecules (14). HRS recruits ESCRT-I through tumor
susceptibility gene 101 protein, and then ESCRT-I recruits
ESCRT-III through ESCRT-II and ALIX (15). ESCRT-III forms exosomes by cutting
MVBs (16), These exosomes are
then released from the cells by fusion with the cell membrane.
Rabphilin (RAB)27A/B, a member of the RAB family, is the main
motion controller of exosomes on the cytoskeleton (17) Additionally, vesicle-associated
membrane protein 7 of the Soluble N-etylmaleimide-sensitive fusion
protein attachment protein receptor proteins family plays the main
role in promoting the fusion between exosomes and cell membranes to
secrete exosomes (18). Some
studies have found that Dicer and Argonaute 2, the key components
of miRNA processing, are functionally present in exosomes. Although
it has yet to be confirmed, these findings suggest that exosomes
are not only passive vehicles but also miRNA factories (19,20). Wang et al (21) reported that chemotherapeutic
agents can stimulate the production or release of exosomes. Wei
et al (22) found that
pyruvate kinase M2, a key enzyme in aerobic glycolysis (known as
the Warburg effect) of tumor cells, can promote the phosphorylation
of synaptosomal-associated protein 23 and the subsequent release of
exosomes. VPS33B is also a protein that regulates the release of
exosomes, the deficiency of which will cause maturation and
secretion disorder of exosomes (23).
Exosomes were thought of as cellular metabolism
waste products for a long time. With an increasing number of
studies on exosomes, researchers have discovered that exosomes are
important in cellular communication (24-26). Moreover, exosomes can affect tumor
growth, angiogenesis, invasion and metastasis (27,28). Exosomes can promote the formation
of the tumor microenvironment (TME) (29), which is comprised of tumor blood
vessels, the extracellular matrix (ECM) and other non-malignant
cells such as stromal cells, fibroblasts and inflammatory cells
(30,31). Therefore, therapies that target
the TME may be an effective treatment for cancer (32,33). Differing from that in normal
cells, the metabolism of tumor cells relies on aerobic glycolysis
even under normoxic conditions. This can generate more lactic acid
and thereby lower the pH of the TME (34,35).
Exosomes can also influence the proliferation and
differentiation of cells in the TME. Osteosarcoma-derived exosomes
can promote osteoclast differentiation and bone resorption
activity. In glioblastoma, exosomes derived from cancer cells that
contain CD171 can promote glioma cell invasiveness, motility and
proliferation (36). In breast
cancer, MDA-MB-231 cell-derived exosomes can transfer miR-20a-5p to
bone marrow macrophages and promote the proliferation and
differentiation of osteoclasts by targeting SRC kinase signaling
inhibitor 1 (37). Exosomes
derived from mesenchymal stem cell (MSC)-differentiated adipocytes
are actively taken up by breast cancer MCF7 cells. This
subsequently promotes MCF7 cell proliferation and migration, as
well as protecting the cells from serum deprivation or
chemotherapeutic drug-induced apoptosis in vitro (38). Cancer cell-derived exosomes can
promote differentiation of healthy fibroblasts or MSCs into
cancer-associated fibroblasts (CAFs), which in turn activate cancer
cell invasion and migration (39,40). Cancer cell loss of tumor protein
p53 can result in adrenergic trans-differentiation of
tumor-associated sensory nerves by EVs (41).
Cancer cell resistance to chemotherapeutics and
monoclonal antibodies is one of the difficult issues affecting the
successful treatment of patients. The mechanism behind this
resistance is quite complicated (Table I).
In experimental studies on exosomes, it was found
that drug-resistant tumor cells can transfer their drug resistance
to drug-sensitive cells through exosomes. For example, the highly
tumorigenic and drug-resistant cell subpopulations in glioblastoma
multiforme (GBM) can regulate other subpopulations through
microRNAs (miRNAs/miRs) in EVs (42). Long non-coding (lnc)RNA regulator
of Akt signaling associated with hepatocellular carcinoma and renal
cell carcinoma exosomes can competitively bind miR-34/miR-449 to
increase the expression of lipoxin A4 receptor and
cellular-mesenchymal epithelial transition factor in RCC cells,
regulating the resistance of these cells to sunitinib (43). Vemurafenib can induce expression
of miR-211-5p in melanoma cells, while cells overexpressing this
miRNA will deliver miR-211-5p to other melanoma cells through
exosomes, and thereby enhance drug resistance (44). Gemcitabine promotes the expression
of miR-155 in exosomes secreted by pancreatic ductal adenocarcinoma
(PDAC) cells and can deliver it to other PDAC cells to increase
their drug resistance (45).
Another study reported that exosomes derived from ontogenically
transformed, mesenchymal human bronchial epithelial cells (HBECs)
could transfer chemoresistance to the parental, epithelial HBECs
and increase zinc finger E-box binding homeobox 1 mRNA, a master
epithelial-mesenchymal transition (EMT) transcription factor, in
the recipient cells (46).
Moreover, exosomes secreted from stromal cells in
the TME can also activate tumor-related pathways to enhance tumor
cell resistance to drugs. The decreased sensitivity of breast
cancer to anthracycline chemotherapy may be from exosomes secreted
by peripheral cells promoting the expression of miR-125b in breast
cancer cells (BCCs) (47). EVs
from bone marrow stromal cells can enhance the resistance of
chronic lymphocytic leukemia B cells to several chemotherapy drugs,
such as fludarabine, ibrutinib, idelaisib and venetoclax (48). Exosomal miR-92a-3p derived from
cancer-associated fibroblasts (CAFs) inhibits F-box/WD
repeat-containing protein 7 and modulator of apoptosis 1 by
activating the Wnt/β-catenin pathway, and ultimately inhibits
mitochondria-related apoptosis (49). In PDAC, after the first-line
gemcitabine treatment, release of CAF exosomes can increase the
expression of epithelial chemotherapy resistance factor Snail and
promote tumor resistance and proliferation (50). Physical factors will also affect
the exosomes secreted by tumor cells. In a previous study, the
phosphorylation of anaplastic lymphoma kinase (ALK) in exosomes
released by the non-small lung cancer H3122 cell line after
irradiation was enhanced, and H3122 cells treated with these
exosomes showed significant resistance to ALK-specific inhibitors
such as crizotinib, ceritinib and TAE684 (51).
Finally, EV-VEGF90K derived from BCCs can interact
with heat shock protein (Hsp)90, blocking the antivascular effect
of bevacizumab, which can be eliminated by Hsp inhibitors (52).
Exosomes act as a communication tool in the TME and
promote tumor growth and invasion. Firstly, the exosomes produced
by stromal cells in tumors have an impact on tumor cells. For
example, Lazar et al (53)
found that, in the presence of adipocyte exosomes, fatty acid
oxidation is increased in melanoma cells, leading to increased
migration and invasion. Secondly, exosomes secreted by tumor cells
can also affect stromal cells to promote tumor cell metastasis and
invasion. α-smooth muscle actin is a common biomarker of CAFs.
Highly metastatic (HCC) cell-derived exosomal miR-1247-3p directly
targets β-1, 4-galactosyltransferase III, resulting in activation
of the β1-integrin-nuclear factor-κB signaling pathway in
fibroblasts. Activated CAFs further promote cancer progression by
secreting pro-inflammatory cytokines, including interleukin (IL)-6
and IL-8 (54). Additionally,
exosomal miR-105 derived from BCCs repress the expression of
MAX-interacting protein 1 (an antagonist of MYC transcriptional
activity) in CAFs, thereby inducing its MYC activation-related gene
expression and promoting tumor cell growth (55). Notably, McAtee et al
(56) found that treating
prostate stromal cells with tumor exosomes robustly stimulated
their migration in a manner dependent on hyaluronidases 1 (Hyal1)
catalytic activity, which explains why high levels of Hyal1 promote
prostate cancer progression. In a study of the interaction between
cancer cells and fibroblasts, Becker et al (57) found that tumor-derived EVs can
cause differentiation of fibroblasts into myofibroblasts, which
release matrix metalloproteinase (MMP) and remodel the ECM.
Exosomes secreted by tumor cells can change the
characteristics of vascular endothelial cells and promote
angiogenesis. For example, in the case of tumor cell hypoxia,
MSC-derived EVs regulate the formation of blood vessels through
various miRNAs, such as miR-126, miR-214 and miR-296 (58). miR-23a from nasopharyngeal
carcinoma is rich in exosomes, which can promote the growth,
migration and microtubule formation of human umbilical vein
endothelial cells (59). In
breast cancer, stromal fibroblasts can acquire the hallmarks of
CAFs as a result of the loss of p85α expression. Exosomes derived
from p85α-deficient fibroblasts can promote cancer progression via
EMT induced by the canonical Wnt pathway (60).
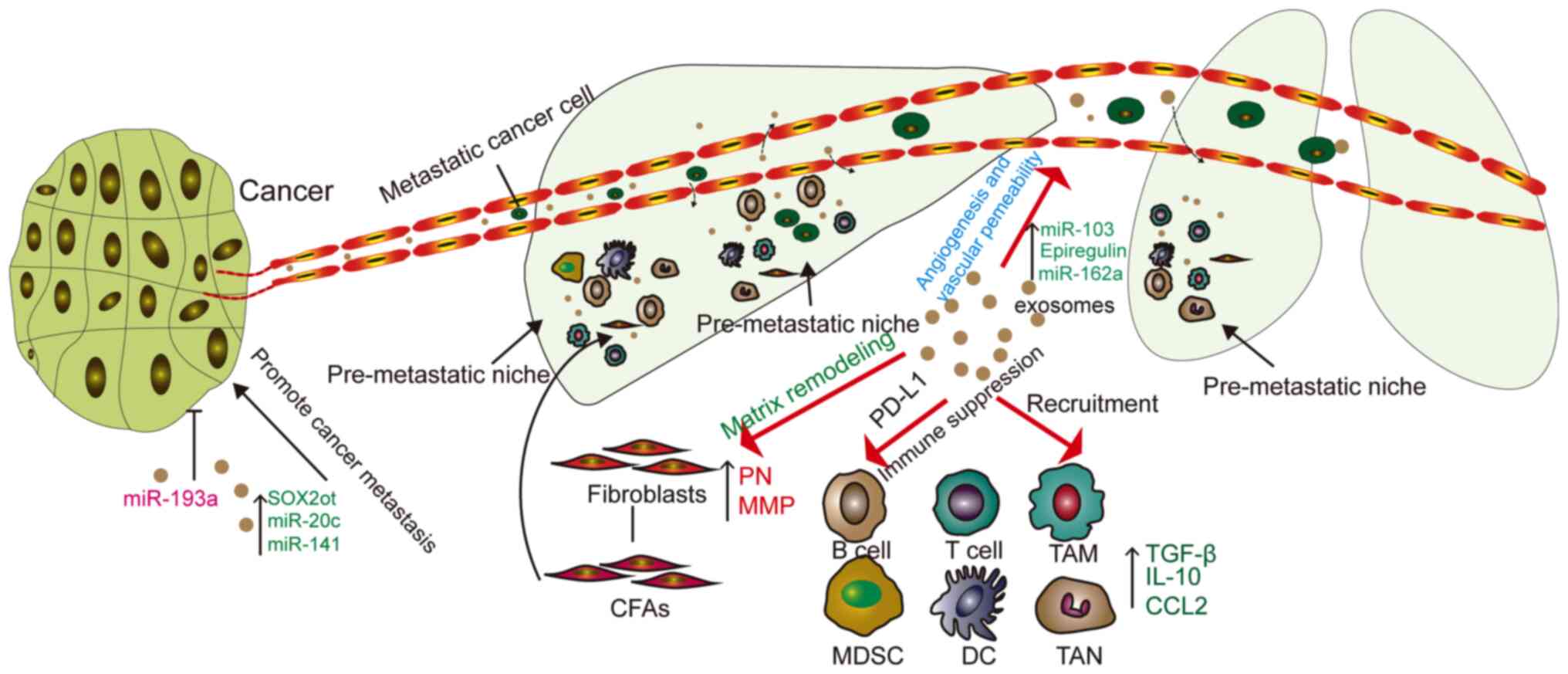
The cancer metastasis process involves several
steps; it starts with the local infiltration of cancer cells, after
which they enter the circulation through the lymphatic system or
blood vessels. From the circulation, cancer cells need to enter and
exit from the remote organs. Exosomes are involved in all steps of
this process (Fig. 2).
Exosomes can increase the EMT effect of tumor cells.
For example, in a previous study, the levels of miR-200c and
miR-141 in the plasma of patients with metastatic breast cancer
were significantly higher than those in the healthy group.
Additionally, exosomal miR-200c and miR-141 derived from metastatic
breast cancer also transferred to other breast cancer cells to
promote their metastasis and EMT (61).
Lung cancer cells produce more exosomes under
hypoxic conditions, and miR-23a is significantly upregulated in the
exosomes from these cells. This leads to hypoxia-inducible
factor-1α (HIF-1α) accumulation in endothelial cells, which
promotes the formation and permeability of blood vessels, as well
as tumor migration (62).
Moreover, hypoxia also increases gastric cancer (GC) exosome
release and miR-301α-3p expression in a HIF-1α-dependent manner,
which promotes GC progression and metastasis (63). The expression level of miR-103 of
exosomes in highly invasive HCC cells is significantly higher
compared with that in non-metastatic HCC cells. Endothelial cells
treated with highly invasive HCC cell-derived exosomes also show
greater permeability (64).
Unexpectedly, doxorubicin induced bone marrow stem cells to secrete
exosomes containing IL-13R and miR-126a in breast cancer-bearing
mice, which further induced IL13+ T-helper (Th)2 cells
to promote tumor angiogenesis (65).
Exosomes secreted by tumors can promote the
formation of pre-metastatic niches (PMNs). EVs secreted by tumor
cells form metastatic niches at remote sites of metastasis
(57). The matrix environment of
the niche prior to transfer is mainly composed of fibroblasts,
endothelial cells and ECM. Fibroblasts not only induce inflammation
and growth factors, but also express fibronectin and MMP (66). In a mouse model, CAF-derived EVs
induced PMN formation in mouse lungs, increasing salivary adenoid
cystic carcinoma lung metastasis (67). Exosomes in gastric cancer cells
can be delivered to the liver and integrate on the plasma membrane
of liver stromal cells. Exosomal epidermal growth factor receptor
can inhibit the expression of miR-26a/b and activates liver cell
growth factors, providing a supportive environment for gastric
cancer cells (68). The study by
Liu et al (69)
demonstrated that lung epithelial cells are critical for initiating
neutrophil recruitment and lung metastatic niche formation by
sensing tumor exosomal RNAs via Toll-like receptor 3 (TLR3),
providing suitable conditions for tumor metastasis.
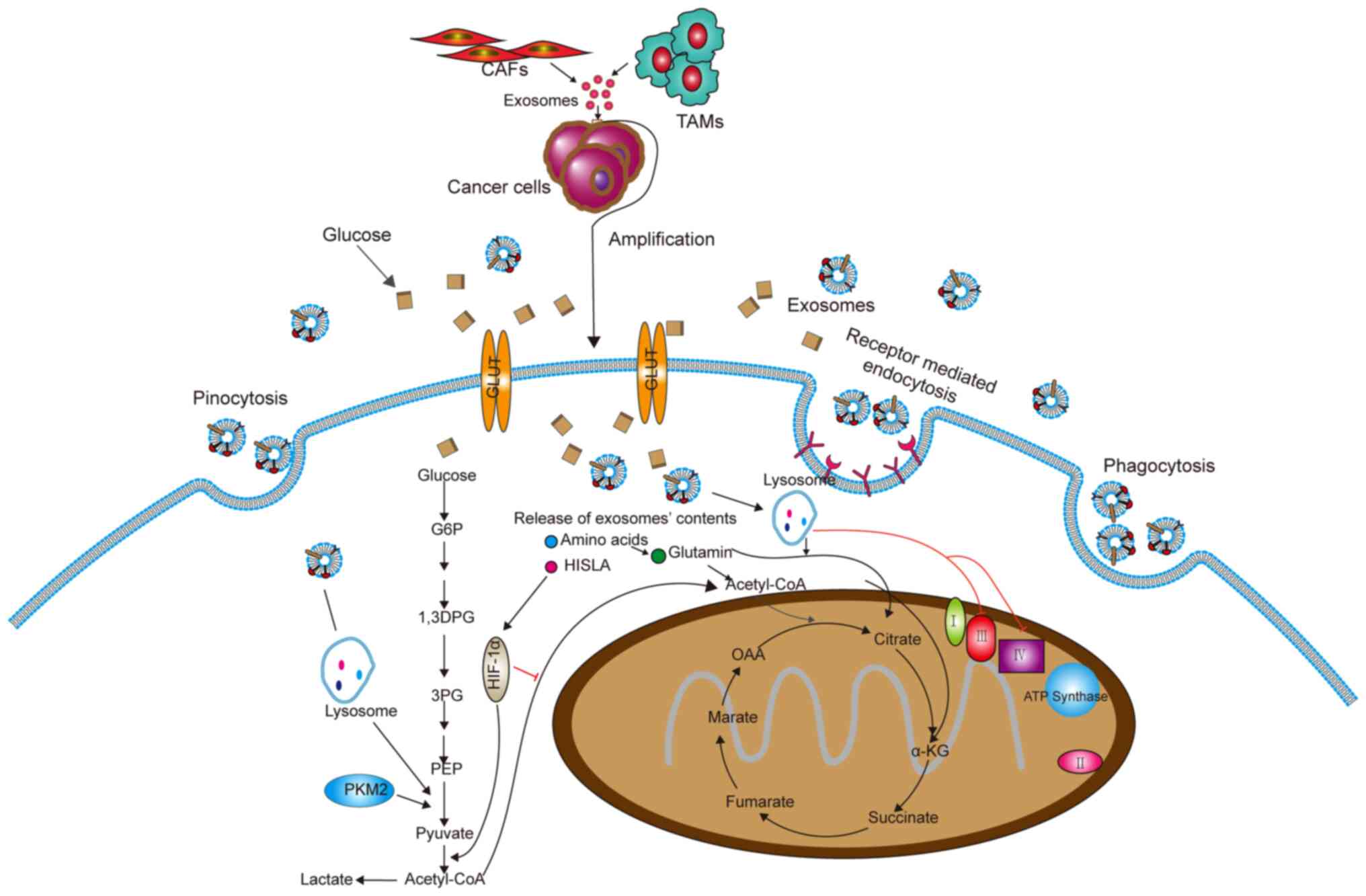
Metabolic reprogramming is an important
characteristic of cancer cells, and this is usually caused by the
TME (70). Cellular events
related to the metabolic pathway include the Warburg effect,
changes in Krebs cycle metabolites and the rate of oxidative
phosphorylation, which may provide energy and structural
requirements for the development and invasiveness of cancer cells
(71) (Fig. 3). Researchers have found that
exosomes secreted by CAFs in tumors can rebuild tumor metabolism.
Specifically, CAF-derived exosomes could inhibit mitochondrial
oxidative phosphorylation, thereby increasing glycolysis and
glutamine-dependent reductive carboxylation (72). In breast cancer, the exosomes
secreted by CAFs containing lncRNA small nucleolar RNA host gene 3
can sponge miR-330-5p, and miR-330-5p can target pyruvate kinase
M1/M2 (PKM1/2). Therefore, exosomes from CAFs can positively
regulate PKM expression, inhibit mitochondrial oxidative
phosphorylation, increase glycolytic carboxylation and enhance
breast tumor cell proliferation (73).
In addition, exosomes from tumor-associated
macrophages (TAMs) contain HIF-1α-stabilizing lncRNA (HISLA). HISLA
blocks the interaction of prolyl-4-hydroxylase domain-containing
proteins 2 and HIF-1α to inhibit the hydroxylation and degradation
of HIF-1α, thereby promoting cancer cell oxyglycolysis (74). These findings provide a new idea
for targeting exosomes to treat tumors.
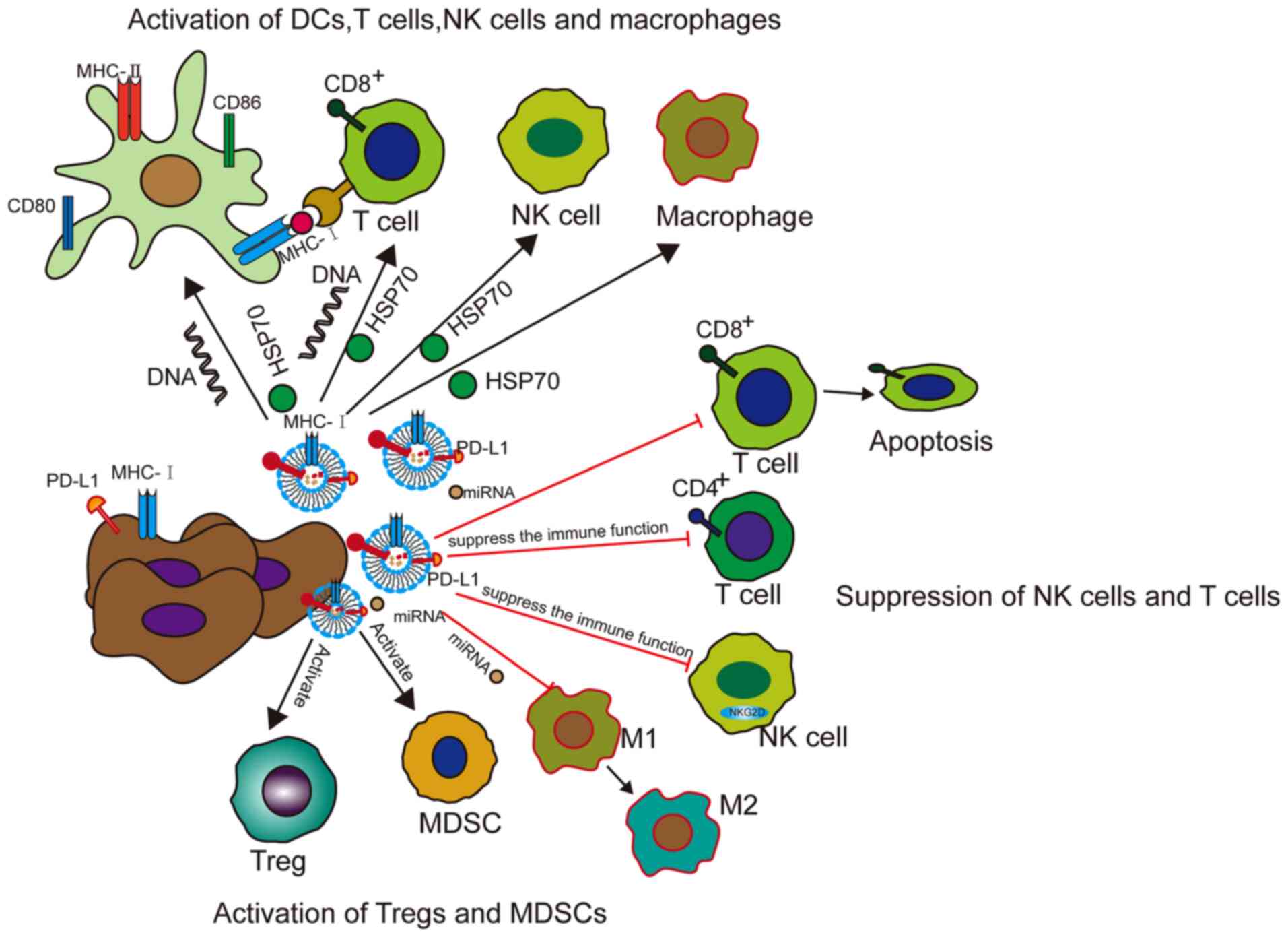
Exosomes not only affect the growth, invasion and
metastasis of tumor cells, but can also modulate the immune system
of the body. Moreover, exosomes can promote anti-tumor immunity
and, in a number of ways, inhibit the immune system within the TME
(75) (Fig. 4).
Tumor-derived exosomes or EVs can activate immune
responses. According to various reports, exosomes from tumor cells
present neoantigens and/or major histocompatibility complex (MHC)
peptide complexes, trigger and activate T cells through direct
presentation and cross presentation through dendritic cells (DCs),
or directly activate natural killer (NK) cells or macrophages
(76-78). For example, HSP70-80 and MHC-I
molecules of exosomes derived from tumor antigens and tumors can
interact with DCs to induce effective CD8+-dependent
anti- tumor effects on mouse tumor T cells (79). In addition, studies have reported
that tumor antigen proteins can be encapsulated by vesicles
secreted by DCs, drained to the lymph nodes and then transferred
between subpopulations of DCs. Synapses are formed between
interacting DCs. Vesicle metastasis occurs in the case of exosomes.
Studies have shown that DCs containing vesicles can activate T
cells, whereas DCs lacking vesicles cannot. These results help
predict the immune response and provide new methods for
immunotherapy (80-82).
Treating BCCs with topotecan, an antitumor drug that
causes DNA double-strand breaks, can induce the release of
exosomes. Consequently, exosomal DNA derived from cancer cells
treated with topotecan can trigger DC activation and subsequent
CD8+ T-cell activation through the cyclic
GMP-AMP/stimulator of interferon genes signaling pathway (83).
The ability of exosomes to express antigen and MHC
complexes, as well as induce helper T-cell immune responses,
increases the possibility that exosomes can be used as anti-cancer
vaccines. It is worth noting that HSP is an effective Th1 adjuvant.
Heat stress can induce the expression of HSPs and MHC-1 in tumor
cells, which can result in an increase in the immunogenicity of
these cells. Exosomes prepared with heat-stressed carcinoembryonic
antigen (CEA)-positive tumor cells can significantly induce
cytotoxic T lymphocyte (CTL) responses, indicating that exosomes
derived from heat-stressed tumor cells can be used as an effective
vaccine for cancer immunotherapy (84).
Exosomes not only play a role in antitumor immunity,
but they also inhibit the immune function in tumors. Unexpectedly,
exosomes participate in all known mechanisms by which cancer can
evade the immune system (85).
Firstly, exosomes can modulate regulatory T cells
(Tregs). For example, exosomes in the plasma of patients with head
and neck cancer (HNC) carry immunosuppressive molecules and
interfere with the functions of immune cells via downregulation of
NK cell group 2D expression in NK cells. Exosomes in the plasma of
patients with active disease significantly induce the apoptosis of
CD8+ T cells, suppress CD4+ T-cell
proliferation and upregulate Treg suppressor functions to promote
tumor growth (86). In a study on
liver cancer cells, the content of 14-3-3ζ in exosomes secreted by
these cells increased significantly. This content increased after
being taken up by tumor-infiltrating T lymphocytes (TILs). The TILs
then showed significant immune activation (CD69+),
proliferation (Ki-67+) and decreased antitumor activity
(87). Moreover, EVs can recruit
and activate Tregs and myeloid-derived suppressor cells (MDSCs),
and can inhibit CD8+ T-cell-mediated apoptosis of tumor
cells (56). Wen et al
(88) probed exosomes secreted by
mouse metastatic BCCs and found that they largely accumulated in
the mouse lungs, and suppressed the immune cell functions,
inhibited T-cell proliferation and decreased NK cell toxicity.
Overall, exosomes have both immune activation and
immune suppression functions in cancer. The effects of activating
immunity mainly depend on the antigen presentation of exosomes,
while the immunosuppressive effects of exosomes mainly depend on
the ligands, proteins and miRNAs they carry, which inhibit the
activity of cytotoxic T cells or increase immunosuppressive cells.
Understanding the underlying mechanisms of these two functions can
help lead to the development of exosomes as a novel method to treat
cancer.
Early diagnosis of tumors has always been a key
factor in tumor treatment. The clinical treatment of benign tumors
generally has a good prognosis. As exosomes exist in various body
fluids of the human body, including the saliva, urine and blood,
they can possibly be used as biomarkers for disease diagnosis and
prognosis (98) (Table II).
Increased levels of circulating EVs are found in
some common liver diseases such as hepatitis and liver cancer
(99). In lung cancer,
tumor-derived exosomal miRNAs include adenocarcinoma-specific
miR-181-5p, miR-30a-3p, miR-30e-3p and miR-361-5p, as well as
squamous cell carcinoma-specific miR-10b-5p, miR-15b-5p and
miR-320b. These miRNAs can be used as early diagnosis markers of
lung cancer (100). The
expression level of lncRNA-urothelial cancer-associated 1
(lncRNA-UCA1) in the exosomes from hypoxic bladder cancer cells was
significantly increased, and the expression level of lncRNA-UCA1 in
the serum exosomes of patients with bladder cancer was also
significantly increased. This lncRNA can therefore be used as a
diagnostic biomarker for bladder cancer (101). Prostate cancer is a common
cancer in men. The gold standard for diagnosis is a biopsy, which
can easily cause infection and bleeding. Evaluating the recurrence
by assessing the prostate-specific antigen (PSA) content in the
blood is insufficient. However, examination of prostate cancer
using exosomes in urine is particularly simple and fast. Moreover,
the exosomes secreted by the tumor increase based on the acidic
environment of this cancer, so that patients can express CD81 and
PSA nanovesicles at high levels (102). In one study, after the removal
of the prostate tumor, the glucuronic acid, D-ribose-5-phosphate
and isobutyryl-L-carnitine contents in urine EVs from patients were
all 2-26 times lower compared with those prior to surgery (103). Although the treatment success
rate has reached 80-90% in acute lymphoblastic leukemia (ALL), BM
biopsy still causes great pain to patients. Johnson et al
(104) found that exosomes
secreted by ALL cells contain high levels of CD19 compared with
normal cells, which can be used to diagnose ALL. Moreover, cancer
risk factors can influence the synthesis and release of exosomes.
Radiation can induce the release of exosome by activating p53
transcription, which can stimulate the expression of tumor
suppressor activated pathway-6 (105). In a study conducted by Wu et
al (106), it was found that
smoking can induce the release of EVs, >90% of which are
exosomes. When neurons are infected with Zika virus, expression of
neutral sphingomy-elinase-2, an important molecule that regulates
the production and release of exosomes, is activated (107). Numerous cancer types usually
show defects in the structure and number of centrosomes. Research
reports have shown that the presence of excessive centrosomes can
increase the secretion of exosomes, and that lysosomal dysfunction
causes cells with extra centrosomes to secrete more exosomes in
PDAC (108,109).
Exosomes can also be used as a prognostic indicator
of treatment. In a prospective study of patients with esophageal
squamous cell carcinoma, the level of GOLM1-NAA35 chimeric RNA in
the saliva of patients was used as a biomarker to evaluate
treatment response, recurrence and early detection (110). Exosomes from the malignant
ascites of patients with GC can promote the invasion of human
gastric adenocarcinoma (AGS) cells. In mouse abdominal xenograft
models using AGS cells, the addition of exosomes separated from
malignant ascites decreased the median survival time of the mice
significantly. Additionally, miR-196, miR-92 and miR-1307 are
highly expressed in malignant ascites exosomes (111). Manier et al (112) showed that a cohort of 156
patients with newly diagnosed and uniformly treated multiple
myeloma had exosomes containing miR-18a and miR-let-7b in the
serum. Levels of these miRNAs were significantly associated with
patient progression-free survival (PFS) and overall survival times.
Exosomal miR-24-3p secreted by nasopharyngeal carcinoma cells can
target fibroblast growth factor 11 to inhibit T-cell function.
Studies have shown that patients with lower serum level of exosomal
miR-24-3p have a longer PFS time (113-115). In osteosarcoma, Xu et al
(116) found that in patients
with OS and a poor chemotherapeutic response compared with in those
with a good chemotherapeutic response, miR-133a, miR-124,
miR-199-3p and miR-385 levels were significantly lower, while
miR-135b, miR-148a, miR-27a and miR-9 were greatly overexpressed in
serum exosomes. These data indicate that various exosomal RNA
molecules may serve as reliable biomarkers for osteosarcoma tumors
with different chemotherapeutic sensitivities.
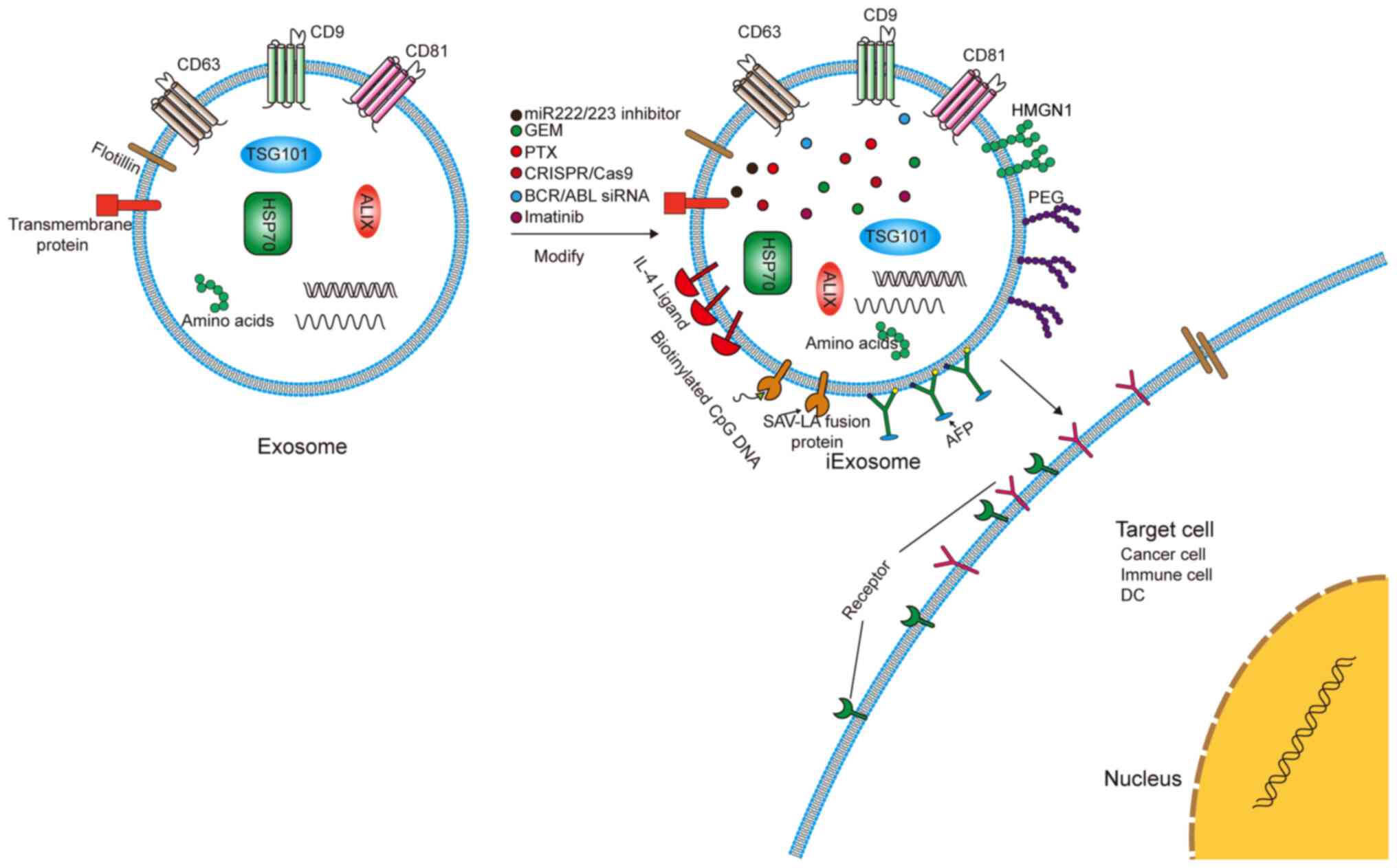
Exosomes, as bioactive nanovesicle substances
secreted by cells, can play an important role in targeted therapy
(Fig. 5). The current clinical
trials of exosomes in patients with cancer are summarized in
Table III. Exosomes are stable,
membrane-permeable, and can even pass through the blood-brain
barrier. Additionally, exosomes can be combined with a number of
physical materials and can therefore be used as natural
nanoparticles for drug delivery and the delivery of miRNAs/small
interfering RNAs (siRNAs) to treat diseases and tumors (117).
Exosomes have a double-layer membrane structure and
nanometer size, which can protect them from clearance by the cell.
This prolongs their circulating half-life and improves their
biological activity (118).
However, the half-life of an exosome in plasma is only 2-4 min
(119). A variety of strategies
have been proposed to improve the tumor cell targeting specificity
and tumor uptake efficiency of exosomes. For example, the
engineered exosomes, iExosomes, can target the Kirsten rat sarcoma
viral oncogene protein for treating pancreatic cancer (120). In lung cancer, exosomes modified
with polyethylene glycol can improve the circulation time of
paclitaxel-containing exosomes in the blood. Researchers have
inserted RNA nanoparticles into the membrane of exosomes with tails
that carry specific ligands. This enables the exosomes to target
specific tumor cells and deliver the loaded siRNA into the cells
(121). Notably, exosomes can
also treat drug-resistant tumor cells. Imatinib can improve the
prognosis of patients with CML; however, acquired resistance is
still being encountered. Bellavia et al (122) attempted to transfer imatinib or
breakpoint cluster region-c-abl oncogene 1 siRNA into exosomes
fused with IL-3, which targeted CML cells and inhibited tumor
growth in vitro and in vivo. Bliss et al
(123) reported that BCCs can
prime MSCs to release exosomes containing distinct miRNA contents,
such as miR-222/223, which in turn promote quiescence and confer
drug resistance in a subset of cancer cells. Building on these
results, a novel, nontoxic therapeutic strategy was developed to
target dormant BCCs based on systemic administration of MSCs loaded
with antagomiR-222/223. In an immunodeficient mouse model of
dormant breast cancer, this therapy sensitized BCCs to
carboplatin-based therapy and increased host survival times.
Cancer-derived exosomes function as natural carriers that can
efficiently deliver clustered regularly interspaced short
palindromic repeats (CRISPR)/CRISPR associated protein 9 plasmids
to cancer cells, inhibit the expression of poly(ADP-ribose)
polymerase family member 1 and induce the death of ovarian cancer
cells (124). Milk-derived
exosomes have been investigated for oral delivery of the
chemotherapeutic drug paclitaxel as an alternative to conventional
intravenous therapy for improved efficacy and reduced toxicity
(125).
Another valuable application of exosomes is their
use as anticancer vaccines. Exosomes secreted by M1 macrophages can
promote an inflammatory response. Encapsulating the
tyrosinase-related protein 2 vaccine with lipid calcium phosphate
nanoparticle enhances its activity and induces a stronger
antigen-specific cytotoxic T-cell response (126). In HCC, tumor-derived exosomes
(TDEs) can elicit a stronger immune response than cell lysates
in vitro and in vivo (127). Exosomes derived from
α-fetoprotein (AFP)-expressing DCs (DEXAFP) and TDEs painted with
the functional domain of high mobility group nucleosome-binding
protein 1 elicited strong antigen-specific immune responses.
Researchers found that HCC mice treated with DEXAFP had more
γ-interferon (IFN-γ)-expressing CD8+ T lymphocytes, high
levels of IFN-γ and IL-2, fewer CD25+Foxp3+
Tregs, and lower levels of IL-10 and TGFβ in the tumors (128,129). Japanese researchers transfected
B16BL6 cells with plasmids containing streptavidin (SAV), a
high-affinity knot and biotin protein, and lactobacillus adhesion
(LA), an exosomal promoting protein, to produce exosomes with
expression of SAV-LA. This effectively enhanced the antigen
presentation capabilities of the exosomes (130).
Exosomes can contain biologically active substances
such as proteins, nucleic acids and lipids. Exosomes can promote
cancer cell drug resistance, invasion and metastasis. Therefore,
the bioactive molecules contained within the exosomes can be used
as a targeted therapy for treating tumors. In the clinic, the use
of exosomes in body fluids to diagnose diseases and assess patient
prognosis has also made significant progress. However, despite
numerous studies and literature reports on exosomes, it remains a
great challenge to translate these basic science studies to
clinical applications. Most of the effects of exosomes on tumor
progression or immunity have mainly been determined by
'gain-of-function' experiments, which may not fully represent the
mechanism of action of exosomes in tumors. Secondly, as a biomarker
for tumor diagnosis and prognosis, the extraction of exosomes is
still a cumbersome task. How to extract them efficiently, rapidly
and in large quantities remains a problem that needs to be
addressed. Moreover, as a treatment method, the biological safety
of exosomes still requires careful investigation. The next step is
to decrease the cost of the extraction of exosomes, improve the
safety of exosomes in clinical applications and develop a method to
produce exosome-mimetic vesicles.
Although there are a number of challenges in their
clinical application, exosomes are considered valid diagnostic
biomarkers and potential therapeutic tools in cancer. Moreover,
along with chemical, cellular and genetic engineering techniques,
exosomal modification strategies may be promising for the
development of clinical therapies.
Not applicable.
XW and YZ contributed equally to this study. KD is
the correspondence author. XW was the main contributor in writing
the manuscript. YZ was the main contributor in making tables and
figures. KD was the main contributor in revising the manuscript
critically for important content. All authors have read and
approved the manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
No funding was received.
|
1
|
Wolf P: The nature and significance of
platelet products in human plasma. Br J Haematol. 13:269–288. 1967.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pan BT and Johnstone RM: Fate of the
transferrin receptor during maturation of sheep reticulocytes in
vitro: Selective externalization of the receptor. Cell. 33:967–978.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnstone RM, Adam M, Hammond JR, Orr L
and Turbide C: Vesicle formation during reticulocyte maturation.
Association of plasma membrane activities with released vesicles
(exosomes). J Biol Chem. 262:9412–9420. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bonifacino JS: Vesicular transport earns a
nobel. Trends Cell Biol. 24:3–5. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jadli AS, Ballasy N, Edalat P and Patel
VB: Inside(sight) of tiny communicator: Exosome biogenesis,
secretion, and uptake. Mol Cell Biochem. 467:77–94. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ruivo CF, Adem B, Silva M and Melo SA: The
biology of cancer exosomes: Insights and new perspectives. Cancer
Res. 77:6480–6488. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kok VC and Yu CC: Cancer-derived exosomes:
Their role in cancer biology and biomarker development. Int J
Nanomedicine. 15:8019–8036. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Plebanek MP, Angeloni NL, Vinokour E, Li
J, Henkin A, Martinez-Marin D, Filleur S, Bhowmick R, Henkin J,
Miller SD, et al: Pre-metastatic cancer exosomes induce immune
surveillance by patrolling monocytes at the metastatic niche. Nat
Commun. 8:13192017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim MS, Haney MJ, Zhao Y, Mahajan V,
Deygen I, Klyachko NL, Inskoe E, Piroyan A, Sokolsky M, Okolie O,
et al: Development of exosome-encapsulated paclitaxel to overcome
MDR in cancer cells. Nanomedicine. 12:655–664. 2016. View Article : Google Scholar :
|
|
10
|
Möller A and Lobb RJ: The evolving
translational potential of small extracellular vesicles in cancer.
Nat Rev Cancer. 20:697–709. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kalluri R and LeBleu VS: The biology,
function, and biomedical applications of exosomes. Science.
367:eaau69772020. View Article : Google Scholar :
|
|
12
|
Simons M and Raposo G: Exosomes-vesicular
carriers for intercellular communication. Curr Opin Cell Biol.
21:575–581. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fujita Y, Kosaka N, Araya J, Kuwano K and
Ochiya T: Extracellular vesicles in lung microenvironment and
pathogenesis. Trends Mol Med. 21:533–542. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hoshino D, Kirkbride KC, Costello K, Clark
ES, Sinha S, Grega-Larson N, Tyska MJ and Weaver AM: Exosome
secretion is enhanced by invadopodia and drives invasive behavior.
Cell Rep. 5:1159–1168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Colombo M, Raposo G and Thery C:
Biogenesis, secretion, and intercellular interactions of exosomes
and other extracellular vesicles. Annu Rev Cell Dev Biol.
30:255–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Frydrychowicz M, Koleckabednarczyk A,
Madejczyk M, Yasar S and Dworacki G: Exosomes-structure, biogenesis
and biological role in non-small-cell lung cancer. Scand J Immunol.
81:2–10. 2015. View Article : Google Scholar
|
|
17
|
Frühbeis C, Fröhlich D, Kuo WP, Amphornrat
J, Thilemann S, Saab AS, Kirchhoff F, Möbius W, Goebbels S, Nave
KA, et al: Neurotransmitter-triggered transfer of exosomes mediates
oligodendrocyte-neuron communication. PLoS Biol. 11:e10016042013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zylbersztejn K and Galli T: Vesicular
traffic in cell navigation. FEBS J. 278:4497–4505. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tran N: Cancer exosomes as miRNA
factories. Trends Cancer. 2:329–331. 2016. View Article : Google Scholar
|
|
20
|
Melo SA, Sugimoto H, O'Connell JT, Kato N,
Villanueva A, Vidal A, Qiu L, Vitkin E, Perelman LT, Melo CA, et
al: Cancer exosomes perform cell-independent microRNA biogenesis
and promote tumorigenesis. Cancer Cell. 26:707–721. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Yeung BZ, Cui M, Peer CJ, Lu Z,
Figg WD, Guillaume Wientjes M, Woo S and Au JL: Exosome is a
mechanism of intercellular drug transfer: Application of
quantitative pharmacology. J Control Release. 268:147–158. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei Y, Wang D, Jin F, Bian Z, Li L, Liang
H, Li M, Shi L, Pan C, Zhu D, et al: Pyruvate kinase type M2
promotes tumour cell exosome release via phosphorylating
synaptosome-associated protein 23. Nat Commun. 8:140412017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gu H, Chen C, Hao X, Wang C, Zhang X, Li
Z, Shao H, Zeng H, Yu Z, Xie L, et al: Sorting protein VPS33B
regulates exosomal autocrine signaling to mediate hematopoiesis and
leukemogenesis. J Clin Invest. 126:4537–4553. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan Q, Yang L, Zhang X, Peng X, Wei S, Su
D, Zhai Z, Hua X and Li H: The emerging role of exosome-derived
non-coding RNAs in cancer biology. Cancer Lett. 414:107–115. 2018.
View Article : Google Scholar
|
|
25
|
Bobrie A, Colombo M, Raposo G and Théry C:
Exosome secretion: Molecular mechanisms and roles in immune
responses. Traffic. 12:1659–1668. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li I and Nabet BY: Exosomes in the tumor
microenvironment as mediators of cancer therapy resistance. Mol
Cancer. 18:322019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wortzel I, Dror S, Kenific CM and Lyden D:
Exosome-mediated metastasis: Communication from a distance. Dev
Cell. 49:347–360. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sung BH, Ketova T, Hoshino D, Zijlstra A
and Weaver AM: Directional cell movement through tissues is
controlled by exosome secretion. Nat Commun. 6:71642015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Tesic Mark M, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumour exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Milane L, Singh A, Mattheolabakis G,
Suresh M and Amiji MM: Exosome mediated communication within the
tumor microenvironment. J Control Release. 219:278–294. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Junttila MR and de Sauvage FJ: Influence
of tumour micro-environment heterogeneity on therapeutic response.
Nature. 501:346–354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tsukamoto H, Fujieda K, Senju S, Ikeda T,
Oshiumi H and Nishimura Y: Immune-suppressive effects of
interleukin-6 on T-cell-mediated anti-tumor immunity. Cancer Sci.
109:523–530. 2018. View Article : Google Scholar
|
|
33
|
Nishikawa H and Sakaguchi S: Regulatory T
cells in cancer immunotherapy. Curr Opin Immunol. 27:1–7. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Warburg O, Wind F and Negelein E: The
metabolism of tumors in the body. J Gen Physiol. 8:519–530. 1927.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gatenby RA, Gawlinski ET, Gmitro AF,
Kaylor BM and Gillies RJ: Acid-mediated tumor invasion: A
multidisciplinary study. Cancer Res. 66:5216–5223. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pace KR, Dutt R and Galileo DS: Exosomal
L1CAM stimulates glioblastoma cell motility, proliferation, and
invasiveness. Int J Mol Sci. 20:39822019. View Article : Google Scholar :
|
|
37
|
Guo L, Zhu Y, Li L, Zhou S, Yin G, Yu G
and Cui H: Breast cancer cell-derived exosomal miR-20a-5p promotes
the proliferation and differentiation of osteoclasts by targeting
SRCIN1. Cancer Med. 8:5687–5701. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang S, Su X, Xu M, Xiao X, Li X, Li H,
Keating A and Zhao RC: Exosomes secreted by mesenchymal
stromal/stem cell-derived adipocytes promote breast cancer cell
growth via activation of Hippo signaling pathway. Stem Cell Res
Ther. 10:1172019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee FT, Mountain AJ, Kelly MP, Hall C,
Rigopoulos A, Johns TG, Smyth FE, Brechbiel MW, Nice EC, Burgess AW
and Scott AM: Enhanced efficacy of radioimmunotherapy with
90Y-CHX-A′-DTPA-hu3S193 by inhibition of epidermal growth factor
receptor (EGFR) signaling with EGFR tyrosine kinase inhibitor
AG1478. Clin Cancer Res. 11:7080s–7086s. 2005. View Article : Google Scholar
|
|
40
|
Luo F, Sun Z, Han Q, Xue C and Bai C:
Effect of human hepatocellular carcinoma HepG2 Cell-derived exosome
on the differentiation of mesenchymal stem cells and their
interaction. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 39:312–317.
2017.PubMed/NCBI
|
|
41
|
Amit M, Takahashi H, Dragomir MP,
Lindemann A, Gleber-Netto FO, Pickering CR, Anfossi S, Osman AA,
Cai Y, Wang R, et al: Loss of p53 drives neuron reprogramming in
head and neck cancer. Nature. 578:449–454. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Godlewski J, Ferrerluna R, Rooj AK, Mineo
M, Ricklefs F, Takeda YS, Nowicki MO, Salińska E, Nakano I, Lee H,
et al: MicroRNA signatures and molecular subtypes of glioblastoma:
The role of extracellular transfer. Stem Cell Reports. 8:1497–1505.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Qu L, Ding J, Chen C, Wu ZJ, Liu B, Gao Y,
Chen W, Liu F, Sun W, Li XF, et al: Exosome-transmitted lncARSR
promotes sunitinib resistance in renal cancer by acting as a
competing endogenous RNA. Cancer Cell. 29:653–668. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lunavat TR, Cheng L, Einarsdottir BO,
Olofsson Bagge R, Veppil Muralidharan S, Sharples RA, Lässer C, Gho
YS, Hill AF, Nilsson JA and Lötvall J: BRAFV600
inhibition alters the microRNA cargo in the vesicular secretome of
malignant melanoma cells. Proc Natl Acad Sci SA. 114:E5930–E5939.
2017. View Article : Google Scholar
|
|
45
|
Mikamori M, Yamada D, Eguchi H, Hasegawa
S, Kishimoto T, Tomimaru Y, Asaoka T, Noda T, Wada H, Kawamoto K,
et al: MicroRNA-155 controls exosome synthesis and promotes
gemcitabine resistance in pancreatic ductal adenocarcinoma. Sci
Rep. 7:423392017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lobb RJ, Van Amerongen R, Wiegmans AP, Ham
S, Larsen JE and Möller A: Exosomes derived from mesenchymal
non-small cell lung cancer cells promote chemoresistance. Int J
Cancer. 141:614–620. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bach D, Hong J, Park HJ and Lee SK: The
role of exosomes and miRNAs in drug-resistance of cancer cells. Int
J Cancer. 141:220–230. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Crompot E, Van Damme M, Pieters K,
Vermeersch M, Perez-Morga D, Mineur P, Maerevoet M, Meuleman N,
Bron D, Lagneaux L and Stamatopoulos B: Extracellular vesicles of
bone marrow stromal cells rescue chronic lymphocytic leukemia B
cells from apoptosis, enhance their migration and induce gene
expression modifications. Haematologica. 102:1594–1604. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hu JL, Wang W, Lan X, Zeng ZC, Liang YS,
Yan YR, Song FY, Wang FF, Zhu XH, Liao WJ, et al: CAFs secreted
exosomes promote metastasis and chemotherapy resistance by
enhancing cell stemness and epithelial-mesenchymal transition in
colorectal cancer. Mol Cancer. 18:912019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Richards KE, Zeleniak AE, Fishel ML, Wu J,
Littlepage LE and Hill R: Cancer-associated fibroblast exosomes
regulate survival and proliferation of pancreatic cancer cells.
Oncogene. 36:1770–1778. 2017. View Article : Google Scholar :
|
|
51
|
Wu H, Zeng C, Ye Y, Liu J, Mu Z, Xie Y,
Chen B, Nong Q and Wu D: Exosomes from irradiated nonsmall cell
lung cancer cells reduced sensitivity of recipient cells to
anaplastic lymphoma kinase inhibitors. Mol Pharm. 15:1892–1900.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Feng Q, Zhang C, Lum D, Druso JE, Blank B,
Wilson KF, Welm A, Antonyak MA and Cerione RA: A class of
extracellular vesicles from breast cancer cells activates VEGF
receptors and tumour angiogenesis. Nat Commun. 8:144502017.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lazar I, Clement E, Dauvillier S, Milhas
D, Ducoux-Petit M, LeGonidec S, Moro C, Soldan V, Dalle S, Balor S,
et al: Adipocyte exosomes promote melanoma aggressiveness through
fatty acid oxidation: A novel mechanism linking obesity and cancer.
Cancer Res. 76:4051–4057. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Fang T, Lv H, Lv G, Li T, Wang C, Han Q,
Yu L, Su B, Guo L, Huang S, et al: Tumor-derived exosomal
miR-1247-3p induces cancer-associated fibroblast activation to
foster lung metastasis of liver cancer. Nat Commun. 9:1912018.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yan W, Wu X, Zhou W, Fong MY, Cao M, Liu
J, Liu X, Chen CH, Fadare O, Pizzo DP, et al: Cancer-cell-secreted
exosomal miR-105 promotes tumour growth through the MYC-dependent
metabolic reprogramming of stromal cells. Nat Cell Biol.
20:597–609. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
McAtee CO, Booth C, Elowsky C, Zhao L,
Payne J, Fangman T, Caplan S, Henry MD and Simpson MA: Prostate
tumor cell exosomes containing hyaluronidase Hyal1 stimulate
prostate stromal cell motility by engagement of FAK-mediated
integrin signaling. Matrix Biol. 78-79:165–179. 2019. View Article : Google Scholar
|
|
57
|
Becker A, Thakur BK, Weiss JM, Kim HS,
Peinado H and Lyden D: Extracellular vesicles in cancer:
Cell-to-cell mediators of metastasis. Cancer Cell. 30:836–848.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Todorova D, Simoncini S, Lacroix R,
Sabatier F and Dignat-George F: Extracellular vesicles in
angiogenesis. Circ Res. 120:1658–1673. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Bao L, You B, Shi S, Shan Y, Zhang Q, Yue
H, Zhang J, Zhang W, Shi Y, Liu Y, et al: Metastasis-associated
miR-23a from nasopharyngeal carcinoma-derived exosomes mediates
angiogenesis by repressing a novel target gene TSGA10. Oncogene.
37:2873–2889. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chen Y, Zeng C, Zhan Y, Wang H, Jiang X
and Li W: Aberrant low expression of p85α in stromal fibroblasts
promotes breast cancer cell metastasis through exosome-mediated
paracrine Wnt10b. Oncogene. 36:4692–4705. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang G, Zhang W, Li B, Stringer-Reasor E,
Chu C, Sun L, Bae S, Chen D, Wei S, Jiao K, et al: MicroRNA-200c
and microRNA-141 are regulated by a FOXP3-KAT2B axis and associated
with tumor metastasis in breast cancer. Breast Cancer Res.
19:732017. View Article : Google Scholar
|
|
62
|
Hsu YL, Hung J, Chang W, Lin YS, Pan YC,
Tsai PH, Wu CY and Kuo PL: Hypoxic lung cancer-secreted exosomal
miR-23a increased angiogenesis and vascular permeability by
targeting prolyl hydroxylase and tight junction protein ZO-1.
Oncogene. 36:4929–4942. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Xia X, Wang S, Ni B, Xing S, Cao H, Zhang
Z, Yu F, Zhao E and Zhao G: Hypoxic gastric cancer-derived exosomes
promote progression and metastasis via MiR-301a-3p/PHD3/HIF-1α
positive feedback loop. Oncogene. 39:6231–6244. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Fang JH, Zhang ZJ, Shang LR, Luo YW, Lin
YF, Yuan Y and Zhuang SM: Hepatoma cell-secreted exosomal
microRNA-103 increases vascular permeability and promotes
metastasis by targeting junction proteins. Hepatology.
68:1459–1475. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Matei I, Kim HS and Lyden D: Unshielding
exosomal RNA unleashes tumor growth and metastasis. Cell.
170:223–225. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Yamamura Y, Asai N, Enomoto A, Kato T, Mii
S, Kondo Y, Ushida K, Niimi K, Tsunoda N, Nagino M, et al:
Akt-Girdin signaling in cancer-associated fibroblasts contributes
to tumor progression. Cancer Res. 75:813–823. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kong J, Tian H, Zhang F, Zhang Z, Li J,
Liu X, Li X, Liu J, Li X, Jin D, et al: Extracellular vesicles of
carcinoma-associated fibroblasts creates a pre-metastatic niche in
the lung through activating fibroblasts. Mol Cancer. 18:1752019.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhang H, Deng T, Liu R, Bai M, Zhou L,
Wang X, Li S, Wang X, Yang H, Li J, et al: Exosome-delivered EGFR
regulates liver microenvironment to promote gastric cancer liver
metastasis. Nat Commun. 8:150162017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liu Y, Gu Y, Han Y, Zhang Q, Jiang Z,
Zhang X, Huang B, Xu X, Zheng J and Cao X: Tumor exosomal RNAs
promote lung pre-metastatic niche formation by activating alveolar
epithelial TLR3 to recruit neutrophils. Cancer Cell. 30:243–256.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Lane AN, Higashi RM and Fan TW: Metabolic
reprogramming in tumors: Contributions of the tumor
microenvironment. Genes Dis. 7:185–198. 2019. View Article : Google Scholar
|
|
71
|
Ji K, Mayernik L, Moin K and Sloane BF:
Acidosis and proteolysis in the tumor microenvironment. Cancer
Metastasis Rev. 38:103–112. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhao H, Yang L, Baddour J, Achreja A,
Bernard V, Moss T, Marini JC, Tudawe T, Seviour EG, San Lucas FA,
et al: Tumor microenvironment derived exosomes pleiotropically
modulate cancer cell metabolism. Elife. 5:e102502016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Li Y, Zhao Z, Liu W and Li X: SNHG3
functions as miRNA sponge to promote breast cancer cells growth
through the metabolic reprogramming. Appl Biochem Biotechnol.
191:1084–1099. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Tomasetti M, Lee W, Santarelli L and
Neuzil J: Exosome-derived microRNAs in cancer metabolism: Possible
implications in cancer diagnostics and therapy. Exp Mol Med.
49:e2852017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Greening DW, Gopal SK, Xu R, Simpson RJ
and Chen W: Exosomes and their roles in immune regulation and
cancer. Semin Cell Dev Biol. 40:72–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Filipazzi P, Bürdek M, Villa A, Rivoltini
L and Huber V: Recent advances on the role of tumor exosomes in
immunosuppression and disease progression. Semin Cancer Biol.
22:342–349. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wolfers J, Lozier A, Raposo G, Regnault A,
Théry C, Masurier C, Flament C, Pouzieux S, Faure F, Tursz T, et
al: Tumor-derived exosomes are a source of shared tumor rejection
antigens for CTL cross-priming. Nat Med. 7:297–303. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Dai S, Zhou X, Wang B, Wang Q, Fu Y, Chen
T, Wan T, Yu Y and Cao X: Enhanced induction of dendritic cell
maturation and HLA-A*0201-restricted CEA-specific CD8(+) CTL
response by exosomes derived from IL-18 gene-modified CEA-positive
tumor cells. J Mol Med (Berl). 84:1067–1076. 2006. View Article : Google Scholar
|
|
79
|
Altieri SL, Khan AN and Tomasi TB:
Exosomes from plasmacytoma cells as a tumor vaccine. J Immunother.
27:282–288. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Balan S and Bhardwaj N: Cross-presentation
of tumor antigens is ruled by synaptic transfer of vesicles among
dendritic cell subsets. Cancer Cell. 37:751–753. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zitvogel L, Regnault A, Lozier A, Wolfers
J, Flament C, Tenza D, Ricciardi-Castagnoli P, Raposo G and
Amigorena S: Eradication of established murine tumors using a novel
cell-free vaccine: Dendritic cell-derived exosomes. Nat Med.
4:594–600. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Pitt JM, André F, Amigorena S, Soria JC,
Eggermont A, Kroemer G and Zitvogel L: Dendritic cell-derived
exosomes for cancer therapy. J Clin Invest. 126:1224–1232. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Kitai Y, Kawasaki T, Sueyoshi T, Kobiyama
K, Ishii KJ, Zou J, Akira S, Matsuda T and Kawai T: DNA-containing
exosomes derived from cancer cells treated with topotecan activate
a STING-dependent pathway and reinforce antitumor immunity. J
Immunol. 198:1649–1659. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Dai S, Wan T, Wang B, Zhou X, Xiu F, Chen
T, Wu Y and Cao X: More efficient induction of
HLA-A*0201-restricted and carcinoembryonic antigen (CEA)-specific
CTL response by immunization with exosomes prepared from
heat-stressed CEA-positive tumor cells. Clin Cancer Res.
11:7554–7563. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Czernek L and Düchler M: Functions of
cancer-derived extracellular vesicles in immunosuppression. Arch
Immunol Ther Exp (Warsz). 65:311–323. 2017. View Article : Google Scholar
|
|
86
|
Ludwig S, Floros T, Theodoraki MN, Hong
CS, Jackson EK, Lang S and Whiteside TL: Suppression of lymphocyte
functions by plasma exosomes correlates with disease activity in
patients with head and neck cancer. Clin Cancer Res. 23:4843–4854.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wang X, Shen H, Zhangyuan G, Huang R,
Zhang W, He Q, Jin K, Zhuo H, Zhang Z, Wang J, et al: 14-3-3ζ
delivered by hepatocellular carcinoma-derived exosomes impaired
anti-tumor function of tumor-infiltrating T lymphocytes. Cell Death
Dis. 9:1592018. View Article : Google Scholar
|
|
88
|
Wen SW, Sceneay J, Lima LG, Wong CS,
Becker M, Krumeich S, Lobb RJ, Castillo V, Wong KN, Ellis S, et al:
The biodistribution and immune suppressive effects of breast
cancer-derived exosomes. Cancer Res. 76:6816–6827. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Poggio M, Hu T, Pai CC, Chu B, Belair CD,
Chang A, Montabana E, Lang UE, Fu Q, Fong L and Blelloch R:
Suppression of exosomal PD-L1 induces systemic anti-tumor immunity
and memory. Cell. 177:414–427.e13. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Gabrusiewicz K, Li X, Wei J, Hashimoto Y,
Marisetty AL, Ott M, Wang F, Hawke D, Yu J, Healy LM, et al:
Glioblastoma stem cell-derived exosomes induce M2 macrophages and
PD-L1 expression on human monocytes. Oncoimmunology.
7:e14129092018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Razzo BM, Ludwig N, Hong CS, Sharma P,
Fabian KP, Fecek RJ, Storkus WJ and Whiteside TL: Tumor-derived
exosomes promote carcinogenesis of murine oral squamous cell
carcinoma. Carcinogenesis. 41:625–633. 2020. View Article : Google Scholar
|
|
92
|
Haderk F, Schulz R, Iskar M, Cid LL, Worst
T, Willmund KV, Schulz A, Warnken U, Seiler J, Benner A, et al:
Tumor-derived exosomes modulate PD-L1 expression in monocytes. Sci
Immunol. 2:eaah55092017. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Chen X, Zhou J, Li X and Wang X, Lin Y and
Wang X: Exosomes derived from hypoxic epithelial ovarian cancer
cells deliver microRNAs to macrophages and elicit a tumor-promoted
phenotype. Cancer Lett. 435:80–91. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Wang X, Luo G, Zhang K, Cao J, Huang C,
Jiang T, Liu B, Su L and Qiu Z: Hypoxic tumor-derived exosomal
miR-301a mediates M2 macrophage polarization via PTEN/PI3Kγ to
promote pancreatic cancer metastasis. Cancer Res. 78:4586–4598.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Hsieh CH, Tai SK and Yang MH:
Snail-overexpressing cancer cells promote M2-like polarization of
tumor-associated macrophages by delivering MiR-21-abundant
exosomes. Neoplasia. 20:775–788. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Cooks T, Pateras IS, Jenkins LM, Patel KM,
Robles AI, Morris J, Forshew T, Appella E, Gorgoulis VG and Harris
CC: Mutant p53 cancers reprogram macrophages to tumor supporting
macrophages via exosomal miR-1246. Nat Commun. 9:7712018.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Casadei L, Calore F, Creighton CJ,
Guescini M, Batte K, Iwenofu OH, Zewdu A, Braggio DA, Bill KL,
Fadda P, et al: Exosome-derived miR-25-3p and miR-92a-3p stimulate
liposarcoma progression. Cancer Res. 77:3846–3856. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Whiteside TL: Tumor-derived exosomes and
their role in cancer progression. Adv Clin Chem. 74:103–141. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Szabo G and Momen-Heravi F: Extracellular
vesicles in liver disease and potential as biomarkers and
therapeutic targets. Nat Rev Gastroenterol Hepatol. 14:455–466.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Jin X, Chen Y, Chen H, Fei S, Chen D, Cai
X, Liu L, Lin B, Su H, Zhao L, et al: Evaluation of tumor-derived
exosomal miRNA as potential diagnostic biomarkers for early-stage
non-small cell lung cancer using next-generation sequencing. Clin
Cancer Res. 23:5311–5319. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Xue M, Chen W, Xiang A, Wang R, Chen H,
Pan J, Pang H, An H, Wang X, Hou H and Li X: Hypoxic exosomes
facilitate bladder tumor growth and development through
transferring long non-coding RNA-UCA1. Mol Cancer. 16:1432017.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Logozzi M, Angelini DF, Iessi E, Mizzoni
D, Di Raimo R, Federici C, Lugini L, Borsellino G, Gentilucci A,
Pierella F, et al: Increased PSA expression on prostate cancer
exosomes in in vitro condition and in cancer patients. Cancer Lett.
403:318–329. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Puhka M, Takatalo M, Nordberg ME, Valkonen
S, Nandania J, Aatonen M, Yliperttula M, Laitinen S, Velagapudi V,
Mirtti T, et al: Metabolomic profiling of extracellular vesicles
and alternative normalization methods reveal enriched metabolites
and strategies to study prostate cancer-related changes.
Theranostics. 7:3824–3841. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Johnson SM, Dempsey C, Chadwick AL,
Harrison S, Liu J, Di Y, McGinn OJ, Fiorillo M, Sotgia F, Lisanti
MP, et al: Metabolic reprogramming of bone marrow stromal cells by
leukemic extracellular vesicles in acute lymphoblastic leukemia.
Blood. 128:453–456. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Yu X, Harris SL and Levine AJ: The
regulation of exosome secretion: A novel function of the p53
protein. Cancer Res. 66:4795–4801. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wu F, Yin Z, Yang L, Fan J, Xu J, Jin Y,
Yu J, Zhang D and Yang G: Smoking induced extracellular vesicles
release and their distinct properties in non-small cell lung
cancer. J Cancer. 10:3435–3443. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Zhou W, Woodson M, Sherman MB, Neelakanta
G and Sultana H: Exosomes mediate Zika virus transmission through
SMPD3 neutral sphingomyelinase in cortical neurons. Emerg Microbes
Infect. 8:307–326. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Baumann K: Making more exosomes. Nat Rev
Mol Cell Biol. 22:2422021. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Adams SD, Csere J, D'Angelo G, Carter EP,
Romao M, Arnandis T, Dodel M, Kocher HM, Grose R, Raposo G, et al:
Centrosome amplification mediates small extracellular vesicle
secretion via lysosome disruption. Curr Biol. 31:1403–1416.e7.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Lin Y, Dong H, Deng W, Lin W, Li K, Xiong
X, Guo Y, Zhou F, Ma C, Chen Y, et al: Evaluation of salivary
exosomal chimeric GOLM1-NAA35 RNA as a potential biomarker in
esophageal carcinoma. Clin Cancer Res. 25:3035–3045. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Hu Y, Qi C, Liu X, Zhang C, Gao J, Wu Y,
Yang J, Zhao Q, Li J, Wang X and Shen L: Malignant ascites-derived
exosomes promote peritoneal tumor cell dissemination and reveal a
distinct miRNA signature in advanced gastric cancer. Cancer Lett.
457:142–150. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Manier S, Liu CJ, Avet-Loiseau H, Park J,
Shi J, Campigotto F, Salem KZ, Huynh D, Glavey SV, Rivotto B, et
al: Prognostic role of circulating exosomal miRNAs in multiple
myeloma. Blood. 129:2429–2436. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Ye SB, Zhang H, Cai TT, Liu YN, Ni JJ, He
J, Peng JY, Chen QY, Mo HY, Jun Cui, et al: Exosomal miR-24-3p
impedes T-cell function by targeting FGF11 and serves as a
potential prognostic biomarker for nasopharyngeal carcinoma. J
Pathol. 240:329–340. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
He L, Ping F, Fan Z, Zhang C, Deng M,
Cheng B and Xia J: Salivary exosomal miR-24-3p serves as a
potential detective biomarker for oral squamous cell carcinoma
screening. Biomed Pharmacother. 121:1095532020. View Article : Google Scholar
|
|
115
|
Zou X, Zhu D, Zhang H, Zhang S, Zhou X, He
X, Zhu J and Zhu W: MicroRNA expression profiling analysis in serum
for nasopharyngeal carcinoma diagnosis. Gene. 727:1442432020.
View Article : Google Scholar
|
|
116
|
Xu JF, Wang YP, Zhang SJ, Chen Y, Gu HF,
Dou XF, Xia B, Bi Q and Fan SW: Exosomes containing differential
expression of microRNA and mRNA in osteosarcoma that can predict
response to chemotherapy. Oncotarget. 8:75968–75978. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Wang J, Li W, Zhang L, Ban L, Chen P, Du
W, Feng X and Liu BF: Chemically edited exosomes with dual ligand
purified by microfluidic device for active targeted drug delivery
to tumor cells. ACS Appl Mater Interfaces. 9:27441–27452. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
He C, Zheng S, Luo Y and Wang B: Exosome
theranostics: Biology and translational medicine. Theranostics.
8:237–255. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Charoenviriyakul C, Takahashi Y, Morishita
M, Matsumoto A, Nishikawa M and Takakura Y: Cell type-specific and
common characteristics of exosomes derived from mouse cell lines:
Yield, physicochemical properties, and pharmacokinetics. Eur J
Pharm Sci. 96:316–322. 2017. View Article : Google Scholar
|
|
120
|
Kamerkar S, LeBleu VS, Sugimoto H, Yang S,
Ruivo CF, Melo SA, Lee JJ and Kalluri R: Exosomes facilitate
therapeutic targeting of oncogenic KRAS in pancreatic cancer.
Nature. 546:498–503. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Pi F, Binzel DW, Lee TJ, Li Z, Sun M,
Rychahou P, Li H, Haque F, Wang S, Croce CM, et al: Nanoparticle
orientation to control RNA loading and ligand display on
extracellular vesicles for cancer regression. Nat Nanotechnol.
13:82–89. 2018. View Article : Google Scholar :
|
|
122
|
Bellavia D, Raimondo S, Calabrese G, Forte
S, Cristaldi M, Patinella A, Memeo L, Manno M, Raccosta S, Diana P,
et al: Interleukin 3-receptor targeted exosomes inhibit in vitro
and in vivo chronic myelogenous leukemia cell growth. Theranostics.
7:1333–1345. 2017. View Article : Google Scholar :
|
|
123
|
Bliss SA, Sinha G, Sandiford OA, Williams
LM, Engelberth DJ, Guiro K, Isenalumhe LL, Greco SJ, Ayer S, Bryan
M, et al: Mesenchymal stem cell-derived exosomes stimulate cycling
quiescence and early breast cancer dormancy in bone marrow. Cancer
Res. 76:5832–5844. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Kim SM, Yang Y, Oh SJ, Hong Y, Seo M and
Jang M: Cancer-derived exosomes as a delivery platform of
CRISPR/Cas9 confer cancer cell tropism-dependent targeting. J
Control Release. 266:8–16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Agrawal AK, Aqil F, Jeyabalan J, Spencer
WA, Beck J, Gachuki BW, Alhakeem SS, Oben K, Munagala R, Bondada S
and Gupta RC: Milk-derived exosomes for oral delivery of
paclitaxel. Nanomedicine. 13:1627–1636. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Cheng L, Wang Y and Huang L: Exosomes from
M1-Polarized macrophages potentiate the cancer vaccine by creating
a pro-inflammatory microenvironment in the lymph node. Mol Ther.
25:1665–1675. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Rao Q, Zuo B, Lu Z, Gao X, You A, Wu C, Du
Z and Yin H: Tumor-derived exosomes elicit tumor suppression in
murine hepatocellular carcinoma models and humans in vitro.
Hepatology. 64:456–472. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Zuo B, Qi H, Lu Z, Chen L, Sun B, Yang R,
Zhang Y, Liu Z, Gao X, You A, et al: Alarmin-painted exosomes
elicit persistent antitumor immunity in large established tumors in
mice. Nat Commun. 11:17902020. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Lu Z, Zuo B, Jing R, Gao X, Rao Q, Liu Z,
Qi H, Guo H and Yin H: Dendritic cell-derived exosomes elicit tumor
regression in autochthonous hepatocellular carcinoma mouse models.
J Hepatol. 67:739–748. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Morishita M, Takahashi Y, Matsumoto A,
Nishikawa M and Takakura Y: Exosome-based tumor antigens-adjuvant
co-delivery utilizing genetically engineered tumor cell-derived
exosomes with immunostimulatory CpG DNA. Biomaterials. 111:55–65.
2016. View Article : Google Scholar : PubMed/NCBI
|