Introduction
Retinoblastoma (RB) is the most common type of
primary intraocular malignant tumor that occurs either genetically
in children or sporadically in adults (1). Apart from its occult onset, RB can
easily metastasize, and can rapidly fill much of the eyeball and
can subsequently spread to the other parts of body, such as the
brain and lymph nodes (2).
Although marked improvements have been made in the treatment of RB,
including enucleation, radiation, focal techniques and systemic
chemotherapy, RB is associated with a high metastatic rate and poor
prognosis (3,4). Therefore, there is an urgent need to
explore the underlying mechanisms involved in RB tumorigenesis,
development and prognosis.
MicroRNAs (miRNAs or miRs), normally containing
~21-25 nucleotides, are a class of small non-coding RNA molecules
involved in the regulation of gene expression by targeting mRNAs
for translational repression or cleavage (5,6).
There is increasing evidence to indicate that miRNAs play critical
roles in regulating cell apoptosis, proliferation, invasion and
migration (7). A number of
studies have indicated the role of miRNAs in various types of
tumors (8). MicroRNA-153-3p
(miR-153), an extensively studied miRNA, has been reported to serve
as a tumor suppressor in a variety of cancer types, such as breast
cancer (9), gastric cancer
(10), osteosarcoma (11) and lung cancer (12). For example, miR-153 has been shown
to target the inhibitor of growth protein 2 to inhibit acute
lymphoblastic leukemia cell proliferation, migration and invasion
(13). It has also been reported
that miR-153 can inhibit the MCL1 gene, thus suppressing the
progression of ovarian carcinoma in vitro and in vivo
(14). In addition, miR-153 has
been reported to inhibit the migration and the tube formation of
endothelial cells by blocking the paracrine of angiopoietin 1 in
breast cancer cells (15).
However, the function of miR-153 in RB remains unknown.
In the present study, miR-153 expression was
analyzed in RB tissues and cell lines by reverse
transcription-quantitative PCR (RT-qPCR) assays. In vitro
experiments were performed to investigate the functional role of
miR-153 in RB cells and the underlying mechanisms. The findings of
these experiments suggested that miR-15 may be a potential
diagnostic and therapeutic target for RB treatment.
Materials and methods
Tissue samples
A total of 50 RB tissues samples were obtained from
patients who received enucleation or enucleation + chemotherapy ±
radiation therapy at the Department of Ophthalmology, the First
People's Hospital of Shangqiu, Shangqiu, China between February,
2017 and November, 2018. A total of 10 normal retinas were obtained
from patients who had passed away due to conditions other than
ophthalmologic diseases at the First People's Hospital of Shangqiu.
Written consent for tissue donation for research purposes was
obtained from the donor or family members prior to tissue
collection. The present study was approved by the Research Ethics
Committee of the First People's Hospital of Shangqiu. The samples
were snap-frozen in liquid nitrogen and stored at −80°C.
RT-qPCR
Total RNA was extracted from tissues and cells by
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). cDNA was synthesized using the PrimeScript RT reagent kit
(Promega Corporation). For the detection of miR-153, qPCR was
conducted using the MicroRNAs Quantitation PCR kit [Sangon Biotech
(Shanghai) Co., Ltd.]. U6 was used as an internal control for the
detection of miR-153 expression. The sequences of the primers used
for miR-153 and U6, as well as those for insulin-like growth factor
1 receptor (IGF1R) and GAPDH are listed in Table I. The thermocycling conditions
were as follows: 50°C for 2 min and 95°C for 10 min, followed by 40
cycles of 95°C for 15 sec and 60°C for 10 min. Fold changes in the
expression of each gene were calculated using the 2−∆∆Cq
method (16).
 | Table ISequences of the primers used in the
present study. |
Table I
Sequences of the primers used in the
present study.
| Gene | Primer
sequence |
|---|
| miR-153-3p | F:
5′-ACACTCCAGCTGGGTTGCATAGTCACAAA-3′ |
| R:
5′-CAGTGCGTGTCGTGGAGT-3′ |
| U6 | F:
5′-TCGCACAGACTTGTGGGAGAA-3′ |
| R:
5′-CGCACATTAAGCCTCTATAGTTACTAGG-3′ |
| IGF1R | F:
5′-GGGGCTCCTGTTTCTCTCC-3′ |
| R:
5′-GCCTTGGAGATGAGCAGGAT-3′ |
| GAPDH | F:
5′-AGCTTGTCATCAACGGGAAG-3′ |
| R:
5′-TTTGATGTTAGTGGGGTCTCG-3′ |
Cell lines and cell culture
WERI-RB-1 (ATCC® HTB-169) and Y79
(ATCC® HTB-18), a normal retinal epithelial cell line
(ARPE-19) (ATCC® CRL-2302) cells were obtained from
American Type Culture Collection (ATCC), while the RB cell line
SO-RB50 was obtained from the Zhongshan Ophthalmic Center, Sun
Yat-sen University (Zhongshan, China). All the cells were cultured
in Dulbecco's modified Eagle's medium (DMEM, HyClone; Cytiva)
supplemented with 10% fetal bovine serum (HyClone; Cytiva), 100
U/ml penicillin and 100 μg/ml streptomycin at 37°C in a 5%
CO2 atmosphere.
Cell transfection
miR-153 mimics (5′-UUGCAUAGUCACAAAAGUGAUC-3′),
mimics negative control (NC) (5′-UUCUCCGAACGUGUCACGUTT-3′), miR-153
inhibitor (5′-GAUCACUUUUGUGACUAUGCAA-3′) and inhibitor NC
(5′-CAGUACUUUUGUGUAGUACAA-3′) were obtained from Shanghai
GenePharma Co., Ltd. pcDNA3.1-IGF1R, pcDNA3.1 vector, as well as
small interfering RNA (si-RNA) specific for IGF1R (si-IGF1R)
(5′-AGGCGGGCTTCCGGGAGGTCTCCTT-3′) or si-Scramble
(5′-GGAAGAGGTGGAGGCTCGCTCGCGG-3′) were obtained from Invitrogen;
Thermo Fisher Scientific, Inc.. The WERI-RB-1 and Y79 cells
(5×105/well) were seeded in a six-well plate overnight,
and the cells were then transfected with miR-153 mimics (20 nM),
mimics NC (20 nM), miR-153 inhibitor (20 nM), inhibitor NC (20 nM),
pcDNA-IGF1R (2 μg) or si-IGF1R (100 nM) (Shanghai GenePharma
Co., Ltd.) using Lipofectamine 2000® (Invitrogen; Thermo
Fisher Scientific, Inc.). At 48 h following transfection, cells
were harvested, and protein and RNA were then extracted for
analyses.
Microarray datasets
The miRNA dataset (GSE7072) was searched and
downloaded from the Gene Expression Omnibus (GEO) database
(https://www.ncbi.nlm.nih.gov/geo/).
GSE7072 was analyzed through the GPL4879 platform (Agilent-019118
Human miRNA Microarray 2.0) and consisted of three samples.
Differentially expressed miRNAs (DEmiRNAs) between normal and RB
samples were screened based on GEO2R (www.ncbi.nlm.nih.gov/geo/geo2r/), an interactive web
tool. DE-miRNAs were then identified based on the fold change in
expression. A heatmap of DE-miRNAs was created using a method of
hierarchical clustering by GeneSpring GX, version 7.3 (Agilent
Technologies, Inc.).
Cell viability assay
At 48 h following transfection, cell viability in
96-well plates was evaluated using the Cell Counting Kit-8 (CCK8;
Dojindo Laboratories, Inc.) assay. Briefly, 10 μl CCK-8
solution (Dojindo Laboratories, Inc). were added to each well and
incubated at 37°C for a further 2 h. The absorbance at 560 nm was
measured using a microplate reader (Bio-Rad Laboratories, Inc.).
The experiments were repeated three times.
Detection of apoptosis by flow
cytometry
Cell apoptosis was evaluated using an Annexin
V/propidium iodide (PI) apoptosis-detection kit (KeyGen BioTech,
Co., Ltd.) according to the manufacturer's protocols. Briefly, at
48 h following trans- fection, cells were centrifuged at 4°C and
washed with PBS, and stained with Annexin V and PI for 15 min at
room temperature in the dark. The stained cells were then analyzed
with EPICS XL-MCL FACScan (Becton-Dickinson and Company).
Bioinformatics analysis
miRNA target prediction tools, including PicTar
version 2007 (https://pictar.mdc-berlin.de/) and TargetScan Release
7.0 (http://targetscan.org/) were used to
search for the putative targets of miR-153.
Vector construction and luciferase
assays
Luciferase reporters were generated based on the
Firefly luciferase expressing vector pGL3-control (Promega
Corporation). The 3′-UTR fragment of the IGF1R gene and its mutant
of the theoretical miR-153 binding site were cloned into the pGL3
control vector (Promega Corporation) to form the reporter vector,
wild-type (wt) and mutant-type (mut) of IGF1R, respectively. To
construct pGL3-IGF1R-3′UTR, a partial 3′UTR of the IGF1R segment of
human IGF1R mRNA containing the putative miR-153 binding sites was
amplified and cloned into the vector pGL3-control. Mutations within
potential miR-153 binding sites were introduced using the
QuikChange Site-Directed Mutagenesis kit (Life Technologies; Thermo
Fisher Scientific, Inc.). When the Y79 cells grew to 60-70%
confluency, the cells were co-transfected with 100 ng luciferase
plasmid and 50 ng Renilla luciferase plasmid along with 100
ng miR-153 mimics/inhibitor or miR-NC using Lipofectamine
2000® (Invitrogen; Thermo Fisher Scientific, Inc.).
Following incubation for 48 h at 37°C, the luciferase activity was
assessed using the dual luciferase reporter kit (Beyotime Institute
of Biotechnology). Renilla activity was used to normalize
Firefly luciferase activity.
Western blot analysis
At 48 h following transfection, the total protein
from cells was obtained using RIPA lysis buffer (Santa Cruz
Biotechnology, Inc.) and quantified using a BCA protein assay kit
(Pierce; Thermo Fisher Scientific, Inc.). The proteins in the
lysates were separated by 10% SDS-PAGE gels and transferred to PVDF
membranes (GE Healthcare; Cytiva). After being blocked with a 5%
skim milk solution for 1 h at room temperature, the membranes were
incubated with specific primary antibodies at 4°C overnight,
including E-cadherin (cat. no. ab40772, 1:1,000 dilution),
N-cadherin (cat. no. ab202030, 1:1,000 dilution), Vimentin (cat.
no. ab45939, 1:1,000 dilution), fibronectin (cat. no. ab2413,
1:1,000 dilution), Snail (cat. no. ab216347, 1:1,000 dilution),
Twist (cat. no. ab50581, 1:1,000 dilution), zinc finger E-box
binding homeobox 1 (ZEB1; cat. no. ab203829, 1:1,000 dilution) (all
from Abcam), IGF1R (cat. no. sc81464, 1:1,000 dilution), PI3K (cat.
no. S365290, 1:1,000 dilution), phosphorylated (p)-AKT (cat. no.
sc-514032, 1:1,000 dilution), p-MEK1/2 (cat. no. sc-81503, 1:1,000
dilution) (both from Santa Cruz Biotechnology, Inc.), p-Raf/1
(Ser338, 1:1,000; cat. no. 9427; Cell Signaling Technology, Inc.)
and β-actin (cat no. sc-84322, Santa Cruz Biotechnology, Inc.,
1:1,000 dilution). Subsequently, the mouse anti-rabbit IgG-HRP
secondary antibody (cat no. sc2537, Santa Cruz Biotechnology, Inc.,
1:1,000 dilution) was added to the membranes followed by incubation
for 2 h at room temperature. The protein bands were visualized
using an ECL detection system (Thermo Fisher Scientific, Inc.).
Semi-quantification was performed using ImageJ version 1.46
(NIH).
Immunofluorescence analysis and
immunohistochemistry (IHC)
For immunofluorescence, the cells on the coverslips
were fixed with 4% paraformaldehyde for 10 min at 48 h following
transfection, and subsequently, the cells were incubated at 4°C
overnight in a solution containing primary antibodies specific for
caspase-3 (cat. no. sc-7272, Santa Cruz Biotechnology, Inc.,
1:200). The cells were then stained with mouse anti-goat IgG-FITC
(cat. no. sc2356, Santa Cruz Biotechnology, Inc., 1:200). The cells
were counterstained with DAPI for 15 min at room temperature to
identify the nuclei and imaged with a confocal laser-scanning
microscope (Axiovert 200 M, Zeiss GmbH).
For IHC, the expression of caspase-3 was evaluated
by IHC staining as previously described (17) with the following primary
antibodies: Caspase-3 (cat. no. sc-7272, Santa Cruz Biotechnology,
Inc., 1:100). Samples were photographed under a light Leica DMD 108
microscope (Leica Microsystems GmbH).
Transwell assay
Briefly, a total of 3×104 transfected
cells in DMEM without serum were added to the top Matrigel-coated
chambers (pore size, 8 μm; Corning, Inc.), while DMEM with
20% FBS (600 μl) was then added to the lower chamber.
Following 24 h of incubation at 37°C, cells on the upper side of
each membrane were cleaned with a cotton swab. The membranes were
then fixed in 20% methanol for 15 min and stained with 0.2% crystal
violet (Sigma-Aldrich; Merck KGaA, Germany) at room temperature for
30 min. A total of five visual fields of each insert were randomly
selected and photographed under a light microscope (Olympus
Corporation) at ×200 magnification.
Wound healing assay
At 48 h following transfection, the cell layer at
80-90% confluency was gently and slowly scratched with a new 200
μl pipette tip across the center of the well. The cells were
then washed twice with phosphate-buffered saline (PBS) and
incubated in free-serum DMEM. The wound healing images
(magnification, ×200) were obtained at the time of wounding (time
0), and at 24 and 48 h after scratching. The wound closure process
was observed and photographed under a microscope (Olympus
Corporation).
Statistical analysis
The SPSS 23.0 software package (SPSS Inc.) was
applied to analyze the data. All data are presented as the mean ±
SD. An unpaired Student's t-test was performed for two-group
comparisons. Comparisons between multiple groups were analyzed by
one-way ANOVA followed by Tukey's post hoc test. The correlation
between IGF1R and miR-153 expression was analyzed using Pearson's
correlation coefficient. The Chi-squared test was used for
association analysis between clinicopathological features of
patients with retinoblastoma and miR-153-3p expression profiles.
The Kaplan-Meier method was applied to calculate the 5-year overall
survival (OS), and the log-rank test was used to evaluate whether
there were statistically significant differences in OS. P<0.05
was considered to indicate a statistically significant
difference.
Results
miR-153 expression is downregulated in RB
tissues and cell lines
To identify RB-associated miRNAs, the differentially
expressed miRNAs (DEmiRNAs) in the GSE dataset (GSE7072) were
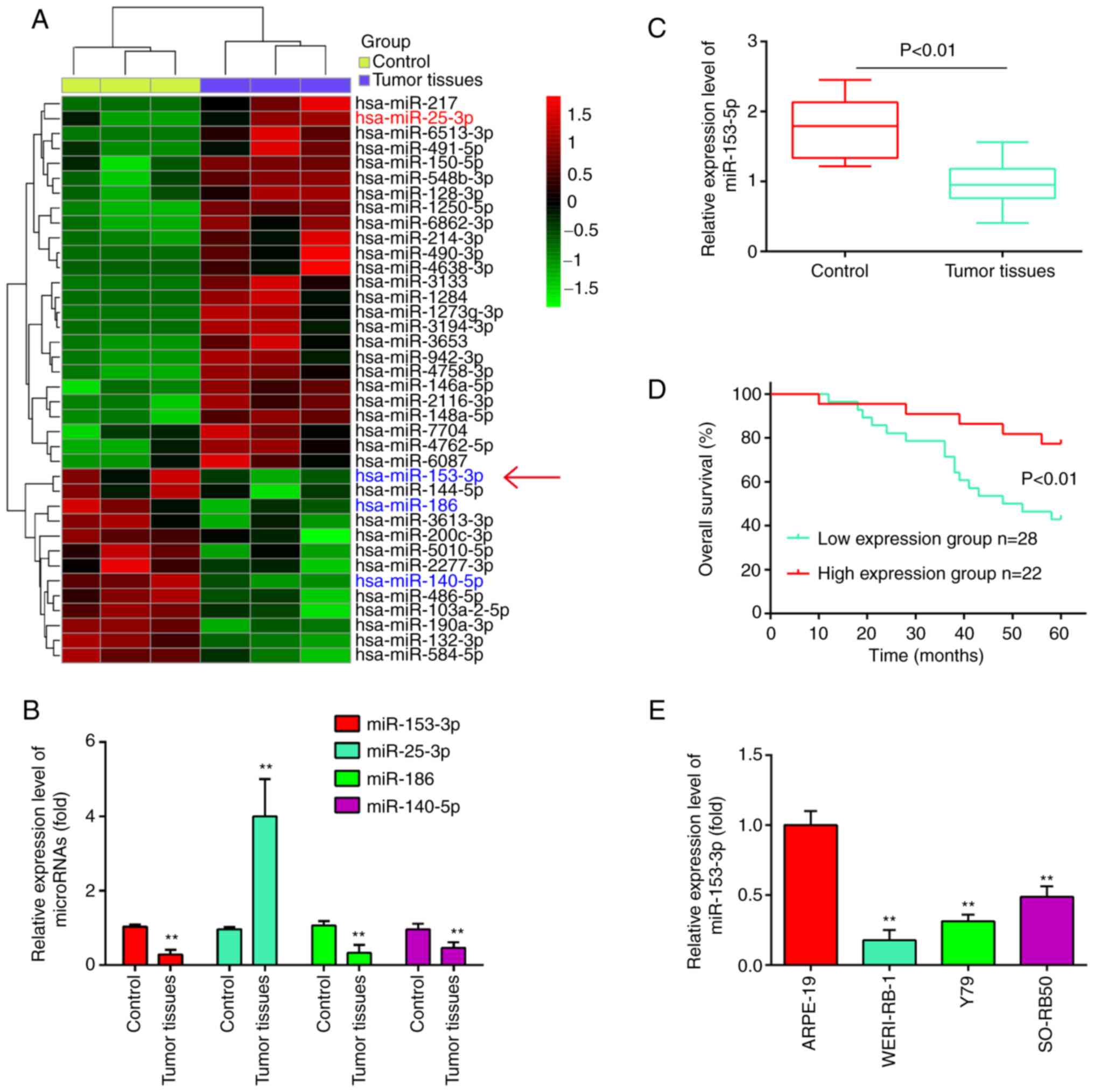
analyzed using bioinformatics analysis. As shown in Fig. 1A, a number of miRNAs were found to
be differentially expressed between these RB tissues and normal
retinal tissues. Out of these miRNAs, it was found that miR-25-3p
was upregulated, while miR-186 and miR-140-5p were downregulated in
the RB tissues (Fig. 1B), which
was consistent with the findings of previous studies (18-20). Of particular interest, miR-153 was
one of the most significantly downregulated miRNA in the RB tissues
(Fig. 1B). Previous studies have
indicated that miR-153 functions as a tumor suppressor in several
malignancies, such as non-small cell lung cancer (NSCLC) (21) and ovarian carcinoma (OC) (14). Therefore, miR-153 was selected for
further analyses.
Using RT-qPCR assay, miR-153 was detected in 50 RB
tissues and 10 normal retinal tissues. As shown in Fig. 1C, the results revealed that the
expression levels of miR-153 were significantly lower in the tumor
tissues than those in the normal tissues (P<0.01). Moreover, the
patients with RB were divided into a high miR-153 expression group
and a low expression group based on the median value of miR-153
expression as a cut-off point. As shown in Fig. 1D, patients with a low expression
of miR-153 had an evidently shorter overall survival time than
those patients with a high expression of miR-153 (P<0.01). In
addition, the expression levels of miR-153 in RB cancer cell lines
were measured by RT-qPCR. Compared with that in ARPE-19 cells,
miR-153 expression was significantly downregulated in the three RB
cancer cell lines (Fig. 1E).
Based on these findings, miR-153 may be involved in the development
of RB.
Subsequently, the association between miR-153
expression and the clinicopathological characteristics of patients
with RB was analyzed. As shown in Table II, a low expression of miR-153
was significantly associated with the largest tumor base and
differentiation; however, no significant association was observed
between the miR-153 expression levels and the patient age, sex,
tumor enucleation location, T classification and clinical stage. On
the whole, these data indicated that miR-153 expression in RB tumor
tissues may serve as a diagnostic and prognostic marker for RB.
| Table IIAssociation between miR-153-3p and
clinicopathological features of patients with retinoblastoma. |
Table II
Association between miR-153-3p and
clinicopathological features of patients with retinoblastoma.
| Feature | Total n=50 | miR-153-3p
expression
| P-value |
|---|
| High | Low |
|---|
| Sex | | | | 0.4741 |
| Male | 21 | 8 | 13 | |
| Female | 29 | 14 | 15 | |
| Age at presentation
(years) | | | | 0.1212 |
| ≤5 | 31 | 11 | 20 | |
| >5 | 19 | 11 | 8 | |
| Tumor enucleation
location | | | | 0.1443 |
| Right | 24 | 8 | 16 | |
| Left | 26 | 14 | 12 | |
| Largest tumor base
(mm) | | | | 0.0199a |
| ≤15 | 32 | 18 | 14 | |
| >15 | 18 | 4 | 14 | |
| T
classification | | | | 0.1830 |
| T1-2 | 28 | 10 | 18 | |
| T3-4 | 22 | 12 | 10 | |
| Clinical stage | | | | 0.2008 |
| I-II | 20 | 11 | 9 | |
| III-IV | 30 | 11 | 19 | |
|
Differentiation | | | | 0.0025b |
| Well and
moderate | 16 | 12 | 4 | |
| Poor | 34 | 10 | 24 | |
miR-153 overexpression inhibits RB cell
proliferation and promotes apoptosis
To examine the effects of miR-153 on RB cell
proliferation and apoptosis, miR-153 mimics and mimics NC were
transfected into the WERI-RB-1 and Y79 cell lines, which exhibited
the lowest levels of miR-153 expression among the four cell lines
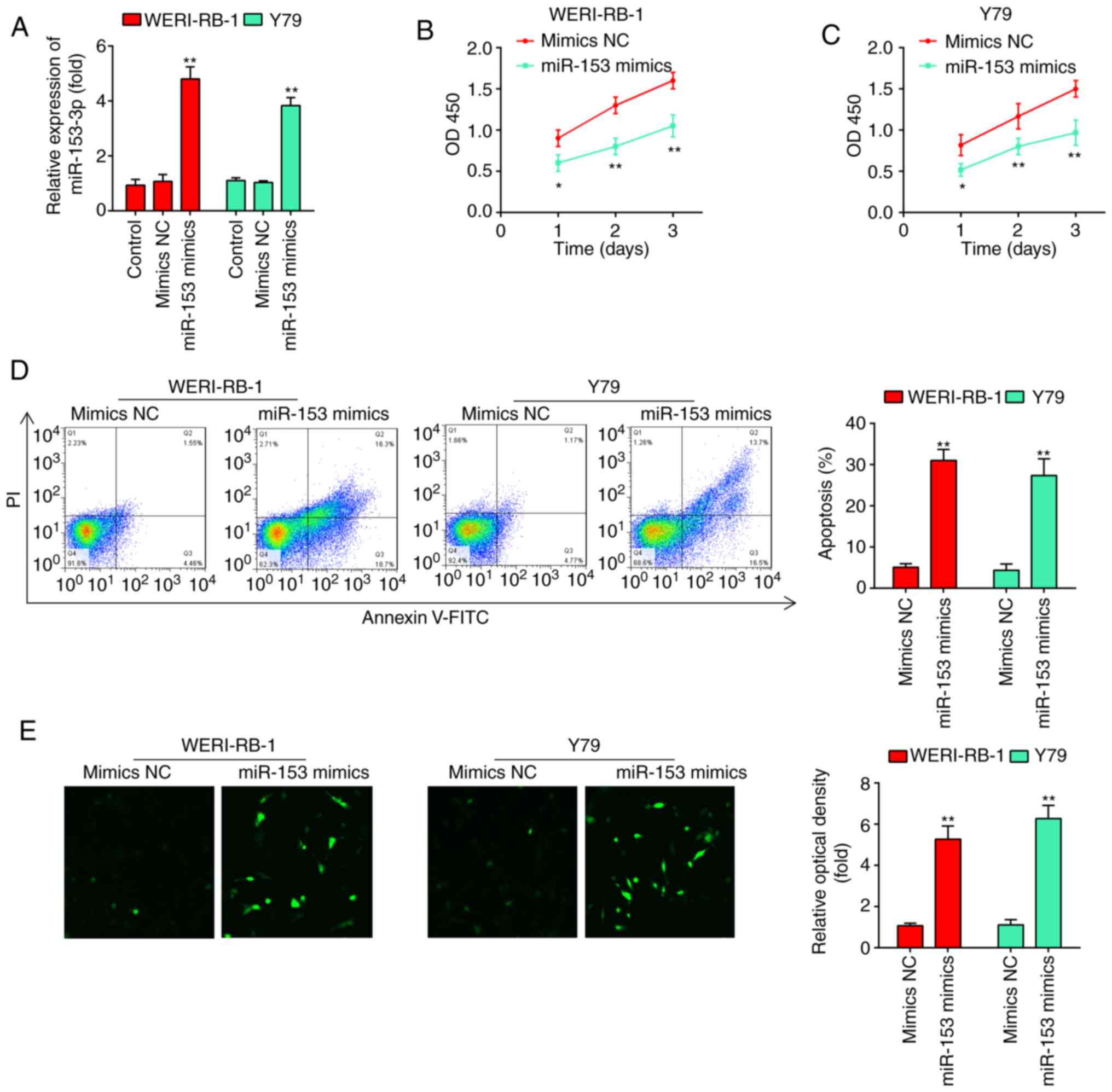
examined. As shown in Fig. 2A,
miR-153 expression was significantly increased compared with that
in the mimics NC-transfected cells (P<0.01). According to the
results of the CCK-8 assay, miR-153 overexpression significantly
inhibited the proliferation of the WERI-RB-1 and Y79 cells compared
with that in the mimics NC group (Fig. 2B and C). Flow cytometric analyses
were subsequently performed to examine the effects of miR-153 on
cell apoptosis. It was found that miR-153 overexpression
significantly increased the percentage apoptosis in comparison with
the mimics NC group (WERI-RB-1 cells: 30.8±1.49 vs. 4.76±0.191%,
P=0.01; Y79 cells: 26.8±1.13 vs. 3.36±0.179%, P=0.01; Fig. 2D). In addition, whether miR-153
modulates the expression of apoptosis-associated proteins, such as
caspase-3 was determined by immunofluorescence. The expression
level of caspase-3 in the cells transfected with miR-153 mimics was
evidently increased compared with that in the mimics NC-transfected
cells (Fig. 2E). Taken together,
these results suggest that miR-153 inhibits the proliferation and
induces the apoptosis of RB cell lines.
Overexpression of miR-153 inhibits
invasion, migration and the epithelial-mesenchymal transformation
(EMT) process in RB cells
It is well known that cell invasion, migration and
the EMT process are crucial for tumor development. Thus, in the
present study, Transwell and wound healing assays were performed to
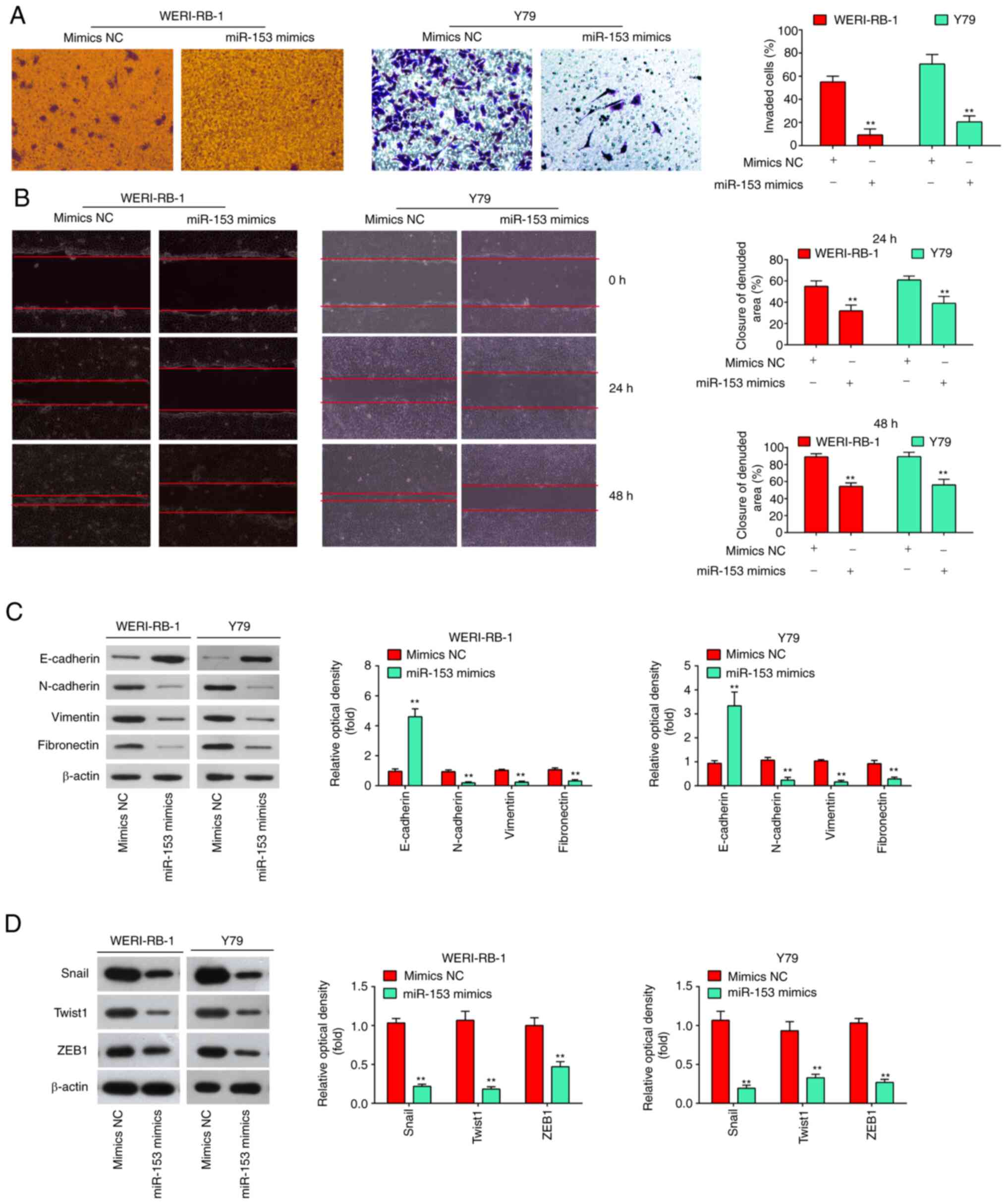
examine the effects of miR-153 on cell invasion and migration. It
was found that the overexpression of miR-153 in WERI-RB-1 and Y79
cells markedly suppressed cell invasion, compared with the cells
transfected with mimics NC (P<0.01, Fig. 3A). As was expected, the WERI-RB-1
and Y79 cells transfected with miR-153 mimics also exhibited a
decreased migration rate, compared with the mimics NC group
(P<0.01, Fig. 3B). E-cadherin
and vimentin are the major markers of EMT, and EMT plays an
important role in tumor metastases (22). Thus, the present study first
detected whether miR-153 modulates the expression of EMT markers.
It was observed that the level of E-cadherin (epithelial marker)
was significantly increased, while the levels of N-cadherin,
Vimentin, fibronectin (mesenchymal markers) were markedly decreased
in the miR-153 mimics group compared with the mimics NC group
(Fig. 3C). It is well known that
EMT-promoting transcription factors, such as Snail, Twist and ZEB1
play a key role in the regulation of the EMT process (23). Thus, the present study also
examined the effects of miR-153 on these EMT-promoting
transcription factors. As shown in Fig. 3D, the expression levels of Snail,
Twist and ZEB1 were significantly decreased in the miR-153 mimics
group compared with the mimics NC group. Taken together, these
findings demonstrated that miR-153 suppresses the invasion and
migration of RB cells in vitro probably by inhibiting the
EMT process.
IGF1R is a direct target of miR-153 in RB
cells
To explore the potential mechanisms through which
miR-153 functions as a tumor suppressor in RB, potential target
genes of miR-153 were screened using TargetScan 7.0 and PicTar.
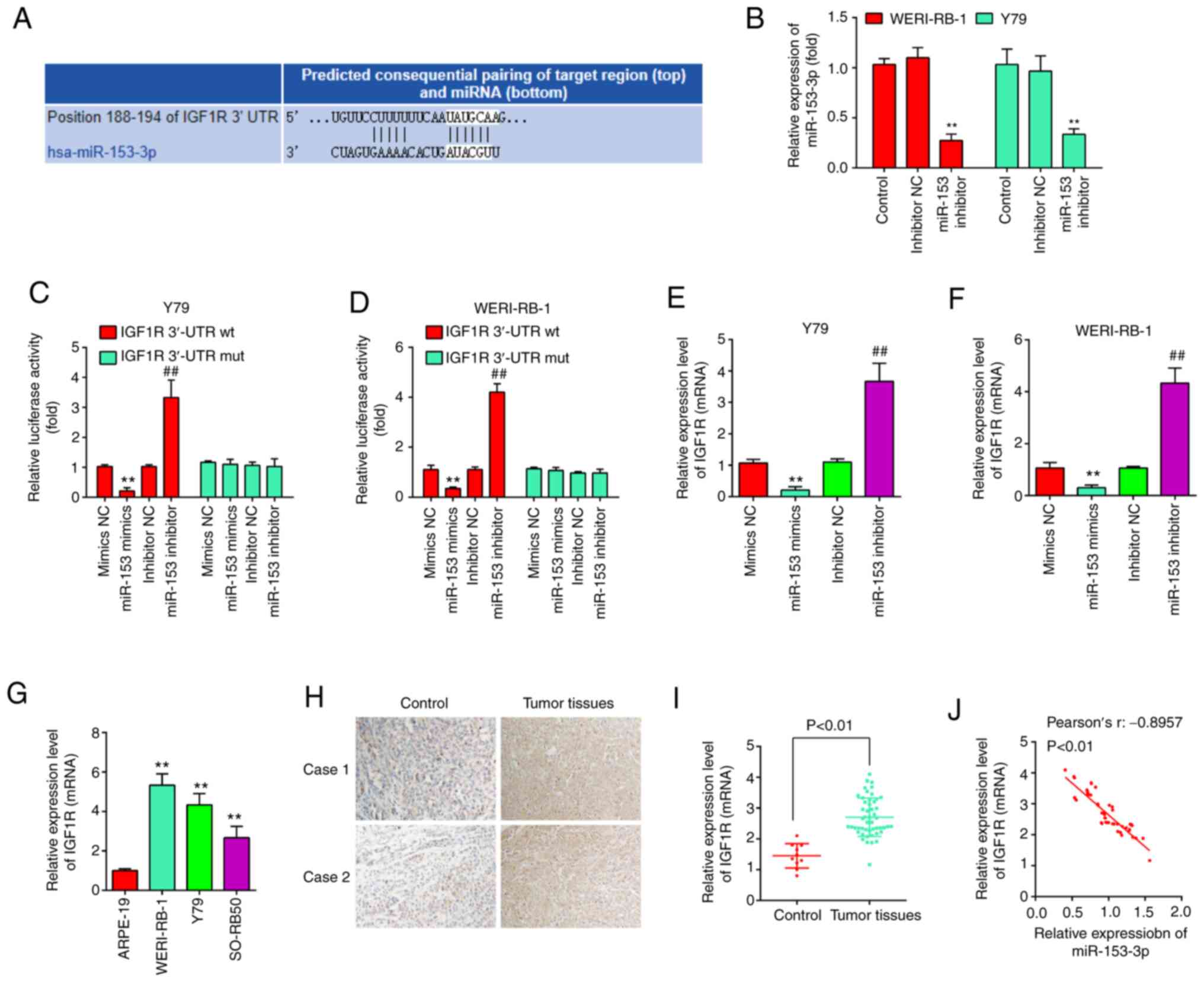
Bioinformatics analysis indicated that one potential binding site
for miR-153 was found in the 3′-UTR region of IGF1R mRNA (Fig. 4A). To verify whether miR-153
directly binds to IGF1R, a dual luciferase reporter assay was
performed using the Y79 and WERI-RB-1 cells. First, the WERI-RB-1
and Y79 cells were transfected with miR-153 inhibitor and
inhibitor-NC as a control, and RT-qPCR was then performed to
determine the transfection efficiency. The results revealed that
miR-153 expression was significantly decreased in the WERI-RB-1 and
Y79 cells following transfection with miR-153 inhibitor (Fig. 4B). The results of luciferase
reporter assay then revealed that transfection with miR-153 mimics
significantly inhibited the luciferase activity combined with the
IGF1R-3′-UTR wt reporter, while transfection with miR-153 inhibitor
led to an increase in luciferase activity; however, no significant
changes were observed using the IGF1R 3′-UTR mut reporter with
miR-153 mimics or inhibitor (Fig. 4C
and D). In addition, to confirm whether miR-153 regulate IGF1R
expression, qRT-PCR was performed. It was observed that IGF1R was
significantly reduced following miR-153 mimics transfection,
whereas increased by miR-153 inhibitor in Y79 cells and WERI-RB-1
cells (Fig. 4E and F). In
addition, the expression levels of IGF1R were detected in RB
tissues from two patients with RB by IHC assay. Compared with the
control group, IGF1R expression was markedly increased in RB
tissues (Fig. 4G). Moreover, the
IGF1R levels were also measured in RB cell lines, as well as in 50
pairs of RB tissues and normal tissue by RT-qPCR. The results
revealed that the IGF1R expression level was higher in both RB
cells lines and tumor tissues compared with the APRE-19 cells and
normal tissues (Fig. 4H and I).
Additionally, an obvious inverse correlation was observed between
the expression of IGF1R and miR-153 in the tissue samples
(r=-0.8957, P<0.01, Fig. 4J).
On the whole, these data suggest that IGF1R may be a functional
target of miR-153.
Knockdown of IGF1R reverses the
phenotypes induced by miR-153
As is known, the IGF1R oncogene is involved in the
carcinogenesis of several human cancers (24-26). In the present study, given that
IGF1R was upregulated in RB tissues, the possible effects of IGF1R
on RB cell biological behaviors were then investigated. The results
of RT-qPCR revealed that the expression of IGF1R was knocked down
by transfection of the WERI-RB-1 and Y79 cells with siRNA (Fig. 5A). CCK-8 assay revealed that the
knockdown of IGF1R led to a marked reduction in the viability of
WERI-RB-1 (Fig. 5B) and Y79 cells
(Fig. 5C). Additionally, as shown
by flow cytometric analysis, the cell apoptotic rate was
significantly increased following transfection with si-IGF1R (from
4.24±0.332% to 32.7±1.27% for WERI-RB-1 cells, and from 3.83±0.311%
to 25.1±1.31% for Y79 cells, P<0.01; Fig. 5D). As was expected, the invasive
abilities of the WERI-RB-1 and Y79 cells were also inhibited
following transfection with si-IGF1R (Fig. 5E). Collectively, these findings
suggested that miR-153 probably inhibited RB cell proliferation and
invasion, and promoted apoptosis by downregulating IGF1R.
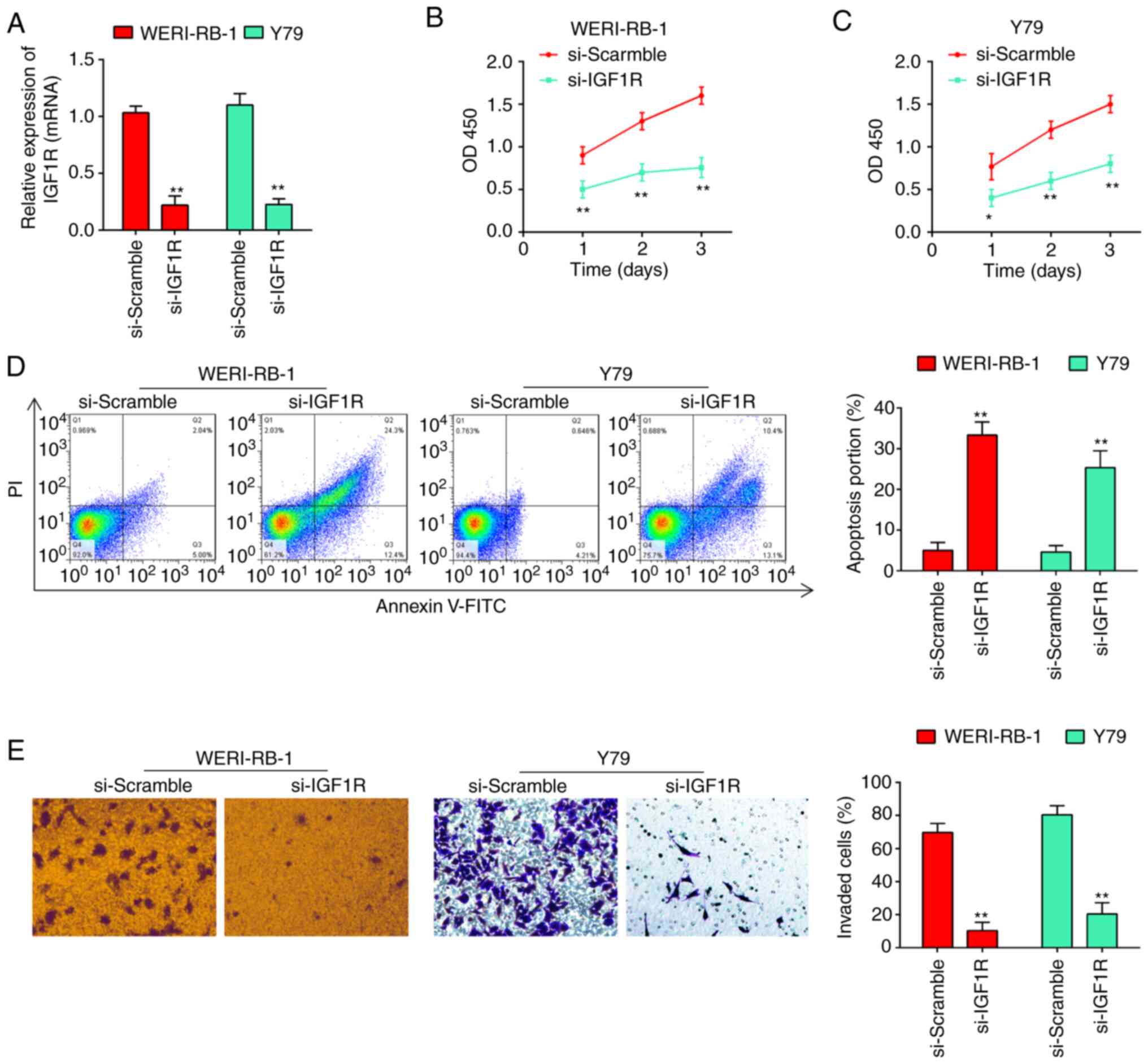 | Figure 5Silencing of IGF1R reverses the
phenotypes caused by miR-153. (A) siRNA efficiency of si-Scramble,
si-IGF1R, respectively, in WERI-RB-1 and Y79 cells. (B) Viability
of WERI-RB-1 cells was measured following transfection with
si-Scramble or si-IGF1R by CCK-8 assay. The results (mean ± SD) are
from three independent experiments (**P<0.01. vs.
si-Scramble). (C) Viability of Y79 cells was measured following
transfection with si-Scramble or si-IGF1R by CCK-8 assay. The
results (mean ± SD) are from three independent experiments
(*P<0.05, **P<0.01. vs. si-Scramble).
(D) Apoptosis of WERI-RB-1 and Y79 cells following transfection
with si-Scramble or si-IGF1R was determined by flow cytometric
analysis. Shown in the right panel are the percentages of apoptotic
cells (**P<0.01. vs. si-Scramble). Prior to flow
cytometry, the tested cells were firstly stained with Annexin
V-fluorescein isothiocyanate. (E) Silencing of IGF1R attenuated the
invasion of RB cells. Transwell assay was used to determine the
invasion of RB cells following transfection of si-Scramble, and
si-IGF1R, respectively, in WERI-RB-1 and Y79 cells. The results
(mean ± SD) are from three independent experiments
(**P<0.01. vs. si-Scramble). RB, retinoblastoma;
IGF1R, insulin-like growth factor 1 receptor. |
miR-153 inhibits retinoblastoma
proliferation, invasion and migration by targeting IGF1R
To explore whether IGF1R mediates the antitumor
effects of miR-153, the WERI-RB-1 and Y79 cells were co-transfected
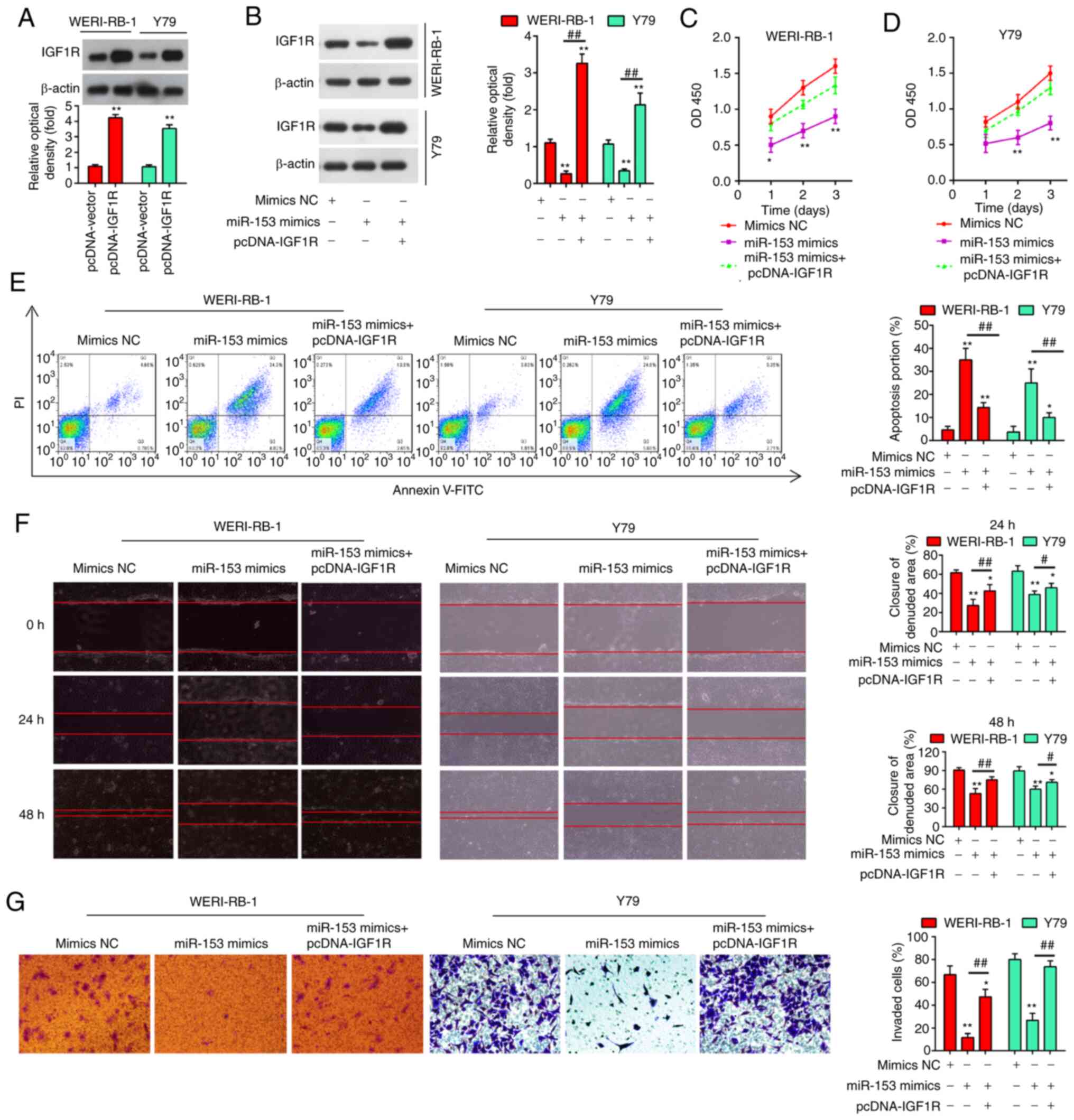
with miR-153 mimics and pcDNA-IGF1R. Firstly, the protein
expression of IGF1R was examined by western blot analysis in the
WERI-RB-1 and Y79 cells following pcDNA-IGF1R transfection. As
shown in Fig. 6A, in both the
cell lines tested, the expression levels of IGF1R were
significantly increased compared with the pcDNA-vector-transfected
cells. Moreover, IGF1R expression was significantly suppressed in
the cells transfected with miR-153 mimics compared with the mimics
NC-transfected cells (P<0.01), while the suppression of IGF1R
was fully reversed in the cells transfected with pcDNA-IGF1R, with
a significantly upregulated expression level of IGF1R (Fig. 6B). Consistently, the suppressive
effects of miR-153 on cell proliferation were evidently impaired by
transfection with pcDNA-IGF1R (Fig.
6C and D), while the apoptotic rate of the RB cells was
markedly decreased (Fig. 6E).
Additionally, the results of Transwell and wound healing assay
demonstrated that the suppression of RB cell invasion and migration
by miR-153 was also significantly attenuated by the overexpression
of IGF1R (Fig. 6F and G). These
findings thus demonstrate that IGF1R is involved in the
tumor-suppressive roles of miR-153 in RB cells.
miR-153-3p deactivates the PI3K/Akt and
MEK pathways in RB via IGF1R
IGF1R has recently been implicated in the activation
of the PI3K/Akt pathway and Raf1/MEK pathway, which affects the
carcinogenesis and development of numerous types of cancer
(27,28). Therefore, the present study
attempted to identify whether miR-153-3p is able to deactivate the
PI3K/AKT and Raf1/MEK pathways in RB cells. The results of western
blot analysis revealed that the expression levels of IGF1R, PI3K,
p-AKT, p-Raf/1 and p-MEK1/2 in the WERI-RB-1 and Y79 cells were
significantly decreased following miR-153 mimics transfection
(Fig. 7A and B), suggesting that
miR-153 may have inactivated the PI3K/Akt and MEK pathway in RB
cells by targeting IGF1-R. It was also found that a lower miR-153
expression in RB tissues was associated with an increased p-AKT and
p-MEK1/2 expression (P<0.01, Fig.
7C and D). Collectively, these data suggest that miR-153
probably suppresses RB cell growth and metastasis via the
suppression of the IGF1R/Raf/MEK and IGF1R/PI3K/AKT signaling
pathways.
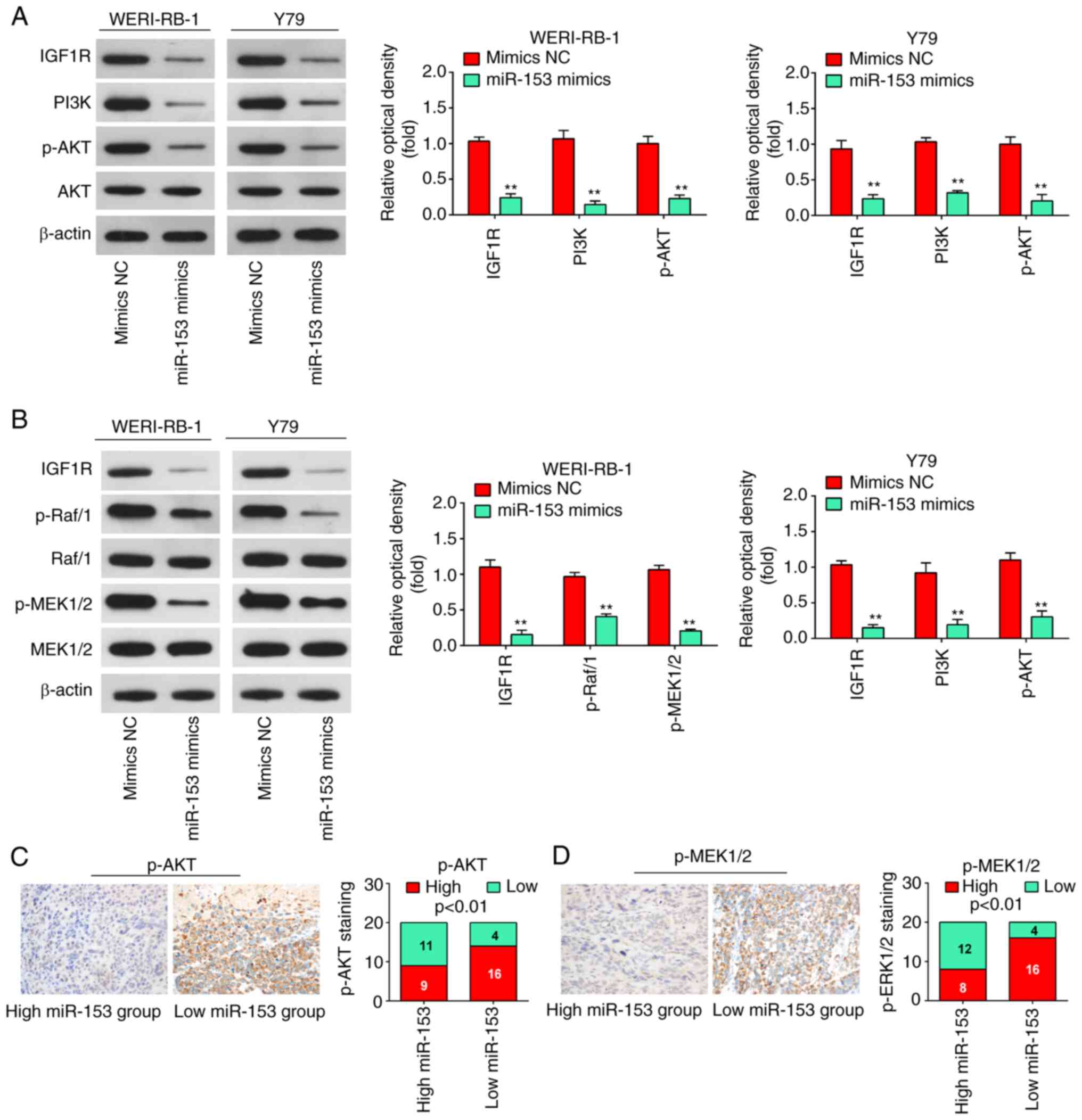 | Figure 7miR-153 deactivates the PI3K/AKT and
Raf/MEK pathway in WERI-RB-1 and Y79 cells by decreasing IGF1R
expression. (A) Following transfection, western blot analysis was
performed to measure IGF1R, PI3K, p-Akt and Akt expression. The
bands were semi-quantitatively analyzed by using ImageJ software,
normalized to β-actin density. **P<0.01 vs. mimics NC
group. (B) Following transfection, western blot analysis was
performed to measure IGF1R, Raf/1, p-Raf/1, p-MEK1/2, and MEK1/2
expression. The bands were semi-quantitatively analyzed by using
ImageJ software, normalized to β-actin density.
**P<0.01 vs. mimics NC group. (C)
Immunohistochemistry was conducted to detect p-AKT in RB tissues
with high or low miR-153 expression. Bar graphs demonstrated a
significant inverse association between miR-153 and p-AKT
expression in RB tissues (n=50). Data are presented as the mean ±
standard deviation of three individual experiments. (D)
Immunohistochemistry was conducted to detect p-MEK1/2 in RB tissues
with high or low miR-153 expression. Bar graphs demonstrated a
significant inverse association between miR-153 and p-MEK1/2
expression in RB tissues (n=50). Data are presented as the mean ±
standard deviation of three individual experiments. RB,
retinoblastoma; IGF1R, insulin-like growth factor-1 receptor; MEK,
mitogen activated protein kinase kinase; miR, microRNA; p,
phosphorylated. |
Discussion
In the present study, it was found that miR-153 was
significantly downregulated in RB tissues and cell lines, and a low
miR-153 expression was closely associated with a larger tumor base
and differentiation. The overexpression of miR-153 inhibited RB
cell proliferation and promoted apoptosis, and suppressed the
migration and invasion of the WERI-RB-1 and Y79 cell lines. In
addition, IGF1R was identified as a direct target of miR-153 in RB
cells. Notably, the results of the present study demonstrate that
miR-153 may exert its antitumor effects on RB cells by blocking the
activation of the PI3K/AKT and MEK pathway. The findings presented
herein provide valuable clues towards the understanding of the
specific tumor suppressive function and regulatory mechanisms of
miR-153 in RB.
Accumulating evidence has indicated that miRNAs,
such as miR-124 (29) and
miR-25-3p (9), play important
roles in the development of human RB. However, the functional
significance of the unique role of miRNAs has yet to be elucidated
in RB. In the present study, miRNA microarray profiling was
performed using a GSE dataset (GSE7072), which was searched and
downloaded from the GEO database (www.ncbi.nlm.nih.gov/geo). miR-153 was found to be one
of the most significantly downregulated miRNAs in RB tissues.
Furthermore, it was confirmed that miR-153 was frequently
downregulated in RB tissues and cell lines. In addition, the
association between miR-153 expression and the clinicopathological
parameters of patients with RB was investigated, and the data
indicated that miR-153 in RB was significantly associated with a
larger tumor base and differentiation, while no significant
association was observed between miR-153 expression levels and
patient age, sex, tumor enucleation location, T classification and
clinical stage. Moreover, a low tumorous miR-153 expression was
found to be associated with a poor overall survival of patients
with RB. Taken together, these data suggest that miR-153 may be an
effective biomarker for RB prognosis.
Several studies have focused on comprehensive
investigations into the biological functions of miR-153 in various
types of cancer. For example, Li et al found that miR-153
inhibited EMT by targeting metadherin in breast cancer (30). Zuo et al demonstrated that
miR-153 may play a suppressive role in breast tumor growth and
metastasis via direct targeting of RUNX2 (9). Another study demonstrated that the
upregulation of miR-153 inhibited the proliferation, invasion and
migration of triple-negative breast cancer (TNBC) cells (31). However, the specific function of
miR-153 in RB progression has not been fully elucidated. In the
present study, the results revealed that the overexpression of
miR-153 inhibited the proliferation and invasion, and promoted the
apoptosis of RB cells. The present study, for the first time, to
the best of our knowledge, confirmed that miR-153 functions as a
tumor suppressor in RB. These findings are consistent with those of
previous studies demonstrating the tumor suppressive role of
miR-153 in other types of tumors (32,33).
It has been well documented that IGF1R is involved
in the carcinogenesis of a number of human cancers by activating
the PI3K/AKT and MEK pathways (27,34). Recent studies have demonstrated
that it acts as an important target of several miRNAs in RB,
including miR-98 (35) and
miR-145 (36). Recently, using a
microarray assay, Song et al found that miR-153 was
significantly downregulated in venous smooth muscle cells (VSMCs)
under conditions of stretch stress, and an enhanced miR-153
expression reduced IGF-1R expression, contributing to VSMC
proliferation (37). However, the
association between miR-153 and IGF1R in RB has not yet been
clarified. In the present study, miR-153 exerted a potent
inhibitory effect on the luciferase activity of the pGL3-report
vector linked to IGF1R 3′UTR. In addition, it was found that the
levels of IGF1R were significantly upregulated in RB tissues and
cell lines, and its expression was inversely correlated with
miR-153 expression in RB tissues. Furthermore, the overexpression
of IGF1R reversed the suppressive effects induced by miR-153
upregulation in RB cells. Taken together, these data indicate that
miR-153 exerts its antitumor effects by targeting IGF1R.
As a vital oncogene in the development and
maintenance of cancers, IGF1R triggers numerous downstream
signaling cascades, including the PI3K/AKT and Raf1/MEK1/2/ERK1/2
signaling pathways, which are involved in the tumorigenesis,
apoptosis and metastasis of a variety of cancer types (38-40). In the present study, the results
revealed that the upregulation of miR-153 reduced the levels of key
PI3K/AKT and Raf1/MEK1/2/ERK1/2 pathway proteins by suppressing
IGF1R. The data suggested that miR-153 suppressed IGF1R to inhibit
PI3K/AKT and Raf1/MEK1/2/ERK1/2 pathway activity, thus suppressing
the malignancy of RB cells.
Retinoblastoma gene (RB1), located at chromosome
13q14, was the first described tumor suppressor gene and its
critical role in cancer has attracted increasing attention
(41). It has been reported that
the loss of the RB1 gene in RB confers limitless replicative
potential to retinoblasts and is a rate-limiting step for RB
tumorigenesis (42). When both
alleles of the RB1 gene in RB are lost, the function of RB protein
(pRB) is curtailed, which results in abnormal cell proliferation
and tumor formation (43,44). Previous studies have reported that
the loss of function of pRB is not only associated with RB, but
also with multiple other non-ocular malignancies, such as prostate
cancer and hepatocellular carcinoma (45,46). In view of the association between
RB1 genes and RB, the authors aim to further study whether miR-153
exert antitumor effects through the regulation of the RB1 gene in
RB in the future.
There are some limitations to the present study. For
example, IGF1R is not unique as a target gene of miR-153; there are
other genes as well. In the future, other target genes of miR-153
or other differentially expressed miRNAs found in the present study
also need to be carefully examined for their role in RB
development. In addition, the expression level of miR-153 was
detected in the RB cell lines, Y79 and WERI-RB-1, which have
somewhat different characteristics, with the Y79 cells exhibiting
inherent metastatic properties and the WERI-RB-1 cells exhibiting
non-metastatic properties (47).
The results of the present study revealed that the expression
levels of miR-153 in the Y79 cells were higher than those in the
Weri-RB1 cells (Fig. 1E),
suggesting that the miR-153 expression level may not be associated
with the aggressiveness of those cell lines. In the future, the
authors aim to examine the expression of miR-153 in more metastatic
and non-metastatic RB tissues samples in order to assess whether
miR-153 is directly associated with the aggressiveness of RB.
In conclusion, the present study demonstrated that
miR-153 inhibited the proliferation, migration and invasion, and
promoted the apoptosis of WERI-RB-1 and Y79-45 cell lines by
inhibiting the IGF1R/PI3K/AKT and IGF1R/Raf/MEK signaling pathways.
These findings suggest that miR-153 may serve as a potential
biomarker for the prognosis and a therapeutic target for patients
with RB.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LG was responsible for main conception of the study
and the draft of the manuscript. YB, TN, YL, RC, SJ and SL assisted
in the design of the study and performed the statistical analysis.
YB and TN assisted with the revision of the manuscript and
participated in its design. LG and YB confirm the authenticity of
all the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
All individuals provided written informed consent
for the use of human specimens for clinical research. The present
study was approved by The First People's Hospital of Shangqiu
Ethics Committee.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
No funding was received.
References
|
1
|
Dimaras H, Dimba EA and Gallie BL:
Challenging the global retinoblastoma survival disparity through a
collaborative research effort. Br J Ophthalmol. 94:1415–1416. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Narang S, Mashayekhi A, Rudich D and
Shields CL: Predictors of long-term visual outcome after
chemoreduction for management of intraocular retinoblastoma. Clin
Exp Ophthalmol. 40:736–742. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shields CL and Shields JA: Basic
understanding of current classification and management of
retinoblastoma. Curr Opin Ophthalmol. 17:228–234. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen HM, Ong SJ, Chao AN, Liou KL, Jung SM
and Kao LY: Histopathologic findings after selective ophthalmic
arterial injection of melphalan for retinoblastoma. Taiwan J
Ophthalmol. 9:262–266. 2019. View Article : Google Scholar
|
|
5
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shukla GC, Singh J and Barik S: MicroRNAs:
Processing, maturation, target recognition and regulatory
functions. Mol Cell Pharmacol. 3:83–92. 2011.PubMed/NCBI
|
|
8
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zuo Z, Ye F, Liu Z, Huang J and Gong Y:
MicroRNA-153 inhibits cell proliferation, migration, invasion and
epithelial-mesenchymal transition in breast cancer via direct
targeting of RUNX2. Exp Ther Med. 17:4693–4702. 2019.PubMed/NCBI
|
|
10
|
Zhang Z, Sun J, Bai Z, Li H, He S, Chen R
and Che X: MicroRNA-153 acts as a prognostic marker in gastric
cancer and its role in cell migration and invasion. Onco Targets
Ther. 8:357–364. 2015.PubMed/NCBI
|
|
11
|
Niu G, Li B, Sun L and An C: MicroRNA-153
inhibits osteosarcoma cells proliferation and invasion by targeting
TGF-β2. PLoS One. 10:e01192252015. View Article : Google Scholar
|
|
12
|
Chen WJ, Zhang EN, Zhong ZK, Jiang MZ,
Yang XF, Zhou DM and Wang XW: MicroRNA-153 expression and prognosis
in non-small cell lung cancer. Int J Clin Exp Pathol. 8:8671–8675.
2015.PubMed/NCBI
|
|
13
|
Jiang J, Liu Y, Zhao Y, Tian F and Wang G:
miR-153-3p suppresses inhibitor of growth protein 2 expression to
function as tumor suppressor in acute lymphoblastic leukemia.
Technol Cancer Res Treat. 18:15330338198529902019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li C, Zhang Y, Zhao W, Cui S and Song Y:
miR-153-3p regulates progression of ovarian carcinoma in vitro and
in vivo by targeting MCL1 gene. J Cell Biochem. 120:19147–19158.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang H, Ge F, Xu Y, Xiao J, Zhou Z, Liu R
and Chen C: miR-153 inhibits the migration and the tube formation
of endothelial cells by blocking the paracrine of angiopoietin 1 in
breast cancer cells. Angiogenesis. 21:849–860. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
17
|
Villar J, Cabrera-Benitez NE, Valladares
F, García-Hernández S, Ramos-Nuez Á, Martín-Barrasa JL, Muros M,
Kacmarek RM and Slutsky AS: Tryptase is involved in the development
of early ventilator-induced pulmonary fibrosis in sepsis-induced
lung injury. Crit Care. 19:1382015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wan W, Wan W, Long Y, Li Q, Jin X, Wan G,
Zhang F, Lv Y, Zheng G, Li Z and Zhu Y: MiR-25-3p promotes
malignant phenotypes of retinoblastoma by regulating PTEN/Akt
pathway. Biomed Pharmacother. 118:1091112019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu S, Han M and Zhang C: Overexpression of
microRNA-186 inhibits angiogenesis in retinoblastoma via the
hedgehog signaling pathway by targeting ATAD2. J Cell Physiol.
234:19059–19072. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liao Y, Yin X, Deng Y and Peng X:
MiR-140-5p suppresses retinoblastoma cell growth via inhibiting
c-Met/AKT/mTOR pathway. Biosci Rep. 38:BSR201807762018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shan N, Shen L, Wang J, He D and Duan C:
MiR-153 inhibits migration and invasion of human non-small-cell
lung cancer by targeting ADAM19. Biochem Biophys Res Commun.
456:385–391. 2015. View Article : Google Scholar
|
|
22
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guarino M: Epithelial-mesenchymal
transition and tumour invasion. Int J Biochem Cell Biol.
39:2153–2160. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hakuno F and Takahashi SI: IGF1 receptor
signaling pathways. J Mol Endocrinol. 61:T69–T86. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu S, Soutto M, Chen Z, Blanca Piazuelo
M, Kay Washington M, Belkhiri A, Zaika A, Peng D and El-Rifai W:
Activation of IGF1R by DARPP-32 promotes STAT3 signaling in gastric
cancer cells. Oncogene. 38:5805–5816. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Z, Pan W, Shen Y, Chen Z, Zhang L,
Zhang Y, Luo Q and Ying X: IGF1/IGF1R and microRNA let-7e
down-regulate each other and modulate proliferation and migration
of colorectal cancer cells. Cell Cycle. 17:1212–1219. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kooijman R, Lauf JJ, Kappers AC and
Rijkers GT: Insulin-like growth factor induces phosphorylation of
immunoreactive insulin receptor substrate and its association with
phosphatidylinositol-3 kinase in human thymocytes. J Exp Med.
182:593–597. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fu HW, Lin X, Zhu YX, Lan X, Kuang Y, Wang
YZ, Ke ZG, Yuan T and Chen P: Circ-IGF1R has pro-proliferative and
anti-apoptotic effects in HCC by activating the PI3K/AKT pathway.
Gene. 716:1440312019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang J, Yang Y, Fang F and Liu K: MALAT1
modulates the autophagy of retinoblastoma cell through
miR-124-mediated stx17 regulation. J Cell Biochem. 119:3853–3863.
2018. View Article : Google Scholar
|
|
30
|
Li W, Zhai L, Zhao C and Lv S: MiR-153
inhibits epithelial-mesenchymal transition by targeting metadherin
in human breast cancer. Breast Cancer Res Treat. 150:501–509. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shi D, Li Y, Fan L, Zhao Q, Tan B and Cui
G: Upregulation of miR-153 inhibits triple-negative breast cancer
progression by targeting ZEB2-mediated EMT and contributes to
better prognosis. Onco Targets Ther. 12:9611–9625. 2019. View Article : Google Scholar
|
|
32
|
Zuo J, Zhao M, Fan Z, Liu B, Wang Y, Li Y,
Lv P, Xing L, Zhang X and Shen H: MicroRNA-153-3p regulates cell
proliferation and cisplatin resistance via Nrf-2 in esophageal
squamous cell carcinoma. Thorac Cancer. 11:738–747. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zeng HF, Yan S and Wu SF: MicroRNA-153-3p
suppress cell proliferation and invasion by targeting SNAI1 in
melanoma. Biochem Biophys Res Commun. 487:140–145. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Scheidegger KJ, Du J and Delafontaine P:
Distinct and common pathways in the regulation of insulin-like
growth factor-1 receptor gene expression by angiotensin II and
basic fibroblast growth factor. J Biol Chem. 274:3522–3530. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Guo L, Bai Y, Ji S and Ma H: MicroRNA-98
suppresses cell growth and invasion of retinoblastoma via targeting
the IGF1R/k-Ras/Raf/MEK/ERK signaling pathway. Int J Oncol.
54:807–820. 2019.PubMed/NCBI
|
|
36
|
Chen Z, Yang H, Nie Y and Xing Y: miR-145
regulates the proliferation and apoptosis of Y79 human
retinoblastoma cells by targeting IGF-1R. Int J Clin Exp Pathol.
11:4331–4338. 2018.PubMed/NCBI
|
|
37
|
Song L, Duan P, Guo P, Li D, Li S, Xu Y
and Zhou Q: Downregulation of miR-223 and miR-153 mediates
mechanical stretch-stimulated proliferation of venous smooth muscle
cells via activation of the insulin-like growth factor-1 receptor.
Arch Biochem Biophys. 528:204–211. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Knowlden JM, Hutcheson IR, Barrow D, Gee
JM and Nicholson RI: Insulin-like growth factor-I receptor
signaling in tamoxifen-resistant breast cancer: A supporting role
to the epidermal growth factor receptor. Endocrinology.
146:4609–4618. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rozen F and Pollak M: Inhibition of
insulin-like growth factor I receptor signaling by the vitamin D
analogue EB1089 in MCF-7 breast cancer cells: A role for
insulin-like growth factor binding proteins. Int J Oncol.
15:589–594. 1999.PubMed/NCBI
|
|
40
|
LeRoith D, Werner H, Neuenschwander S,
Kalebic T and Helman LJ: The role of the insulin-like growth
factor-I receptor in cancer. Ann N Y Acad Sci. 766:402–408. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhu L: Tumour suppressor retinoblastoma
protein Rb: A transcriptional regulator. Eur J Cancer.
41:2415–2427. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mu G, Liu H, Zhou F, Xu X, Jiang H, Wang Y
and Qu Y: Correlation of overexpression of HMGA1 and HMGA2 with
poor tumor differentiation, invasion, and proliferation associated
with let-7 down-regulation in retinoblastomas. Hum Pathol.
41:493–502. 2010. View Article : Google Scholar
|
|
43
|
Dalgard CL, Gonzalez M, de Niro JE and
O'Brien JM: Differential microRNA-34a expression and tumor
suppressor function in retinoblastoma cells. Invest Ophthalmol Vis
Sci. 50:4542–4551. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ganguly A and Shields CL: Differential
gene expression profile of retinoblastoma compared to normal
retina. Mol Vis. 16:1292–1303. 2010.PubMed/NCBI
|
|
45
|
Maddison LA, Sutherland BW, Barrios RJ and
Greenberg NM: Conditional deletion of Rb causes early stage
prostate cancer. Cancer Res. 64:6018–6025. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hui AM, Li X, Makuuchi M, Takayama T and
Kubota K: Over-expression and lack of retinoblastoma protein are
associated with tumor progression and metastasis in hepatocellular
carcinoma. Int J Cancer. 84:604–608. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Guihurt Santiago J, Burgos-Tirado N,
Lafontaine DD, Mendoza Sierra JC, Camacho RH, Vecchini Rodríguez
CM, Morales-Tirado V and Flores-Otero J: Adhesion G protein-coupled
receptor, ELTD1, is a potential therapeutic target for
retinoblastoma migration and invasion. BMC Cancer. 21:532021.
View Article : Google Scholar : PubMed/NCBI
|