1. Introduction
Gliomas are the most common malignancy of the
central nervous system, accounting for 80% of all primary brain
tumours. The recurrence and mortality rates of gliomas are high
(1). Glioblastoma multiforme
(GBM) is a grade 4 highly aggressive tumour. Although a variety of
high-intensity treatment regimens (such as a combination of surgery
and chemoradiotherapy) have been adopted, the median survival time
for patients with GBM remains at only 12-15 months and the 5-year
survival rate is only 3-5% (1). A
randomized phase III clinical study of temozolomide (TMZ) indicated
that the median survival time for patients with GBM treated with a
combination of TMZ and concurrent radiotherapy was 14.6 months and
that the median survival time for patients treated with
radiotherapy alone was 12 months (2). Although TMZ is currently the
standard first-line chemotherapeutic drug for malignant gliomas,
TMZ resistance has become a substantial obstacle to the
chemotherapeutic treatment of gliomas and frequently leads to
treatment failure and poor prognosis. Therefore, the regulatory
processes leading to the development of drug resistance in gliomas
have attracted extensive attention and become an emerging research
hotspot. However, the specific molecular mechanisms by which
noncoding (nc)RNAs act as key molecules to regulate the drug
resistance of gliomas remain to be fully elucidated and remain a
key scientific task that urgently requires to be resolved to
improve the clinical treatment of gliomas.
In recent years, the development of next-generation
sequencing technology and bioinformatics analysis has revealed
information that was previously inaccessible. The human genome
contains an estimated 20,000-25,000 protein-coding genes,
accounting for <2% of the entire genome. The remaining 98% of
the genome encodes functional RNA molecules that cannot be
translated into proteins (3).
Most of the ncRNAs involved in glioma drug resistance are microRNAs
(miRNAs), long noncoding RNAs (lncRNAs) and circular RNAs
(circRNAs), and they are being gradually uncovered. An increasing
amount of evidence indicates that ncRNAs are not only biomarkers of
tumour proliferation, invasion, apoptosis, autophagy and immune
response, but also regulatory factors involved in glioma drug
resistance. ncRNAs may act as oncogenes or tumour suppressor genes
to mediate chemoresistance during the treatment of tumours
(3,4). Therefore, ncRNAs have become key
molecules in glioma drug resistance and the targeted regulation of
ncRNAs may become a novel therapeutic strategy to reverse drug
resistance in gliomas. In the present review, the relevant
mechanisms of drug resistance in gliomas were discussed and the
regulatory functions and mechanisms of ncRNAs in glioma drug
resistance were systematically summarized. A series of hot issues
and research trends of tumour drug resistance, ncRNA and vector
modification are then briefly summarized, aiming to further analyze
the potential significance of ncRNAs as novel therapeutic targets
for the reversal of glioma drug resistance, and providing a new
direction for basic research on tumor precision treatment.
2. Mechanisms of chemotherapy resistance in
gliomas
In the past, chemotherapies for gliomas
overemphasized the blood-brain barrier (BBB), which was thought to
limit the access of anticancer drugs to tumour tissue, resulting in
chemotherapy failure (1). In
fact, scholars have demonstrated that the BBB in patients with
malignant gliomas is frequently damaged to varying degrees. The
tolerance of gliomas themselves to anticancer drugs is the most
essential cause of chemotherapy failure (5). Therefore, revealing the mechanisms
of resistance is of crucial importance for reducing or eliminating
the phenomenon of chemoresistance. In the following chapter,
several of the most recognized mechanisms of drug resistance in
gliomas were summarized (Fig.
1).
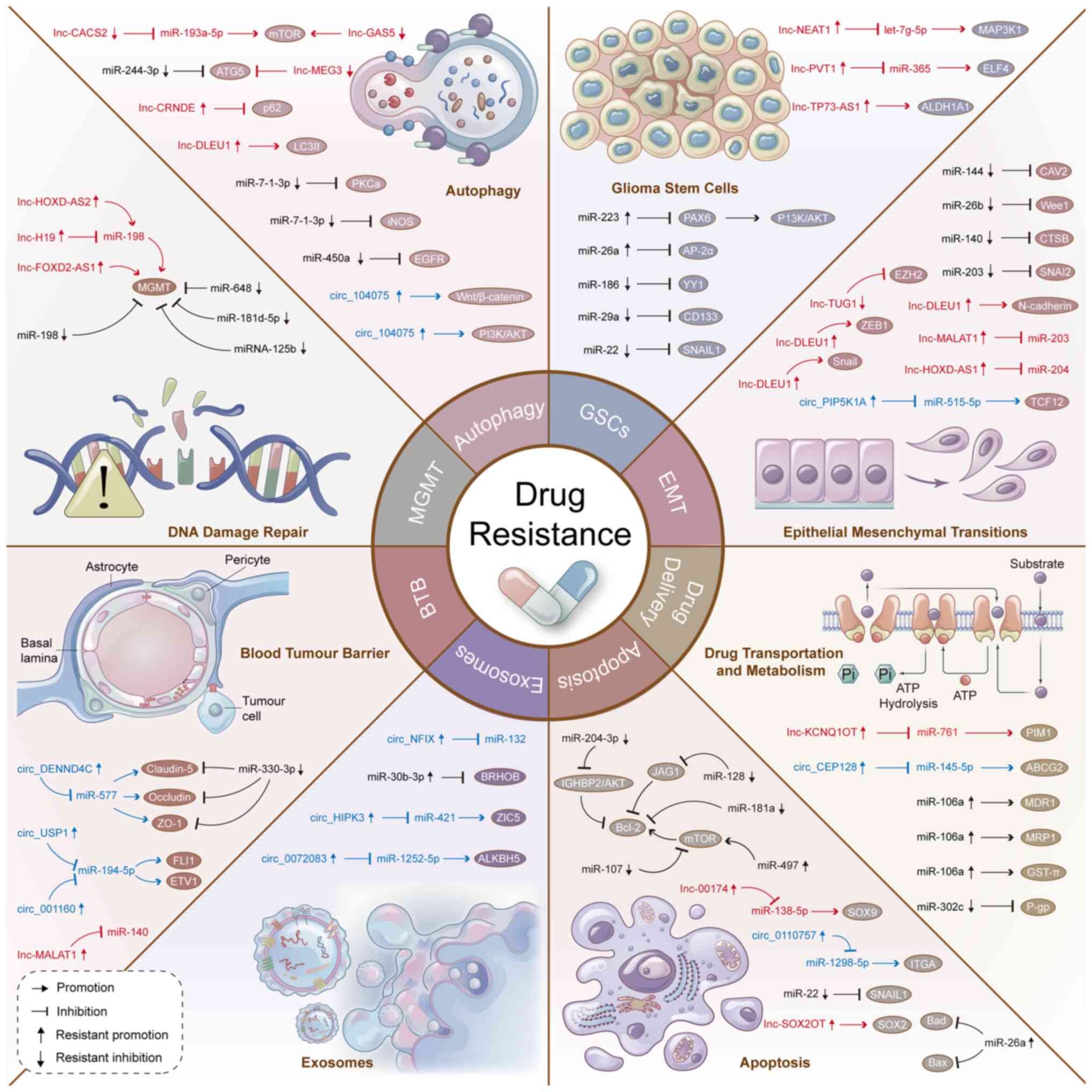 | Figure 1Novel ncRNAs and their downstream
targets related to drug resistance mechanisms in glioma. Different
ncRNAs (miRNAs, lncRNAs and circRNAs) and their downstream targets
promote (or inhibit) chemotherapy resistance of glioma cells by
regulating GSCs phenotype, EMT, drug transport, apoptosis,
exosomes, blood tumour barrier, DNA repair and autophagy. EMT,
epithelial-mesenchymal transition; GSC, glioma stem cell; lncRNA,
long non-coding RNA; miR, microRNA; circRNA, circular RNA; MGMT,
O6-methylguanine-DNA methyltransferase. |
Drug transport and metabolism
After reaching their therapeutic targets, numerous
antitumour drugs are actively pumped out of tumour cells against
the concentration gradient by transporter proteins, reducing the
intracellular accumulation of drugs and causing drug resistance.
This is a classic feature of pharmacokinetics. The 3 main proteins
that cause drug resistance are P-glycoprotein (P-gp), multidrug
resistance-associated protein (MRP) and breast cancer resistance
protein (BCRP) (6). P-gp is an
ATP-dependent drug efflux pump that binds to antitumour drugs and
ATP simultaneously. It utilizes the energy from ATP to transport
chemotherapeutic drugs out of glioma cells, resulting in drug
resistance (7). In addition, P-gp
is also expressed in vascular endothelial cells and participates in
drug resistance caused by the blood-tumour barrier (BTB). MRP and
P-gp have the same function. These proteins specifically recognize
hydrophobic chemotherapy drugs. They bind to glutathione to form a
glutathione S-conjugate pump and indirectly transport weakly basic
chemotherapy drugs (7,8). In a study by Marinho et al
(9), MRP1 and MRP3 were observed
to be highly expressed in glioma cells and related to resistance to
chemotherapy drugs, e.g. etoposide and vincristine. BCRP was
initially isolated from multidrug-resistant breast cancer cells.
BCRP is also highly expressed in glioma cells. It affects the
efficacy of >20 anticancer drugs (including daunorubicin and
mitoxantrone), resulting in drug resistance (10). In addition to the 3 types of
transporters described above, major vault protein (MVP) and lung
resistance-related protein (LRP) also induce drug resistance by
interfering with drug transport (6). Studies have demonstrated that both
MVP and LRP are highly expressed in drug-resistant glioma cells,
particularly in tumour blood vessels.
Inhibition of apoptosis
When drug resistance occurs in tumours, the
apoptotic/anti-apoptotic mechanism is in an unbalanced state. P53
is a classic tumour suppressor gene. Wild-type p53 directly induces
the apoptosis of tumour cells. The mechanism by which glioma cells
develop resistance in the process of TMZ chemotherapy frequently
involves the deletion or mutation of wild-type p53 and the
significantly increased expression of O6-methylguanine-DNA
methyltransferase (MGMT). As a result, apoptosis cannot be induced
in glioma cells (2,11). The B-cell lymphoma-2 (Bcl-2) gene
is a well-known anti-apoptotic gene and causes drug resistance in
tumour cells by inhibiting apoptosis. Therefore, Bcl-2 is also
recognized as a novel drug resistance gene. In addition, the
apoptosis system includes a key factor, the homeobox (HOX) gene.
HOX is a transcription factor. This gene is likely to be highly
expressed during chemotherapy and directly induces TMZ resistance
through the PI3K/AKT signalling pathway. Furthermore, HOX increases
the expression of MGMT through the nuclear factor κB (NF-κB)
signalling pathway, which indirectly induces the development of
drug resistance (12).
DNA damage repair
The destruction of the DNA structure of tumour cells
to promote apoptosis is one of the most common mechanisms of action
of drugs. Conversely, this strategy leads to the failure of
chemotherapeutic drugs if glioma cells acquire an enhanced ability
to repair damaged DNA during chemotherapy. MGMT is one of the
predictive indicators of the prognosis and chemosensitivity of
gliomas. The main function of MGMT is to repair intracellular DNA
damage caused by alkylating agents. Therefore, MGMT induces
resistance to antitumour drugs by preventing the formation of DNA
crosslinking (13). Mismatch
repair (MMR) is mainly involved in the repair of DNA replication
errors. It prevents gene mutations and thus inhibits the
occurrence, development and drug resistance of tumours. In
addition, a complex regulatory network exists between MMR and MGMT.
Studies have indicated that there is a negative correlation between
them in glioma cells (11,14).
Topoisomerase II (Top II) has also been an important focus of
studies on chemoresistance in recent years. Top II maintains DNA
stability and genome integrity in glioma cells by reducing drug
efficacy (15). In addition,
another study reported that the sensitivity of glioma cells to TMZ
increases when the expression of poly(ADP-ribose) polymerase-1,
base excision repair enzymes or high mobility group AT-hook 2
decreases (16).
Autophagy
Autophagy may exert opposite regulatory effects on
tumours. It has either a promoting or an inhibitory effect and the
final result depends on the type and state of the tumours. When
tumour cells are stimulated by chemotherapeutic drugs, autophagy is
activated as a stress response. Autophagy may provide sufficient
energy for tumour cell metabolism through degrading proteins,
thereby promoting tumour development and reducing the efficacy of
chemotherapeutic drugs. Studies have indicated that a number of
signalling pathways are involved in TMZ-induced autophagy in glioma
cells, including the PI3K/Akt/mammalian target of rapamycin (mTOR),
hypoxia-inducible factor 1α (HIF-1α)/C-X-C chemokine receptor type
4 and Ras/Raf/MAPK kinase pathways (17). Therefore, inhibition of tumour
drug resistance by regulating autophagy-related target genes may
become a novel strategy for the treatment of drug resistance in
gliomas.
Glioma stem cells (GSCs)
GSCs are present in glioma tissue. These cells have
strong proliferation and differentiation abilities, and
conventional radiotherapy and chemotherapy are virtually
ineffective against them. Therefore, GSCs are considered to be the
culprit of glioma recurrence. In addition, the powerful resistance
mechanisms and long survival time of GSCs allow tumour drug
resistance to increase with the intracellular accumulation of
related mutations. Relevant mechanistic studies also indicate that
glioma chemoresistance is closely related to the high expression of
drug resistance genes in GSCs, including the multidrug resistance
(MDR), MGMT and ATP-binding cassette (ABC) transporter genes
(18).
Epithelial-mesenchymal transition
(EMT)
An increasing amount of evidence indicates that
glioma cells may be converted to cells with stem-like properties
through EMT. Furthermore, GSCs may overexpress EMT marker molecules
and undergo the EMT process. These events eventually lead to the
doubling of the number of GSCs, resulting in the aggravation of
glioma biological behaviours such as invasion and drug resistance
(19). EMT, GSCs and
chemoresistance are regarded by certain scholars as key factors for
malignant progression of gliom and are assumed to be the
fundamental reason these tumours are difficult to cure. Therefore,
targeted intervention or the reversal of EMT may serve as a novel
research approach for the treatment of glioma chemoresistance.
Mutations at drug target sites
Abnormal expression caused by target gene mutations
is one of the common mechanisms of chemoresistance in tumours. In
gliomas, Top II is an important target of chemotherapeutic drugs.
Its enzymatic activity mediates the unwinding of DNA double strands
for replication. Certain studies have indicated that Top II in
patients with recurrent gliomas is prone to mutation, which
frequently leads to the loss of drug targeting and the development
of drug resistance (15,20). In addition, a study by Santangelo
et al (21) demonstrated
that metallothionein is highly expressed in gliomas, another
important cause of chemoresistance and poor prognosis.
Tumour microenvironment
The glioma microenvironment is composed of not only
tumour cells, fibroblasts, immune cells and inflammatory cells, but
also the surrounding intercellular matrix, microvessels and
infiltrating biological molecules. Hypoxia caused by the high
proliferation of tumour cells represents a common glioma
microenvironment. Hypoxia stimulates the high expression of P-gp
and MDR in glioma cells, thereby enhancing the vitality and drug
resistance of tumour cells (22).
Similarly, cytokines secreted by stromal cells, such as
transforming growth factor (TGF) and vascular endothelial growth
factor (VEGF), also inhibit the cytotoxicity of chemotherapeutic
drugs (23).
3. miRNAs and glioma drug resistance
As the most extensively studied regulatory factors
in ncRNA research, miRNAs have been indicated to target and bind to
the 3′-untranslated region (3′-UTR) of mRNAs, resulting in the
direct degradation of mRNAs or the blocking of translation. Studies
have indicated that through such regulatory mechanisms, miRNAs
directly or indirectly participate in tumour proliferation,
metastasis, drug resistance and immunity. A number of abnormally
expressed miRNAs have been detected in various drug-resistant
glioma cells, indicating that miRNAs have a key role in glioma
chemoresistance. The following is a summary of the research on the
involvement of miRNAs in the resistance of gliomas to TMZ,
cisplatin (DDP), nitrosourea and plant-derived anticancer drugs
(Fig. 2).
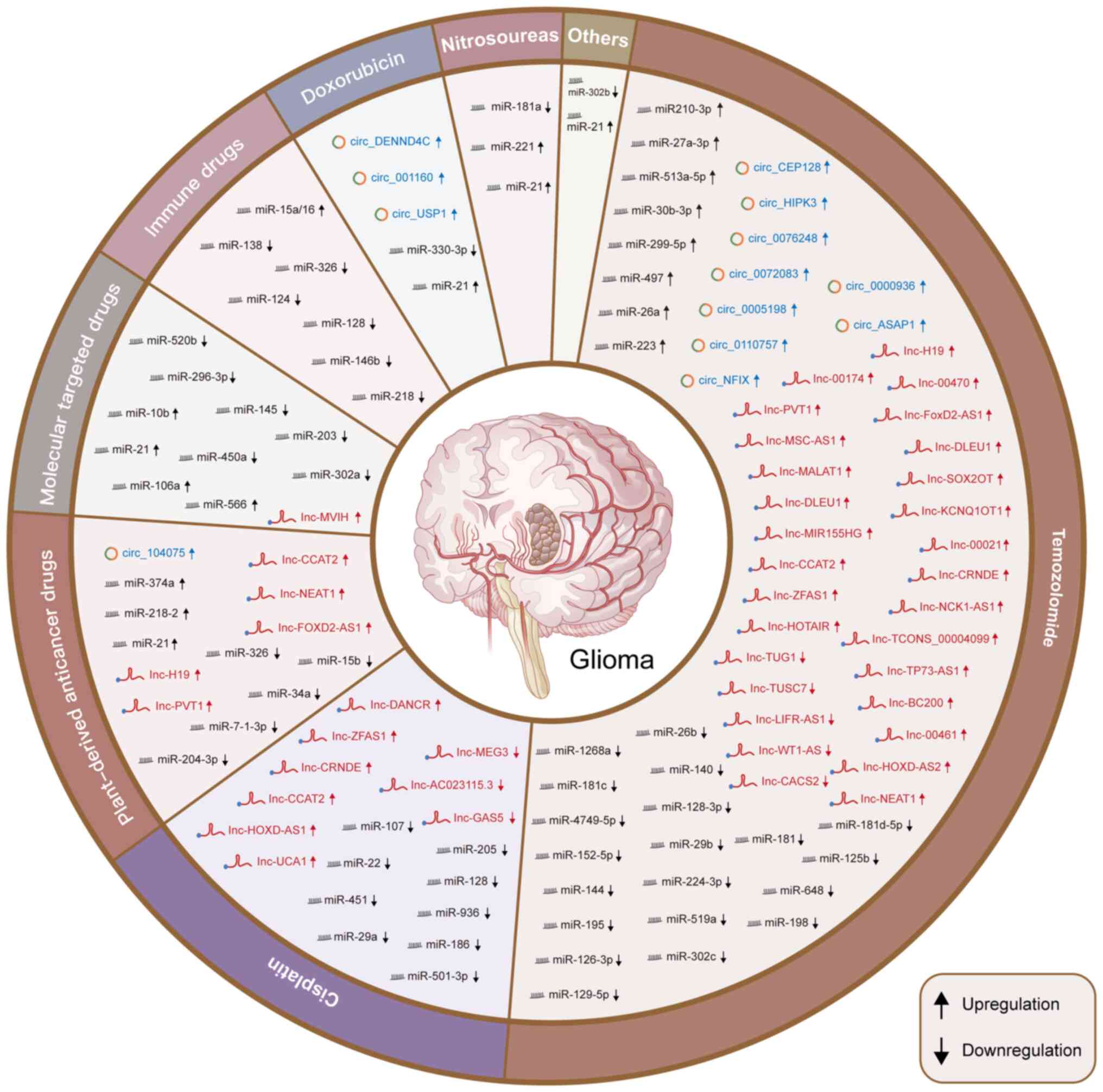 | Figure 2Novel ncRNAs related to antitumor
drugs (including the proportion of genes involved in various drugs;
temozolomide has a dominant role). Certain ncRNAs have differential
expression and regulatory functions in the formation of drug
resistance to various anti-tumor drugs (temozolomide, cisplatin,
plant-derived anticancer drugs, molecular targeted drugs,
immunotherapeutic drugs, doxorubicin and nitrosourea drugs), which
may be used as a potential target for drug resistance treatment of
glioma. NcRNA, noncoding RNA; lncRNA, long non-coding RNA; miR,
microRNA; circRNA, circular RNA. |
miRNAs are involved in TMZ
resistance
The main mechanisms underlying the cytotoxicity of
TMZ are the destruction of the DNA structure in glioma cells,
prevention of DNA repair and induction of apoptosis. As a
second-generation alkylating agent, this drug has good
bioavailability and central nervous system permeability. At
present, TMZ-based chemotherapy regimens are widely used in the
postoperative adjuvant treatment of high-grade gliomas and the
salvage treatment of recurrent gliomas. Although the clinical
application of TMZ has a 'milestone' significance, resistance to
TMZ has become an important reason limiting the efficacy of
chemotherapy. TZM resistance is a key scientific problem that
requires to be urgently resolved in the clinical chemotherapy
treatment of gliomas. To date, numerous studies have demonstrated a
relationship between miRNAs and TMZ resistance in gliomas (Table I).
 | Table ImiRNAs involved in temozolomide
resistance. |
Table I
miRNAs involved in temozolomide
resistance.
| miRNA | Direction of
differential expression | Genes and
pathways | Mechanism | (Refs.) |
|---|
| miR-210-3p | ↑ | NeuroD2, HIF3A | - | (24) |
| miR-27a-3p | ↑ | NF1 | - | (25) |
| miR-513a-3p | ↑ | NEDD4L, IGF-1 | - | (26) |
| miR-26a | ↑ | Bad, Bax,
AP-2α | Apoptosis,
GSCs | (27,31) |
| miR-299-5p | ↑ | GOLPH3,
MAPK/ERK | Apoptosis | (28) |
| miR-497 | ↑ | IGFIR/IRS1,
mTOR/Bcl-2 | Apoptosis | (29) |
| miR-30b-3p | ↑ | RHOB | GSCs | (30) |
| miR-223 | ↑ | PAX6/PI3K/AKT | GSCs | (32) |
| miR-181c | ↓ | RPN2, Tcf-4 | - | (33) |
| miR-4749-5p | ↓ | RFC2 | - | (34) |
| miR-152-5p | ↓ | FBXL7 | - | (35) |
| miR-144 | ↓ | FGF7, CAV2 | - | (36) |
| miR-195 | ↓ | CCNE1 | Apoptosis | (37) |
| miR-126-3p | ↓ |
SOX2/Wnt/β-catenin | Apoptosis | (38) |
| miR-181 | ↓ | SELK | Apoptosis | (39) |
| miR-648,
miRNA-125b | ↓ | MGMT | DNA damage
repair | (40) |
| miR-181d-5p | ↓ | MGMT | DNA damage
repair | (41) |
| miR-198 | ↓ | MGMT | DNA damage
repair | (42) |
| miR-26b | ↓ | Wee1 | EMT | (43) |
| miR-140 | ↓ | CTSB | EMT | (44) |
| miR-128-3p | ↓ | C-met, PDGFRα,
Notch1, Slug | EMT | (45) |
| miR-29b | ↓ | - | Autophagy | (46) |
| miR-224-3p | ↓ | ATG5 | Autophagy | (47) |
| miR-519a | ↓ |
STAT3/Bcl-2/Beclin-1 | Autophagy | (48) |
| miR-302c | ↓ | P-gp | Drug transport and
metabolism | (49) |
| miR-1268a | ↓ | ABCC1 | Drug transport and
metabolism | (50) |
| miR-129-5p | ↓ | Wnt5a | Drug transport and
metabolism | (51) |
Oncogenic miRNAs promote the resistance of gliomas
to TMZ. Studies have indicated that in the hypoxic
microenvironment, the HIF1α/HIF2α-miR210-3p network forms a
positive feedback loop with epidermal growth factor (EGF) and
jointly promotes the malignant progression and TMZ resistance of
GBM. Jana et al (24)
further explored a novel strategy that used tachyplesin (Tpl) as a
nanocarrier for anti-miR-210 in the treatment of GBM. The results
indicated that this Tpl complex significantly inhibited the
expression of miR-210 and induced the expression of tumour
suppressor genes neuronal differentiation 2 and HIF3A in GBM cells
in a targeted manner, thereby enhancing the TMZ sensitivity of GBM
(24). miR-27a-3p is highly
expressed in glioma tissues and various glioma cell lines. In
addition, miR-27a-3p induces chemotherapy resistance of glioma
cells to TMZ by degrading the downstream target neurofibromatosis
type 1 (NF1). Of note, restoration of the expression of NF1
reverses this drug resistance effect (25). NEDD4 like E3 ubiquitin protein
ligase (NEDD4L) has been demonstrated to be a direct target gene
downstream of carcinogenic miR-513a-5p. The inhibitory effect of
miR-513a-5p on NEDD4L is directly involved in the activation of the
WNT/β-catenin signalling pathway by insulin-like growth factor 1
(IGF-1) and significantly reduces the cytotoxicity of TMZ to
gliomas (26).
Studies have indicated that certain miRNAs regulate
apoptosis-related signalling pathways and induce tumour cell escape
from apoptosis, resulting in the resistance of gliomas to TMZ. For
instance, HIF-1α-mediated miR-26a is a hypoxia-sensitive miRNA. The
expression of miR-26a is significantly upregulated in GBM cells
under hypoxic conditions. The upregulation of miR-26a directly
induces the protective response of mitochondria and enhances TMZ
resistance in vivo and in vitro. In addition, miR-26a
inhibits the expression of Bcl-2-associated X protein (Bax) and
Bcl-2-associated agonist of cell death, thereby reducing
TMZ-induced apoptosis (27).
Similarly, miR-299-5p, which is significantly expressed in glioma
tissues and cell lines, regulates the MAPK/ERK signaling axis by
targeting Golgi phosphoprotein 3, thereby inhibiting apoptosis and
enhancing the chemoresistance of gliomas to TMZ (28). In addition, miR-497 upregulates
the protein expression of mTOR and Bcl-2 through the IGF1
receptor/insulin receptor substrate 1 pathway, thereby inducing TMZ
apoptosis resistance in glioma cells (29). GSCs have an important role in
glioma chemotherapy through a variety of complex interlaced
signalling networks. An increasing amount of evidence indicates
that there is an association between miRNAs and GSC-driven drug
resistance in gliomas. The expression of miR-30b-3p is
significantly increased in hypoxic GSC-derived exosomes. miR-30b-3p
may be transported in exosomes into recipient GBM cells, where it
directly targets and binds to Ras homolog family member B to
promote TMZ chemoresistance (30). In addition, high expression of
exogenous miR-26a and miR-223 promotes the proliferative ability
and sphere formation of GSCs, thereby inducing chemoresistance in
glioma cells. Evidence indicates that miR-26a induces the
proliferation of GSCs by targeting and binding to the 3′-UTR of
activator protein 2α (AP-2α), thereby promoting the resistance of
gliomas to TMZ (31). The
expression of miR-223 is significantly increased in GBM cells.
miR-223 directly binds to and negatively regulates paired box 6 and
promotes the proliferation and differentiation of GSCs by
regulating the PI3K/Akt signalling pathway, thereby reducing TMZ
sensitivity and the growth inhibitory effect of TMZ (32).
By contrast, miRNAs may also act as tumour
suppressors to reverse TMZ resistance in gliomas. In general, the
expression of these miRNAs is downregulated in gliomas. The
restoration of their expression is conducive to the enhancement of
the sensitivity of tumour cells to chemotherapeutic drugs. For
instance, overexpression of miR-181c inhibits the activity of
β-catenin/transcription factor 4 (TCF-4) by targeting and binding
to ribophorin II, thereby reducing the resistance of GBM cells to
TMZ (33). Upregulation of
miR-4749-5p causes the degradation of replication factor C subunit
2 (RFC2) and hinders RFC2 from performing DNA repair, thereby
promoting TMZ-mediated cytotoxicity (34). In U251 glioma cells, knockdown of
miR-152-5p expression inhibited the chemoresistance of cells to TMZ
through a targeted reduction in F-box/LRR-repeat protein 7
(35). In addition, in
vitro molecular experiments have demonstrated that fibroblast
growth factor 7 (FGF7) and caveolin 2 (CAV2) are target genes of
miR-144. Knockdown of FGF7 induced apoptosis of U251 cells by
activating the Akt/reactive oxygen species (ROS) pathway and
knockdown of CAV2 inhibited the EMT of U251 cells. These events
jointly increase the sensitivity of U251 cells to TMZ (36). In addition, certain tumour
suppressor miRNAs enhance the sensitivity of glioma cells to TMZ by
inducing apoptosis. The expression of miR-195 is significantly
decreased in TMZ-resistant glioma cells and is negatively
associated with the drug resistance index. miR-195 reverses the TMZ
resistance of gliomas by targeting and binding to cyclin E1 and
inducing apoptosis (37).
Similarly, decreased expression of miR-126-3p has been detected in
TMZ-resistant glioma tissues and cell lines. Furthermore miR-126-3p
directly inhibits the expression of SRY-box 2 (SOX2) to induce
Wnt/β-catenin inactivation, thereby promoting the apoptosis of
glioma cells and enhancing the cytotoxicity of TMZ (38). In addition, the human miR-181
family has 8 members that have an important role in the occurrence
and development of a variety of malignant tumours. Overexpression
of miR-181 increases the Bax/Bcl-2 ratio and inhibits the
proliferation of glioma cells by targeting and binding to the
3′-UTR of selenoprotein K, thereby enhancing the sensitivity of
tumours to TMZ (39).
MGMT is a DNA repair enzyme that has been indicated
to cause chemoresistance in tumours. The basic function of MGMT is
to repair damaged guanine nucleotides by transferring the methyl
group at the O6 site of guanine to a cysteine residue, thereby
preventing gene mutation and cell death caused by alkylating
agents. Through an examination of patients with MGMT methylation,
Jesionek-Kupnicka et al (40) observed that the expression of MGMT
was downregulated and negatively correlated with the expression of
miR-648 and miR-125b. Another study reported that after delivering
miRNA mimics into glioma cells, highly expressed miRNA-181d
reverses the resistance of glioma cells to TMZ by targeting and
binding to MGMT. Moreover, a genome-wide microarray analysis noted
that miR-181d-5p only targets and binds to MGMT and that this
specificity is of great value for the prediction of TMZ resistance
(41). In addition, miR-198 also
reduces the expression of MGMT in glioma cells, thereby enhancing
the cytotoxicity of TMZ. However, the overexpression of TGF-β1
hinders the splicing and maturation of miR-198 by inhibiting
KH-type splicing regulatory protein (KSRP) in human epidermal
keratinocytes, thereby promoting MGMT demethylation and glioma drug
resistance (42). EMT is a marker
of malignant tumours. Its biological process has a vital role in
the evaluation of malignant phenotypes and the clinical treatment
of gliomas. miRNAs also regulate the EMT process, thereby reversing
the resistance of gliomas to TMZ. For instance, miR-26b
overexpression reverses the EMT of drug-resistant glioma cells by
targeting and binding to WEE1 G2 checkpoint kinase, thereby
increasing TMZ sensitivity in drug-resistant cells (43). Cathepsin B (CTSB) has been
demonstrated to be a direct target of miR-140. miR-140 inhibits the
EMT of tumour cells and enhances the cytotoxicity of TMZ by
reducing the expression of CTSB (44). In another study, miR-128-3p
expression was observed to be downregulated in glioma tissues and
cell lines. Overexpression of miR-128-3p downregulated the
expression of the EMT proteins C-Met, platelet-derived growth
factor receptor α, Notch1 and Slug; as a result, the sensitivity of
glioma cells to TMZ was enhanced (45).
Autophagy is a turnover process based on the
lysosomal degradation of intracellular substances. Autophagy has
been indicated to have an important role in numerous aspects of
gliomas, particularly in drug-induced stress. miRNA-mediated
autophagy has a key regulatory role in glioma drug resistance.
Studies have indicated that overexpression of miR-29b enhances the
sensitivity of U87MG and U251 cells to TMZ by inducing apoptosis
and autophagy in these tumour cells. Consistent with the
conclusions of in vitro experiments, the results of an in
vivo nude mouse model of xenograft tumours confirmed that
overexpression of miR-29b effectively inhibited tumour growth and
enhanced the cytotoxicity of TMZ (46). Another study reported that under
hypoxic conditions, HIF-1α affected the chemoresistance of tumour
cells by negatively regulating the expression of miR-224-3p.
Autophagy-related gene 5 (ATG5) is a direct target of miR-224-3p.
High miR-224-3p expression reverses chemoresistance in LN229 cells
and U251 cells by inhibiting ATG5-mediated hypoxic autophagy
(47). In addition, high miR-519a
expression increases the sensitivity of glioma cells to TMZ
chemotherapy and such chemosensitization is achieved through
miR-519a-mediated inhibition of the STAT3/Bcl-2/beclin-1 pathway
and the regulation of autophagy and apoptosis (48). There is a large and complex
interaction network between drug transport and TMZ resistance.
Strategies to reverse the resistance of gliomas to TMZ through
miRNA-mediated transporters have been widely studied. For instance,
upregulation of miR-302c enhances the cytotoxicity of TMZ through
the targeted inhibition of the transporter protein P-gp in
drug-resistant glioma cells (49). Another similar study reported that
the expression of ABC subfamily C member 1 (ABCC1) was upregulated
in drug-resistant tumour cells. Overexpression of miR-1268a
reversed this upregulation and inhibited the translation of ABCC1,
thereby enhancing the chemosensitivity of drug-resistant cells to
TMZ (50). Wnt5a is the key
target of miR-129-5p. Overexpression of miR-129-5p blocks the
activation of the protein kinase C (PKC)/ERK/NF-κB and c-Jun
N-terminal kinase signalling pathways by inhibiting Wnt5a, thereby
reversing the resistance of glioma cells to TMZ (51).
miRNAs are involved in DDP
resistance
DDP is the most commonly used chemotherapeutic drug
in the salvage treatment of recurrent gliomas. The main target of
DDP is DNA. DDP covalently binds to the purine bases guanine and
adenine to form intrastrand adducts, thus inhibiting DNA
replication and transcription and causing DNA damage. Gliomas are
highly sensitive to DDP. However, with the initiation of various
repair mechanisms in tumour cells, drug resistance has seriously
limited the clinical efficacy of DDP. Therefore, the development of
strategies to reduce DDP resistance during treatment has become a
focus of research. To date, a number of studies have explored
miRNA-mediated DDP resistance in gliomas (Table II).
| Table IImiRNAs involved in cisplatin
resistance. |
Table II
miRNAs involved in cisplatin
resistance.
| miRNA | Direction of
differential expression | Genes and
pathways | Mechanism | (Refs.) |
|---|
| miR-936 | ↓ | - | - | (52) |
| miR-205 | ↓ | E2F1 | - | (53) |
| miR-451 | ↓ | MMP-2 | - | (54) |
| miR-186 | ↓ | YY1 | GSCs | (55) |
| miR-29a | ↓ | CD133 | GSCs | (56) |
| miR-22 | ↓ | SNAIL1 | Apoptosis | (57) |
| miR-107 | ↓ | mTOR | Apoptosis | (58) |
| miR-501-3p | ↓ | MYCN | Apoptosis | (59) |
| miR-128 | ↓ | JAG1/Bcl-2 | Apoptosis | (60) |
A variety of miRNAs (such as miR-936, miR-205 and
miR-451) are able to act as tumour suppressors to reverse DDP
resistance in gliomas. According to a previous study, the
expression of miR-936 was reduced in glioma tissues and cell lines
and the restoration of miR-936 expression directly inhibited the
chemoresistance of gliomas to DDP (52). Li et al (53) performed a similar study and their
results indicated that DDP treatment decreases miR-205 expression
in glioma cells and that the expression of miR-205 was lower in
DDP-resistant cell lines than in DDP-sensitive cell lines. The
overexpression of miR-205 in drug-resistant U87MG cells directly
enhances apoptosis and cell cycle arrest and restores the
sensitivity of tumour cells to DDP by targeting and binding to the
3′UTR of E2F1 (53). In addition,
Alural et al (54)
reported that erythropoietin enhanced the proliferation, invasion
and DDP chemoresistance of glioma cells by downregulating the
expression of miR-451. Restoration of the expression of miR-451
significantly reversed these effects. In this process, matrix
metallopeptidase 2 (MMP-2) is the direct target of miR-451
(54). GSCs are a group of cells
with self-renewal ability. New evidence indicates that GSC-related
miRNAs are the key mediators of glioma chemoresistance. The
expression of miR-186 is significantly reduced in glioma tissues.
Yin Yang 1 (YY1) is a molecular marker of GSCs. The overexpression
of miR-186 inhibits the development of the GSC phenotype by
targeting and binding to YY1, thereby reversing the DDP resistance
of gliomas (55). In addition,
CD133-positive GSCs exhibit stronger resistance to DDP treatment.
The overexpression of miR-29a improves CD133-mediated
chemoresistance and increases the sensitivity of gliomas to DDP
(56).
Certain tumour suppressor genes are able to reverse
the DDP resistance of gliomas, mainly by regulating apoptosis.
Exogenous miR-22 mimics induce cell cycle arrest in tumour cells by
targeting and binding to SNAIL1, thereby enhancing the sensitivity
of U87MG cells to DDP (57).
Furthermore, the expression of miR-107 was observed to be decreased
in glioma tissue and the expression of mTOR was significantly
increased. Compared with those in parental U251 cells, the
expression levels of miR-107 and mTOR exhibited similar trends in
drug-resistant U251 strains. Overexpression of miR-107 in
vitro promoted apoptosis of drug-resistant cells by inhibiting
the expression of mTOR and survivin, thus significantly enhancing
DDP-mediated cytotoxicity (58).
Similarly, miR-501-3p targets and binds to the 3′-UTR of MYCN to
promote DDP-induced apoptosis and proliferation arrest of glioma
cells. Restoration of MYCN expression reversed the promoting effect
of miR-501-3p (59). In addition,
low miR-128 expression in glioma tissues and cell lines was able to
reverse DDP resistance. The potential mechanism was determined to
be as follows: Restoration of miR-128 expression increases the
expression of Bax and decreases the expression of Bcl-2 by
targeting and binding to the jagged canonical notch ligand 1
molecular locus, which promotes tumour cell apoptosis and S-phase
arrest and eventually enhances DDP-mediated cytotoxicity (60).
miRNAs are involved in mono drug
resistance to nitrosoureas
Nitrosourea drugs have gradually become second-line
drugs in glioma chemotherapy due to the wide application of TMZ and
the limitations of their efficacy and side effects. However,
monochemotherapy with nitrosoureas (formustine, carmustine and
nimustine) or combined chemotherapy with PVC (procarbazine +
lomustine + vincristine) remains the best choice for the treatment
of recurrent and TMZ-resistant gliomas (61). As alkylating agents with
broad-spectrum antitumour activity, nitrosourea drugs may penetrate
the BBB due to their nonionic properties and high lipid solubility.
Nitrosourea drugs alkylate tumour cell DNA at multiple sites,
inducing the formation of DNA crosslinking and the occurrence of
single-strand or double-strand breaks, inhibiting DNA repair and
ultimately altering the structure of DNA and proteins (61,62). Various miRNAs have been reported
to be related to nitrosourea resistance in gliomas (Table III).
| Table IIImiRNAs involved in mono drug
resistance to nitrosoureas. |
Table III
miRNAs involved in mono drug
resistance to nitrosoureas.
| miRNA | Direction of
differential expression | Genes and
pathways | Drug | (Refs.) |
|---|
| miR-21 | ↑ | Spry2 | Carmustine | (63) |
| miR-221 | ↑ | PTEN | Carmustine | (64) |
| miR-181a | ↓ |
Bcl-2/Caspase-9 | Carmustine | (65) |
Several carcinogenic miRNAs have been indicated to
promote the chemoresistance of glioma cells to nitrosoureas. For
instance, the expression of miR-21 is significantly increased in
drug-resistant glioma cells and the abnormal expression of miR-21
is the major cause of drug resistance in gliomas. In this process,
miR-21 increases the resistance of gliomas to carmustine (BCNU) by
reducing the expression of Spry2 (63). Similarly, miR-221 has been
indicated to be highly expressed in glioma cells and BCNU-resistant
cells. miR-221 targets and binds to the 3′-UTR of phosphatase and
tensin homologue and promotes the proliferation and BCNU
chemoresistance of tumour cells through the PI3K/AKT signalling
pathway (64). miR-181a, which is
expressed at a low level in U373 cells, binds to BCNU drug
molecules, thus inhibiting cell migration and proliferation by
downregulating MMP-2 and BTB domain and CNC homologue 1. In
addition, miR-181a has been reported to increase BCNU-induced
apoptosis and cell cycle arrest of U373 cells by regulating the
expression of caspase-9, Bcl-2 and sirtuin 1 (SIRT1) (65).
miRNAs are involved in the resistance to
plant-derived anti-cancer drugs
For thousands of years, Chinese herbal medicine has
been used to provide remedies for various diseases. It has been
widely applied in disease prevention and treatment. The antitumour
effects of Chinese herbal medicine have always been a medical
issue. With continuous developments in science and technology, the
pharmacodynamic components of natural plants have gradually been
revealed and researched. Since curcumin, paclitaxel and β-elemene
have been proved to have significant antitumor effects, the
anti-glioma effect of plant extracts has attracted significant
attention from medical researchers (66). A large number of clinical studies
have indicated that these natural extracts may not only have
antitumour activities but also alleviate adverse reactions to
chemoradiotherapy, improve the quality of life of patients and
reduce the recurrence rate of tumours (67). In addition, an increasing number
of in vivo and in vitro studies have explored the
relationship between miRNAs and tumour sensitivity to Chinese
herbal extracts (Table IV).
| Table IVmiRNAs involved in the resistance to
plant-derived anticancer drugs. |
Table IV
miRNAs involved in the resistance to
plant-derived anticancer drugs.
| miRNA | Direction of
differential expression | Genes and
pathways | Drug | (Refs.) |
|---|
| miR-374a | ↑ | FOXO1 | Etoposide | (68) |
| miR-218-2 | ↑ | CDC27 | β-lapachone | (69) |
| miR-21 | ↑ | LRRFIP1 | Teniposide | (70) |
| miR-326 | ↓ | SHH/GLI1 | Curcumin | (71) |
| miR-34a | ↓ | PD-L1 | Paclitaxel | (72) |
| miR-204-3p | ↓ |
IGFBP2/AKT/Bcl-2 | Xanthohumol | (73) |
| miR-7-1-3p | ↓ | PKCa, iNOS | Luteolin,
Silibinin | (74) |
| miR-15b | ↓ | MMP-9 | Mangiferin | (75) |
The expression of certain carcinogenic miRNAs
directly affects the efficacy of chemotherapy. For instance, the
expression of miR-374a is significantly increased in malignant
gliomas. Knockdown of miR-374a directly increased the expression of
forkhead box protein O1 (FOXO1) in glioma cells, enhancing the
cytotoxicity of etoposide (68).
miR-218 is a tumour suppressor gene. However, miR-218-2 is a
cancer-promoting gene that is highly expressed in glioma tissues
and cells and is positively associated with the growth, invasion,
migration and β-lapachone resistance of glioma cells. In terms of
the underlying mechanism, miR-218-2 promotes the chemoresistance of
U87MG and U251 cells to β-lapachone by reducing the expression of
the cell division cycle 27 (CDC27) gene (69). In addition, miR-21 acts as a
modulator of teniposide chemoresistance. LRR binding friend
leukaemia integration 1 transcription factor (FLI1) interacting
protein 1 (LRRFIP1) has been identified as a direct target of
miR-21, and miR-21 enhances the resistance of glioma cells to
teniposide by targeting and binding to the 3′-UTR of LRRFIP1
(70).
Yin et al (71) reported that overexpression of
miR-326 increased the chemosensitivity of malignant glioma cells to
the anticancer drug curcumin. The results of in vitro
experiments demonstrated that high miR-326 expression reduces the
viability of tumour cells by inhibiting the sonic hedgehog/GLI
family zinc finger 1 signalling pathway, thereby enhancing the
cytotoxicity of curcumin to U87MG and U251 cells (71). The expression of programmed
death-ligand 1 (PD-L1) is significantly increased in
paclitaxel-resistant U87MG cells. By directly acting on the 3′-UTR
of PD-L1, miR-34a inhibits the progression and chemoresistance of
tumour cells (72). Similarly,
the expression of miR-204-3p is decreased in glioma cell lines.
miR-204-3p targets and binds to the 3′-UTR of IGF binding protein 2
(IGFBP2) and enhances xanthohumol-induced tumour cell apoptosis by
inhibiting the IGFBP2/AKT/Bcl-2 pathway (73). The application of rapamycin
created a starvation state in glioma cells and induced autophagy.
High miR-7-1-3p expression enhanced the antitumour activity of
synergistic silibinin and luteolin treatment by inhibiting the
expression of PKCα and inducible nitric oxide synthase in GBM
cells. More importantly, miR-7-1-3p promoted apoptosis by
inhibiting rapamycin-induced autophagy (74). In addition, mangiferin inhibited
the expression of MMP-9 in U87MG cells by upregulating miR-15b,
thereby promoting apoptosis. Conversely, the use of MMP-9 agonists
and anti-miR-15b inhibitors significantly reduced the inhibitory
effect of mangiferin on U87MG cells (75).
miRNAs are involved in resistance to
molecular targeted drugs
Tumour-related molecular targeted therapy has become
an important research field. In particular, molecular targeted
therapy has achieved great success in the treatment of non-small
cell lung cancer, malignant melanoma and chronic myeloid leukaemia
(76), findings that have
important guiding significance for the application of molecular
targeted drugs in the treatment of malignant gliomas. Compared with
traditional drugs, molecular targeted drugs are less toxic and only
inhibit tumour cells. In terms of their mechanism of action,
molecular targeted drugs accurately regulate specific receptors,
key genes and regulatory molecules in tumour cells. Therefore, the
application of molecular targeted drugs has become a novel
treatment strategy for gliomas and is expected to improve the
prognosis of patients with glioma. The mechanisms underlying the
resistance of gliomas to molecular targeted drugs remain to be
fully elucidated. However, certain miRNAs have been identified to
directly participate in the regulation of the cytotoxicity of
molecular targeted drugs (Table
V).
| Table VmiRNAs involved in resistance to
molecular targeted drugs. |
Table V
miRNAs involved in resistance to
molecular targeted drugs.
| miRNAs | Direction of
differential expression | Genes and
pathways | Drug | (Refs.) |
|---|
| miR-21,
miR-10b | ↑ | - | Bevacizumab | (78) |
| miR-145 | ↓ | P-gp, Bcrp | Sunitinib | (79) |
| miR-302a,
miR-520b | ↓ | AKT1, PIK3CA,
SOS1 | Sunitinib | (80) |
| miR-106a | ↑ | MDR1, MRP1,
GST-π | Gefitinib | (81) |
| miR-450a | ↓ | EGFR | Gefitinib | (82) |
| miR-566 | ↑ | VHL | Nimotuzumab | (83) |
| miR-21 | ↑ | VHL/β-catenin,
PPARα/AP | Nimotuzumab | (84) |
| miR-203 | ↓ | SNAI2 | Imatinib | (85) |
| miR-296-3p | ↓ | EAG1 | Imatinib | (86) |
Bevacizumab is a recombinant monoclonal
immunoglobulin G (IgG) antibody. It specifically blocks the binding
of VEGF-A to the VEGF receptor, thereby reducing angiogenesis and
inhibiting tumour growth. In 2009, bevacizumab was approved by the
Food and Drug Administration (FDA) of the US for the treatment of
recurrent GBM (77). Siegal et
al (78) performed a
longitudinal study of the circulating miRNA levels in patients with
glioma treated with bevacizumab. During treatment, the levels of
miR-10b and miR-21 were negatively associated with changes in
tumour diameter and bevacizumab had poor efficacy in the treatment
of patients with high miR-10b and miR-21 expression (78). However, the specific underlying
mechanisms of action remain elusive and require to be further
investigated. Sunitinib is an anti-angiogenic tyrosine kinase
inhibitor. Transfection with miR-145 mimics promoted the cytotoxic
effects of sunitinib on GBM by directly acting on the 3′-UTR of
P-gp and breast cancer resistance protein (Bcrp) (79). Similarly, miR-302a and miR-520b
mimics directly targeted and bound to the 3′-UTR of AKT1, PI3K
catalytic subunit alpha (PIK3CA) and son of sevenless homologue 1
(SOS1). These miRNAs inhibit the expression of receptor tyrosine
kinase mediators (AKT1, PIK3CA and SOS1) in U87MG cells, thereby
enhancing the sensitivity of tumour cells to sunitinib and
increasing apoptosis. However, this regulatory effect of miR-302a
and miR-520b has not been observed for TMZ (80).
Growth factor receptors are important regulatory
proteins in the signalling networks of malignant gliomas.
Resistance to novel drugs that target the epidermal growth factor
receptor and platelet-derived growth factor receptor has been a
focus of scientific exploration. miR-106a is an important molecule
involved in drug resistance. miR-106a enhances the drug efflux
ability and anti-apoptotic ability of drug-resistant U87MG cells by
promoting the expression of Twist, AP-1 and Snail. Furthermore,
miR-106a positively regulates the expression of glutathione
S-transferase π to enhance the drug detoxification ability of
drug-resistant U87MG cells. Therefore, miR-106a increases the
resistance of glioma cells to gefitinib synergistically through the
abovementioned regulatory mechanisms (81). Compared with that in normal glial
cells, the expression of miR-450a was observed to be significantly
reduced in GBM. High miR-450a expression was indicated to regulate
the PI3K/AKT/mTOR signalling pathway by inhibiting the translation
of epidermal growth factor receptor (EGFR), thereby regulating
apoptosis and autophagy and enhancing the sensitivity of glioma
cells to gefitinib. In addition, the abovementioned responses
induced by miR-450a were able to be reversed by the knockdown of WD
repeat domain phosphoinositide-interacting protein 1 (WIPI1)
(82). Inhibition of the highly
expressed miR-566 in glioma cell lines negatively regulated von
Hippel-Lindau (VHL), thereby increasing the sensitivity of tumour
cells to nimotuzumab (83). In
addition, miR-21 was indicated to regulate the EGFR/AKT signal
conduction pathway through VHL/β-catenin and peroxisome
proliferator-activated receptor α/AP and blocking this regulatory
loop significantly inhibits the chemoresistance of glioma cells to
nimotuzumab. More importantly, the combined therapeutic effect of
nimotuzumab and a miR-21 inhibitor was superior to that of single
drug treatment (84). In
addition, increased Snail family transcriptional repressor 2
(SNAI2) expression was observed in imatinib-resistant GBM cells and
silencing of SNAI2 directly inhibited the EMT process and drug
resistance of tumour cells. Furthermore, miR-203 mimics increased
the sensitivity of drug-resistant tumour cells to imatinib by
targeting and binding to SNAI2 (85). As a tumour suppressor gene,
miR-296-3p reduced the proliferative activity of GBM cells by
inhibiting the proliferation regulator ether-à-go-go1, which was
conducive to reducing the resistance of GBM cells to imatinib
(86).
miRNAs are involved in resistance to
immunotherapeutic drugs
Immune checkpoint inhibitors are monoclonal
anti-bodies developed specifically for immune checkpoints. These
inhibitors induce sustained antitumour immune responses by blocking
the inhibitory effects of tumour cells on immune cells. The immune
checkpoint inhibitors approved by the FDA include ipilimumab
[cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) inhibitor],
nivolumab (PD-1 inhibitor) and avelumab (PD-L1 inhibitor) (87). The successful clinical application
of these agents has brought new life to the immunotherapeutic
treatment of malignant gliomas. According to research reports,
certain miRNAs regulate immune checkpoints in glioma cells. For
instance, compared with that in a wild-type nude mouse xenograft
model, the growth of gliomas was significantly inhibited in
miR-15a/16-deficient nude mice and the survival of the nude mice
was significantly prolonged. More importantly, a large number of
highly active and proliferative CD8+ T cells accumulated in the
tumours of the miR-15a/16-deficient nude mice. This differential
phenotype was induced by the miR-15a/16 deficiency-mediated low
expression of PD-1, T cell immunoglobulin and mucin-domain
containing-3 and lymphocyte-activation gene 3 (88). In addition, miR-138 targets and
binds to CTLA-4, PD-1 and FOXP3 in CD4+ T and CD8+ T cells,
resulting in the regression of glioma tissues in immunocompetent
nude mice (89). Tumour vaccines
have also been a hotspot of cancer research in recent years. The
mechanism of action of tumour vaccines is to introduce
tumour-related antigens into patients; this approach activates
cellular and humoural immunity and induces inflammatory responses
to enhance antitumour effects. Li et al (90) reported that miR-326 reduced the
exocrine secretion of TGF-β1 in glioma cells through the smoothened
(SMO)/Gli2 pathway, thereby improving the activity and killing
ability of T cells and enhancing the cytotoxicity of the EGFR
variant III-dendritic cell (EGFRvIII-DC) vaccine. More importantly,
compared with the EGFRvIII-DC vaccine alone, combined
administration of the EGFRvIII-DC vaccine and miR-326 achieved
better therapeutic efficacy and exerted a stronger killing effect
on U87MG cells (90) (Table VI).
| Table VImiRNAs involved in resistance to
immunotherapeutic drugs. |
Table VI
miRNAs involved in resistance to
immunotherapeutic drugs.
| miRNAs | Direction of
differential expression | Genes and
pathways | Drug | (Refs.) |
|---|
| miR-15a/16 | ↑ | PD-1, Tim-3,
LAG-3 | PD-L1
inhibitor | (88) |
| miR-138 | ↓ | CTLA-4, PD-1,
FoxP3 | CTLA-4 inhibitor,
PD-1 inhibitor | (89) |
| miR-326 | ↓ | SMO/Gli2 | EGFRvIII-DC
vaccine | (90) |
| miR-124, miR-128,
miR-146b, miR-218 | ↓ | E1A | OA-4MREs | (91) |
With the continuous development of bioengineering
technology, researchers have used gene editing technology to
transform viruses with strong replication ability into oncolytic
viruses. Various oncolytic adenoviruses promote the apoptosis of
glioma cells, with low toxicity in normal cells. These viruses
display high specificity and selectivity. To improve the
cytotoxicity and specificity of oncolytic adenoviruses, Yao et
al (91) combined the miRNA
response elements (MRE) of 4 glioma inhibitory miRNAs (miR-124,
miR-128, miR-146b and miR-218) with an oncolytic adenovirus to
construct a recombinant oncolytic adenovirus (OA-4MREs). In
vivo and in vitro experiments demonstrated that the MREs
regulated the replication ability of the virus by targeting and
binding to adenovirus early region 1A (E1A). Compared with the
proliferative adenovirus, OA-4MREs had a stronger cytotoxic effect
on glioma cells. Of note, it was also observed that OA-4MREs had no
significant cytotoxicity in normal tissues and cells and only
produced a limited number of viral offspring (91). Therefore, OA-4MREs exhibited high
safety, allowing further testing in clinical practice.
miRNAs are involved in resistance to
other drugs
Doxorubicin (Dox) has a wide range of toxic effects
on Top II. It is a highly effective broad-spectrum anticancer drug.
The mechanism of action involves the intercalation of Dox into the
DNA of tumour cells, thereby inhibiting nucleic acid replication.
An increasing amount of evidence indicates that miRNAs are related
to Dox resistance in gliomas. miR-330-3p inhibitors reduce the
proliferative, migratory and invasive capabilities of glioma cells.
The combined application of a miR-330-3p inhibitor and a low dose
of human endothelial monocyte activating peptide II increased the
permeability of the BTB by reducing the expression of zonula
occludens-1 (ZO-1), occludin and claudin-5, thereby enhancing the
chemotherapeutic efficacy of Dox on gliomas (92). As one of the first miRNAs
detected, miR-21 has been demonstrated to be highly expressed in
gliomas and to be involved in the regulation of tumour drug
resistance. For instance, therapies combining Dox and miR-21
inhibitors induced apoptosis of T98G cells resistant to Dox
chemotherapy, thereby increasing the sensitivity of T98G cells to
Dox (93). Similarly, in another
study, the highly synergistic effect of Dox and miR-21 inhibitors
was indicated to block the EMT of U87MG and LN299 cells by
enhancing the expression of the tumour suppressor genes E-cadherin,
reversion-inducing cysteine-rich protein with kazal motifs, VHL and
P21, thereby reversing Dox resistance in tumour cells (94). In addition to the above-mentioned
drugs, knockdown of miR-21 reduced the resistance of glioma cells
to tamoxifen and 5-fluorouracil (5-FU) (95,96). By contrast, knockdown of miR-302b
significantly increased the resistance of glioma cells to all-trans
retinoic acid (97) (Table VII).
| Table VIImiRNAs involved in resistance to
other drugs. |
Table VII
miRNAs involved in resistance to
other drugs.
| miRNA | Direction of
differential expression | Genes and
pathways | Drug | (Refs.) |
|---|
| miR-330-3p | ↓ | ZO-1, Occludin,
Claudin-5 | Doxorubicin | (92) |
| miR-21 | ↑ | E-cadherin, RECK,
VHL, P21 | Doxorubicin,
Tamoxifen, 5-fluorouracil | (93-96) |
| miR-302b | ↓ | - | All-trans retinoic
acid | (97) |
4. LncRNAs and glioma drug resistance
LncRNAs are ncRNAs with a length of >200
nucleotides. A number of studies have indicated that lncRNAs are
composed of promoter regions, exons, antisense sequences, enhancer
sequences, 3′- and 5′-UTRs, introns, and intergenic and intragenic
regions of the genome. LncRNAs have key roles in physiological
activities, including epigenetic regulation, cell cycle regulation
and cell differentiation regulation through multiple pathways and
mechanisms (98). These RNAs have
become a focus of genetic research. More importantly, lncRNAs have
been indicated to serve as biomarkers for the diagnosis, treatment,
prognostication, drug response prediction and other pathological
processes of gliomas by interacting with miRNAs to form a competing
endogenous RNA (ceRNA) network, participating in histone
modification and directly binding to target genes (98). In this chapter, the roles of
related lncRNAs in the development of chemotherapy resistance in
gliomas are discussed and a sufficient theoretical basis for the
comprehensive understanding of the mechanisms of tumour drug
responses are provided.
LncRNAs are involved in TMZ
resistance
A number of carcinogenic lncRNAs have been indicated
to induce TMZ resistance in gliomas. For instance, knockdown of
lncRNA zinc finger antisense 1 (ZFAS1), lncRNA LINC00021 and lncRNA
NCK1-AS1 not only inhibited tumour proliferation, migration and
invasion, but also reversed the TMZ resistance of gliomas (99-101). LncRNA TCONS_00004099 is a newly
discovered ncRNA. It inhibits TMZ-induced apoptosis by regulating
the downstream target protein tyrosine phosphatase receptor type F
(PTPRF) (102). LncRNA HOX
antisense intergenic RNA (HOTAIR) is an oncogene. It was recently
discovered that knocking out lncRNA HOTAIR significantly reduced
TMZ resistance in gliomas by targeting calcium binding and
coiled-coil domain 1 and zinc finger CCCH-type containing 10
(103). As a ceRNA, lncRNA
HOTAIR may act as an miRNA sponge. It promotes the malignant
development and TMZ resistance of gliomas by adsorbing miR-125/HK2
(104). In vitro, lncRNA
BC200 promotes TMZ resistance in tumours; this process is induced
by antagonizing miR-218-5p (105). High lncRNA miR155HG expression
is negatively associated with the quality of life of patients with
gliomas. Knockdown of miR-155HG inhibits the Wnt/β-catenin pathway
by effectively downregulating the expression of polypyrimidine
tract-binding protein 1 in tumour cells, thereby reducing the
resistance of gliomas to TMZ (106). In addition, another ceRNA
molecule, lncRNA colon cancer associated transcript 2 (CCAT2), has
been indicated to regulate the expression of checkpoint kinase 1
(Chk1) through the sponge adsorption of miR-424, thereby promoting
TMZ resistance in gliomas (107)
(Table VIII).
| Table VIIILncRNAs involved in temozolomide
resistance. |
Table VIII
LncRNAs involved in temozolomide
resistance.
| LncRNAs | Direction of
differential expression | Genes and
pathways | Mechanism | (Refs.) |
|---|
| ZFAS1 | ↑ | - | - | (99) |
| LINC00021 | ↑ | - | - | (100) |
| NCK1-AS1 | ↑ | - | - | (101) |
| TCONS_00004099 | ↑ | PTPRF | - | (102) |
| HOTAIR | ↑ | CALCOCO1, ZC3H10,
miR-125/HK2 | - | (103,104) |
| BC200 | ↑ | miR-218-5p | - | (105) |
| MIR155HG | ↑ |
PTBP1/Wnt/β-catenin | - | (106) |
| CCAT2 | ↑ | miR-424/Chk1 | - | (107) |
| MSC-AS1 | ↑ |
miR-373-3p/CPEB4 | Apoptosis | (108) |
| SOX2OT | ↑ | SOX2 | Apoptosis | (109) |
| LINC00461 | ↑ | miR-216a/AQP4 | Apoptosis | (110) |
| 00174 | ↑ |
miR-138-5p/SOX9 | Apoptosis | (111) |
| FOXD2-AS1 | ↑ | MGMT | DNA damage
repair | (112) |
| H19 | ↑ | miR-198/MGMT | DNA damage repair,
EMT | (113,116) |
| HOXD-AS2 | ↑ | miR-198/MGMT | DNA damage
repair | (113) |
| TP73-AS1 | ↑ | ALDH1A1 | GSCs | (114) |
| NEAT1 | ↑ |
let-7g-5p/MAP3K1 | GSCs | (115) |
| PVT1 | ↑ | miR-365/ELF4 | GSCs | (116) |
| DLEU1 | ↑ | ZEB1, N-cadherin,
Snail, P62, ATG7 | EMT, Autophagy | (117) |
| MALAT1 | ↑ |
miR-203/miR-140 | EMT | (118) |
| CRNDE | ↑ | P62 | Autophagy | (119) |
| LINC00470 | ↑ |
miR-134/MYC/ABCC1 | Drug transport and
metabolism | (120) |
| KCNQ1OT | ↑ | miR-761/PIM1 | Drug transport and
metabolism | (121) |
| LIFR-AS1 | ↓ | miR-4262/NF-κB | - | (122) |
| WT1-AS | ↓ |
miR-494-3p/p-AKT | - | (123) |
| CACS2 | ↓ |
miR-193a-5p/mTOR | Autophagy | (124) |
| TUG1 | ↓ | EZH2 | EMT, GSCs | (125) |
| TUSC7 | ↓ | MDR1, miR-10a | Apoptosis | (126) |
Certain lncRNAs may also act as regulators of
apoptosis and MGMT to mediate chemotherapy resistance in gliomas.
For instance, knockdown of lncRNA MSC-AS1 regulated the
miR-373-3p/cytoplasmic polyadenylation element binding protein 4
axis in glioma cells through the PI3K/Akt signalling pathway,
significantly enhancing the induction of apoptosis during TMZ
treatment and reducing the half-maximal inhibitory concentration
(IC50) of TMZ (108).
LncRNA SOX2 overlapping transcript (SOX2OT) is not only a biomarker
for predicting the high recurrence risk and poor prognosis of
gliomas, but also an important regulator of chemoresistance to TMZ.
In addition, in vivo and in vitro studies have
indicated that lncRNA SOX2OT directly regulates the SOX2-bound AlkB
homolog 5 (ALKBH5) protein. As a result, SOX2 is demethylated,
further inhibiting apoptosis and enhancing the resistance of tumour
cells to TMZ chemotherapy (109). miR-216a acts as a tumour
suppressor. Silencing of lncRNA LINC00461 regulates cellular
aquaporin-4 by upregulating the expression of miR-216a, ultimately
leading to apoptosis and the antagonization of TMZ resistance in
gliomas (110). In addition,
lncRNA 00174, as a ceRNA molecule, acts as a 'molecular sponge' in
the process of drug treatment; it directly adsorbs miR-138-5p to
upregulate SOX9 protein expression and promote the development of
TMZ resistance in gliomas (111). High MGMT expression reduced
TMZ-mediated cytotoxicity. Furthermore, MGMT induces its own
methylation during the process of guanine demethylation and
immediately becomes inactivated. Therefore, it is called a 'suicide
enzyme'. LncRNA FOXD2-AS1 is overexpressed in recurrent gliomas and
its high expression is significantly associated with poor prognosis
of patients. Silencing lncRNA FOXD2-AS1 not only reduced the
proliferative and metastatic abilities of glioma cells but also
induced hypermethylation of the MGMT promoter region and enhanced
the sensitivity of glioma cells to TMZ (112). In addition, lncRNA H19 and
lncRNA HOXD-AS2 regulated functional mechanisms different from the
abovementioned ceRNA mechanism. As an RNA-binding protein, KSRP
binds to follistatin-like 1 and promotes the formation of mature
miR-198. TGF-β1 upregulates the expression of H19 and HOXD-AS2
through SMAD signalling and prevents the maturation of miR-198,
ultimately decreasing the expression of miR-198 and increasing the
expression of MGMT, significantly enhancing the resistance of
gliomas to TMZ chemotherapy (113).
GSCs are an important factor in glioma heterogeneity
and are closely related to tumour recurrence and treatment
resistance. An increasing number of lncRNAs have been indicated to
regulate tumour drug resistance through stemness-maintaining
transcription factors and stem cell-related regulatory pathways.
LncRNA TP73-AS1 is an oncogene that is overexpressed in GBM
tissues, particularly in GSC tissues. It promotes the TMZ
resistance of GSCs through the targeted regulation of the metabolic
gene ALDH1A1 (114). LncRNA
nuclear-enriched abundant transcript 1 (NEAT1) has been confirmed
to have a cancer-promoting effect. In addition, it acts as a ceRNA
and interferes with the binding of the tumour suppressor gene
let-7g-5p and MAPK kinase kinase 1, thus promoting the malignant
development of GSCs during TMZ treatment (115). The lncRNA plasmacytoma variant
translocation 1 (PVT1) promotes the proliferation, migration and
invasion of glioma cells by activating SOX2. In addition, lncRNA
PVT1 positively regulates the expression of E74-like ETS
transcription factor 4 (ELF4) in GCSs through the sponge adsorption
of miR-365. High ELF4 expression further induces asymmetric cell
division of GCSs, thereby enhancing TMZ resistance (116). EMT is closely related to glioma
invasion, migration and chemotherapy resistance, and lncRNAs may
act as key factors that regulate these biological behaviours.
LncRNA H19 has a central role in inducing TMZ chemoresistance in
gliomas. In vitro, the induction of EMT, the activation of
oncogenic signalling pathways and the alteration of the tumour
microenvironment are all important mechanisms by which H19
participates in chemotherapy resistance (117). In addition, silencing lncRNA
deleted in lymphocytic leukaemia 1 (DLEU1) inhibits the TMZ-induced
EMT process by regulating the expression of marker proteins (i.e.,
zinc finger E-box-binding homeobox 1, N-cadherin and Snail) in
glioma cells, thereby increasing the sensitivity of tumour cells to
TMZ (118). Similarly, a study
by Baspinar et al (119)
indicated that lncRNA metastasis associated lung adenocarcinoma
transcript 1 (MALAT1) reduces the expression of miR-203, thereby
inducing EMT by enhancing the synthesis of thymidylate synthase and
ultimately promoting the resistance of glioma cells to TMZ.
Furthermore, knocking out MALAT1 enhanced the permeability of the
BTB by upregulating the expression of miR-140, which markedly
improved the targeting effect and biological efficacy of TMZ in the
treatment of tumours and provides novel gene therapy strategies for
gliomas (119).
Autophagy is a highly conserved process. Numerous
molecular complexes and oncoproteins have key roles in various
stages of autophagy. Therefore, it is not surprising that lncRNAs
directly participate in the regulation of autophagy by regulating
the activity and expression of the abovementioned molecules. The
lncRNA colorectal neoplasia differentially expressed (CRNDE) is
highly expressed in TMZ-resistant patients and knocking down lncRNA
CRNDE significantly enhanced the TMZ chemosensitivity of glioma
cells. In terms of the underlying mechanisms, silencing CRNDE
directly reduces the expression of light chain (LC)3II/I, beclin1
and autophagy-related 5 (ATG5), thereby inhibiting autophagy
related to PI3K/AKT/mTOR pathway activation and ABCG2 expression
(120). In the study by Lv et
al (118), lncRNA deleted in
lymphocytic leukemia 1 (DLEU1) was confirmed to have a crucial role
in GBM progression and TMZ resistance. Knocking down DLEU1
inhibited autophagy by negatively regulating the expression of P62
and ATG7 in GBM cells and increased the sensitivity of tumour cells
to TMZ by inducing apoptosis. The reduction in the accumulation of
chemotherapeutic drugs is one of the main mechanisms of drug
resistance. For instance, lncRNA LINC00470 directly targets the
miR-134/MYC pathway to enhance the resistance of glioma cells to
TMZ. During TMZ treatment, the miR-134/MYC pathway upregulates the
expression of ABCC1 in glioma cells and there is a positive
correlation between MYC and ABCC1 expression (121). The interaction between lncRNA
KCNQ1 opposite strand/antisense transcript 1 (KCNQ1OT1) and miR-761
also has an important role in the development, progression and
chemoresistance of GBM. KCNQ1OT1 upregulates PIM1 expression
through the sponge adsorption of miR-761. Overexpression of PIM1
directly induces the activation of multidrug resistance mutation 1
(MDR1), c-MYC and survivin, thereby enhancing TMZ resistance
(122).
A variety of tumour suppressor lncRNAs reverse TMZ
resistance in gliomas. For instance, in glioma tissues and cell
lines, low expression of lncRNA leukaemia inhibitory factor
receptor-AS1 enhances the sensitivity of tumour cells to TMZ by
targeting the miR-4262/NF-κB axis (123). In U87MG cells overexpressing
lncRNA Wilms tumour 1 AS, the IC50 of the
chemotherapeutic agent TMZ is significantly reduced. The underlying
mechanism involves the inhibition of phosphorylating AKT activation
through the sponge adsorption of miR-494-3p, reversing chemotherapy
resistance in GBM (124). Jiang
et al (125) reported
that lncRNA cancer susceptibility 2 (CACS2) may be used as a
biomarker for the diagnosis and treatment of gliomas. Furthermore,
lncRNA CACS2 acts as a ceRNA molecule to reverse the resistance of
tumour cells to TMZ, primarily through the miR-193a-5p/mTOR
signalling pathway-mediated inhibition of autophagy. The results of
in vivo and in vitro experiments indicated that
lncRNA taurine upregulated gene 1 inhibits the EMT process and GSC
phenotype of gliomas by downregulating the expression of enhancer
of zeste homologue 2 (EZH2), thereby reducing TMZ resistance
(126). In addition, lncRNAs
have been indicated to affect the apoptosis of TMZ-resistant glioma
cells. For instance, lncRNA tumour suppressor candidate 7 (TUSC7)
is a tumour suppressor gene that reduces the TMZ resistance of
glioma cells by inhibiting the expression of MDR1. Furthermore,
lncRNA TUSC7 induces apoptosis through targeted silencing of
miR-10a expression, thereby increasing the sensitivity of
drug-resistant cells to TMZ (127).
LncRNAs are involved in DDP
resistance
Evidence indicates that certain carcinogenic or
tumour suppressor lncRNAs mediate the resistance of gliomas to DDP
chemotherapy. For instance, the lncRNA differentiation antagonizing
non-protein coding RNA, which is highly expressed in drug-resistant
glioma cell lines, activates the PI3K/AKT/NF-κB signalling pathway
by inducing the expression of AXL in drug-resistant tumour cells,
thereby promoting resistance to DDP. Similar results were obtained
in an in vivo experiment (128). LncRNA CCAT2-mediated promotion
of tumour cell chemoresistance is the major cause of the poor
overall survival and progression-free survival of patients with
GBM. In terms of the mechanism, CCAT2 acts as a sponge for miR-424.
It enhances the chemotherapy resistance of GBM cells to DDP by
inhibiting the molecular degradation of Chk1 by miR-424 (107). Similarly, the role of lncRNA
ZFAS1/miR-432-5p in glioma chemoresistance has also been confirmed
(129). LncRNA HOXD cluster AS1
(HOXD-AS1) is an oncogene. High HOXD-AS1 expression has been
observed in DDP-resistant glioma clinical specimens and cell lines.
LncRNA HOXD-AS1 jointly promotes the resistance of tumour cells to
DDP by competitively binding to miR-204. Furthermore, knocking down
HOXD-AS1 inhibits the GSC phenotype and the EMT process, thereby
enhancing DDP sensitivity and reducing tumour cell proliferation,
migration and invasion (130).
In addition, certain lncRNAs affect the DDP resistance of glioma
cells by regulating apoptosis-related pathways. For instance,
lncRNA urothelial carcinoma associated 1, which is highly expressed
in drug-resistant glioma cell lines, reduces DDP-induced apoptosis
and cell cycle arrest by activating Wnt/β-catenin signalling,
thereby promoting the chemoresistance of tumour cells to DDP
(131). Similarly, the lncRNA
CRNDE has been reported to inhibit DDP-induced apoptosis by
competitively binding to miR-29c-3p in glioma cells (132) (Table IX).
| Table IXLncRNAs involved in cisplatin
resistance. |
Table IX
LncRNAs involved in cisplatin
resistance.
| LncRNA | Direction of
differential expression | Genes and
pathways | Mechanism | (Refs.) |
|---|
| DANCR | ↑ | AXL | - | (127) |
| CCAT2 | ↑ | miR-424/Chk1 | - | (107) |
| ZFAS1 | ↑ | miR-432-5p | - | (128) |
| HOXD-AS1 | ↑ | miR-204 | EMT, GSCs | (129) |
| UCA1 | ↑ | Wnt/β-catenin | Apoptosis | (130) |
| CRNDE | ↑ | miR-29c-3p | Apoptosis | (131) |
| GAS5 | ↓ | mTOR | Autophagy | (132) |
| MEG3 | ↓ | ATG5 | Autophagy | (133) |
| AC023115.3 | ↓ | miR-26a/GSK3β | Autophagy | (134) |
In addition to the oncogenic lncRNAs related to DDP
resistance, there are several tumour suppressor lncRNAs that
enhance the sensitivity of tumour cells to DDP. All of these tumour
suppressor lncRNAs are closely related to autophagy in glioma
cells. The expression of mTOR is downregulated in drug-resistant
glioma cell lines, promoting DDP-induced autophagy. The
downregulation of mTOR ultimately mediates an increase in the
expression of autophagy-related protein LC3II and a decrease in the
expression of autophagy substrate p62 in tumour cells, thereby
enhancing chemotherapy resistance to DDP. Of note, overexpression
of lncRNA growth arrest specific 5 activates mTOR signalling and
restores the sensitivity of glioma cells to DDP (133). LncRNA maternally expressed 3
enhances the chemical sensitivity of U87MG cells to DDP by
inhibiting the expression of ATG5. In terms of the mechanism, ATG5
is a key molecule in the formation of autophagic vacuoles.
Silencing ATG5 significantly inhibits autophagy in tumour cells and
ultimately increases the cytotoxic effect of DDP in U87MG cells
(134). LncRNA AC023115.3 is
another lncRNA that sensitizes glioma cells to DDP in vivo
and in vitro. It inhibits tumour cell autophagy and promotes
DDP-induced apoptosis through the miR-26a/glycogen synthase kinase
3β signalling pathway (135).
LncRNAs are involved in resistance to
other drugs
In addition to mediating the resistance of gliomas
to TMZ and DDP, certain lncRNAs are also involved in the resistance
of gliomas to other chemotherapeutic drugs, such as natural
anticancer drugs and molecular targeted drugs. Both lncRNA CCAT2
and Chk1 are highly expressed in teniposide-resistant glioma cells.
The potential mechanism of drug resistance is the sponge adsorption
of miR-424 by CCAT2, which enhances the chemoresistance of glioma
cells to teniposide through the upregulation of Chk1 expression
(107). A recent study showed
that the overexpression of lncRNA H19 plays an important role in
the development of curcumin resistance. Knocking down lncRNA H19
reverses curcumin resistance by regulating vitamin D receptor
(VDR), and VDR is a direct target of miR-143 in gliomas (136). Lv et al (137) performed a similar study,
according to which high lncRNA FOXD2-AS1 expression increased the
me3 modification ability of H3K27 by mediating the activation of
EZH2 in glioma cells, thereby promoting the chemoresistance of
tumour cells to curcumin. Curcumin is a natural antitumour drug. Of
note, combined therapy with curcumin and TMZ significantly enhanced
the cytotoxicity of the drugs (137). Silencing lncRNA NEAT1 promoted
the sensitivity of U87MG cells to isoliquiritigenin by inhibiting
tumour angiogenesis. Knocking down NEAT1 significantly enhanced the
inhibitory effect of miR-194-5p on the AKT-fibroblast growth factor
2/TGF-β/VEGF signalling pathway, thereby reversing the resistance
of glioma cells to isoliquiritigenin (138). Compared with that in parental
SHG-44 cells, lncRNA PVT1 is highly expressed in drug-resistant
SHG-44 cells. Overexpression of PVT1 promotes paclitaxel
resistance; the primary mechanism is the enhancement of paclitaxel
sensitivity through the regulation of apoptosis (139). In addition, lncRNA microvascular
invasion in hepatocellular carcinoma (MVIH) was reported to be
involved in the resistance of glioma cells to cediranib. It
enhanced the resistance of gliomas to cediranib through the sponge
inhibition of miR-137. By contrast, knocking down MVIH
significantly reduced glycolysis and cell proliferation and
sensitized glioma cells to cediranib (140) (Table X).
| Table XLncRNAs involved in resistance to
other drugs. |
Table X
LncRNAs involved in resistance to
other drugs.
| LncRNA | Direction of
differential expression | Genes and
pathways | Drug | (Refs.) |
|---|
| CCAT2 | ↑ | miR-424 | Teniposide | (107) |
| H19 | ↑ | miR-143/VDR | Curcumin | (135) |
| FOXD2-AS1 | ↑ | EZH2 | Curcumin | (136) |
| NEAT1 | ↑ | miR-194-5p |
Isoliquiritigenin | (137) |
| PVT1 | ↑ | - | Paclitaxel | (138) |
| MVIH | ↑ | miR-137 | Cediranib | (139) |
5. CircRNAs and drug resistance of
gliomas
In recent years, circRNAs have received widespread
attention as a new type of ncRNA. Unlike traditional linear RNAs,
the 3′ and 5′ ends of circRNAs are covalently connected. Therefore,
circRNAs form covalently closed loop structures that resist
cleavage by ribonucleases and display stronger stability,
conservation and tissue/cell specificity (141). Due to these characteristics,
circRNAs, as molecular targets for the malignant progression of
gliomas, have demonstrated great potential in the regulation of
tumourigenesis, tumour proliferation, tumour invasion and
prognosis. For instance, circPIP5K1A has been indicated to be
upregulated in gliomas. It induces the expression of TCF12 in
glioma cells by competitively binding to miR-515-5p, thereby
promoting tumour proliferation, invasion and EMT (142). Similarly, circCDC45 promotes the
expression of colony stimulating factor 1 through the sponge
adsorption of miR-485-5p. It has a key role in the entire malignant
progression of GBM and has become a potential marker for the
diagnosis and treatment of gliomas (143). In addition, with the development
of chemoresistance and the need for chemotherapy, an increasing
amount of evidence indicates that circRNAs also have important
roles in the drug resistance mechanisms of gliomas.
CircRNAs are involved in TMZ
resistance
A number of studies have demonstrated that
carcinogenic circRNAs are involved in TMZ chemoresistance of
gliomas. For instance, the expression of circRNA_ankyrin repeat and
Pleckstrin homology domain 1 (circ_ASAP1) is significantly
upregulated in recurrent GBM tissues and TMZ-resistant cell lines.
The sponge function of circ_ASAP1 directly inhibits the targeted
binding of miR-502-5p to the 3′-UTR of neuroblastoma ras viral
oncogene homologue (NRAS), thereby increasing the expression of
NRAS and promoting the resistance of GBM cells to TMZ (144). Circ_0000936 promotes the TMZ
resistance and proliferation of glioma cells by competitively
binding to miR-1294. By contrast, restoration of miR-1294
eliminates the promoting effect of circ_0000936 on TMZ resistance
in drug-resistant cells (145).
Another carcinogenic circRNA, circ_0076248, induces the expression
of p53 and SIRT1 in tumour cells through the sponge adsorption of
miR-181a. It has been demonstrated to reduce the sensitivity of
gliomas to TMZ (146). Exosomes
derived from TMZ-resistant glioma cells mediate the transfer of
chemoresistance to TMZ-sensitive glioma cells via circ_0042003
(147). Therefore, exosomal
circRNAs have an undeniable role in TMZ resistance. These circRNAs
directly participate in the development of chemoresistance by
targeting multiple mechanisms. Exosomal circRNA_homeodomain
interacting protein kinase 3 (circ_HIPK3) promotes TMZ resistance
in glioma cells by inhibiting apoptosis. Specifically, circ_HIPK3
regulates the miR-421/Zic family member 5 signalling pathway,
resulting in drug resistance (148). In a clinical study from Ding
et al (149), the
expression of exosomal circ_0072083 was increased in the body
fluids of TMZ-resistant patients, which was negatively associated
with the overall survival of the patients. Furthermore,
circ_0072083 promotes AlkB homolog 5 (ALKBH5)-mediated
demethylation by targeting miR-1252-5p, thereby upregulating Nanog
homeobox (NANOG) expression and conferring TMZ chemoresistance in
glioma cells in vitro and xenograft tumour tissues in
vivo (149). CircRNA_nuclear
factor I/X (circ_NFIX) is not only a member of the exosomal circRNA
family but also an inducer of TMZ resistance in GBM cells. A study
by the same authors indicated that exosome-mediated circ_NFIX
significantly reduced the expression of miR-132 in recipient cells
through the sponge adsorption of miR-132, ultimately leading to the
acquisition of drug resistance in TMZ-sensitive glioma cells
(150). In summary, emerging
research has revealed key exosomal circRNAs and the related
pathways that regulate chemoresistance in glioma cells, findings
that may improve the clinical benefits of TMZ treatment for
patients with glioma (Table
XI).
| Table XICircRNAs involved in temozolomide
resistance. |
Table XI
CircRNAs involved in temozolomide
resistance.
| CircRNA | Direction of
differential expression | Genes and
pathways | Mechanisms | (Refs.) |
|---|
| ASAP1 | ↑ |
miR-502-5p/NRAS | - | (143) |
| 0000936 | ↑ | miR-1294 | - | (144) |
| 0076248 | ↑ | miR-181a, p53,
SIRT1 | - | (145) |
| HIPK3 | ↑ | miR-421/ZIC5,
miR-524-5p/KIF2A | Exosome,
apoptosis | (147,151) |
| 0072083 | ↑ |
miR-1252-5p/ALKBH5 | Exosome | (148) |
| NFIX | ↑ | miR-132 | Exosome | (149) |
| 0110757 | ↑ |
miR-1298-5p/ITGA | Apoptosis | (150) |
| 0005198 | ↑ | miR-198/TRIM14 | Apoptosis | (152) |
| CEP128 | ↑ |
miR-145-5p/ABCG2 | Drug transport and
metabolism | (153) |
Evidence indicates that several circRNAs induce TMZ
resistance by regulating apoptosis-related genes. Circ_0110757,
formed by the reverse splicing of exons of myeloid-cell leukaemia
1, has been indicated to be upregulated in TMZ-resistant glioma
cell lines. The underlying drug resistance mechanism is that
circ_0110757 acts as a molecular sponge for miRNA and inhibits
TMZ-induced apoptosis through the miR-1298-5p/ITGA signalling
pathway (151). Similarly,
circ_HIPK3 stimulates the upregulation of kinesin family member 2A
(KIF2A) expression in glioma cells by interacting with miR-524-5p,
thereby inhibiting TMZ sensitivity and apoptosis. Of note, KIF2A
not only acts as a downstream target of miR-524-5p but also
activates the PI3K/AKT signalling pathway in the entire drug
resistance mechanism induced by circ_HIPK3 (152). Circ_0005198 is a ceRNA highly
expressed in glioma tissues, serum samples and TMZ-resistant cells.
It competitively binds to miR-198 to regulate the expression of
tripartite motif-containing 14. Knocking down circ_0005198
expression directly limits the biological behaviours of
drug-resistant gliomas and significantly increases tumour
sensitivity to TMZ (153). In
addition, circRNAs target ABC transporters, generating drug
resistance through the promotion of drug excretion. A study by Hua
et al (154) demonstrated
that silencing circRNA_centrosomal protein 128 (circ_CEP128)
increases the concentration of TMZ in glioma cells by reducing the
expression of ATP-binding cassette superfamily G member 2 (ABCG2),
thereby enhancing cytotoxicity. In summary, circRNAs activate
proteins and pathways in glioma cells. They are expected to become
key targets for the treatment of TMZ resistance.
CircRNAs are involved in resistance to
other drugs
During the treatment of gliomas, the presence of
the BTB seriously hinders the effective delivery of antitumour
drugs to the central nervous system. The following studies use the
BTB as the breakthrough point to explore the key role of multiple
carcinogenic circRNAs in the formation of Dox resistance in-depth.
Gao et al (155) reported
that circRNA_ubiquitin specific protease 1 (circ_USP1) is highly
expressed in glioma-derived microvascular endothelial cells
(GDMECs) in vitro. It acts as a sponge of miRNAs and binds
to miR-194-5p. Therefore, knocking down circ_USP1 directly reduces
the expression of tight junction-related proteins (claudin-5,
occludin and ZO-1) in GDMECs by mediating the miR-194-5p/FLI1 axis,
thereby destroying the integrity of the BTB, increasing the
permeability of Dox and eventually promoting the Dox-induced
apoptosis of glioma cells (155). Another study indicated that the
RNA-binding protein K homology (KH) domain-containing, RNA-binding,
signal transduction-associated protein 3 (KHDRBS3) is upregulated
in GDMECs. KHDRBS3 binds to circRNA_DENN/MADD domain containing 4C
(circ_DENND4C) to form a ribonucleoprotein complex, increasing its
stability. Similarly, circ_DENND4C acts as a molecular sponge of
miR-577, which also affects the permeability of the BTB. Knocking
down circ_DENND4C significantly enhanced the degradation of
downstream target genes (claudin-1, occludin and ZO-1) by miR-577,
promoted the penetration of Dox across the BTB and ultimately led
to the apoptosis of glioma cells (156). CircRNA_001160 is considered an
important regulator of GDMEC growth. It enhances Dox resistance.
The underlying mechanism is as follows: CircRNA_001160 upregulates
the expression of ETS variant transcription factor 1 (ETV1) through
the sponge adsorption of miR-195-5p; overexpressed ETV1 binds to
the promoters of tight junction-related proteins, thereby
increasing the expression of tight junction-related proteins and
ultimately blocks the delivery of Dox to glioma cells and inhibits
Dox-induced apoptosis (157). In
addition, the role of circRNA_104075 in the chemotherapy resistance
of gliomas has been confirmed. Regarding the mechanism,
circRNA_104075 induces glioma cell autophagy by activating the
Wnt/β-catenin and the PI3K/AKT signalling pathways, thereby
reducing the cytotoxicity of matrine (158). In summary, the regulatory
mechanism of circRNAs may provide novel strategies for treating
glioma chemoresistance (Table
XII).
| Table XIICircRNAs involved in resistance to
other drugs. |
Table XII
CircRNAs involved in resistance to
other drugs.
| CircRNA | Direction of
differential expression | Genes and
pathways | Drug | (Refs.) |
|---|
| USP1 | ↑ |
miR-194-5p/FLI1 | Doxorubicin | (154) |
| DENND4C | ↑ |
miR-577/Claudin-5,Occludin, ZO-1 | Doxorubicin | (155) |
| 001160 | ↑ |
miR-195-5p/ETV1 | Doxorubicin | (156) |
| 104075 | ↑ | Wnt/β-catenin,
PI3K/AKT | Matrine | (157) |
6. Future perspectives
Tumour drug resistance is a multifactorial and
complex biological process. To completely eliminate tumour
chemoresistance, it is necessary to reveal its molecular mechanism
from an aetiological perspective to develop new antitumour drugs.
To this end, the present study proposes the research prospects of
tumour drug resistance, ncRNA and drug delivery systems, aimed to
reveal the molecular mechanism of tumour drug resistance from an
epigenetic perspective and clarified the role of ncRNA and
nanocarriers in the treatment of tumours via multiple channels,
targets and levels to confer a novel basis and strategy for the
clinical management of tumour chemoresistance.
Ferroptosis, a newly defined type of programmed
cell death, is closely related to oxidative stress and the
disruption of redox homeostasis is also crucial for the
chemoresistance of tumour cells (159). Wu et al (160) indicated that nanodrugs with
silica particles used as carriers are able to dissolve and release
Fe3O4 particles and glutaminase inhibitors
under acidic conditions, thus consuming glutathione in tumour cells
and reversing the drug resistance of tumour cells to oxaliplatin by
inducing ROS-dependent ferroptosis. Wang et al (161) indicated that ovarian tumour
domain-containing ubiquitin aldehyde-binding protein 1 (OTUB1) and
solute carrier family 7 member 11 (SLC7A11) directly bind to
circ_BGN as RNA binding proteins, and circ_BGN significantly
enhances OTUB1-mediated SLC7A11 deubiquitination, thereby
suppressing ferroptosis. Erastin, a ferroptosis inducer, is able to
reverse the above molecular mechanisms and restore the sensitivity
of drug-resistant breast cancer cells to trastuzumab. The
combination of everolimus and erastin is able to synergistically
reduce the viability of renal cell carcinoma cells and induce
ferroptosis, thereby reversing the drug resistance of tumour cells
to everolimus by inhibiting the activation of the mTOR/4E-binding
protein 1 (4EBP1) signalling pathway (162). Furthermore, the development of
novel drugs based on active ingredients from Traditional Chinese
Medicine is also of great value to solve tumour chemoresistance.
Statistics indicate that >75% of 200 approved drugs for the
treatment of malignant tumours contain active ingredients from
Traditional Chinese Medicine (163). It is well accepted that an
increase in MGMT is able to enhance the chemoresistance of
glioblastoma and tubeimoside-I, a natural extract of Bolbostemma
paniculatum, induces glioblastoma cell apoptosis by reducing
MGMT expression and sensitizes TMZ-resistant glioblastoma cells to
chemotherapy (164). Similarly,
cinobufotalin has been identified as an active antitumour
ingredient that attenuates resistance to cisplatin by inducing
enkurin expression to suppress non-muscle myosin heavy chain
IIA-mediated c-Myc deubiquitination in lung adenocarcinoma
(165).
ncRNAs are able to regulate a variety of biological
processes, representing a class of promising biomarkers for tumour
diagnosis and prognosis. The traditional view is that ncRNA is a
functional RNA molecule that does not encode any protein. However,
with the significant advancements of translatomics and mass
spectrometry in recent years, researchers determined that certain
ncRNAs have a coding function and are able to translate into
functionally independent micropeptides in tumour cells (166). For instance, lncRNA-encoded
peptide-AP attenuates the pentose phosphate pathway by interacting
with transaldolase 1, causing ROS accumulation in colorectal cancer
cells and inducing apoptosis, which eventually sensitizes
colorectal cancer cells to oxaliplatin (167). Aberrant activation of the
Hedgehog (Hh) pathway is able to drive the occurrence and
development of multiple tumour types. Wu et al (168) confirmed that circ_SMO is able to
encode a novel protein, SMO-193a.a., which is critical for Hh
signal transduction and drives malignant progression of
glioblastoma. CircMAPK14-175aa, a novel tumour suppressor protein
encoded by circRNA_mitogen-activated protein kinase 14
(circ_MAPK14), inhibits colorectal cancer progression and
metastasis by reducing the nuclear translocation of MAPK14 via
competitively binding to MAPK kinase 6 to facilitate
ubiquitin-mediated degradation of FOXC1 (169). N6-methyladenosine
(m6A) methylation is the most prevalent
post-transcriptional RNA modification in the transcriptome.
Previous studies have unveiled the critical importance of the
interaction between m6A methylation and ncRNA in tumour
proliferation, metastasis, drug resistance and immunity (170). Pan et al (171) revealed that
methyltransferase-like 3 (METTL3)-dependent m6A
methylation promotes the processing and maturation of miR-181b-5p
by DiGeorge syndrome critical region 8, thus reducing the
expression of neurocalcin δ and inhibiting the 5-FU sensitivity of
colorectal cancer cells. In addition, high expression of METTL3 has
been indicated to promote m6A modification and enhance
lncRNA ABHD11-AS1 transcript stability and expression, resulting in
a poor prognosis of patients with non-small cell lung cancer
(172). In addition,
dysregulation of ncRNAs affects m6A modification. Xie
et al (173) identified
circRNA_ protein tyrosine phosphatase receptor type A (circ_PTPRA)
as a tumour suppressor that abolishes the promoting effect of IGF-2
mRNA-binding protein 1 (IGF2BP1) on breast cancer cell
proliferation, migration and invasion. Mechanistically, circ_PTPRA
interacts with the KH domains of IGF2BP1 and blocks the recognition
of IGF2BP1 by m6A-modified RNAs, resulting in the
downregulation of fatty acid amide hydrolase and MYC mRNA
stability.
With the wide application of nanomedicine in tumour
treatment, researchers have developed various drug delivery systems
to increase the in vivo delivery of therapeutic drugs or
nucleic acids. Of note, these delivery carriers have unique
molecular structures and application advantages, which may enhance
therapeutic effects by improving the retention time of the contents
in vivo and the bioavailability, but they still lack
selectivity and affinity. To address the above shortcomings, the
surface of carrier materials may be modified and the high affinity
between ligands and receptors and the differential expression of
receptors or antigens between tumour cells and normal cells may be
utilized to achieve more accurate targeted therapies. At present,
proteins and peptides are commonly used for active modification of
carrier materials. For instance, lactoferrin (Lf), a
multifunctional glycoprotein, is only expressed in brain
capillaries. Qi et al (174) indicated that a targeted delivery
system modified by Lf is able to markedly improve the ability of
docetaxel liposomes to cross the BBB and enhance brain targeting
effects, demonstrating its potential as a promising
chemotherapeutic drug delivery system for glioblastoma. Angiopep-2
(Ang) is able to specifically bind to receptor-related protein 1
(LRP-1), which is highly expressed on the glioma cell membrane.
Nanoparticles with addition of the Ang polypeptide possess obvious
brain targeting characteristics and do not destroy the integrity of
the BBB. However, the targeting effect of Ang polypeptide
modification is ineffective in normal glial cells with LRP-1
deficiency (175). Mahmoudi
et al (176) attempted to
use arginine-glycine-aspartate (RGD) peptide-modified liposomes as
delivery carriers of curcumin for the targeted treatment of glioma.
They indicated that, compared with free curcumin, RGD-modified
curcumin liposomes significantly improved the tissue targeting,
water solubility and biocompatibility of drugs (176). Borneol, a crystal extracted from
the resin and volatile oil of Dipterocarpus aromatica
Gaertn. f., was able to change the brain neurotransmitter level by
inhibiting P-gp expression and then improve the physiological BBB
permeability. Conjugation of the drug delivery system surface with
borneol not only reduce the cytotoxicity of the drug delivery
system itself but also effectively improved the penetration of 5-FU
into the brain (177).
Furthermore, menthol extracted from the leaves and stems of
peppermint has also been demonstrated to enhance the ability of
drugs to cross various physiological barriers, such as the BBB,
gastrointestinal mucosa barrier and skin. Menthol-modified casein
nanoparticles are able to significantly increase the distribution
of drugs in the brain and their targeting efficiency for glioma
cells is even higher than that of transferrin-modified
nanoparticles (178).
7. Discussion
At present, the most common strategy for the
chemotherapeutic treatment of gliomas is to induce apoptosis by
destroying the DNA structure. The mechanisms of drug resistance
tend to be diverse, including drug excretion, DNA repair,
apoptosis, EMT and the GSC phenotype. In recent years,
RNA-sequencing (RNA-Seq) technology has developed rapidly and
become an indispensable tool for analysing differential genes at
the transcriptome level. Due to its high-throughput, high-accuracy
and high-sensitivity characteristics, RNA-Seq technology has
further uncovered the link between ncRNAs and the epigenetics of a
variety of cancers and opened the door for ncRNA research to move
towards clinical practice.
Numerous studies have demonstrated that ncRNAs are
related to chemotherapy response. In the complex regulatory
network, ncRNAs participate in glioma chemoresistance by activating
multiple signalling pathways. As the first ncRNA to be studied,
miRNAs bind to the 3′-UTR of target genes and exert their
regulatory activity by inhibiting the translation of downstream
proteins. The studies related to the involvement of lncRNAs and
circRNAs in chemoresistance mainly focus on their role as sponge
molecules of miRNAs, regulating drug resistance through the ceRNA
mechanism. By contrast, only a small number of studies focused on
functional mechanisms related to RNA-binding proteins. Furthermore,
2 mechanisms (namely, regulation of transcription and regulation of
translation) have not been reported in relation to drug resistance.
Relevant studies indicate that numerous protein targets are not
able to interact with drug molecules due to the lack of a suitable
protein structure or transcription factors and thus are 'not ready
for medicine' (179).
Encouragingly, compared with protein-coding genes widely used as
targets for glioma treatment, ncRNAs have inherent application
potential. The molecular fragments of ncRNAs account for nearly 98%
of the entire human genome, providing a sufficient target selection
scope for glioma chemotherapy (3). In addition, although almost all
traditional drugs used for glioma chemotherapy face the challenge
of drug resistance, there is currently no report on ncRNA drug
resistance. Furthermore, a series of chemical modifications may be
added to ncRNA drugs so that the circulation half-life of ncRNA
drugs will be longer than that of small-molecule or antibody
drugs.
In summary, ncRNAs have a key regulatory role in
glioma chemoresistance. Targeted correction of the dysregulated
expression of endogenous ncRNAs during the development of drug
resistance using ncRNA antagonists or mimics may be an effective
treatment strategy for the reversal of glioma chemoresistance.
Researchers have developed a variety of miRNA-based therapeutic
tools, such as anti-miRs, antagomiRs, sponge inhibitors, mimics,
agomiRs and pre-miRs. MRX34 is a miR-34a mimic encapsulated by
liposomal nanoparticles. As the first mono nucleic acid drug to
enter clinical trials, MRX34 has achieved satisfactory clinical
results in the treatment of renal cell carcinoma, acral melanoma
and hepatocellular carcinoma. However, the trial was stopped as
patients developed serious adverse reactions (180). Cobomarsen is an anti-miR-155
based on locked nucleic acid modification. It regulates cell
proliferation and differentiation by targeting carcinogenic miR-155
in patients. In phase II clinical trials that enrolled patients
with lymphoma and leukaemia, patients had good tolerance to
systemically administered and intratumourally administered
cobomarsen (181). In terms of
GBM treatment, Regulus Therapeutics Inc. announced a novel
anti-miR-10b-based drug candidate, RGLS5579, in 2019; however, it
is still in the preclinical stage (182). In addition, the expression of
carcinogenic lncRNAs and circRNAs may be inhibited by specific
small interfering RNAs (siRNAs) or short hairpin RNAs, and the
overexpression of tumour suppressor lncRNAs and circRNAs in tumour
cells may be achieved by delivery with specific vectors. However,
as lncRNAs and circRNAs are emerging research targets, the related
mechanisms require to be further explored. Therefore, lncRNA- and
circRNA-based treatments are still in the early stages.
Although ncRNA therapy has broad prospects, it is
still a great challenge to select key target ncRNAs from a large
number of candidates and choose the appropriate delivery system.
ncRNAs are able to simultaneously target multiple functional genes
in a regulatory network. However, while the ncRNAs exert powerful
regulatory functions, off-target effects cannot be avoided. Target
ncRNAs may exert effects on genes other than target genes,
resulting in nonspecific regulation and unexpected effects.
Therefore, target ncRNAs must be selected based on strict
standards, with the comprehensive consideration of the overall
response of ncRNA drugs in vivo. Limited delivery efficiency
is also an important factor hindering ncRNA therapy. To date, a
variety of efficient novel delivery systems have been developed,
e.g. virus-based delivery systems, cationic lipid delivery systems,
polymer delivery systems and antibodies (183). These delivery systems not only
enhance the delivery efficiency of ncRNAs but also improve their
biological stability and pharmacokinetic distribution. However,
while maintaining a high delivery efficiency of therapeutic ncRNAs,
it is necessary to consider the potential damage to human organs
caused by the carrier itself. Viral vectors and cationic
nanoparticles not only have certain toxicity but also induce
immunogenic responses. Furthermore, excessive cationic components
readily cause decomposition of the carrier particles at the
glomerular basement membrane, resulting in renal clearance of
ncRNAs (175). Although
scientific researchers have made great efforts to design a variety
of promising noncationic nucleic acid carriers (octameric
ribonucleoprotein, exosomes, spherical nucleic acids and siRNA
conjugates) to load ncRNAs (183,184), viral vectors and cationic
nanoparticles are still the most commonly used delivery methods in
ncRNA therapy. In other words, the toxicity and immunogenicity of
nucleic acid carrier systems cannot be overcome at present.
Therefore, developing an efficient and nontoxic carrier system is
key to the success of glioma ncRNA therapy. In addition, with the
development of clinical techniques such as liquid biopsy and
secretion examination in recent years, the use of ncRNAs as an
early screening index of drug sensitivity has become another
potential research direction. More interestingly, the combined
administration of ncRNAs and chemotherapeutic drugs for the
sensitization of gliomas also appears to be promising and is worthy
of further translational research and clinical trials. In
conclusion, there is still a long way to go before the
comprehensive application of ncRNA therapy in clinical practice.
However, the development of ncRNA-based therapies may be an
effective supplement to traditional therapy and may overcome the
chemoresistance of tumour cells and thereby improve the prognosis
of patients with glioma (Fig.
3).
 | Figure 3Novel nanoparticles loaded with
targeting nucleic acid sequences for the treatment of glioma drug
resistance. Various new nano-delivery systems (endogenous carriers,
inorganic particles, polymer-based formulas and lipid-based
carriers) may be loaded with a variety of nucleic acid treatment
tools [ASO, LNA, miRNA, siRNA, shRNA and CRISPR-cas9] that may be
delivered into glioma cells to correct the unbalanced expression of
endogenous noncoding RNAs during the formation of drug resistance
by releasing inclusions to cause a reversal of chemoresistance in
gliomas. Due to their accuracy, they have markedly improved the
delivery efficiency of glioma nucleic acid therapy. siRNA, small
interfering RNA; ASO, antisense oligonucleotide; LNA, locked
nucleic acid; miRNA, microRNA; shRNA, short hairpin RNA; sgRNA,
single guide RNA; CRISPR, clustered regularly interspaced short
palindromic repeats; cas9, CRISPR-associated system 9. |
8. Conclusion
In conclusion, chemotherapy is an important
component of comprehensive glioma treatment, but chemoresistance
markedly limits the prognosis of patients with glioma. The present
review focuses on the functions and mechanisms of miRNAs, lncRNAs
and circRNAs in glioma chemoresistance and ultimately illustrates
that ncRNA-mediated precision therapy may be an ideal therapeutic
strategy to overcome chemoresistance in gliomas. Although these
ncRNAs have great potential for monitoring and reversing anticancer
drug resistance, there are numerous ncRNAs that are closely
associated with drug resistance that remain elusive and require to
be identified and further explored.
In addition, more scientific research and clinical
trials are urgently required to develop ncRNA-based therapies to
improve the prognosis of patients with glioma. Thus, ncRNAs will
have an even more important role in future glioma research and
treatment.
Availability of data and materials
Not applicable.
Authors' contributions
ZMZ and YYC were the major contributors to the
writing and revision of the manuscript. XCG and YHZ performed the
literature search, XCW and QYY participated in discussions. TTW, CL
and YX revised this article critically for important intellectual
content. JCD, KBZ and ZWS gave approval for the final version of
the manuscript. All authors have read and approved the final
manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to give special thanks to
Mrs. Ruijuan Ren, the curator of Hebei University library (Baoding,
China), for her guidance and assistance in the literature retrieval
process for this paper.
Funding
The present study was financially supported by grants from the
National Natural Science Foundation of China (grant nos. 81930091,
81773462, 81973077 and 21805197); Graduate Innovative Ability
Training Projects of Hebei Province (grant no. CXZZBS 2022015);
Government-funded Clinical Medicine Outstanding Talent Training
Project (grant no. 360017); Medical Science Foundation of Hebei
University (grant no. 2020A11); and the Foundation Project of the
Affiliated Hospital of Hebei University (grant no. 2019Z003).
References
|
1
|
Francis SS, Ostrom QT, Cote DJ, Smith TR,
Claus E and Barnholtz-Sloan JS: The epidemiology of central nervous
system tumors. Hematol Oncol Clin North Am. 36:23–42. 2022.
View Article : Google Scholar
|
|
2
|
Tomar MS, Kumar A, Srivastava C and
Shrivastava A: Elucidating the mechanisms of Temozolomide
resistance in gliomas and the strategies to overcome the
resistance. Biochim Biophys Acta Rev Cancer. 1876:1886162021.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li D, Zhang Z, Xia C, Niu C and Zhou W:
Non-Coding RNAs in glioma microenvironment and angiogenesis. Front
Mol Neurosci. 14:7636102021. View Article : Google Scholar
|
|
4
|
Shahzad U, Krumholtz S, Rutka JT and Das
S: oncoding RNAs in Glioblastoma: Emerging biological concepts and
potential therapeutic implications. Cancers (Basel). 13:15552021.
View Article : Google Scholar
|
|
5
|
Alomari S, Zhang I, Hernandez A, Kraft CY,
Raj D, Kedda J and Tyler B: Drug Repurposing for glioblastoma and
current advances in drug Delivery-A comprehensive review of the
literature. Biomolecules. 11:18702021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Khunweeraphong N and Kuchler K: Multidrug
resistance in mammals and fungi-from MDR to PDR: A rocky road from
atomic structures to transport mechanisms. Int J Mol Sc.
22:48062021. View Article : Google Scholar
|
|
7
|
Bartolini D, Torquato P, Piroddi M and
Galli F: Targeting glutathione S-transferase P and its interactome
with selenium compounds in cancer therapy. Biophys Acta Gen Subj.
1863:130–143. 2019. View Article : Google Scholar
|
|
8
|
Wiese M and Stefan SM: The A-B-C of
small-molecule ABC transport protein modulators: From inhibition to
activation-a case study of multidrug resistance-associated protein
1 (ABCC1). Med Res Rev. 39:2031–2081. 2019. View Article : Google Scholar
|
|
9
|
Marinho MAG, da Silva Marques M, Lettnin
AP, de Souza Votto AP, de Moraes Vaz Batista Filgueira D and Horn
AP: Interaction between near-infrared radiation and temozolomide in
a glioblastoma multiform cell line: A treatment strategy? Cell Mol
Neurobiol. 41:91–104. 2021. View Article : Google Scholar
|
|
10
|
Chaves C, Declèves X, Taghi M, Menet MC,
Lacombe J, Varlet P, Olaciregui NG, Carcaboso AM and Salvatore C:
Characteri zation of the Blood-brain barrier integrity and the
brain trans cisterninoport of SN-38 in an orthotopic xenograft rat
model of diffuse intrinsic pontine glioma. Pharmaceutics.
12:3992020. View Article : Google Scholar
|
|
11
|
Zhang X, Katsakhyan L, LiVolsi VA, Roth
JJ, Rassekh CH, Bagley SJ and Nasrallah MP: TP53 Mutation and
extraneural metastasis of glioblastoma: Insights from an
institutional experience and comprehensive literature review. Am J
Surg Pathol. 45:1516–1526. 2021. View Article : Google Scholar
|
|
12
|
Se YB, Kim SH, Kim JY, Kim JE, Dho YS, Kim
JW, Kim YH, Woo HG, Kim SH, Kang SH, et al: Underexpression of
HOXA11 is associated with treatment resistance and poor prognosis
in glioblastoma. Cancer Res Trea. 49:387–398. 2017. View Article : Google Scholar
|
|
13
|
Zampieri LX, Sboarina M, Cacace A, Grasso
D, Thabault L, Hamelin L, Vazeille T, Dumon E, Rossignol R,
Frédérick R, et al: Olaparib is a mitochondrial complex i inhibitor
that kills temozolomide-resistant human glioblastoma cells. Int J
Mol Sci. 22:119382021. View Article : Google Scholar
|
|
14
|
Cho YA, Kim D, Lee B, Shim JH and Suh YL:
Incidence, clinicopathologic, and genetic characteristics of
mismatch repair gene-mutated glioblastomas. J Neurooncol.
153:43–53. 2021. View Article : Google Scholar
|
|
15
|
Tomé M, Tchorz J, Gassmann M and Bettler
BJ: Constitutive activation of Notch2 signalling confers
chemoresistance to neural stem cells via transactivation of
fibroblast growth factor receptor-1. Stem Cell Res. 35:1013902019.
View Article : Google Scholar
|
|
16
|
Jia WQ, Zhu JW, Yang CY, Ma J, Pu TY, Han
GQ, Zou MM and Xu RX: Verbascoside inhibits progression of
glioblastoma cells by promoting Let-7g-5p and down-regulating HMGA2
via Wnt/beta-catenin signalling blockade. J Cell Mol Med.
24:2901–2916. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chuang HY, Hsu LY, Pan CM, Pikatan NW,
Yadav VK, Fong I, Chen HC, Yeh CT and Chiu SC: The E3 Ubiquitin
Ligase NEDD4-1 Mediates Temozolomide-resistant glioblastoma through
PTEN attenuation and redox imbalance in Nrf2-HO-1 Axis. Int J Mol
Sci. 22:102472021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Uribe D, Torres Á, Rocha JD, Niechi I,
Oyarzún C, Sobrevia L, Martín RS and Quezada C: Multidrug
resistance in glioblastoma stem-like cells: Role of the hypoxic
microenvironment and adenosine signaling. Mol Aspects Med.
55:140–151. 2017. View Article : Google Scholar
|
|
19
|
Lah TT, Novak M and Breznik B: Brain
malignancies: Glioblastoma and brain metastases. Semin Cancer Biol.
60:262–273. 2020. View Article : Google Scholar
|
|
20
|
Macieja A, Kopa P, Galita G, Pastwa E,
Majsterek I and Poplawski T: Comparison of the effect of three
different topoisomerase II inhibitors combined with cisplatin in
human glioblastoma cells sensitized with double strand break repair
inhibitors. Mol Biol Rep. 46:3625–3636. 2019. View Article : Google Scholar
|
|
21
|
Santangelo R, Rizzarelli E and Copani A:
Role for Metallothionein-3 in the resistance of human U87
glioblastoma cells to temozolomide. ACS Omega. 5:17900–17907. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
De Trizio I, Errede M, d'Amati A, Girolamo
F and Virgintino D: Expression of P-gp in Glioblastoma: What we can
learn from brain development. Curr Pharm Des. 26:1428–1437. 2020.
View Article : Google Scholar
|
|
23
|
Mir H and Singh S: CCL25 Signaling in the
Tumor Microenvironment. Adv Exp Med Biol. 1302:99–111. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jana A, Narula P, Chugh A and Kulshreshtha
R: Efficient delivery of anti-miR-210 using Tachyplesin, a cell
penetrating peptide, for glioblastoma treatment. Int J Pharm.
572:1187892019. View Article : Google Scholar
|
|
25
|
Lu H, Liu P and Pang Q:
MiR-27a-3p/miR-27b-3p promotes neurofibromatosis type 1 via
targeting of NF1. J Mol Neurosci. 71:2353–2363. 2021. View Article : Google Scholar
|
|
26
|
Chen KC, Chen PH, Ho KH, Shih CM, Chou CM,
Cheng CH and Lee CC: IGF-1-enhanced miR-513a-5p signaling
desensitizes glioma cells to temozolomide by targeting the
NEDD4L-inhibited Wnt/β-catenin pathway. PLoS One. 14:e02259132019.
View Article : Google Scholar
|
|
27
|
Ge X, Pan MH, Wang L, Li W, Jiang CF, He
J, Abouzid K, Liu LZ, Shi ZM and Jiang BH: Hypoxia-mediated
mitochondria apoptosis inhibition induces temozolomide treatment
resistance through miR-26a/Bad/Bax axis. Cell Death Dis.
9:11282018. View Article : Google Scholar
|
|
28
|
Peng Y, He X, Chen H, Duan H, Shao B, Yang
F, Li H, Yang P, Zeng Y, Zheng J, et al: Inhibition of
microRNA-299-5p sensitizes glioblastoma cells to temozolomide via
the MAPK/ERK signaling pathway. Biosci Rep. 38:BSR201810512018.
View Article : Google Scholar
|
|
29
|
Zhu D, Tu M, Zeng B, Cai L, Zheng W, Su Z
and Yu Z: Up-regulation of miR-497 confers resistance to
temozolomide in human glioma cells by targeting mTOR/Bcl-2. Cancer
Med. 6:452–462. 2017. View Article : Google Scholar
|
|
30
|
Yin J, Ge X, Shi Z, Yu C, Lu C, Wei Y,
Zeng A, Wang X, Yan W, Zhang J and You Y: Extracellular vesicles
derived from hypoxic glioma stem-like cells confer temozolomide
resistance on glioblastoma by delivering miR-30b-3p. Theranostics.
11:1763–1779. 2021. View Article : Google Scholar
|
|
31
|
Huang W, Zhong Z, Luo C, Xiao Y, Li L,
Zhang X, Yang L, Xiao K, Ning Y, Chen L, et al: The
miR-26a/AP-2α/Nanog signaling axis mediates stem cell self-renewal
and temozolomide resistance in glioma. Theranostics. 9:5497–5516.
2019. View Article : Google Scholar
|
|
32
|
Huang BS, Luo QZ, Han Y, Huang D, Tang QP
and Wu LX: MiR-223/PAX6 axis regulates glioblastoma stem cell
proliferation and the chemo resistance to TMZ via regulating
PI3K/Akt pathway. J Cell Biochem. 118:3452–3461. 2017. View Article : Google Scholar
|
|
33
|
Sun J, Ma Q, Li B, Wang C, Mo L, Zhang X,
Tang F, Wang Q, Yan X, Yao X, et al: RPN2 is targeted by miR-181c
and mediates glioma progression and temozolomide sensitivity via
the wnt/β-catenin signaling pathway. Cell Death Dis. 11:8902020.
View Article : Google Scholar
|
|
34
|
Ho KH, Kuo TC, Lee YT, Chen PH, Shih CM,
Cheng CH, Liu AJ, Lee CC and Chen KC: Xanthohumol regulates
miR-4749-5p-inhibited RFC2 signaling in enhancing temozolomide
cytotoxicity to glioblastoma. Life Sci. 254:1178072020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kong S, Fang Y, Wang B, Cao Y, He R and
Zhao Z: miR-152-5p suppresses glioma progression and tumorigenesis
and potentiates temozolomide sensitivity by targeting FBXL7. J Cell
Mol Med. 24:4569–4579. 2020. View Article : Google Scholar
|
|
36
|
Liu ZQ, Ren JJ, Zhao JL, Zang J, Long QF,
Du JJ, Jia XT, Gu NB, Di ZL, Qian YH and Li SZ: MicroRNA-144
represses gliomas progression and elevates susceptibility to
Temozolomide by targeting CAV2 and FGF7. Sci Rep. 10:41552020.
View Article : Google Scholar :
|
|
37
|
Wang H, Ren S, Xu Y, Miao W, Huang X, Qu
Z, Li J, Liu X and Kong P: MicroRNA-195 reverses the resistance to
temozolomide through targeting cyclin E1 in glioma cells.
Anticancer Drugs. 30:81–88. 2019. View Article : Google Scholar
|
|
38
|
Luo W, Yan D, Song Z, Zhu X, Liu X, Li X
and Zhao S: miR-126-3p sensitizes glioblastoma cells to
temozolomide by inactivating Wnt/β-catenin signaling via targeting
SOX2. Life Sci. 226:98–106. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu CH, Xiao LM, Zeng EM, Chen LK, Zheng
SY, Li DH and Liu Y: MicroRNA-181 inhibits the proliferation, drug
sensitivity and invasion of human glioma cells by targeting
Selenoprotein K (SELK). Am J Transl Res. 11:6632–6640. 2019.
|
|
40
|
Jesionek-Kupnicka D, Braun M,
Trąbska-Kluch B, Czech J, Szybka M, Szymańska B, Kulczycka-Wojdala
D, Bieńkowski M, Kordek R and Zawlik I: MiR-21, miR-34a, miR-125b,
miR-181d and miR-648 levels inversely correlate with MGMT and TP53
expression in primary glioblastoma patients. Arch Med Sci.
15:504–512. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen YY, Ho HL, Lin SC, Ho TD and Hsu CY:
Upregulation of miR-125b, miR-181d, and miR-221 predicts poor
prognosis in MGMT Promoter-unmethylated glioblastoma patients. Am J
Clin Pathol. 149:412–417. 2018. View Article : Google Scholar
|
|
42
|
Nie E, Jin X, Wu W, Yu T, Zhou X, Shi Z,
Zhang J, Liu N and You Y: MiR-198 enhances temozolomide sensitivity
in glioblastoma by targeting MGMT. J Neurooncol. 133:59–68. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang L, Su J, Zhao Z, Hou Y, Yin X, Zheng
N, Zhou X, Yan J, Xia J and Wang Z: MiR-26b reverses temozolomide
resistance via targeting Wee1 in glioma cells. Cell Cycle.
16:1954–1964. 2017. View Article : Google Scholar :
|
|
44
|
Ho KH, Cheng CH, Chou CM, Chen PH, Liu AJ,
Lin CW, Shih CM and Chen KC: miR-140 targeting CTSB signaling
suppresses the mesenchymal transition and enhances temozolomide
cytotoxicity in glioblastoma multiforme. Pharmacol Res.
147:1043902019. View Article : Google Scholar
|
|
45
|
Zhao C, Guo R, Guan F, Ma S, Li M, Wu J,
Liu X, Li H and Yang B: MicroRNA-128-3p enhances the
chemosensitivity of temozolomide in glioblastoma by targeting c-Met
and EMT. Sci Rep. 10:94712020. View Article : Google Scholar
|
|
46
|
Xu JX, Yang Y, Zhang X and Luan XP:
Micro-RNA29b enhances the sensitivity of glioblastoma multiforme
cells to temozolomide by promoting autophagy. Anat Rec (Hoboken).
304:342–352. 2021. View Article : Google Scholar
|
|
47
|
Huang S, Qi P, Zhang T, Li F and He X: The
HIF/1α/miR-224-3p/ATG5 axis affects cell mobility sand
chemoensitivity by regulating hypoxia-induced protective autophagy
in glioblastoma and astrocytoma. Oncol Rep. 41:1759–1768. 2019.
|
|
48
|
Li H, Chen L, Li JJ, Zhou Q, Huang A, Liu
WW, Wang K, Gao L, Qi ST and Lu YT: miR-519a enhances
chemosensitivity and promotes autophagy in glioblastoma by
targeting STAT3/Bcl2 signaling pathway. J Hematol Oncol. 11:702018.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wu Y, Yao Y, Yun Y, Wang M and Zhu R:
MicroRNA-302c enhances the chemosensitivity of human glioma cells
to temozolomide by suppressing P-gp expression. Biosci Rep.
39:BSR201904212019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li Y, Liu Y, Ren J, Deng S, Yi G, Guo M,
Shu S, Zhao L, Peng Y and Qi S: miR-1268a regulates ABCC1
expression to mediate temozolomide resistance in glioblastoma. J
Neurooncol. 138:499–508. 2018. View Article : Google Scholar
|
|
51
|
Zeng A, Yin J, Li Y, Li R, Wang Z, Zhou X,
Jin X, Shen F, Yan W and You Y: miR-129-5p targets Wnt5a to block
PKC/ERK/NF-κB and JNK pathways in glioblastoma. Cell Death Dis.
9:3942018. View Article : Google Scholar
|
|
52
|
Lin XJ, Liu H, Li P, Wang HF, Yang AK, Di
JM, Jiang QW, Yang Y, Huang JR, Yuan ML, et al: miR-936 suppresses
cell proliferation, invasion, and drug resistance of laryngeal
squamous cell carcinoma and targets GPR78. Front Oncol. 10:602020.
View Article : Google Scholar
|
|
53
|
Li FF, Xing C, Wu LL and Xue F: MiR-205
enhances cisplatin sensitivity of glioma cells by targeting E2F1.
Eur Rev Med Pharmacol Sci. 22:299–306. 2018.
|
|
54
|
Alural B, Ayyildiz ZO, Tufekci KU, Genc S
and Genc K: Erythropoietin promotes glioblastoma via miR-451
suppression. Vitam Horm. 105:249–271. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li J, Song J and Guo F: miR-186 reverses
cisplatin resistance and inhibits the formation of the
glioblastoma-initiating cell phenotype by degrading Yin Yang 1 in
glioblastoma. Int J Mol Med. 43:517–524. 2019.
|
|
56
|
Yang L, Li N, Yan Z, Li C and Zhao Z:
MiR-29a-Mediated CD133 expression contributes to cisplatin
resistance in CD133+ glioblastoma stem cells. J Mol
Neurosci. 66:369–377. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang Y, Tu L, Zhou X and Li B:
MicroRNA-22 regulates the proliferation, drug sensitivity and
metastasis of human glioma cells by targeting SNAIL1. J BUON.
25:491–496. 2020.
|
|
58
|
Su PF and Song SQ: Regulation of mTOR by
miR-107 to facilitate glioma cell apoptosis and to enhance
cisplatin sensitivity. Eur Rev Med Pharmacol Sci. 24:114612020.
|
|
59
|
Zhang CG, Yang F, Li YH, Sun Y, Liu XJ and
Wu X: miR-501-3p sensitizes glioma cells to cisplatin by targeting
MYCN. Mol Med Rep. 18:4747–4752. 2018.PubMed/NCBI
|
|
60
|
Yi DY, Su Q, Zhang FC, Fu P, Zhang Q, Cen
YC, Zhao HY and Xiang W: Effect of microRNA-128 on cisplatin
resistance of glioma SHG-44 cells by targeting JAG1. J Cell
Biochem. 119:3162–3173. 2018. View Article : Google Scholar
|
|
61
|
Ducray F and Idbaih A: Neuro-oncology:
Anaplastic oligodendrogliomas-value of early chemotherapy. Nat Rev
Neurol. 9:7–8. 2013. View Article : Google Scholar
|
|
62
|
Yamamuro S, Takahashi M, Satomi K, Sasaki
N, Kobayashi T, Uchida E, Kawauchi D, Nakano T, Fujii T, Narita Y,
et al: Lomustine and nimustine exert efficient antitumor effects
against glioblastoma models with acquired temozolomide resistance.
Cancer Sci. 112:4736–4747. 2021. View Article : Google Scholar
|
|
63
|
Wang GB, Liu JH, Hu J and Xue K: MiR-21
enhanced glioma cells resistance to carmustine via decreasing Spry2
expression. Eur Rev Med Pharmacol Sci. 21:5065–5071. 2017.
|
|
64
|
Xie Q, Yan Y, Huang Z, Zhong X and Huang
L: MicroRNA-221 targeting PI3-K/Akt signaling axis induces cell
proliferation and BCNU resistance in human glioblastoma.
Neuropathology. 34:455–464. 2014. View Article : Google Scholar
|
|
65
|
Rezaei T, Hejazi M, Mansoori B, Mohammadi
A, Amini M, Mosafer J, Rezaei S, Mokhtarzadeh A and Baradaran B:
MicroRNA-181a mediates the chemo-sensitivity of glioblastoma to
carmustine and regulates cell proliferation, migration, and
apoptosis. Eur J Pharmacol. 888:1734832020. View Article : Google Scholar
|
|
66
|
Zhang Y, Yang Y, Yan C, Li J, Zhang P, Liu
R, He J and Chang Y: A review of the ethnopharmacology,
phytochemistry and pharmacology of Cynanchumatratum. J
Ethnopharmacol. 284:1147482022. View Article : Google Scholar
|
|
67
|
Khan I, Mahfooz S and Hatiboglu MA: Herbal
medicine for glioblastoma: Current and future prospects. Med Chem.
16:1022–1043. 2020. View Article : Google Scholar
|
|
68
|
Ni W, Luo L, Zuo P, Li R, Xu X, Wen F and
Hu D: miR-374a inhibitor enhances etoposide-induced cytotoxicity
against glioma cells through upregulation of FOXO1. Oncol Res.
27:703–712. 2019. View Article : Google Scholar
|
|
69
|
Feng Z, Zhang L, Zhou J, Zhou S, Li L, Guo
X, Feng G, Ma Z, Huang W and Huang F: mir-218-2 promotes
glioblastomas growth, invasion and drug resistance by targeting
CDC27. Oncotarget. 8:6304–6318. 2017. View Article : Google Scholar :
|
|
70
|
Li Y, Li W, Yang Y, Lu Y, He C, Hu G, Liu
H, Chen J, He J and Yu H: MicroRNA-21 targets LRRFIP1 and
contributes to VM-26 resistance in glioblastoma multiforme. Brain
Res. 1286:13–18. 2009. View Article : Google Scholar
|
|
71
|
Yin S, Du W, Wang F, Han B, Cui Y, Yang D,
Chen H, Liu D, Liu X, Zhai X and Jiang C: MicroRNA-326 sensitizes
human glioblastoma cells to curcumin via the SHH/GLI1 signaling
pathway. Cancer Biol Ther. 19:260–270. 2018. View Article : Google Scholar
|
|
72
|
Wang Y and Wang L: miR-34a attenuates
glioma cells progression and chemoresistance via targeting PD-L1.
Biotechnol Lett. 39:1485–1492. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Chen PH, Chang CK, Shih CM, Cheng CH, Lin
CW, Lee CC, Liu AJ, Ho KH and Chen KC: The miR-204-3p-targeted
IGFBP2 pathway is involved in xanthohumol-induced glioma cell
apoptotic death. Neuropharmacology. 110:362–375. 2016. View Article : Google Scholar
|
|
74
|
Chakrabarti M and Ray SK: Anti-tumor
activities of luteolin and silibinin in glioblastoma cells:
Overexpression of miR-7-1-3p augmented luteolin and silibinin to
inhibit autophagy and induce apoptosis in glioblastoma in vivo.
Apoptosis. 21:312–328. 2016. View Article : Google Scholar
|
|
75
|
Xiao J, Liu L, Zhong Z, Xiao C and Zhang
J: Mangiferin regulates proliferation and apoptosis in glioma cells
by induction of microRNA-15b and inhibition of MMP-9 expression.
Oncol Rep. 33:2815–2820. 2015. View Article : Google Scholar
|
|
76
|
Bartoušková M and Melichar B: Precision
medicine in medical oncology: Hope, disappointment and reality.
Clin Chem Lab Med. 58:1427–1431. 2020. View Article : Google Scholar
|
|
77
|
Sousa F, Costa-Pereira AI, Cruz A,
Ferreira FJ, Gouveia M, Bessa J, Sarmento B, Travasso RDM and Pinto
IM: Intratumoral VEGF nanotrapper reduces gliobastoma
vascularization and tumor cell mass. J Control Release.
339:381–390. 2021. View Article : Google Scholar
|
|
78
|
Siegal T, Charbit H, Paldor I, Zelikovitch
B, Canello T, Benis A, Wong ML, Morokoff AP, Kaye AH and Lavon I:
Dynamics of circulating hypoxia-mediated miRNAs and tumor response
in patients with high-grade glioma treated with bevacizumab. J
Neurosurg. 125:1008–1015. 2016. View Article : Google Scholar
|
|
79
|
Liu H, Liu Z, Jiang B, Huo L, Liu J and Lu
J: Synthetic miR-145 Mimic enhances the cytotoxic effect of the
antiangiogenic drug sunitinib in glioblastoma. Cell Biochem
Biophys. 72:551–557. 2015. View Article : Google Scholar
|
|
80
|
Cunha PP, Costa PM, Morais CM, Lopes IR,
Cardoso AM, Cardoso AL, Mano M, Jurado AS and Lima MCPD:
High-throughput screening uncovers miRNAs enhancing glioblastoma
cell susceptibility to tyrosine kinase inhibitors. Hum Mol Genet.
26:4375–4387. 2017. View Article : Google Scholar
|
|
81
|
Wang Q, Wang Z, Chu L, Li X, Kan P, Xin X,
Zhu Y and Yang P: The effects and molecular mechanisms of MiR-106a
in multidrug resistance reversal in human glioma U87/DDP and U251/G
Cell Lines. PLoS One. 10:e01254732015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Liu Y, Yang L, Liao F, Wang W and Wang ZF:
MiR-450a-5p strengthens the drug sensitivity of gefitinib in glioma
chemotherapy via regulating autophagy by targeting EGFR. Oncogene.
39:6190–6202. 2020. View Article : Google Scholar :
|
|
83
|
Zhang KL, Zhou X, Han L, Chen LY, Chen LC,
Shi ZD, Yang M, Ren Y, Yang JX, Frank TS, et al: MicroRNA-566
activates EGFR signaling and its inhibition sensitizes glioblastoma
cells to nimotuzumab. Mol Cancer. 13:632014. View Article : Google Scholar
|
|
84
|
Zhang KL, Han L, Chen LY, Shi ZD, Yang M,
Ren Y, Chen LC, Zhang JX, Pu PY and Kang CS: Blockage of a
miR-21/EGFR regulatory feedback loop augments anti-EGFR therapy in
glioblastomas. Cancer Lett. 342:139–149. 2014. View Article : Google Scholar
|
|
85
|
Liao H, Bai Y, Qiu S, Zheng L, Huang L,
Liu T, Wang X, Liu Y, Xu N, Yan X and Guo H: MiR-203 downregulation
is responsible for chemoresistance in human glioblastoma by
promoting epithelial-mesenchymal transition via SNAI2. Oncotarget.
6:8914–8928. 2015. View Article : Google Scholar :
|
|
86
|
Bai Y, Liao H, Liu T, Zeng X, Xiao F, Luo
L, Guo H and Guo L: MiR-296-3p regulates cell growth and multi-drug
resistance of human glioblastoma by targeting ether-à-go-go (EAG1).
Eur J Cancer. 49:710–724. 2013. View Article : Google Scholar
|
|
87
|
Petrelli F, Consoli F, Ghidini A, Perego
G, Luciani A, Mercurio P, Berruti A and Grisanti S: Efficacy of
immune checkpoint inhibitors in rare tumours: A systematic review.
Front Immunol. 12:7207482021. View Article : Google Scholar
|
|
88
|
Yang J, Liu R, Deng Y, Qian J, Lu Z, Wang
Y, Zhang D, Luo F and Chu Y: MiR-15a/16 deficiency enhances
anti-tumor immunity of glioma-infiltrating CD8+ T cells through
targeting mTOR. Int J Cancer. 141:2082–2092. 2017. View Article : Google Scholar
|
|
89
|
Wei J, Nduom EK, Kong LY, Hashimoto Y, Xu
S, Gabrusiewicz K, Ling X, Huang N, Qiao W, Zhou S, et al: MiR-138
exerts anti-glioma efficacy by targeting immune checkpoints. Neuro
Oncol. 18:639–648. 2016. View Article : Google Scholar
|
|
90
|
Li J, Wang F, Wang G, Sun Y, Cai J, Liu X,
Zhang J, Lu X, Li Y, Chen M, et al: Combination epidermal growth
factor receptor variant III peptide-pulsed dendritic cell vaccine
with miR-326 results in enhanced killing on EGFRvIII-positive
cells. Oncotarget. 8:26256–26268. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yao W, Guo G, Zhang Q, Fan L, Wu N and Bo
Y: The application of multiple miRNA response elements enables
oncolytic adenoviruses to possess specificity to glioma cells.
Virology. 458-459:69–82. 2014. View Article : Google Scholar
|
|
92
|
Liu J, Liu L, Chao S, Liu Y, Liu X, Zheng
J, Chen J, Gong W, Teng H, Li Z, et al: The role of
miR-330-3p/PKC-α signaling pathway in low-dose endothelial-monocyte
activating polypeptide-ii increasing the permeability of
blood-tumor barrier. Front Cell Neurosci. 11:3582017. View Article : Google Scholar
|
|
93
|
Giunti L, da Ros M, Vinci S, Gelmini S,
Iorio AL, Buccoliero AM, Cardellicchio S, Castiglione F, Genitori
L, de Martino M, et al: Anti-miR21 oligonucleotide enhances
chemosensitivity of T98G cell line to doxorubicin by inducing
apoptosis. Am J Cancer Res. 5:231–242. 2015.
|
|
94
|
Zhang S, Han L, Wei J, Shi Z, Pu P, Zhang
J, Yuan X and Kang C: Combination treatment with doxorubicin and
microRNA-21 inhibitor synergistically augments anticancer activity
through upregulation of tumor suppressing genes. Int J Oncol.
46:1589–1600. 2015. View Article : Google Scholar
|
|
95
|
Harmalkar M, Upraity S, Kazi S and Shirsat
NV: Tamoxifen-induced cell death of malignant glioma cells is
brought about by oxidative-stress-mediated alterations in the
expression of BCL2 family members and is enhanced on miR-21
inhibition. J Mol Neurosci. 57:197–202. 2015. View Article : Google Scholar
|
|
96
|
Ren Y, Kang CS, Yuan XB, Zhou X, Xu P, Han
L, Wang G, Jia Z, Zhong Y, Yu S, et al: Co-delivery of as-miR-21
and 5-FU by poly(amidoamine) dendrimer attenuates human glioma cell
growth in vitro. J Biomater Sci Polym Ed. 21:303–314. 2010.
View Article : Google Scholar
|
|
97
|
Chen PH, Shih CM, Chang WC, Cheng CH, Lin
CW, Ho KH, Su PC and Chen KC: MicroRNA-302b-inhibited E2F3
transcription factor is related to all trans retinoic acid-induced
glioma cell apoptosis. J Neurochem. 131:731–742. 2014. View Article : Google Scholar
|
|
98
|
Momtazmanesh S and Rezaei N: Long
Non-Coding RNAs in diagnosis, treatment, prognosis, and progression
of glioma: A state-of-the-art review. Front Oncol. 11:7127862021.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Li X, Luo Y, Liu L, Cui S, Chen W, Zeng A,
Shi Y and Luo L: The long noncoding RNA ZFAS1 promotes the
progression of glioma by regulating the miR-150-5p/PLP2 axis. J
Cell Physiol. 235:2937–2946. 2020. View Article : Google Scholar
|
|
100
|
Zhang S, Guo S, Liang C and Lian M: Long
intergenic noncoding RNA 00021 promotes glioblastoma temozolomide
resistance by epigenetically silencing p21 through Notch pathway.
IUBMB Life. 72:1747–1756. 2020. View Article : Google Scholar
|
|
101
|
Chen M, Cheng Y, Yuan Z, Wang F, Yang L
and Zhao H: NCK1-AS1 increases drug resistance of glioma cells to
temozolomide by modulating miR-137/TRIM24. Cancer Biother
Radiopharm. 35:101–108. 2020. View Article : Google Scholar
|
|
102
|
Wang Y, Shan A, Zhou Z, Li W, Xie L, Du B
and Lei B: LncRNA TCONS_00004099-derived microRNA regulates
oncogenesis through PTPRF in gliomas. Ann Transl Med. 9:10232021.
View Article : Google Scholar
|
|
103
|
Zhang L, He A, Chen B, Bi J, Chen J, Guo
D, Qian Y, Wang W, Shi T, Zhao Z, et al: A HOTAIR regulatory
element modulates glioma cell sensitivity to temozolomide through
long-range regulation of multiple target genes. Genome Res.
30:155–163. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Zhang J, Chen G, Gao Y and Liang H:
HOTAIR/miR-125 axis-mediated Hexokinase 2 expression promotes
chemoresistance in human glioblastoma. J Cell Mol Med.
24:5707–5717. 2020. View Article : Google Scholar
|
|
105
|
Su YK, Lin JW, Shih JW, Chuang HY, Fong
IH, Yeh CT and Lin CM: Targeting BC200/miR218-5p signaling axis for
overcoming temozolomide resistance and suppressing glioma stemness.
Cells. 9:18592020. View Article : Google Scholar
|
|
106
|
He X, Sheng J, Yu W, Wang K, Zhu S and Liu
Q: LncRNA MIR155HG promotes temozolomide resistance by activating
the Wnt/β-catenin pathway via binding to PTBP1 in glioma. Cell Mol
Neurobiol. 41:1271–1284. 2021. View Article : Google Scholar
|
|
107
|
Ding J, Zhang L, Chen S, Cao H, Xu C and
Wang X: lncRNA CCAT2 enhanced resistance of glioma cells against
chemodrugs by disturbing the normal function of miR-424. Onco
Targets Ther. 13:1431–1445. 2020. View Article : Google Scholar
|
|
108
|
Li C, Feng S and Chen L: MSC-AS1 knockdown
inhibits cell growth and temozolomide resistance by regulating
miR-373-3p/CPEB4 axis in glioma through PI3K/Akt pathway. Mol Cell
Biochem. 476:699–713. 2021. View Article : Google Scholar
|
|
109
|
Liu B, Zhou J, Wang C, Chi Y, Wei Q, Fu Z,
Lian C, Huang Q, Liao C, Yang Z, et al: LncRNA SOX2OT promotes
temozolomide resistance by elevating SOX2 expression via
ALKBH5-mediated epigenetic regulation in glioblastoma. Cell Death
Dis. 11:3842020. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Peng Y, Wu W, Shang Z, Li W and Chen S:
Inhibition of lncRNA LINC00461/miR-216a/aquaporin 4 pathway
suppresses cell proliferation, migration, invasion, and
chemoresistance in glioma. Open Life Sci. 15:532–543. 2020.
View Article : Google Scholar
|
|
111
|
Li B, Zhao H, Song J, Wang F and Chen M:
LINC00174 down-regulation decreases chemoresistance to temozolomide
in human glioma cells by regulating miR-138-5p/SOX9 axis. Hum Cell.
33:159–174. 2020. View Article : Google Scholar
|
|
112
|
Shangguan W, Lv X and Tian N: FoxD2-AS1 is
a prognostic factor in glioma and promotes temozolomide resistance
in a O6-methylguanine-DNA methyltransferase-dependent
manner. Korean J Physiol Pharmacol. 23:475–482. 2019. View Article : Google Scholar
|
|
113
|
Nie E, Jin X, Miao F, Yu T, Zhi T, Shi Z,
Wang Y, Zhang J, Xie M and You Y: TGF-β1 modulates temozolomide
resistance in glioblastoma via altered microRNA processing and
elevated MGMT. Neuro Oncol. 23:435–446. 2021. View Article : Google Scholar
|
|
114
|
Mazor G, Levin L, Picard D, Ahmadov U,
Carén H, Borkhardt A, Reifenberger G, Leprivier G, Remke M and
Rotblat B: The lncRNA TP73-AS1 is linked to aggressiveness in
glioblastoma and promotes temozolomide resistance in glioblastoma
cancer stem cells. Cell Death Dis. 10:2462019. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Bi CL, Liu JF, Zhang MY, Lan S, Yang ZY
and Fang JS: LncRNA NEAT1 promotes malignant phenotypes and TMZ
resistance in glioblastoma stem cells by regulating
let-7g-5p/MAP3K1 axis. Biosci Rep. 40:BSR202011112020. View Article : Google Scholar
|
|
116
|
Gong R, Li ZQ, Fu K, Ma C, Wang W and Chen
JC: Long noncoding RNA PVT1 promotes stemness and temozolomide
resistance through miR-365/ELF4/SOX2 axis in glioma. Exp Neurobiol.
30:244–255. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Ghafouri-Fard S, Shoorei H, Bahroudi Z,
Abak A and Taheri M: The role of H19 lncRNA in conferring
chemoresistance in cancer cells. Biomed Pharmacother.
138:1114472021. View Article : Google Scholar
|
|
118
|
Lv QL, Wang LC, Li DC, Lin QX, Shen XL,
Liu HY, Li M, Ji YL, Qin CZ and Chen SH: Knockdown lncRNA DLEU1
Inhibits gliomas progression and promotes temozolomide
chemosensitivity by regulating autophagy. Front Pharmacol.
11:5605432020. View Article : Google Scholar
|
|
119
|
Baspinar Y, Elmaci I, Ozpinar A and
Altinoz MA: Long non-coding RNA MALAT1 as a key target in
pathogenesis of glioblastoma. Janus faces or Achilles' heal? Gene.
739:1445182020. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Zhao Z, Liu M, Long W, Yuan J, Li H, Zhang
C, Tang G, Jiang W, Yuan X, Wu M, et al: Knockdown lncRNA CRNDE
enhances temozolomide chemosensitivity by regulating autophagy in
glioblastoma. Cancer Cell Int. 21:4562021. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Wu C, Su J, Long W, Qin C, Wang X, Xiao K,
Li Y, Xiao Q, Wang J, Pan Y and Liu Q: LINC00470 promotes tumour
proliferation and invasion, and attenuates chemosensitivity through
the LINC00470/miR-134/Myc/ABCC1 axis in glioma. J Cell Mol Med.
24:12094–12106. 2020. View Article : Google Scholar
|
|
122
|
Wang W, Han S, Gao W, Feng Y, Li K and Wu
D: Long Noncoding RNA KCNQ1OT1 confers gliomas resistance to
temozolomide and enhances cell growth by retrieving PIM1 from
miR-761. Cell Mol Neurobiol. 42:695–708. 2022. View Article : Google Scholar
|
|
123
|
Ding H, Cui L and Wang C: Long noncoding
RNA LIFR-AS1 suppresses proliferation, migration and invasion and
promotes apoptosis through modulating miR-4262/NF-κB pathway in
glioma. Neurol Res. 43:210–219. 2021. View Article : Google Scholar
|
|
124
|
Qiu G, Tong W, Jiang C, Xie Q, Zou J, Luo
C and Zhao J, Zhang L and Zhao J: Long noncoding RNA WT1-AS inhibit
cell malignancy via miR-494-3p in Glioma. Technol Cancer Res Treat.
19:15330338209197592020. View Article : Google Scholar
|
|
125
|
Jiang C, Shen F, Du J, Fang X, Li X, Su J,
Wang X, Huang X and Liu Z: Upregulation of CASC2 sensitized glioma
to temozolomide cytotoxicity through autophagy inhibition by
sponging miR-193a-5p and regulating mTOR expression. Biomed
Pharmacother. 97:844–850. 2018. View Article : Google Scholar
|
|
126
|
Cao Y, Chai W, Wang Y, Tang D, Shao D,
Song H and Long J: lncRNA TUG1 inhibits the cancer stem cell-like
properties of temozolomide-resistant glioma cells by interacting
with EZH2. Mol Med Rep. 24:5332021. View Article : Google Scholar :
|
|
127
|
Shang C, Tang W, Pan C, Hu X and Hong Y:
Long non-coding RNA TUSC7 inhibits temozolomide resistance by
targeting miR-10a in glioblastoma. Cancer Chemother Pharmaco.
81:671–678. 2018. View Article : Google Scholar
|
|
128
|
Ma Y, Zhou G, Li M, Hu D, Zhang L, Liu P
and Lin K: Long noncoding RNA DANCR mediates cisplatin resistance
in glioma cells via activating AXL/PI3K/Akt/NF-κB signaling
pathway. Neurochem Int. 118:233–241. 2018. View Article : Google Scholar
|
|
129
|
Yang G, Han B and Feng T: ZFAS1 knockdown
inhibits viability and enhances cisplatin cytotoxicity by
up-regulating miR-432-5p in glioma cells. Basic Clin Pharmacol
Toxicol. 125:518–526. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zhou H, Ma Y, Zhong D and Yang L:
Knockdown of lncRNA HOXD-AS1 suppresses proliferation, migration
and invasion and enhances cisplatin sensitivity of glioma cells by
sponging miR-204. Biomed Pharmacother. 112:1086332019. View Article : Google Scholar
|
|
131
|
Zhang B, Fang S, Cheng Y, Zhou C and Deng
F: The long non-coding RNA, urothelial carcinoma associated 1,
promotes cell growth, invasion, migration, and chemo-resistance in
glioma through Wnt/β-catenin signaling pathway. Aging (Albany NY).
11:8239–8253. 2019. View Article : Google Scholar
|
|
132
|
Sun XH, Fan WJ, An ZJ and Sun Y:
Inhibition of long noncoding RNA CRNDE increases chemosensitivity
of medulloblastoma cells by targeting miR-29c-3p. Oncol Res.
28:95–102. 2020. View Article : Google Scholar
|
|
133
|
Huo JF and Chen XB: Long noncoding RNA
growth arrest-specific 5 facilitates glioma cell sensitivity to
cisplatin by suppressing excessive autophagy in an mTOR-dependent
manner. J Cell Biochem. 120:6127–6136. 2019. View Article : Google Scholar
|
|
134
|
Ma B, Gao Z, Lou J, Zhang H, Yuan Z, Wu Q,
Li X and Zhang B: Long non coding RNA MEG3 contributes to cisplatin
induced apoptosis via inhibition of autophagy in human glioma
cells. Mol Med Rep. 16:2946–2952. 2017. View Article : Google Scholar
|
|
135
|
Ma B, Yuan Z, Zhang L, Lv P, Yang T, Gao
J, Pan N, Wu Q, Lou J, Han C and Zhang B: Long non-coding RNA
AC023115.3 suppresses chemoresistance of glioblastoma by reducing
autophagy. Biochim Biophys Acta Mol Cell Res. 1864:1393–1404. 2017.
View Article : Google Scholar
|
|
136
|
Pan JX, Chen TN, Ma K, Wang S, Yang CY and
Cui GY: A negative feedback loop of H19/miR-675/VDR mediates
therapeutic effect of cucurmin in the treatment of glioma. J Cell
Physiol. 235:2171–2182. 2020. View Article : Google Scholar
|
|
137
|
Lv X, Sun J, Hu L, Qian Y, Fan C and Tian
N: Curcumol inhibits malignant biological behaviors and
TMZ-resistance in glioma cells by inhibiting long noncoding RNA
FOXD2-As1-promoted EZH2 activation. Aging (Albany NY).
13:24101–24116. 2021. View Article : Google Scholar
|
|
138
|
Wang C, Chen Y, Wang Y, Liu X, Liu Y, Li
Y, Chen H, Fan C, Wu D and Yang J: Inhibition of COX-2, mPGES-1 and
CYP4A by isoliquiritigenin blocks the angiogenic Akt signaling in
glioma through ceRNA effect of miR-194-5p and lncRNA NEAT1. J Exp
Clin Cancer Res. 38:3712019. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Song T, Yan L, Cai K, Zhao T and Xu M:
Downregulation of long noncoding RNA PVT1 attenuates paclitaxel
resistance in glioma cells. Cancer Biomark. 23:447–453. 2018.
View Article : Google Scholar
|
|
140
|
Cardoso AM, Morais CM, Rebelo O, Tão H,
Barbosa M, Pedroso de Lima MC and Jurado AS: Downregulation of long
non-protein coding RNA MVIH impairs glioblastoma cell proliferation
and invasion through an miR-302a-dependent mechanism. Hum Mol
Genet. 30:46–64. 2021. View Article : Google Scholar
|
|
141
|
Sun W, Zhou H, Han X, Hou L and Xue X:
Circular RNA: A novel type of biomarker for glioma (Review). Mol
Med Rep. 24:6022021. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Zheng K, Xie H, Wu W, Wen X, Zeng Z and
Shi Y: CircRNA PIP5K1A promotes the progression of glioma through
upregulation of the TCF12/PI3K/AKT pathway by sponging miR-515-5p.
Cancer Cell Int. 21:272021. View Article : Google Scholar
|
|
143
|
Liu R, Dai W, Wu A and Li Y: CircCDC45
promotes the malignant progression of glioblastoma by modulating
the miR-485-5p/CSF-1 axis. BMC Cancer. 21:10902021. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Wei Y, Lu C, Zhou P, Zhao L, Lyu X, Yin J,
Shi Z and You Y: EIF4A3-induced circular RNA ASAP1 promotes
tumorigenesis and temozolomide resistance of glioblastoma via
NRAS/MEK1/ERK12-signaling. Neuro Oncol. 23:611–624. 2021.
View Article : Google Scholar
|
|
145
|
Hua L, Huang L, Zhang X and Feng H:
Downregulation of hsa_circ_0000936 sensitizes resistant glioma
cells to temozolomide by sponging miR-1294. J Biosci. 45:1012020.
View Article : Google Scholar
|
|
146
|
Lei B, Huang Y, Zhou Z, Zhao Y, Thapa AJ,
Li W, Cai W and Deng Y: Circular RNA hsa_circ_0076248 promotes
oncogenesis of glioma by sponging miR-181a to modulate SIRT1
expression. J Cell Biochem. 120:6698–6708. 2019. View Article : Google Scholar
|
|
147
|
Si J, Li W, Li X, Cao L, Chen Z and Jiang
Z: Heparanase confers temozolomide resistance by regulation of
exosome secretion and circular RNA composition in glioma. Cancer
Sci. 112:3491–3506. 2021. View Article : Google Scholar
|
|
148
|
Han C, Wang S, Wang H and Zhang J:
Exosomal circ-HIPK3 facilitates tumor progression and temozolomide
resistance by regulating miR-421/ZIC5 axis in glioma. Cancer
Biother Radiopharm. 36:537–548. 2021. View Article : Google Scholar
|
|
149
|
Ding C, Yi X, Chen X, Wu Z, You H, Chen X,
Zhang G, Sun Y, Bu X, Wu X, et al: Warburg effect-promoted exosomal
circ_0072083 releasing up-regulates NANGO expression through
multiple pathways and enhances temozolomide resistance in glioma. J
Exp Clin Cancer Res. 40:1642021. View Article : Google Scholar
|
|
150
|
Ding C, Yi X, Wu X, Bu X, Wang D, Wu Z,
Zhang G, Gu J and Kang D: Exosome-mediated transfer of circRNA
CircNFIX enhances temozolomide resistance in glioma. Cancer Lett.
479:1–12. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Li H, Liu Q, Chen Z, Wu M and Zhang C, Su
J, Li Y and Zhang C: Hsa_circ_0110757 upregulates ITGA1 to
facilitate temozolomide resistance in glioma by suppressing
hsa-miR-1298-5p. Cell Death Dis. 12:2522021. View Article : Google Scholar
|
|
152
|
Yin H and Cui X: Knockdown of circHIPK3
facilitates temozolomide sensitivity in glioma by regulating
cellular behaviors through miR-524-5p/KIF2A-mediated PI3K/AKT
pathway. Cancer Biother Radiopharm. 36:556–567. 2021. View Article : Google Scholar
|
|
153
|
Deng Y, Zhu H, Xiao L, Liu C and Meng X:
Circ_0005198 enhances temozolomide resistance of glioma cells
through miR-198/TRIM14 axis. Aging (Albany NY). 13:2198–2211. 2020.
View Article : Google Scholar
|
|
154
|
Hua L, Huang L, Zhang X, Feng H and Shen
B: Knockdown of circular RNA CEP128 suppresses proliferation and
improves cytotoxic efficacy of temozolomide in glioma cells by
regulating miR-145-5p. Neuroreport. 30:1231–1238. 2019. View Article : Google Scholar
|
|
155
|
Gao Y, Wu P, Ma Y, Xue Y, Liu Y, Zheng J,
Liu X, He Q, Ma J, Liu L and Wang P: Circular RNA USP1 regulates
the permeability of blood-tumour barrier via miR-194-5p/FLI1 axis.
J Cell Mol Med. 24:342–355. 2020. View Article : Google Scholar
|
|
156
|
Wu P, Gao Y, Shen S, Xue Y, Liu X, Ruan X,
Shao L, Liu Y and Wang P: KHDRBS3 regulates the permeability of
blood-tumor barrier via cDENND4C/miR-577 axis. Cell Death Dis.
10:5362019. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Li H, Shen S, Ruan X, Liu X, Zheng J, Liu
Y, Yang C, Wang D, Liu L, Ma J, et al: Biosynthetic CircRNA_001160
induced by PTBP1 regulates the permeability of BTB via the
CircRNA_001160/miR-195-5p/ETV1 axis. Cell Death Dis. 10:9602019.
View Article : Google Scholar :
|
|
158
|
Chi G, Xu D, Zhang B and Yang F: Matrine
induces apoptosis and autophagy of glioma cell line U251 by
regulation of circRNA-104075/BCL-9. Chem Biol Interact.
308:198–205. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Zhang J, Liu Y, Li Q, Xu A, Hu Y and Sun
C: Ferroptosis in hematological malignancies and its potential
network with abnormal tumor metabolism. Biomed Pharmacother.
148:1127472022. View Article : Google Scholar
|
|
160
|
Wu F, Du Y, Yang J, Shao B, Mi Z, Yao Y,
Cui Y, He F, Zhang Y and Yang P: Peroxidase-like Active
Nanomedicine with dual glutathione depletion property to restore
oxaliplatin chemosensitivity and promote programmed cell death. ACS
Nano. 16:3647–3663. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Wang S, Wang Y, Li Q, Li X and Feng X: A
novel circular RNA confers trastuzumab resistance in human
epidermal growth factor receptor 2-positive breast cancer through
regulating ferroptosis. Environ Toxicol. Mar 2–2022. View Article : Google Scholar : Epub ahead of
print.
|
|
162
|
Wang Y, Shi G, Shi S, Jie Y, Rui Y and Yu
R: Everolimus accelerates Erastin and RSL3-induced ferroptosis in
renal cell carcinoma. Gene. 809:1459922022. View Article : Google Scholar
|
|
163
|
Zhang W, Liu C, Li J, Lu Y, Li H, Zhuang
J, Ren X, Wang M and Sun C: Tanshinone IIA: New perspective on the
anti-tumor mechanism of A traditional natural medicine. Am J Chin
Med. 50:209–239. 2022. View Article : Google Scholar
|
|
164
|
Tang Q, Cao H, Tong N, Liu Y, Wang W, Zou
Y, Xu L, Zeng Z, Xu W, Yin Z, et al: Tubeimoside-I sensitizes
temozolomide-resistant glioblastoma cells to chemotherapy by
reducing MGMT expression and suppressing EGFR induced
PI3K/Akt/mTOR/NF-κB-mediated signaling pathway. Phytomedicine.
99:1540162022. View Article : Google Scholar
|
|
165
|
Liu JH, Yang HL, Deng ST, Hu Z, Chen WF,
Yan WW, Hou RT, Li YH, Xian RT, Xie YY, et al: The small molecule
chemical compound cinobufotalin attenuates resistance to DDP by
inducing ENKUR expression to suppress MYH9-mediated c-Myc
deubiquitination in lung adenocarcinoma. Acta Pharmacol Sin. Mar
16–2022. View Article : Google Scholar : Epub ahead of
print.
|
|
166
|
Chen Y, Ho L and Tergaonkar V:
sORF-Encoded MicroPeptides: New players in inflammation,
metabolism, and precision medicine. Cancer Lett. 500:263–270. 2021.
View Article : Google Scholar
|
|
167
|
Wang X, Zhang H, Yin S, Yang Y, Yang H,
Yang J, Zhou Z, Li S, Ying G and Yi Ba: lncRNA-encoded pep-AP
attenuates the pentose phosphate pathway and sensitizes colorectal
cancer cells to Oxaliplatin. EMBO Rep. 23:e531402022. View Article : Google Scholar
|
|
168
|
Wu X, Xiao S, Zhang M, Yang L, Zhong J, Li
B, Li F, Xia X, Li X, Zhou H, et al: A novel protein encoded by
circular SMO RNA is essential for Hedgehog signaling activation and
glioblastoma tumorigenicity. Genome Biol. 22:332021. View Article : Google Scholar
|
|
169
|
Wang L, Zhou J, Zhang C, Chen R, Sun Q,
Yang P, Peng C, Tan Y, Jin C, Wang T, et al: A novel tumour
suppressor protein encoded by circMAPK14 inhibits progression and
metastasis of colorectal cancer by competitively binding to MKK6.
Clin Transl Med. 11:e6132021. View Article : Google Scholar
|
|
170
|
Yao L, Man CF, He R, He L, Huang JB, Xiang
SY, Dai Z, Wang XY and Fan Y: The interaction Between
N6-methyladenosine modification and non-coding RNAs in
gastrointestinal tract cancers. Front Oncol. 11:7841272021.
View Article : Google Scholar
|
|
171
|
Pan S, Deng Y, Fu J, Zhang Y, Zhang Z and
Qin X: N6-methyladenosine upregulates miR-181d-5p in exosomes
derived from cancer-associated fibroblasts to inhibit 5-FU
sensitivity by targeting NCALD in colorectal cancer. Int J Oncol.
60:142022. View Article : Google Scholar :
|
|
172
|
Xue L, Li J, Lin Y, Liu D, Yang Q, Jian J
and Peng J: m6 A transferase METTL3-induced lncRNA
ABHD11-AS1 promotes the Warburg effect of non-small-cell lung
cancer. J Cell Physiol. 236:2649–2658. 2021. View Article : Google Scholar
|
|
173
|
Xie F, Huang C, Liu F, Zhang H, Xiao X,
Sun J, Zhang X and Jiang G: CircPTPRA blocks the recognition of RNA
N6-methyladenosine through interacting with IGF2BP1 to
suppress bladder cancer progression. Mol Cancer. 20:682021.
View Article : Google Scholar
|
|
174
|
Qi N, Zhang S, Zhou X, Duan W, Gao D, Feng
J and Li A: Combined integrin α v β 3 and lactoferrin receptor
targeted docetaxel liposomes enhance the brain targeting effect and
anti-glioma effect. J Nanobiotechnology. 19:4462021. View Article : Google Scholar
|
|
175
|
Jiang T, Qiao Y, Ruan W, Zhang D, Yang Q,
Wang G, Chen Q, Zhu F, Yin J, Zou Y, et al: Cation-Free siRNA
micelles as effective drug delivery platform and potent RNAi
nanomedicines for glioblastoma therapy. Adv Mater. 33:e21047792021.
View Article : Google Scholar
|
|
176
|
Mahmoudi R, Ashraf Mirahmadi-Babaheidri S,
Delaviz H, Hassan Fouani M, Alipour M, Barmak M, Christiansen G and
Bardania H: RGD peptide-mediated liposomal curcumin targeted
delivery to breast cancer cells. J Biomater Appl. 35:743–753. 2021.
View Article : Google Scholar
|
|
177
|
Lin JF, Liu YS, Huang YC, Chi CW, Tsai CC,
Tsai TH and Chen YJ: Borneol and tetrandrine modulate the
blood-brain barrier and blood-tumor barrier to improve the
therapeutic efficacy of 5-fluorouracil in brain metastasis. Integr
Cancer Ther. 21:153473542210776822022. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Gao C, Liang J, Zhu Y, Ling C, Cheng Z, Li
R, Qin J, Lu W and Wang J: Menthol-modified casein nanoparticles
loading 10-hydroxycamptothecin for glioma targeting therapy. Acta
Pharm Sin B. 9:843–857. 2019. View Article : Google Scholar
|
|
179
|
Huang CK, Kafert-Kasting S and Thum T:
Preclinical and clinical development of noncoding RNA therapeutics
for cardiovascular disease. Circ Res. 126:663–678. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Hong DS, Kang YK, Borad M, Sachdev J,
Ejadi S, Lim HY, Brenner AJ, Park K, Lee JL, Kim TY, et al: Phase 1
study of MRX34, a liposomal miR-34a mimic, in patients with
advanced solid tumours. Br J Cancer. 122:1630–1637. 2020.
View Article : Google Scholar
|
|
181
|
Anastasiadou E, Seto AG, Beatty X,
Hermreck M, Gilles ME, Stroopinsky D, Pinter-Brown LC, Pestano L,
Marchese C, Avigan D, et al: Cobomarsen, an oligonucleotide
inhibitor of miR-155, Slows DLBCL tumor cell growth in vitro and in
vivo. Clin Cancer Res. 27:1139–1149. 2021. View Article : Google Scholar
|
|
182
|
Teplyuk NM, Uhlmann EJ, Gabriely G,
Volfovsky N, Wang Y, Teng J, Karmali P, Marcusson E, Peter M, Mohan
A, et al: Therapeutic potential of targeting microRNA-10b in
established intracranial glioblastoma: First steps toward the
clinic. EMBO Mol Med. 8:268–287. 2016. View Article : Google Scholar :
|
|
183
|
Lôbo GCNB, Paiva KLR, Silva ALG, Simões
MM, Radicchi MA and Báo SN: Nanocarriers used in drug delivery to
enhance immune system in cancer therapy. Pharmaceutics.
13:11672021. View Article : Google Scholar
|
|
184
|
Binzel DW, Li X, Burns N, Khan E, Lee WJ,
Chen LC, Ellipilli S, Miles W, Ho YS and Guo P: Thermostability,
tunability, and tenacity of RNA as rubbery anionic polymeric
materials in nanotechnology and nanomedicine-specific cancer
targeting with undetectable toxicity. Chem Rev. 121:7398–7467.
2021. View Article : Google Scholar : PubMed/NCBI
|














