Introduction
Lung cancer, which is usually diagnosed at an
advanced stage, is a major cause of cancer-related mortality, due
to its high incidence, aggressive behavior and lack of major
advancements in treatment strategy (1). Lung cancer was the leading indication
for respiratory surgery (48.9%) in 2011 in Japan (2) and >33,000 patients underwent
surgery for lung cancer at Japanese institutions in the same year
(2). The clinical behavior of
non-small-cell lung cancer (NSCLC) is largely associated with
cancer stage. Curative surgery is only achieved in early-stage
NSCLC (3). Recent advances in
targeted therapy for NSCLC have augmented treatment choices for
lung adenocarcinoma, but chemotherapy remains the therapeutic
mainstay for other types of NSCLC.
Molecular-targeted therapies, such as crizotinib,
which target anaplastic lymphoma kinase fusion gene, and erlotinib
or gefitinib, which target epidermal growth factor receptor gene
mutations, have demonstrated superior single-agent activity in
selected patients compared to standard chemotherapy regimes in lung
cancer treatment (4,5). Recently, a series of new gene fusions
were described in lung cancer patients associated with the kinase
domain of the NTRK1 gene, which encodes the high-affinity nerve
growth factor receptor (TRKA) protein (6). Both the myosin phosphatase
Rho-interacting protein (MPRIP)-NTRK1 and CD74-NTRK1 fusions lead
to constitutive TRKA kinase activity and have been shown to be
oncogenic. In addition, Marchetti et al (7) demonstrated that there are NTRK3
mutations associated with the TRKC receptor and NTRK2 mutations
encoding the NTRK2 (TRKB) receptor in lung large-cell
neuroendocrine carcinoma (LCNEC). Recently, the TRKB signaling
pathway was also reported to be a potential therapeutic target for
lung LCNEC (8).
NTRK1 fusions and NTRK expression in lung cancer may
be promising as a molecular-targeted therapy for future clinical
trials. To expand these findings and determine the prevalence of
these mutations in a cohort of Japanese lung cancer patients, we
investigated the presence of NTRK1 fusions in surgical resection
NSCLC samples (adenocarcinoma, 198 cases; and squamous cell
carcinoma, 70 cases). Surprisingly, using reverse-transcription
polymerase chain reaction (RT-PCR) and direct DNA sequencing, we
were unable to identify fusions in any of those patients. The
immunohistochemical analysis demonstrated that some lung cancer
cases exhibited NTRK2 expression. For this reason, we further
investigated the in vitro antitumor effects of AZD7451 on
the KM12 cell line [colorectal cancer cell line harboring
tropomyosin (TPM)-NTRK1 fusion] (6) and the H460 and H810 cell lines (LCNEC
cell lines exhibiting NTRK2 expression). AZD7451 is a potent
small-molecule pan-TRK inhibitor with a high degree of specificity
and selectivity as compared to other kinases (9). We also performed a PCR using total
mRNA extracted from the three cell lines and examined the total
mRNA expression levels in these cell lines.
Patients and methods
PCR assays for NTRK1 fusions
Total RNA was extracted from lung cancer tissues and
adjacent normal lung tissues using Isogen kit (Nippon Gene, Tokyo,
Japan) according to the manufacturer’s instructions. RNA
concentration was determined using a spectrophotometer (NanoDrop
Technologies, Inc., Rockland, DE, USA) and adjusted to a
concentration of 200 ng/ml. RNA (1 μg) was reverse-transcribed
using Superscript II enzyme (Gibco-BRL, Gaithersburg, MD, USA) with
0.5 μg oligo(dT)12–16 (Amersham Pharmacia Biotech, Inc.,
Piscataway, NJ, USA). DNA concentration was determined by a
NanoDrop spectrophotometer and adjusted at 50 ng/ml. We used 1 ml
of each DNA for the assays. The fusion gene primers were
5′-CTCCCAAGCCTGTGAGCAAGAT-3′ (CD74; forward),
5′-AGGAGATTAGCTCCCTCAAGGAT-3′ (MPRIP; forward) and
5′-GTTGTGGCACTCAGCAAGGAAG-3′ (NTRK1; reverse) and they were
amplified (6). The cycling
conditions were as follows: initial denaturation at 94°C for 1 min,
followed by 40 cycles at 98°C for 10 sec and 68°C for 60 sec,
followed by a final extension at 72°C for 10 min.
Cell lines and cell culture
The KM12 colorectal cancer cell line and the H460
and H810 LCNEC cell lines were purchased from American Type Culture
Collection (Manassas, VA, USA). The KM12 and H460 cell lines were
maintained in RPMI-1640 medium with 10% fetal bovine serum (FBS).
H810 cells were maintained in HITES medium supplemented with 5%
FBS. All the cell lines were grown at 37°C in a humidified
incubator with 5% CO2. The cells were grown to 60–70%
confluence, harvested with trypsin and resuspended to the cell
density required for each assay.
qPCR of NTRK1-3
Total RNA was prepared from the KM12, H460 and H810
cell lines using the miRNeasy mini kit (Qiagen, Hilden, Germany)
and was reverse-transcribed using the High-Capacity cDNA Reverse
Transcription kit (Applied Biosystems, Foster City, CA, USA)
according to the manufacturer’s instructions. Relative qPCR of the
NTRK1-3 tyrosine kinase domains (NTRK1, Hs01021011_m1; NTRK2,
Hs00178811_m1; and NTRK3, 00176797_m1; Applied Biosystems) was used
to evaluate their level of mRNA expression. The relative
quantification method in the 7500 Fast Real-time PCR system
(Applied Biosystems) was used, with actin-β (Applied Biosystems) as
an endogenous internal control. All the samples were evaluated in
triplicate. Approximately 80 lung cancer cases were also evaluated
for NTRK1-3 expression.
Immunohistochemistry
NTRK protein expression was evaluated by
immunohistochemistry using rabbit monoclonal anti-TRKA (Sab76291;
Abcam, Cambridge, MA, USA) and anti-TRKB (80G2; Cell Signaling
Technology, Boston, MA, USA) antibodies. We used a standard
protocol for the immunostaining of samples as previously described
(8). Sections (4 μm) were cut from
paraffin tissue blocks from NSCLC tumors and mounted onto glass
slides. The slides were treated with xylene and then dehydrated in
graded alcohols. For epitope retrieval, the specimens were exposed
to 10 mM citrate buffer (pH 6.0) and heated to ~10 min in
autoclave. Endogenous peroxidase activity was blocked with
H2O2 in methanol. The sections were incubated
with blocking solution (10% Block Ace) and then allowed to react
with anti-TRKA (x100) or anti-TRKB (x80) antibody overnight at 4°C.
After the excess antibody had been washed out with
phosphate-buffered saline (PBS), the samples were incubated with a
peroxidase-conjugated anti-rabbit antibody (rabbit HRP
EnVision™+; Dako, Carpinteria, CA, USA) for 45 min.
After the excess antibody had been washed out with PBS,
3,3-diaminobenzidine substrate (10 min) was used to visualize
antibody binding and the sections were counterstained with
hematoxylin. NTRK staining was evaluated under a light microscope
(Leica DM4000B; Leica microsystems, Wetzler, Germany) with a
magnification of ×400.
Evaluation of growth inhibition by
AZD7451 in vitro
To determine whether NTRK fusions or mutations were
capable of driving cell growth and proliferation in vitro,
we used AZD7451 (AstraZeneca Pharmaceuticals, Waltham, MA, USA) to
inhibit NTRK isoforms in the three cell lines. AZD7451 is a potent
small-molecule pan-TRK inhibitor, which has been reported to have a
similar 2–5 nM in vitro enzyme potency in cell-based assays
(5).
KM12, H460 and H810 cells were plated in 6-well
plates at density of 1.0×106 cell/well at 24 h following
treatment with AZD7451 at variable doses (1, 2.5, 4, 5, 7.5 and 10
nM). Dimethyl sulfoxide (DMSO) was used as a negative control.
After 24 h, the cells were counted using a hemocytometer and the
data are expressed as percent control (% control), which represents
the ratio of cells treated with AZD7451 against control cells
(Fig. 3).
Western blotting and antibodies
Protein extracts from KM12, H460 and H810 cells were
used for western blotting to measure the relative expression of
NTRK protein levels. The cells were cultured at density of
1.0×106 cell/well in 6-well plates as previously
described. KM12 and H460 cells were used after being treated with
AZD7451 concentrations of 0, 1 and 5 nM and incubated for 24 h.
H810 cells were also used after being treated with a concentration
of 0,1 nM as control.
After treatment, the cultured cells were washed,
scraped and resuspended in NuPAGE lithium dodecyl sulfate sample
buffer (Invitrogen Life Technologies, Carlsbad, CA, USA). Equal
amounts of protein were separated by SDS-PAGE and electroblotted
onto polyvinylidene difluoride membranes (Bio-Rad, Hercules, CA,
USA). The membranes were blocked in PBS containing 0.1% Tween-20
and 5% ECL blocking agent (GE Healthcare, Little Chalfont, UK) and
probed with primary antibodies directed against pAkt (Ser473,
1:5,000), pERK1/2 (Thr202/Tyr204, 1:5,000), pTRKA/B (Tyr490/Tyr512,
1:1,000), pTRKA/B (Tyr674/675/Tyr706/707, 1:1,000) and total TRKB
(1:2,000) (Cell Signaling Technology), total TRKA (1:1,000) and
α-tubulin (1:5,000) (Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA). The membranes were washed and incubated with secondary
antibody consisting of horseradish peroxidase-linked anti-rabbit or
anti-mouse IgG (GE Healthcare), at a dilution of 1:5,000.
Results
Absence of NTRK1 fusion genes in Japanese
patients
According to Vaishnavi et al (6), we investigated whether CD74-NTRK1 or
MPRIP-NTRK1 fusions existed in a cohort of 268 primary samples
resected from Japanese NSCLC patients. Within 198 adenocarcinomas,
70 were known driver mutation-negative cases. Using RT-PCR and
direct DNA sequencing, we were unable to identify fusions in any of
these cases.
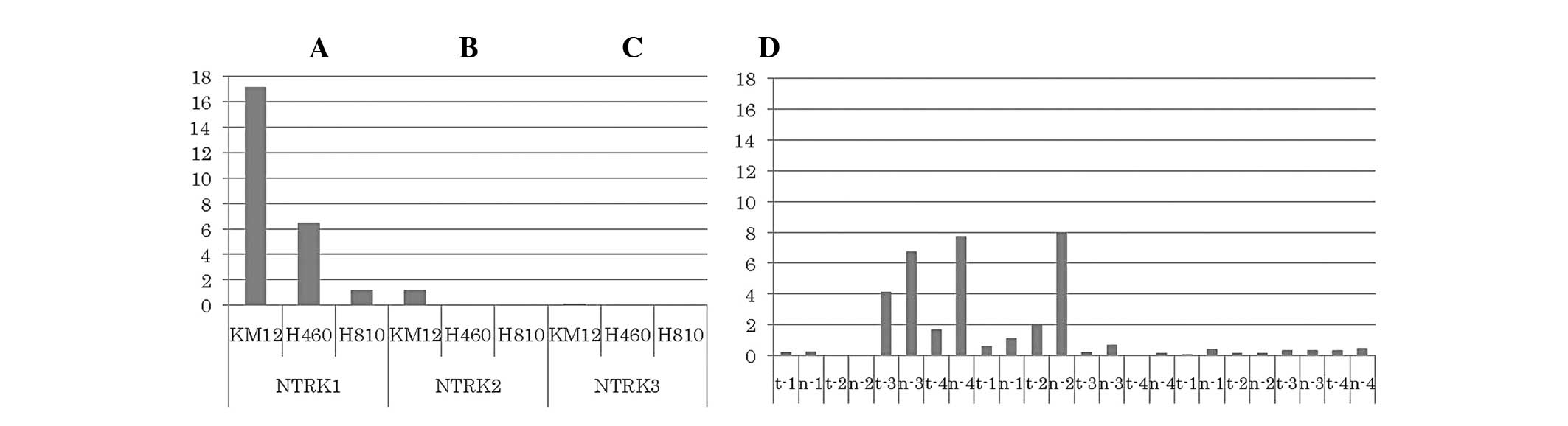
Total mRNA levels of NTRK1-3 tyrosine
kinase domain
Relative qPCR assay was performed to measure the
mRNA levels of the NTRK1-3 tyrosine kinase domain using cDNA
extracted from three cell lines, namely KM12, H460 and H810. Our
results demonstrated that the KM12 cell line exhibited high levels
of NTRK1 mRNA, the H460 cell line exhibited intermediate
expression, while the H810 cell line exhibited relatively low
NTRK1-3 mRNA levels. There was no increase in the NTRK2 mRNA levels
in any of the three cell lines, although KM12 and H460 cells
exhibited a marginal increase in NTKR2 expression (Fig. 1). In addition, relative qPCR was
also performed on 4 primary samples of LCNEC resected at our
institution, but we found no evidence of increased NTRK mRNA levels
in tumor cells compared to those in normal tissue. In
adenocarcinomas, 4/74 exhibited high NTRK1 expression (tumor/normal
ratio, >1), 3/66 exhibited high NTRK2 expression and 1/62
exhibited high NTRK3 expression.
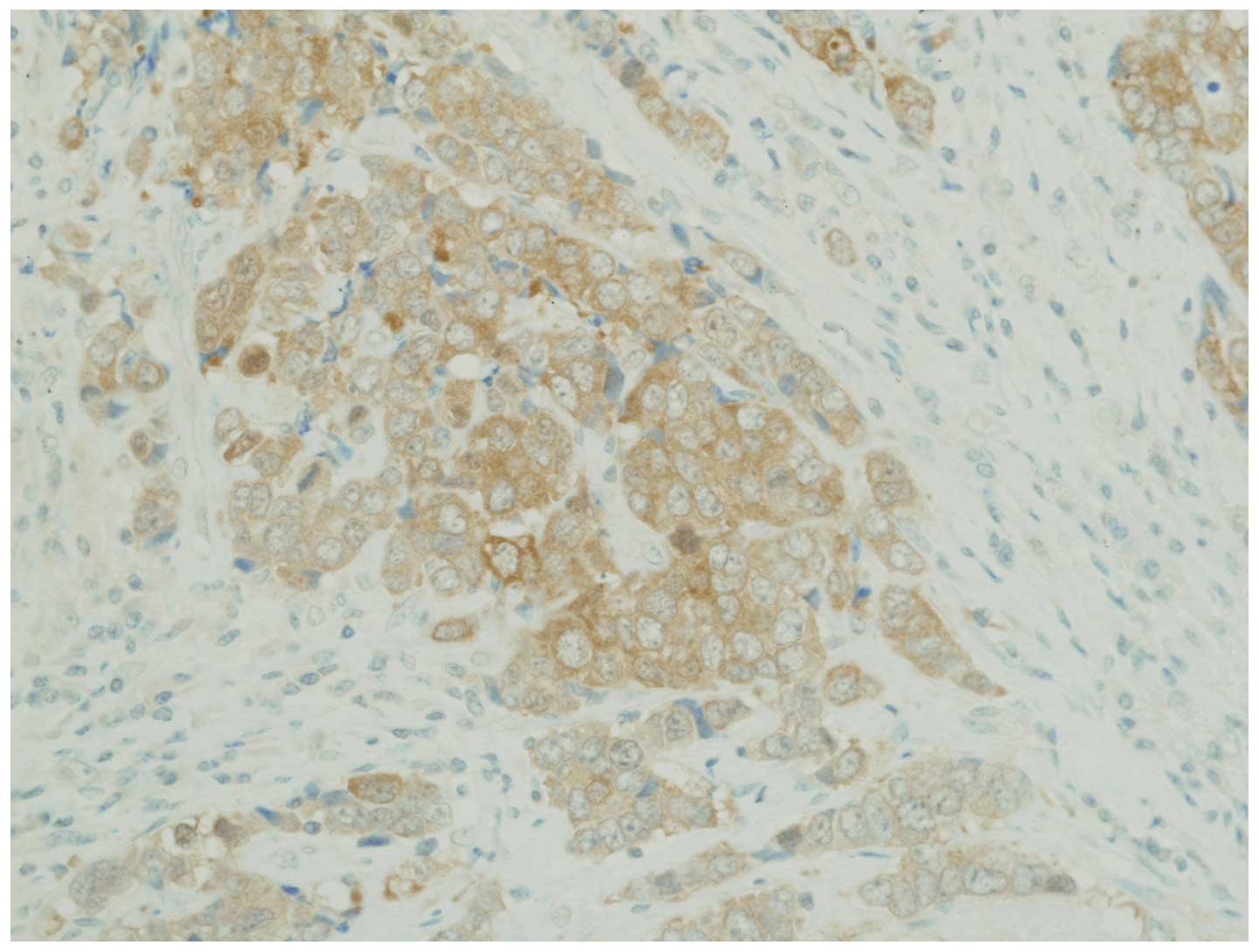
Immunohistochemical analyses for
NTRK
Using the immunohistochemical analysis described
above for NTRK1, we investigated NTRK1 protein expression in 61
lung cancer samples; however, none of the samples exhibited
significant levels of detectable NTRK1 (TRKA) overexpression. In
addition, we investigated NTRK2 (TRKB) expression and found that 7
of the 61 patient samples stained positive for NTRK2. In addition,
1 of the 4 LCNEC samples stained strongly positive for NTKR2
(Fig. 2).
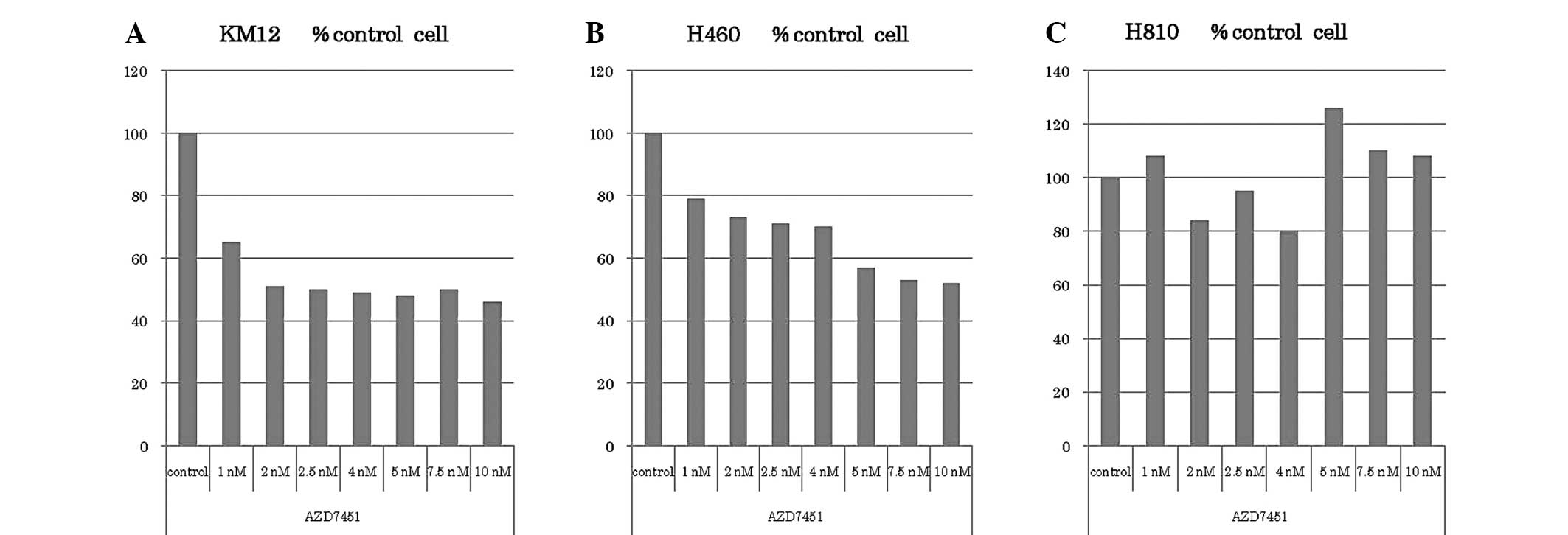
Cytostatic effect of AZD7451 in
vitro
The KM12, H460 and H810 cell lines were treated with
various doses of AZD7451 (0, 1, 2.5, 4.5, 7.5 and 10 nM) and the
results are presented in Fig. 3.
KM12 cell proliferation was potently inhibited at a concentration
of 1 nM, as compared to the DMSO control. Further cell
proliferation was inhibited up to 2 nM, corresponding to a growth
suppression of ~60%; however, there were no further differences in
cell proliferation with increased concentrations of AZD7451. H460
cell proliferation was also inhibited at concentrations ≥1 nM.
Further cell proliferation was inhibited up to 5 nM, also with a
maximal suppression of growth at ~60%. There was no apparent
inhibitory effect on H810 cell proliferation.
Inhibition of phosphorylation of TRKA/B
and downstream signaling in cells treated with AZD7451
In order to determine the phosphorylation of TRKA/B
and the downstream signaling effects on Akt and ERK, we performed
western blotting in all three cell lines treated with AZD7451 at
different doses (KM12 and H460: 0, 1 and 5 nM; and H810: 0 and 5
nM) (Fig. 4). In KM12 cells, the
potent inhibition of pTRKA Tyr490 and pAkt was found to be
concentration-dependent, but inhibition of the expression of pTRKA
Tyr674/675 and pERK was unclear. In H460 cells, total TRKB was
strongly expressed compared to the total TRKA. Therefore, we
considered that H460 cells primarily express NTRK2. In addition,
the expression of pTRKB Tyr706/707 and pERK was inhibited in a
concentration-dependent manner, but inhibition of the expression of
pTRKB Tyr512 and pAkt was unclear in H460 cells. H810 cells
exhibited little to no expression of TRKA/B.
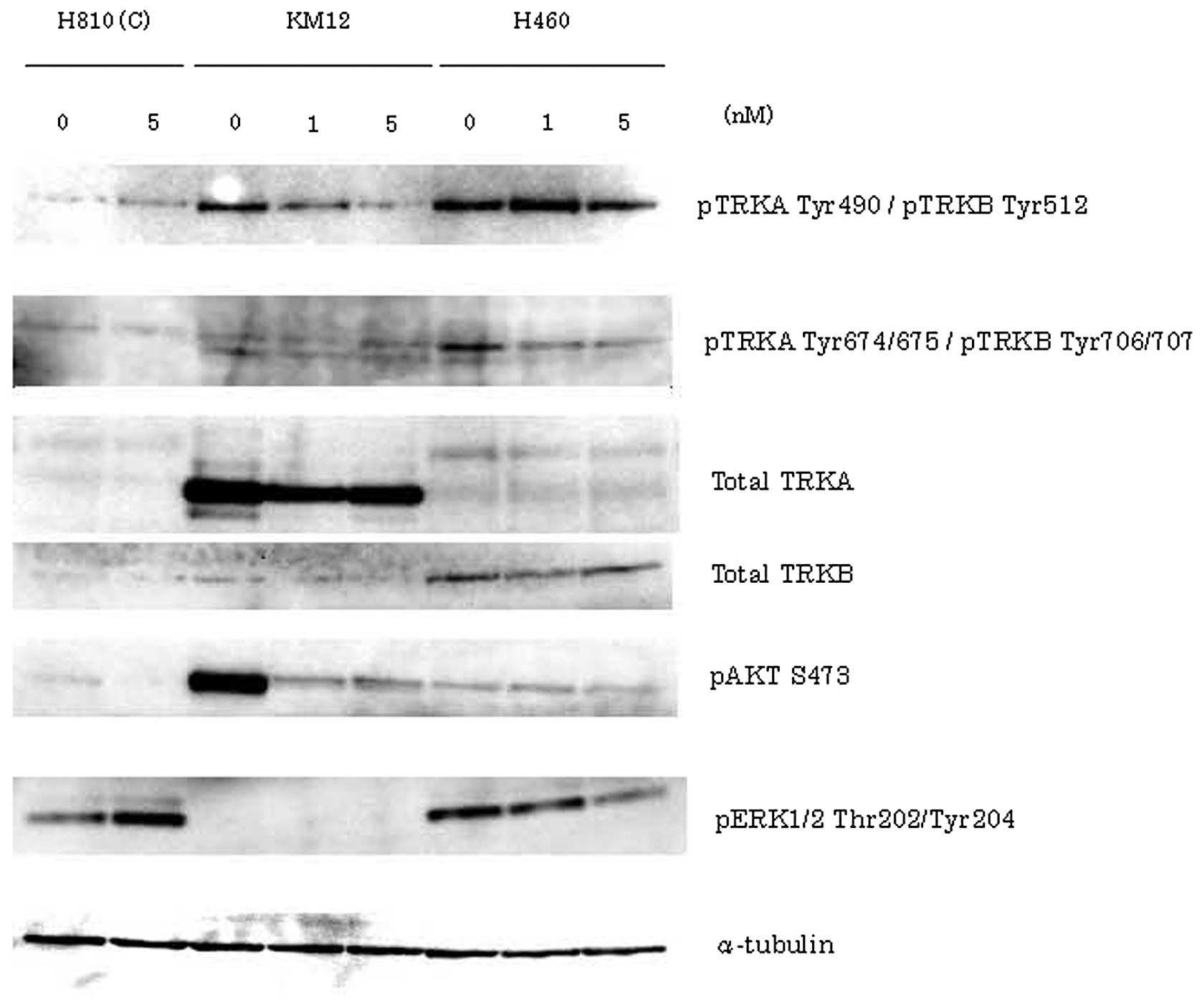 | Figure 4In order to determine TRKA/B
phosphorylation and the downstream signaling effects on Akt and
ERK, western blotting was performed in KM12, H460 and H810 cells.
KM12 and H460 cells were treated with AZD7451 at concentrations of
0, 1 and 5 nM, and incubated for 24 h. H810 cells were also used as
control, after being treated at concentrations of 0 and 5 nM (C).
In KM12 cells, the potent inhibition of pTRKA Tyr490 and pAkt was
found to be concentration-dependent; however, the inhibition of the
expression of pTRKA Tyr674/675 and pERK was unclear. In H460 cells,
total TRKB was strongly expressed compared to total TRKA. In
addition, the expression of pTRKB Tyr706/707 and pERK was inhibited
in a drug concentration-dependent manner, but inhibition of the
expression of pTRKB Tyr512 and pAkt was unclear in H460 cells. H810
cells exhibited little to no expression of TRKA/B. |
Discussion
Vaishnavi et al (6) previously reported the presence of
NTRK1 fusions in 3 of 91 (3.3%) NSCLC cases. In addition, Marchetti
et al (7) also reported the
presence of NTRK2,3 mutations (NTRK2 in 4 and NTRK3 in 5 cases) in
9 of 29 cases (31%) of LCNEC. In this study, we sought to confirm
and extend these analyses and determine the prevalence of these
mutations in a specific cohort of 203 Japanese NSCLC patients.
Understanding the specific molecular segments and proliferation
drivers may improve lung cancer treatment in Japanese patients in
the future. Therefore, we used RT-PCR, direct sequencing,
immunostaining and qPCR in 268 cases of NSCLC specimens from our
institution. Patients with specific mutations (data not shown) or
translocations of the NTRK genes were not identified. There is a
possibility that the frequency of the genetic abnormalities is
extremely low in the Japanese patient population, or that there are
biological differences in the disease subtypes used in this study.
However, to determine the underlying causes of these differences,
more patient samples and additional data associated with
environmental factors are required to further validate and confirm
these results in the future.
NTRK2 (TRKB) has been suggested to play a role in
various neuroendocrine cancers; however, there is no direct
evidence supporting a role for any of the NTRK1-3 receptors in
cancer. The expression of NTKR2 and its ligand brain-derived
neurotrophic factor (BDNF) analyzed by immunohistochemistry for
lung cancer were found to be significantly higher in neuroendocrine
tumors (NET) compared to non-NET (8). In particular, LCNEC, a subtype of
NET, exhibited significantly higher NTRK2 and BDNF compared to
another NET type, small-cell lung cancer; and a significant
correlation between NTRK2 and BDNF expression was noted in LCNEC,
but not in small-cell lung cancer (5). In an in vitro assay, the
addition of exogenous BDNF enhanced the invasion of matrigel by
LCNEC cells, whereas the inhibition of NTRK2 or BDNF suppressed
matrix metalloproteinase activity and tumor cell invasiveness
(8). Exogenous BDNF also increased
anchor-independent colony formation on soft agar gels for LCNEC,
while the inhibition of NTRK2 or BDNF suppressed
anchorage-independent growth. In vivo experiments using
implanted LCNEC cells pretreated with NTRK2-siRNA failed to develop
subcutaneous tumors, while most the control-treated siRNA cells
were capable of forming tumors in nude mice (8,10).
LCNEC is associated with a particularly poor prognosis and an
effective therapeutic strategy has yet to be established. Thus,
BDNF/NTRK2 signaling appears to be involved in malignant
progression, invasiveness and tumorigenicity of LCNEC and may be a
potential target for LCNEC patients without other options for
standard cancer therapy.
We further examined the mRNA levels for the tyrosine
kinase domain of NTRK1-3 and the potential antitumor effects of
AZD7451 in cell lines expressing TPM-NTRK1 fusion and those
exhibiting NTRK2 overexpression. The KM12 and H460 cell lines
exhibited increased expression of NTRK1 mRNA levels, while the H810
cell line did not exhibit significant NTRK1-3 mRNA levels.
Interestingly, none of the three cell lines exhibited increased
mRNA levels of NTRK2,3 based on RT-qPCR data (Fig. 1). Moreover, when the three cell
lines were treated with AZD7451, the proliferation of H460 and KM12
cells was significantly inhibited at concentrations of 1–2 nM, but
the proliferation of H810 cells was not inhibited at significantly
higher doses (Fig. 2). Based on
these results, we hypothesized that the expression of NTRK1 and a
potential proliferation drive may both be associated with NTRK in
the KM12 and H460 cell lines.
To further test this hypothesis, we examined the
phosphorylation of TRKA/B, Akt and ERK1/2. Active signaling was
confirmed by western blotting and subsequent inhibition in the
three cell lines treated with AZD7451 at various doses (KM12 and
H460: 0, 1 and 5 nM; and H810: 0 and 5 nM) (Fig. 4). In KM12, the expression of pTRKA
Tyr490 and pAkt was potently inhibited in a drug
concentration-dependent manner, but inhibition of the expression of
pTRKA Tyr674/675 and pERK was unclear. In H460 cells, total TRKB
was strongly expressed compared to total TRKA. Therefore, we
considered that H460 cells predominantly express NTRK2. In
addition, the expression of pTRKB Tyr706/707 and pERK was inhibited
in a drug concentration-dependent manner, but inhibition of the
expression of pTRKB Tyr512 and pAkt was unclear in H460 cells. H810
cells exhibited low or no expression of TRKA/B.
Based on these results, it seems reasonable to
conclude that, in KM12 cells, following treatment with AZD7451,
phosphorylation of TRKA and cell proliferation were inhibited and
these cells are presumably driven through the NTRK1 fusion in
vitro. In H460 cells, it was confirmed that the expression of
NTRK2 by western blotting and the phosphorylation of TRKB and cell
proliferation were also inhibited by treatment with AZD7451. In
H810 cells, we were unable to detect the expression of any of the
NTRK1-3 isoforms and cell growth was not inhibited by AZD7451.
In conclusion, we were unable to detect NTRK1
fusions or NTRK1-3 mutations in 268 Japanese NSCLC patients at our
institution. One possibility for the discrepancy in these results
may be geographical or ethnic differences associated with these
patient populations; in addition, the incidence of genetic
abnormalities or recurrent polymorphisms in these samples may be
significantly lower in Japan compared to Europe or the United
States. However, we were able to confirm that a potent and
selective inhibitor, such as AZD7451, is able to inhibit the growth
of cells with NTRK1 fusions or NTRK2 expression. Therefore, there
remains a possibility that NTRK mutations or expression may
contribute to the proliferation of lung cancer cells and targeted
therapy with a NTRK inhibitor may be of value in the treatment of
lung cancer, particularly LCNEC, in the future.
Acknowledgements
The authors would like to thank Miss Ito Yamamoto
for her excellent technical assistance. This study was supported by
Grants-in-Aid for Scientific Research, Japan Society for the
Promotion of Science (nos. 26861125, 25293303 and 24592097).
References
|
1
|
Ginsberg RJ, Kris MK and Armstrong JG:
Cancer of the lung. Principles and Practice of Oncology. DeVita VT
Jr, Hellman S and Rosenberg SA: 4th edition. Lippincott;
Philadelphia, PA: pp. 673–682. 1993
|
|
2
|
Amano J, Kuwano H and Yokomise H: Thoracic
and cardiovascular surgery in Japan during 2011: annual report by
the Japanese Association for Thoracic Surgery. Gen Thorac
Cardiovasc Surg. 61:578–607. 2013. View Article : Google Scholar
|
|
3
|
Postmus PE: Chemotherapy for non-small
cell lung cancer: the experience of the Lung Cancer Cooperative
Group of the European Organization for Research and Treatment of
Cancer. Chest. 113(Suppl 1): 28S–31S. 1998. View Article : Google Scholar
|
|
4
|
Shaw AT, Kim DW, Nakagawa K, et al:
Crizotinib versus chemotherapy in advanced ALK-positive lung
cancer. New Engl J Med. 368:2385–2394. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mok TS, Wu YL, Thongprasert S, et al:
Gefitinib or carboplatin-paclitaxel in pulmonary adenocarcinoma.
New Engl J Med. 361:947–957. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vaishnavi A, Capelletti M, Le AT, et al:
Oncogenic and drug-sensitive NTRK1 rearrangements in lung cancer.
Nature Med. 19:1469–1472. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Marchetti A, Felicioni L, Pelosi G, et al:
Frequent mutations in the neurotrophic tyrosine receptor kinase
gene family in large cell neuroendocrine carcinoma of the lung.
Human Mut. 29:609–616. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Odate S, Nakamura K, Onishi H, et al:
TrkB/BDNF signaling pathway is a potential therapeutic target for
pulmonary large cell neuroendocrine carcinoma. Lung Cancer.
79:205–214. 2013. View Article : Google Scholar
|
|
9
|
Ivanov SV, Panaccione A, Brown B, et al:
TrkC signaling is activated in adenoid cystic carcinoma and
requires NT-3 to stimulate invasive behavior. Oncogene.
32:3698–3710. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Odate S, Onishi H, Nakayama K, et al:
Tropomyosin-related kinase B inhibitor has potential for tumor
regression and relapse prevention in pulmonary large cell
neuroendocrine carcinoma. Anticancer Res. 33:3699–3703. 2013.
|