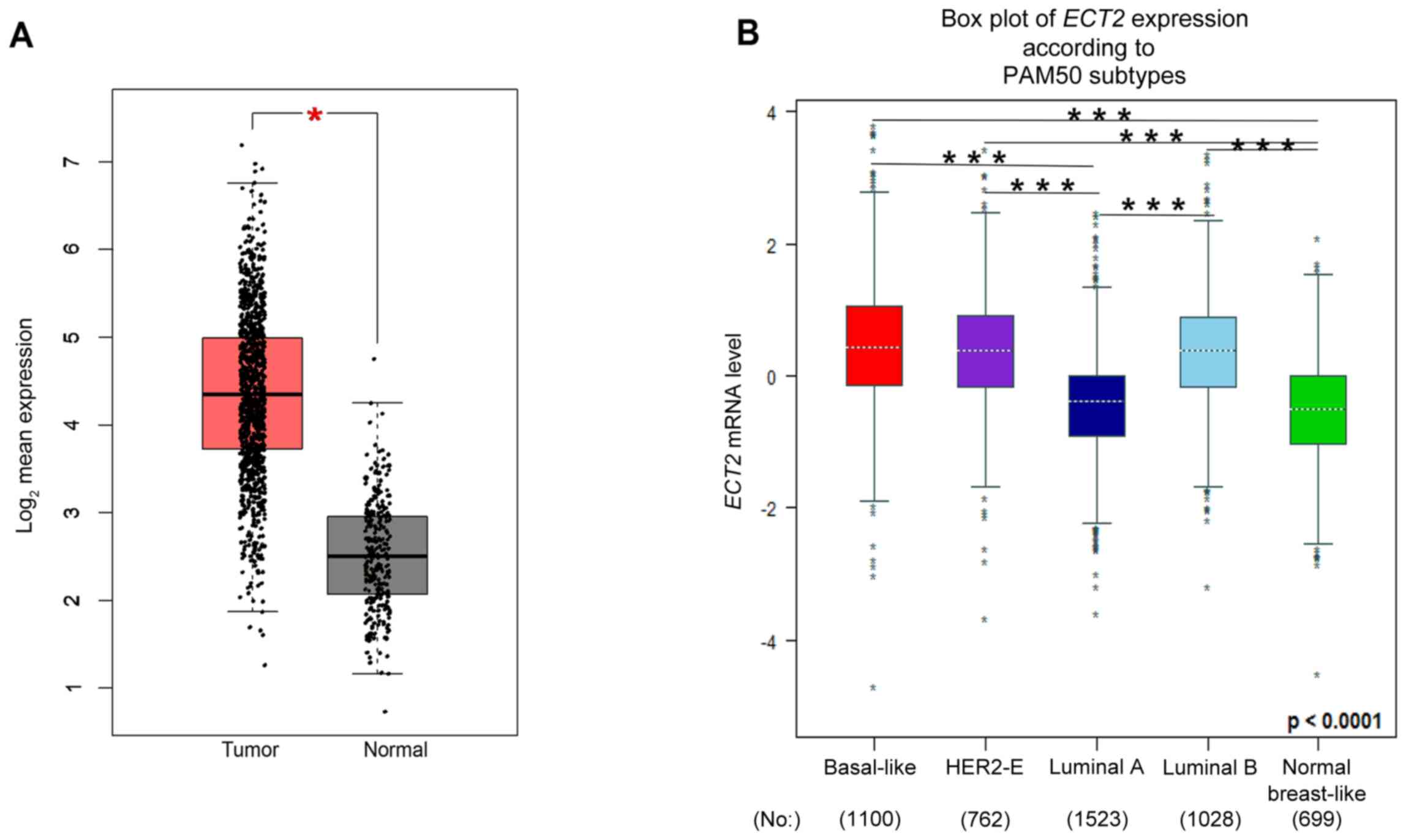
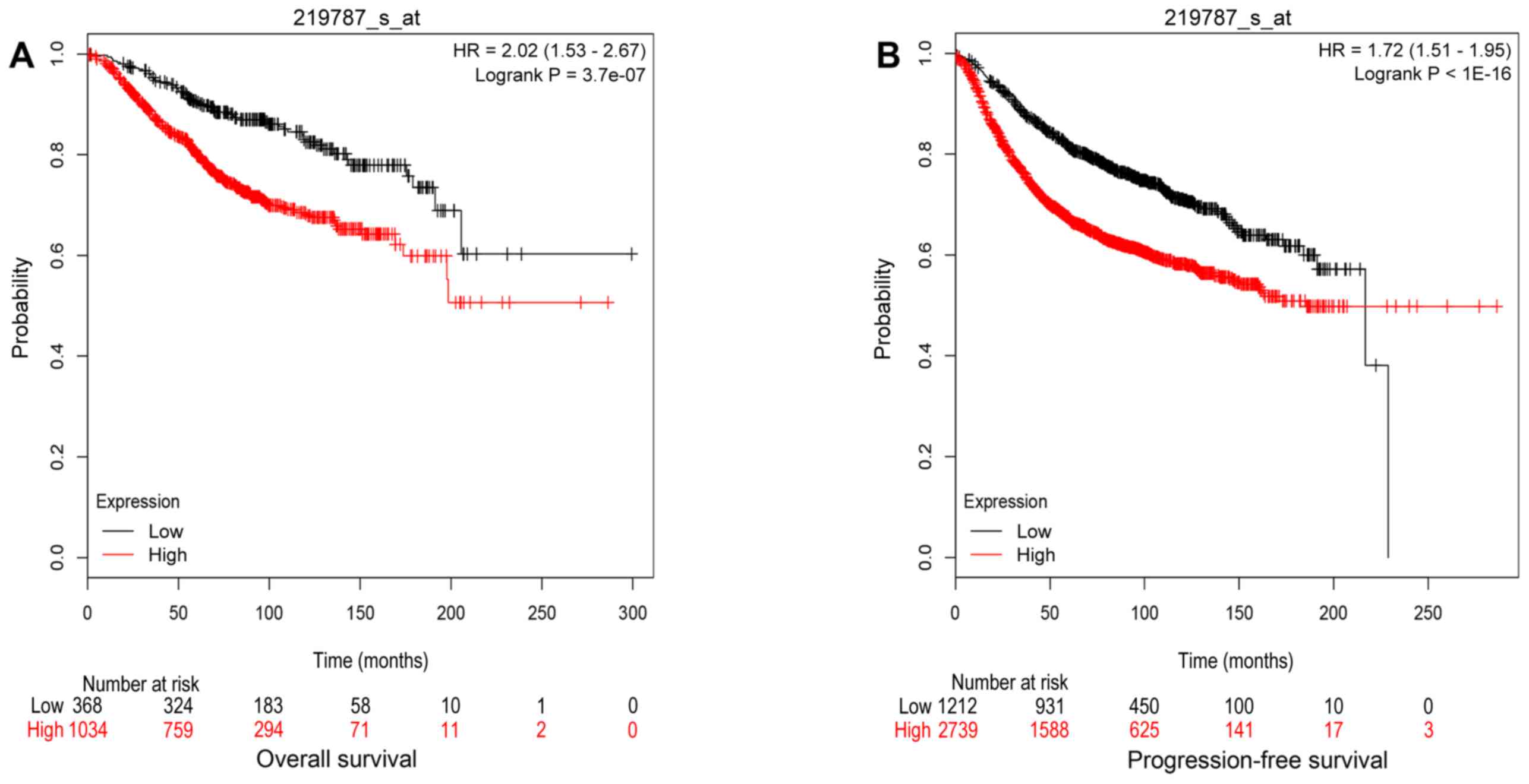
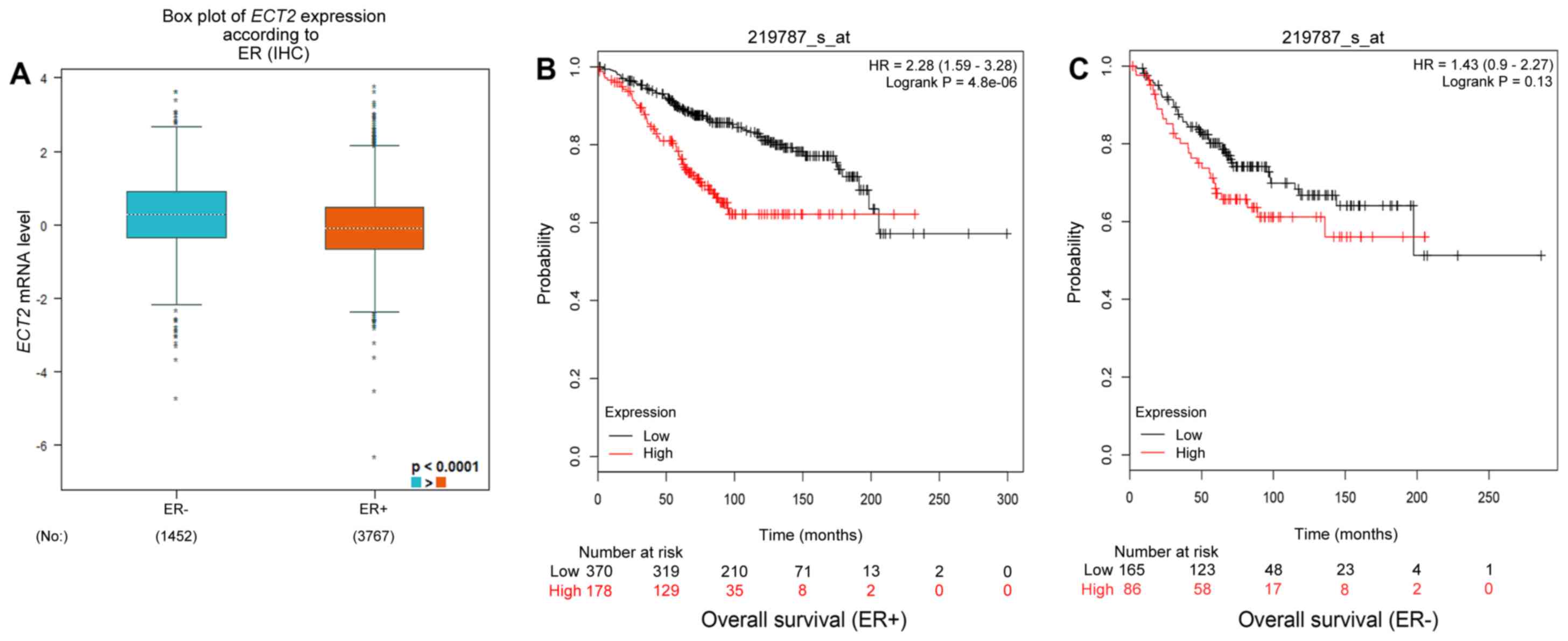
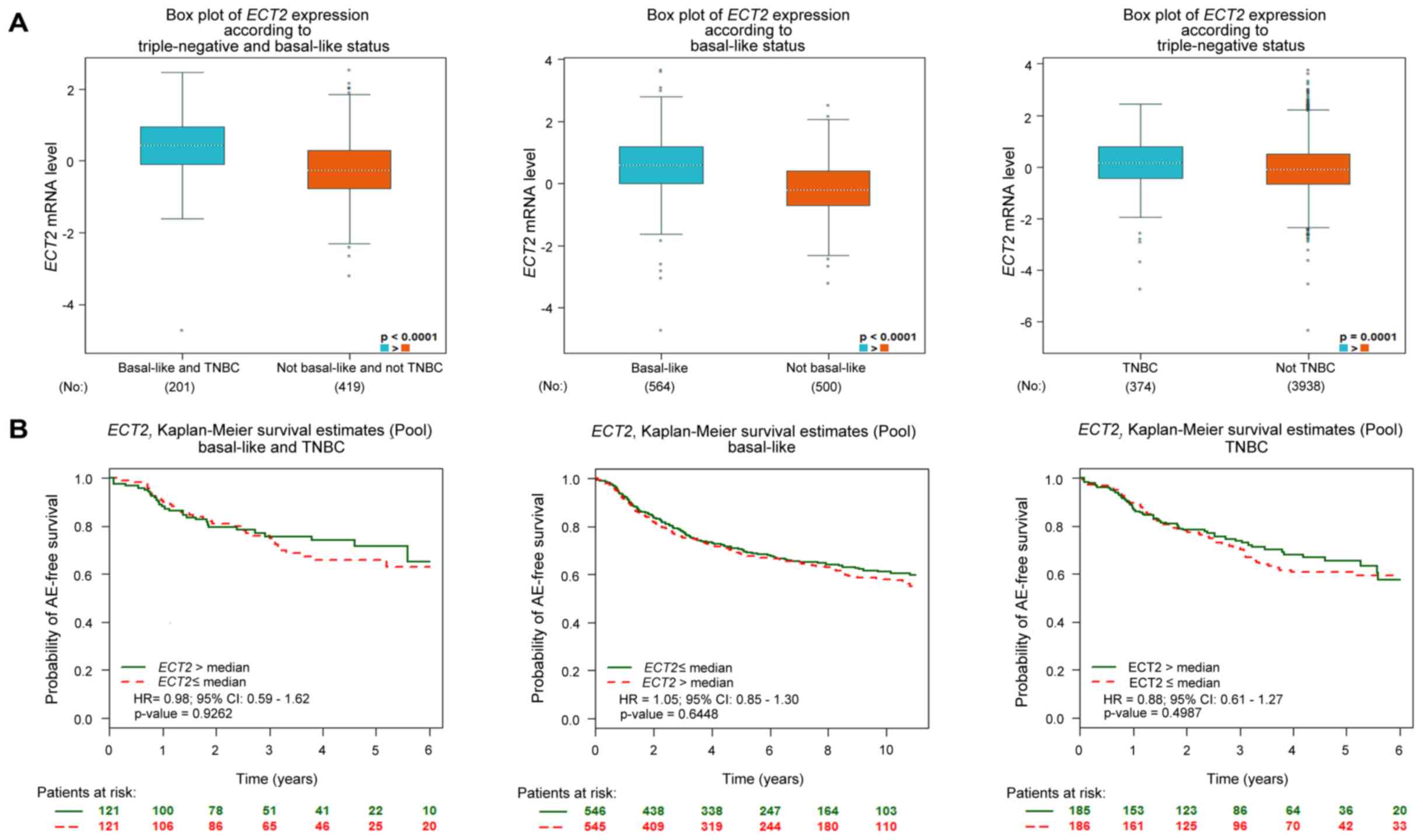
| Biological
process |
|
|
|
| GO:
0051301 | Cell division |
<0.0001a | CDK1, SMC4, CENPW,
CCNB1, MAD2L1, NEK2, BUB1B, NCAPG, KIF18B, FBXO5, KIF11, FAM83D,
CKS2, CCNA2, CDCA8, UBE2C, KIF14, ZWINT, CCNE2, PTTG1, TPX2, CENPF,
OIP5, SMC2, NDC80, KNSTRN, AURKA, CDC20, ERCC6L, NCAPH, CDCA2,
KIF2C, NUF2, GPSM2, CKS1B, CDCA3, SPAG5, BIRC5, MASTL, SGO2, CCNB2,
CENPE, NCAPG2, BUB1, SKA3, CDCA5, KIFC1 |
| GO:
00007062 | Sister chromatid
cohesion |
<0.0001a | MAD2L1, BUB1B,
CDCA8, ZWINT, CENPA, CENPF, NDC80, CDC20, ERCC6L, KIF2C, NUF2,
CENPK, CENPL, BIRC5, CENPU, SGO2, KIF18A, CENPE, CENPM, CENPN,
BUB1, CDCA5 |
| GO:
0000278 | Mitotic cell
cycle |
<0.0001a | CENPW, NEK2, BUB1B,
PBK, KIF18B, KIF11, TPX2, CENPF, NDC80, AURKA, KIF2C, NUF2, KIF15,
MASTL, RAB6C, CENPE, SKA3, CDCA5 |
| GO:
0000070 | Mitotic sister
chromatid segregation |
<0.0001a | SMC4, MAD2L1, NEK2,
NUSAP1, KIF18B, ZWINT, NDC80, KNSTRN, SPAG5, ESPL1, KIFC1 |
| Cellular
component |
|
|
|
| GO:
0005654 | Nucleoplasm |
<0.0001a | CDK1, SMC4, CENPW,
RFC4, ANLN, FANCI, KPNA2, CCNB1, MAD2L1, NEK2, PRC1, UBE2T,
RACGAP1, FBXO5, DTL, RAD51AP1, TOP2A, CCNA2, KIF4A, CDCA8, UBE2C,
KIF23, ZWINT, CCNE2, KIF20A, CENPA, TPX2, CENPF, EZH2, OIP5, SMC2,
NDC80, FOXM1, MCM6, PCNA, ACTL6A, MCM2, ATAD2, AURKA, CDC20, CDCA2,
KIF2C, CENPK, CKS1B, ASF1B, HJURP, PCLAF, CENPL, FEN1, RMI2,
ZNF367, BIRC5, CENPU, DBF4, MCM10, MASTL, SGO2, EXO1, CHEK1, GMNN,
RAD51, NCAPG2, MCM4, CSE1L, DEK, CENPM, TOPBP1, RNASEH2A, CENPN,
BUB1, SNRPD1, CDCA5, NUDCD1 |
| GO:
0000777 | Condensed
chromosome kinetochore |
<0.0001a | CENPW,MAD2L1, NEK2,
BUB1B, ZWINT, NDC80, KNSTRN,ERCC6L, KIF2C, NUF2, CENPK, HJURP,
SPAG5, BIRC5, CENPU, SGO2, CENPM, CENPN, BUB1 |
| GO:
0000775 | Cytosol |
<0.0001a | ECT2, CDK1, SMC4,
RRM2, FANCI, KPNA2, CCNB1, DLGAP5, MAD2L1, NEK2, PRC1, RACGAP1,
BUB1B, NCAPG, KIF18B, FBXO5, DTL, KIF11, CDKN3, CCNA2, DEPDC1B,
KIF4A, CDCA8, UBE2C, KIF23, KIF14, ZWINT, CCNE2, PTTG1, CENPA,
TPX2, CENPF, OIP5, SMC2, NDC80, CKAP2L, HMMR, MCM2, MTHFD2, KNSTRN,
AURKA, CDC20, ERCC6L, NCAPH, CDCA2, KIF2C, NUF2, GPSM2, CENPK,
HJURP, CENPL, CDCA3, RMI2, KIF15, BIRC5, CENPU, SGO2, KIF18A, STIL,
CHEK1, GMNN, ESPL1, TYMS, GMPS, RAD51, CCNB2, RAB6C, CENPE, PDCD10,
CSE1L, CENPM, RNASEH2A, CENPN, BUB1, SNRPD1, PLK4, CDCA5,
NUDCD1 |
| GO:
0030496 | Nucleus |
<0.0001a | PRC1, UBE2T,
RACGAP1, NCAPG, PBK, KIF18B, FBXO5, DTL, CDKN3, RAD51AP1, TOP2A,
CCNA2, CDCA8, KIF23, KIF14, ZWINT, H2AFZ, PTTG1, CENPA, TPX2,
CENPF, EZH2, OIP5, SMC2, NDC80, FOXM1, MCM6, PCNA, ACTL6A, MCM2,
ATAD2, KNSTRN, AURKA, TRIP13, DONSON, NCAPH, NUF2, HJURP, PCLAF,
FEN1, DEPDC1, ZNF367, BIRC5, CENPU, DBF4, MCM10, MASTL, TCF19,
KIF18A, EXO1, CHEK1, GMNN, ESPL1, UHRF1, TYMS, RAD51, CCNB2, RAB6C,
CENPE, MCM4, CSE1L, DEK, TOPBP1, CENPN, SNRPD1, CDCA5, TMPO,
KIFC1 |
| Molecular
function |
|
|
|
| GO:
0008017 | Microtubule
binding |
<0.0001a | PRC1, NUSAP1,
RACGAP1, KIF18B, KIF11, FAM83D, KIF4A, KIF23, KIF14, KIF20A, SPAG5,
KIF15, BIRC5, KIF18A, CENPE, KIFC1 |
| GO:
0005524 | Protein
binding |
<0.0001a | ECT2, CDK1, SMC4,
CENPW, CEP55, MELK, RFC4, RRM2, FANCI, KPNA2, CCNB1, DLGAP5,
MAD2L1, NEK2, PRC1, NUSAP1, UBE2T, RACGAP1, BUB1B, NCAPG, PBK, TTK,
KIF18B, FBXO5, DTL, CDKN3, RAD51AP1, TOP2A, FAM83D, CKS2, CCNA2,
KIF4A, CDCA8, UBE2C, KIF23, KIF14, ZWINT, H2AFZ, CCNE2, PTTG1,
KIF20A, CENPA, TPX2, CENPF, EZH2, OIP5, SMC2, NDC80, FOXM1, MCM6,
HMMR, PCNA, ACTL6A, MCM2, MTFR2, ATAD2, KNSTRN, AURKA, CDC20,
TRIP13, ERCC6L, DONSON, NCAPH, KIF2C, NUF2, GPSM2, CENPK, CKS1B,
ASF1B, HJURP, PCLAF, CENPL, CDCA3, FEN1, SPAG5, DEPDC1, KIF15,
BIRC5, CENPU, DBF4, MCM10, SGO2, TCF19, KIF18A, EXO1, STIL, CHEK1,
GMNN, ESPL1, UHRF1, RAD51, CCNB2, FBXO45, CENPE, PDCD10, MCM4,
CSE1L, SHCBP1, TOPBP1, FAM111B, BUB1, SKA3, SNRPD1, PLK4, CDCA5,
NUDCD1 |
| GO:
0003777 | Microtubule motor
activity |
<0.0001a | KIF18B, KIF11,
KIF4A, KIF23, KIF14, KIF20A, KIF2C, KIF15, CENPE, KIFC1 |
| GO:
0005515 | ATP binding |
<0.0001a | CDK1, SMC4, MELK,
RFC4, NEK2, UBE2T, BUB1B, PBK, TTK, KIF18B, KIF11, TOP2A, KIF4A,
UBE2C, KIF23, KIF14, KIF20A, TPX2, SMC2, MCM6, MCM2, ATAD2, AURKA,
TRIP13, ERCC6L, KIF2C, KIF15, MASTL, KIF18A, CHEK1, GMPS, RAD51,
CENPE, MCM4, BUB1, PLK4, KIFC1 |