Introduction
Cutaneous melanoma is a common malignant skin tumor
(1). The incidence of cutaneous
melanoma is continuing to increase at an annual rate of 2.6%
(2), and the mortality rates are
projected to remain stable through 2019 (3). Currently, surgical or non-surgical
treatment (chemotherapy, cryotherapy) are the gold standard or
second-line options for the treatment of melanoma (4,5), but the
5-year survival rate, recurrence and side effects are major
limitations. Furthermore, the incidence of melanoma and healthcare
costs are expected to increase if suitable preventive measures are
not undertaken.
The hallmark of malignant tumors is abnormal cell
proliferation, and the regulation of the cell cycle is dependent
upon the precise coordination of cyclins and cyclin-dependent
kinases (CDKs) (6,7). CDKs (CDK2, 4, 6 and 1) are key
regulatory enzymes of cell cycle phase transitions (8,9); CDK2,
in particular, plays a crucial role in the regulation of G1-S
transition and modulation of G2 progression (10,11). A
CDK2 activity signature predicts outcome in CDK2-low cancers
(12); hence, developing inhibitors
targeting CDK2 may be a promising approach to cancer therapy
(13). Other studies have indicated
that the role of CDK2 in cancer is debatable, as CDK2 knockdown
failed to inhibit the proliferation of colon cancer cells (14,15), and
cell proliferation may occur in the absence of CDK2 (16,17).
Whether CDK2 controls the cell cycle in cutaneous melanoma remains
unknown.
CRISP/Cas9 technology is a novel tool for gene
editing that has been used for identification of cancer genes
(18,19). Our previous studies demonstrated that
the proliferation of A375 cells was inhibited and apoptosis was
promoted when CDK2 was downregulated by adenovirus-associated virus
(AAV) or lentivirus-mediated shRNA (20,21). In
an attempt to confirm the role of CDK2 in regulating the cell
cycle, apoptosis, or as a therapeutic candidate in human melanoma,
a CDK2-single-guide RNA (sgRNA)-expressing lentivirus was
constructed in the present study, and the effects of
CRISP/Cas9-mediated CDK2 knockout on A375 cell cycle progression
and apoptosis were evaluated, aiming to provide novel insight into
targeted therapy for cutaneous melanoma.
Materials and methods
Cell culture
A375 human melanocytes were obtained from the Cell
Resource Center, Shanghai Institutes for Biological Science,
Chinese Academy of Sciences. A375 cells were cultured in Dulbecco's
modified Eagle's medium (HyClone; GE Healthcare) supplemented with
10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.),
penicillin (100 U/ml) and streptomycin (100 U/ml), in a 5%
CO2 incubator at 37°C.
Lentiviral CDK2-sgRNA vector
construction and packaging
Two sgRNA target sequences [sgCDK2-108
(TGTTCGTACTTACACCCATG) and sgCDK2-110 (CAGAAACAAGTTGACGGGAG)] for
the CDK2 gene (NM_001798) were designed, and a non-silencing sgRNA
sequence (CGCTTCCGCGGCCCGTTCAA) was used as negative control
(sgCDK2-NC). sgRNA constructs were synthesized and cloned into
Lenti-CAS9-sgRNA-EGFP vector with BsmBI sites (GeneChem).
The CDK2-sgRNA plasmids were transfected into 293T cells using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. The packaging
lentivirus particles expressing CDK2-sgRNA were harvested after
transfection for 72 h. Lentivirus was concentrated using the
Centricon Plus-20 centrifugal ultrafiltration device (EMD
Millipore), and stored at −80°C for further use.
Lentivirus infection
A375 cells were seeded in 6-well plates and infected
with sgCDK2 lentivirus or sgCDK2-NC lentivirus at multiplicity of
infection (MOI) of 10, supplemented with polybrene (Santa Cruz
Biotechnology, Inc.) when the confluency of A375 cells reached 30%.
The culture medium was refreshed after 12 h. The lentiviral
infection efficiency was determined through observing GFP
fluorescence expression under a fluorescence microscope (Olympus
Corporation) at 72 h post-infection.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol reagent
(15596026; Thermo Fisher Scientific, Inc.) from A375 cells
following infection by lentivirus for 72 h. cDNA was synthesized by
a high-capacity cDNA reverse transcription kit (4368814; Thermo
Fisher Scientific, Inc.) following the manufacturer's instructions.
The primers used for qPCR were as follows: CDK2, forward
5′-TTCTATGCCTGATTACAAGCC-3′ and reverse 5′-CTGGCTTGGTCACATCCT-3′;
CDK4, forward 5′-CTGGTGACAAGTGGTGGAACAGTC-3′ and reverse
5′-GGTGTAAGTGCCATCTGGTAGCTG-3′; cyclin D1, forward
5′-TACCGCCTCACACGCTTCCTC-3′ and reverse
5′-ACCTCCTCCTCCTCCTCTTCCTC-3′; cyclin A2, forward
5′-AGAAACAGCCAGACATCACTAA-3′ and reverse
5′-TTCAAACTTTGAGGCTAACAGC-3′; and GAPDH, forward
5′-AAGAAGGTGGTGAAGCAGGC-3′ and reverse 5′-TCCACCACCCAGTTGCTGTA-3′.
qPCR was performed using the Quantstudio3 platform (Applied
Biosystems; Thermo Fisher Scientific, Inc.) with FS Universal SYBR
Green Master (4913914001; Roche Diagnostics). The relative
expression of the targets was determined by the 2−∆∆Cq
method in triplicate samples (22).
Western blot analysis
Total proteins were extracted using RIPA lysis
buffer (Beyotime Institute of Biotechnology) supplemented with
protease inhibitors (Roche Diagnostics). Protein concentrations
were determined by a BCA protein assay kit (P0010; Beyotime
Institute of Biotechnology). Proteins (40 µg per lane) were
separated by SDS-PAGE and then transferred onto a polyvinylidene
fluoride membrane. The membrane was blocked in TBST buffer
containing 5% BSA for 2 h at room temperature, and then incubated
with primary antibodies at 4°C overnight. The primary antibodies
used in the study were as follows: CDK2 (1:1,000; ab32147; Abcam),
CDK4 (1:2,000; ab199728; Abcam), cyclin A2 (1:2,000; ab181591;
Abcam), cyclin D1 (1:2,000; ab134175; Abcam), GAPDH (1:1,000,
TA-08; ZsBio). The membrane was then incubated with horseradish
peroxidase (HRP)-goat anti-rabbit IgG (1:5,000; ZB-2301; ZsBio) or
HRP-goat anti-mouse IgG (1:5,000; ZB-2305; ZsBio) after washing
three times with TBST buffer. The blots were developed using the
BeyoECL Plus kit (P0018S; Beyotime Institute of Biotechnology). The
bands of target protein were subjected to densitometric analysis
using Image-Pro Plus 6.0 software (Media Cybernetics).
Cell cycle analysis
A375 cells were seeded in 6-well plates and infected
with sgCDK2-108 or sgCDK2-NC lentivirus. The cells were harvested
by digesting with 0.25% trypsin and centrifuged at 1,000 × g at 4°C
for 5 min, washed twice with PBS and centrifuged at 1,000 × g at
4°C for 5 min to remove the supernatant, then fixed with 70%
ethanol at 4°C overnight. The cells were resuspended in PBS
containing RNase A (100 µg/ml) at 37°C for 30 min, then stained
with propidium iodide (WLA010a; Wanleibio) at 4°C for 30 min. The
stained cells were analyzed using the BD Accuri C6 flow cytometer
(BD Biosciences).
Detection of apoptosis
The A375 cells were handled as described above. The
cells were resuspended in 500 µl binding buffer; subsequently, 5 µl
Annexin V-Light 650 and 10 µl propidium iodide (WLA002c; Wanleibio)
were added and gently mixed. The cells were stained at 37°C for 15
min and the apoptosis was determined using the BD Accuri C6 flow
cytometer (BD Biosciences).
Statistical analysis
All data are expressed as the mean ± standard
deviation of three independent experiments. The data were analyzed
with Student's t-test between two groups, whereas multiple groups
were analyzed with Student Newman Keuls (S-N-K) and one-way ANOVA
using SPSS software, version 13.0 (SPSS, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
CDK2 knockout in A375 cells
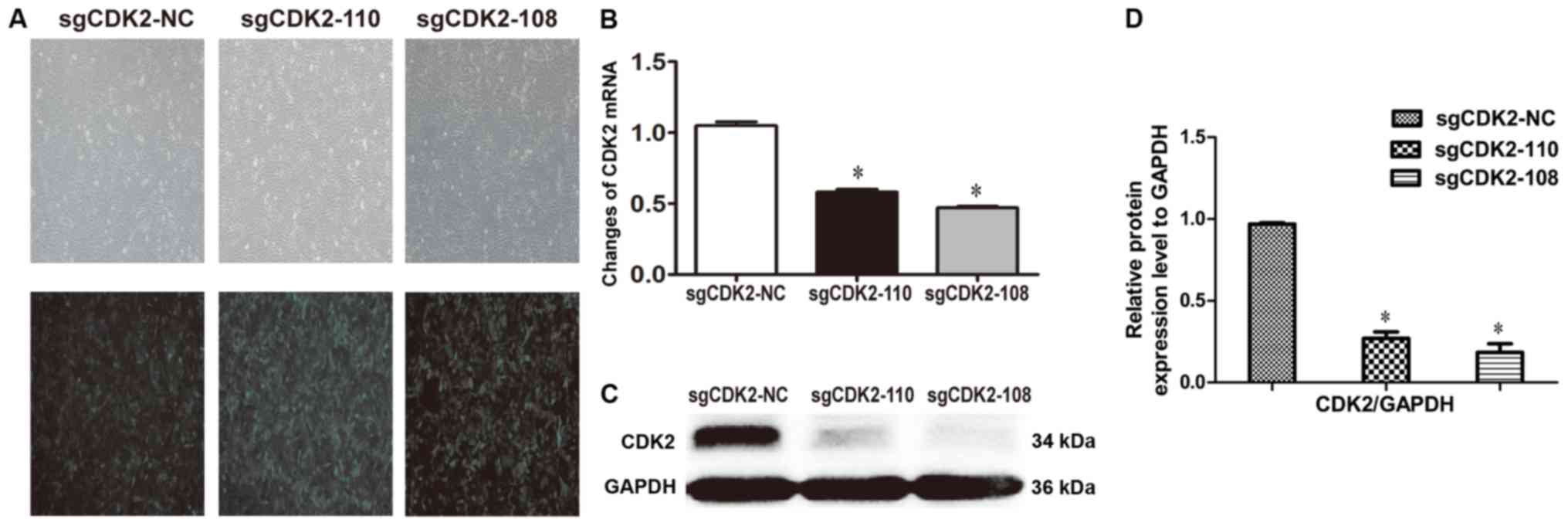
Fluorescence microscopy was used to investigate the
infection efficiency of recombinant lentivirus in A375 cells. The
results demonstrated that >80% of A375 cells expressed GFP among
the sgCDK2-110, sgCDK2-108 and sgCDK2-NC groups (Fig. 1A). The qPCR results revealed that the
mRNA levels of CDK2 were significantly downregulated in A375 cells
infected by lentiviruses sgCDK2-110 and sgCDK2-108 compared with
sgCDK2-NC (P<0.05; Fig. 1B). The
results were further confirmed by western blotting, which
demonstrated that the CDK2 was knocked out in A375 cells infected
by lentiviruses sgCDK2-110 and sgCDK2-108, particularly sgCDK2-108
(P<0.05; Fig. 1C and 1D). These results indicated that CDK2 is
depleted in A375 cells.
CDK2 knockout induces G0/G1 phase
arrest in A375 cells
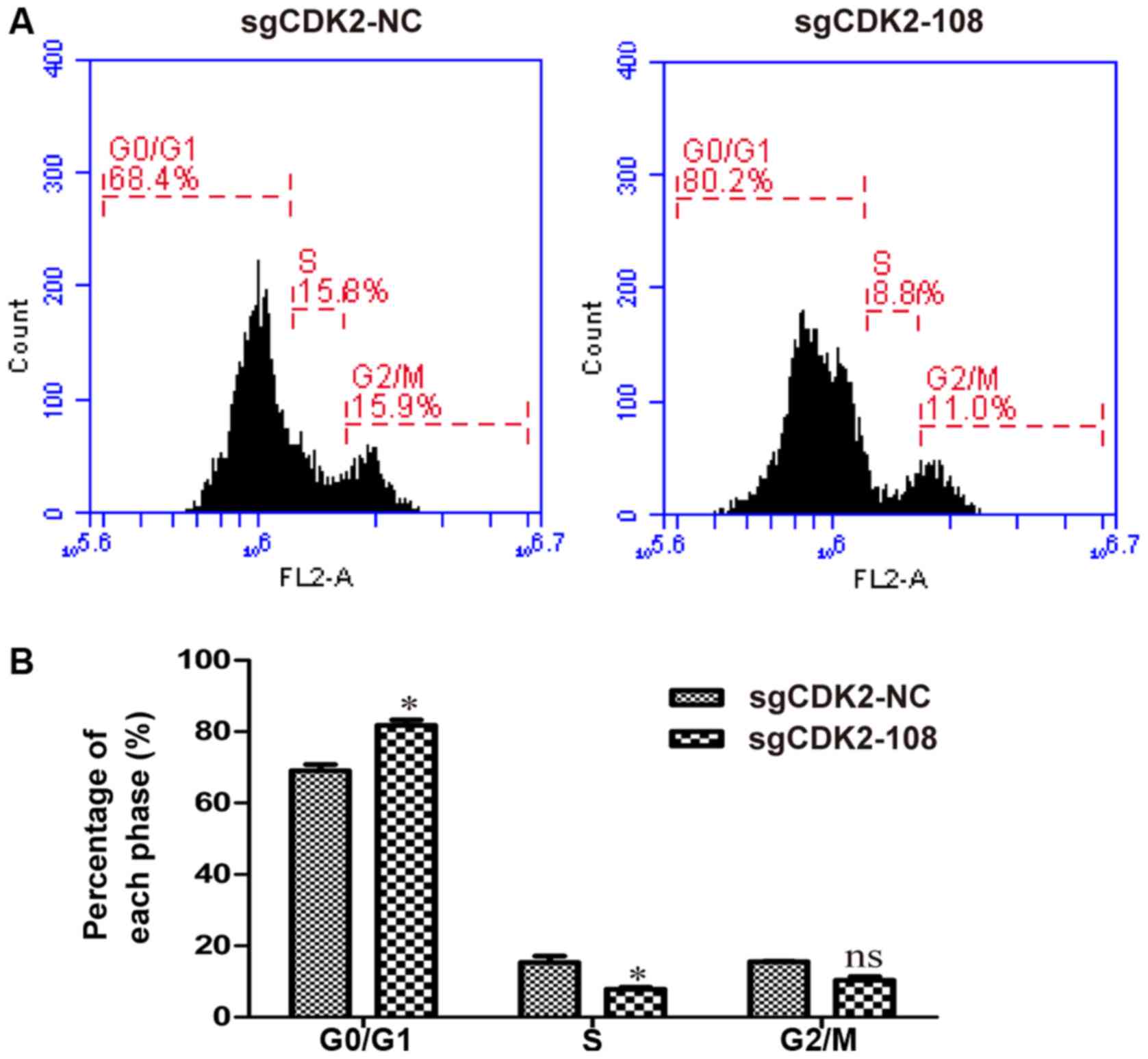
The effect of CDK2 knockout on the cell cycle in
A375 cells was examined by flow cytometric analysis. The lentivirus
sgCDK2-108 was selected for the following experiments. The results
revealed an increased percentage of cells in the G0/G1 phase among
A375 cells infected by lentivirus sgCDK2-108 (Fig. 2A). The percentage of cells in the
G0/G1 phase among lentivirus sgCDK2-108-infected A375 cells was
significantly higher (81.78%) compared with lentivirus sgCDK2-NC
(69.06%); however, the percentage of S phase cells in the
sgCDK2-108 group was significantly reduced (7.85%) compared with
the sgCDK2-NC group (15.38%) (P<0.05; Fig. 2B). These results indicated that CDK2
knockout induces G0/G1 phase arrest in A375 cells.
CDK2 knockout induces early apoptosis
in A375 cells
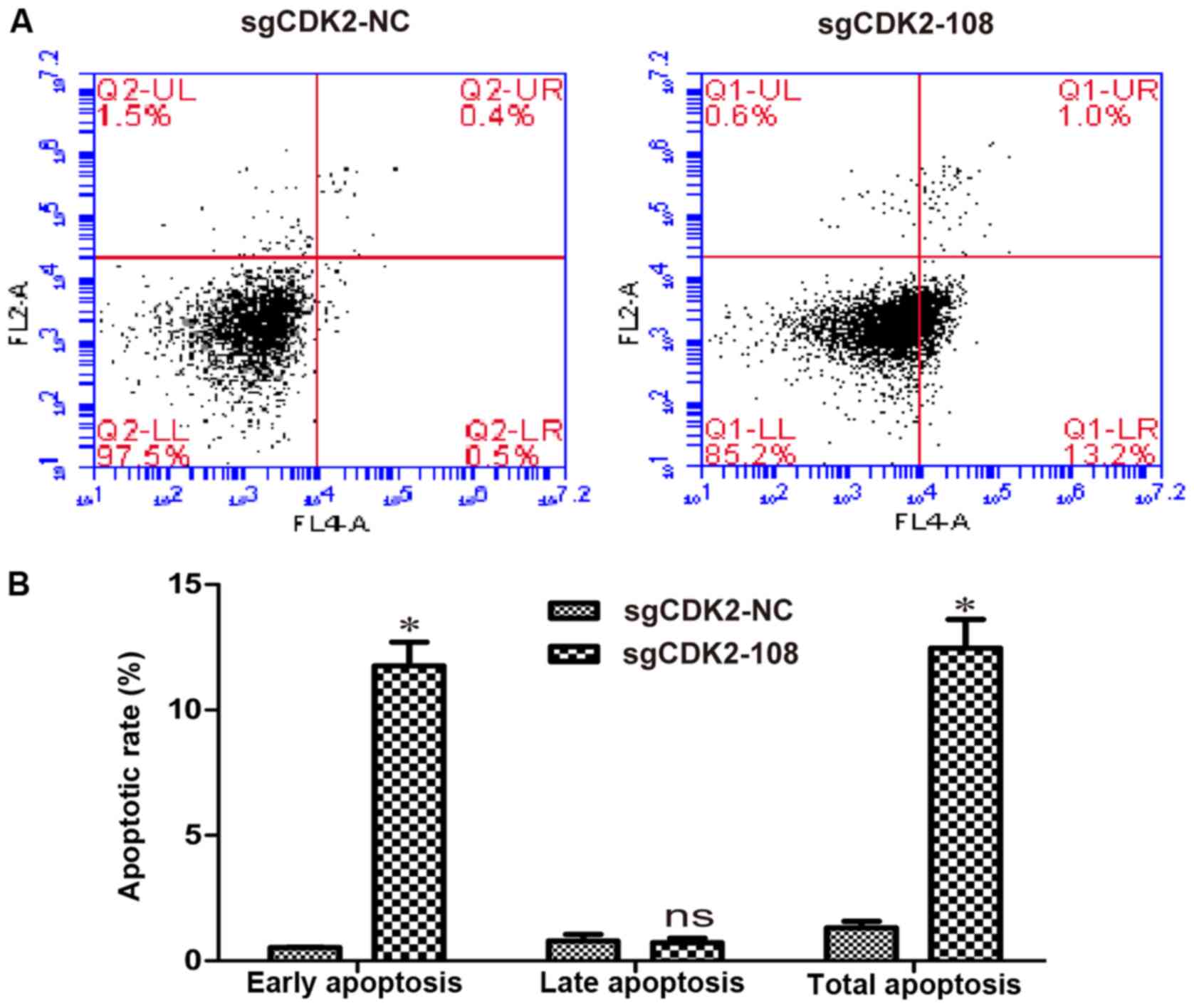
Apoptosis was then analyzed in A375 cells infected
with lentiviruses sgCDK2-108 or sgCDK2-NC. Compared with the
sgCDK2-NC group, early apoptosis of A375 cells infected with
lentivirus sgCDK2-108 was observed (Fig.
3A). The rate of early apoptosis in A375 cells infected with
lentivirus sgCDK2-108 was 11.76%, and the rate of total apoptosis
reached 12.47% (P<0.05; Fig. 3B),
suggesting that knockout of CDK2 induces early apoptosis in A375
cells.
CDK2 knockout alters the expression of
CDK4 and cyclin A2
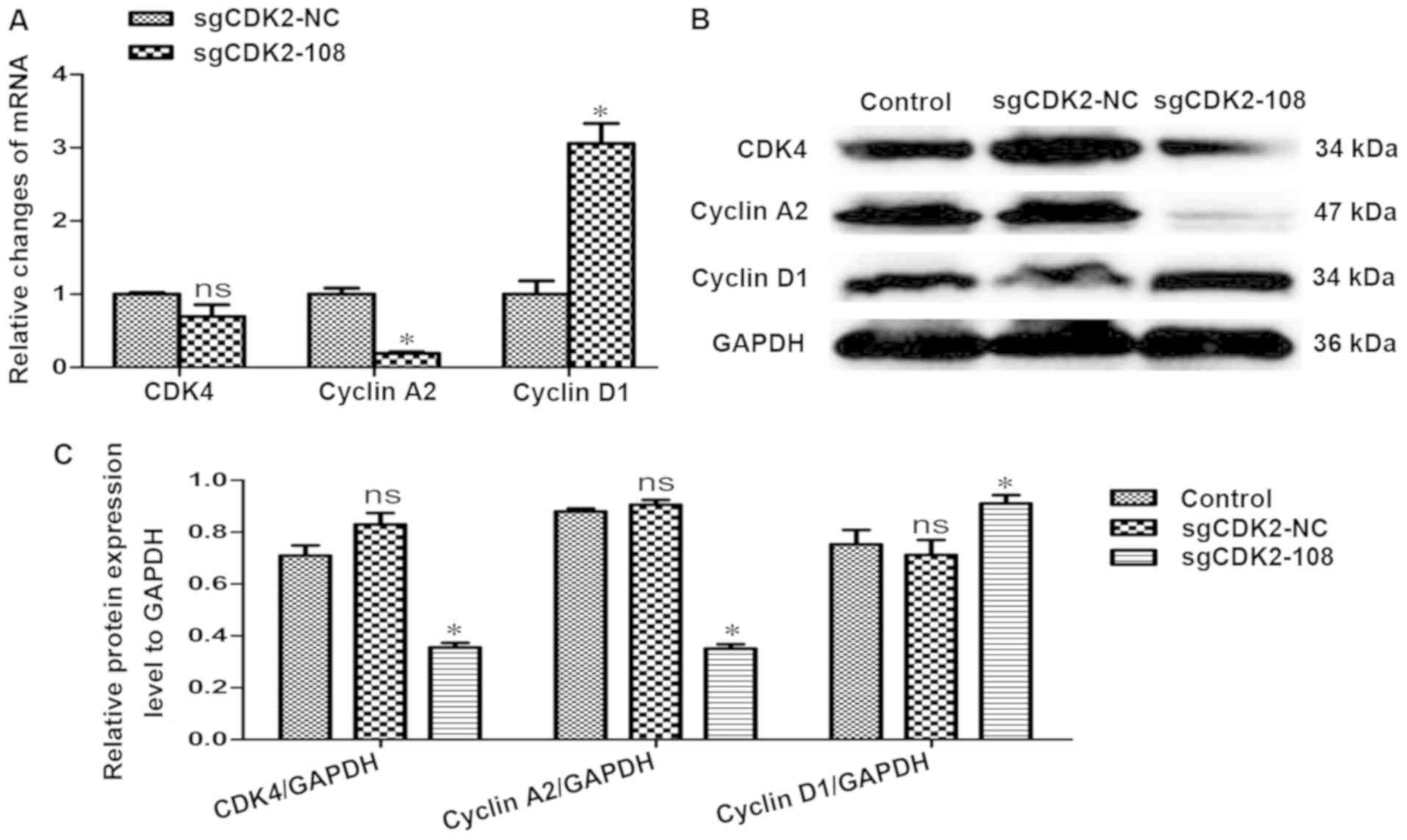
In order to investigate whether the cell
cycle-related proteins were differentially regulated, the relative
changes in CDK4, cyclin A2 and cyclin D1 expression were evaluated
with RT-qPCR and western blot analysis. The results revealed that
the transcriptional level of CDK4 was slightly downregulated
(P>0.05), the level of cyclin A2 was significantly downregulated
(P<0.05), whereas the level of cyclin D1 was significantly
upregulated (P<0.05) (Fig. 4A).
The protein levels of CDK4 and cyclin A2 were downregulated; in
particular, cyclin A2 was significantly decreased. However, the
protein expression of cyclin D1 was significantly upregulated
(P<0.05) (Fig. 4B and 4C).
Discussion
Previous studies demonstrated that the
colony-forming ability and cell viability were markedly inhibited
by knockdown of myosin VI using lentivirus-mediated shRNA in A375
melanocytes (23). Knockdown of
cyclin-dependent kinase-like 1 (CDKL1) by CDKL1-siRNA-expressing
lentivirus in A375 and MV3 cells also significantly inhibited cell
growth and colony-forming ability (24). Therefore, molecular-targeted therapy
using lentivirus-mediated shRNA has become a focus in anti-melanoma
research, as it is not associated with genotoxicity compared with
conventional chemotherapeutic agents. However, the outcome of
lentivirus-mediated gene knockdown may be affected by the
off-target effects using a single shRNA. CRISP/Cas9 technology is a
reliable method for gene editing, and CRISP/Cas9-mediated knockout
of the PDEF gene significantly inhibited the migration and invasion
of AGS human gastric cancer cells by transfection with
pX459-PDEF-sgRNA plasmids (25).
Therefore, CRISP/Cas9-based genome editing may provide novel
insight into cancer therapy (26–28).
CDK2 is a key regulator of the G1/S and S/G2 cell
cycle transitions; however, genetic deletion of CDK2 in p27
(Kip1)-null mice failed to suppress the development of pituitary
tumors (29). CDK2 was also found to
be a key mediator of epidermal growth factor-induced cell
transformation through directly phosphorylating ELK4 and regulating
c-fos expression (30).
Additionally, ablation of CDK2 significantly delayed S-M
progression and downregulated the expression of CDK6 (31). Therefore, the suitability of CDK2 as
a therapeutic target remains controversial.
In order to evaluate the role of CDK2 in regulating
cell cycle and mediating apoptosis of cutaneous melanoma cells, we
selected a single lentiviral vector to deliver nuclease Cas9, a
sgRNA, and a puromycin selection and enhanced green fluorescent
protein (EGFP) markers into target cells. A previous study using a
single lentiviral vector (lentiCRISPR) to deliver Cas9 and sgRNA
into target cells demonstrated that lentiCRISPs could abolish EGFP
fluorescence in 93±8% of infected cells at a low MOI of 0.3 for 11
days; however, lentiviral vectors expressing EGFP-targeting shRNA
were unable to completely knock down EGFP (32). Further study also demonstrated a
significant reduction in the diversity of sgRNAs in surviving human
melanoma A375 cells and human HUES62 stem cells transduced with the
GeCKO library at an MOI of 0.3 (32). This lentiviral SCRISP/Cas9 genome
editing system have been using in human cells (33). Two lentiviruses were constructed to
knock out CDK2 using CRISP/Cas9 technology. The results revealed a
successful lentivirus-mediated knockout of CDK2 using CRISP/Cas9
technology; the expression of CDK2 was also completely knocked out
in A375 cells. Although the nuclease Cas9 and sgRNA (sgCDK2-108)
were delivered into A375 cells by sgCDK2 lentivirus and CDK2
expression was abolished at the mRNA and protein levels, single
colonies and PCR identification at the DNA level were not
conducted. The homozygosity of the cells was not known, and the
lack of a precise genetic investigation is a limitation of the
study.
Further study demonstrated that the loss of CDK2
function significantly increased the percentage of cells in the
G0/G1 phase and induced G0/G1 phase arrest. The percentage of early
apoptotic A375 cells was also increased. These results indicated
that CDK2 plays a pivotal role in the regulation of cell cycle
transition, and may be associated with the progression of cutaneous
melanoma. Our study also demonstrated that the expression of CDK4
and cyclin A2 was downregulated, whereas the expression of cyclin
D1 was upregulated at the transcriptional and translational levels.
This result indicates that G0/G1 phase arrest is induced by
downregulated expression of CDK4 and cyclin A2, and upregulated
expression of cyclin D1. Subsequently, apoptosis occurs as a result
of G0/G1 phase arrest. Apoptosis as a protective mechanism ensures
homeostasis of host cells through cell shrinkage, fragmentation of
cellular DNA and formation of ‘apoptotic bodies’ leading to cell
death. Two pathways, namely ‘extrinsic’ and ‘intrinsic’ pathways,
activate caspases to cleave vital cellular proteins, and BCL-2
protein, as the first inhibitor of apoptosis, controls cell death
first though directly regulating the integrity of the outer
mitochondrial membrane (34,35). In the present study, apoptosis of
A375 cells occurred following knockout of CDK2 by flow cytometry,
but the changes of apoptotic-related proteins, such PARP, caspase-3
and BCL-2, were not evaluated by western blotting, which is another
limitation of this study. Further research will focus on the
mechanism of apoptosis of A375 cells following CDK2 knockout by a
lentiviral CRISP/Cas9 system. Elucidating the changes in whole
cellular proteins by proteomic analysis and investigating the role
of caspases or BCL-2 may provide more evidence regarding the
role(s) of CDK2 in human melanoma.
In conclusion, the results of the present study
demonstrated that CDK2 is crucial for cell cycle regulation through
controlling the G1/S transition in A375 human melanoma cells.
Therefore, knockout of CDK2 by CRISPR/Cas9 technology may provide a
novel therapeutic approach to cutaneous melanoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by Young and
Middle-aged Talent Project of Fujian Provincial Health Commission
(grant no. 2016-ZQN-90), and the Project of Bureau of Economic and
Information Technology of Tongan District, Xiamen (grant no.
2016-xt-01).
Availability of data and materials
All the datasets generated and analyzed in the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
HL conceived, designed, supervised the study and
wrote the manuscript. ZL and SH performed the experiments, QW and
LG analyzed the data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors have no commercial or other associations
that may pose a conflict of interest.
References
|
1
|
Little EG and Eide MJ: Update on the
current state of melanoma incidence. Dermatol Clin. 30:355–361.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Quintanilla-Dieck MJ and Bichakjian CK:
Management of early-stage melanoma. Facial Plast Surg Clin North
Am. 27:35–42. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guy GP Jr, Thomas CC, Thompson T, Watson
M, Massetti GM and Richardson LC; Centers for Disease Control and
Prevention (CDC), : Vital signs: Melanoma incidence and mortality
trends and projections-United States, 1982–2030. MMWR Morb Mortal
Wkly Rep. 64:591–596. 2015.PubMed/NCBI
|
|
4
|
Duffy KL, Truong A, Bowen GM, Andtbacka
RH, Hyngstrom J, Bowles T, Grossmann K, Khong H, Hyde M, Florell
SR, et al: Adequacy of 5-mm surgical excision margins for
non-lentiginous melanoma in situ. J Am Acad Dermatol. 71:835–838.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nadiminti H, Scope A, Marghoob AA, Busam K
and Nehal KS: Use of reflectance confocal microscopy to monitor
response of lentigo maligna to nonsurgical treatment. Dermatol
Surg. 36:177–184. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nurse P, Masui Y and Hartwell L:
Understanding the cell cycle. Nat Med. 4:1103–1106. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hunt T, Nasmyth K and Novák B: The cell
cycle. Philos Trans R Soc Lond B Biol Sci. 366:3494–3497. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Malumbres M: Physiological relevance of
cell cycle kinases. Physiol Rev. 91:973–1007. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hunt T: Nobel Lecture. Protein synthesis,
proteolysis, and cell cycle transitions. Biosci Rep. 22:465–486.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Merrick KA, Wohlbold L, Zhang C, Allen JJ,
Horiuchi D, Huskey NE, Goga A, Shokat KM and Fisher RP: Switching
Cdk2 on or off with small molecules to reveal requirements in human
cell proliferation. Mol Cell. 42:624–636. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guadagno TM and Newport JW: Cdk2 kinase is
required for entry into mitosis as a positive regulator of
Cdc2-cyclin B kinase activity. Cell. 84:73–82. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
McCurdy SR, Pacal M, Ahmad M and Bremner
R: A CDK2 activity signature predicts outcome in CDK2-low cancers.
Oncogene. 36:2491–2502. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chohan TA, Qian H, Pan Y and Chen JZ:
Cyclin-dependent kinase-2 as a target for cancer therapy: Progress
in the development of CDK2 inhibitors as anti-cancer agents. Curr
Med Chem. 22:237–263. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tetsu O and McCormick F: Proliferation of
cancer cells despite CDK2 inhibition. Cancer Cell. 3:233–245. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hochegger H, Dejsuphong D, Sonoda E,
Saberi A, Rajendra E, Kirk J, Hunt T and Takeda S: An essential
role for Cdk1 in S phase control is revealed via chemical genetics
in vertebrate cells. J Cell Biol. 178:257–268. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Berthet C, Aleem E, Coppola V, Tessarollo
L and Kaldis P: Cdk2 knockout mice are viable. Curr Biol.
13:1775–1785. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Enders GH: Mammalian interphase cdks:
Dispensable master regulators of the cell cycle. Genes Cancer.
3:614–618. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Adelmann CH, Wang T, Sabatini DM and
Lander ES: Genome-wide CRISPR/Cas9 screening for identification of
cancer genes in cell lines. Methods Mol Biol. 1907:125–136. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Slipek NJ, Varshney J and Largaespada DA:
CRISPR/Cas9-based positive screens for cancer-related traits.
Methods Mol Biol. 1907:137–144. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu HG, Liu Z, Jiang Y, Xiao JR, Li HY, Li
Z, Huo SS and Yu YJ: Apoptosis of human melanoma A375 cells induced
by lentivirus mediated CDK2-shRNA. Chin J Gerontol. 35:6345–6347.
2015.(In Chinese).
|
|
21
|
Liu HG, Liu Z, Jiang Y, Xiao JR, Li HY, Li
Z, Huo SS and Yu YJ: Apoptosis of human melanoma A375 cells induced
by adenovirus-associated virus mediated CDK2-shRNA. Chin J Control
Endemic Dis. 30:135–136. 2015.(In Chinese).
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li H, Zhou F, Wang H, Lin D, Chen G, Zuo
X, Sun L, Zhang X and Yang S: Knockdown of myosin VI by
lentivirus-mediated short hairpin RNA suppresses proliferation of
melanoma. Mol Med Rep. 12:6801–6806. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song Z, Lin J, Sun Z, Ni J and Sha Y:
RNAi-mediated downregulation of CDKL1 inhibits growth and
colony-formation ability, promotes apoptosis of human melanoma
cells. J Dermatol Sci. 79:57–63. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang YQ, Pei JH, Shi SS, Guo XS, Cui GY,
Li YF, Zhang HP and Hu WQ: CRISPR/Cas9-mediated knockout of the
PDEF gene inhibits migration and invasion of human gastric cancer
AGS cells. Biomed Pharmacother. 111:76–85. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yao S, He Z and Chen C:
CRISPR/Cas9-mediated genome editing of epigenetic factors for
cancer therapy. Hum Gene Ther. 26:463–471. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhen S and Li X: Oncogenic human
papillomavirus: Application of CRISPR/Cas9 therapeutic strategies
for cervical cancer. Cell Physiol Biochem. 44:2455–2466. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Biagioni A, Laurenzana A, Margheri F,
Chilla A, Fibbi G and So M: Delivery systems of CRISPR/Cas9-based
cancer gene therapy. J Biol Eng. 12:332018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Martín A, Odajima J, Hunt SL, Dubus P,
Ortega S, Malumbres M and Barbacid M: Cdk2 is dispensable for cell
cycle inhibition and tumor suppression mediated by p27(Kip1) and
p21(Cip1). Cancer Cell. 7:591–598. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Peng C, Zeng W, Su J, Kuang Y, He Y, Zhao
S, Zhang J, Ma W, Bode AM, Dong Z and Chen X: Cyclin-dependent
kinase 2 (CDK2) is a key mediator for EGF-induced cell
transformation mediated through the ELK4/c-Fos signaling pathway.
Oncogene. 35:1170–1179. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bacevic K, Lossaint G, Achour TN, Georget
V, Fisher D and Dulic V: Cdk2 strengthens the intra-S checkpoint
and counteracts cell cycle exit induced by DNA damage. Sci Rep.
7:134292017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shalem O, Sanjana NE, Hartenian E, Shi X,
Scott DA, Mikkelson T, Heckl D, Ebert BL, Root DE, Doench JG and
Zhang F: Genome-scale CRISPR-Cas9 knockout screening in human
cells. Science. 343:84–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Koike-Yusa H, Li Y, Tan EP,
Velasco-Herrera MC and Yusa K: Genome-wide recessive genetic
screening in mammalian cells with a lentiviral CRISPR-guide RNA
library. Nat Biotechnol. 32:267–273. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kale J, Osterlund EJ and Andrews DW: BCL-2
family proteins: Changing partners in the dance towards death. Cell
Death Differ. 25:65–80. 2018. View Article : Google Scholar
|
|
35
|
Adams JM and Cory S: The BCL-2 arbiters of
apoptosis and their growing role as cancer targets. Cell Death
Differ. 25:27–36. 2018. View Article : Google Scholar
|