Introduction
Clear cell sarcoma (CCS) is characterized by a
chromosomal t(12; 22) (q13; q12) translocation resulting in a
fusion between the Ewing sarcoma (EWSR1) and activating
transcription factor 1 (ATF1) genes (1). Detection of the EWSR1-ATF1
chimera gene product is widely used as a highly sensitive
diagnostic test for CCS (2). CCS
affects the deep soft tissues predominantly in young adults, aged
between 15 and 35 years of age and is known to have high rates of
metastasis, in worldwide (3,4). For
example, Chung and Enzinger (4)
reported that 50 out of 115 patients had died from metastatic
tumors in 1983. Despite progress in the different treatments
available, the prognosis of patients with CCS is still poor, as 30%
of patients have metastases at the time of diagnosis (5). Lymphatic metastasis is rare in other
types of malignant soft tissue tumors; however, is commonly
detected in CCS (4,6). A previous study reported that positive
sentinel nodes were identified in 2 out of 42 patients with
synovial sarcoma, compared with 6 out of 12 patients with CCS
(6,7). Radical surgical resection is the first
line of treatment for CCS; however, the rate of local recurrence
can be as high as 84% and the rate of late metastases (10 years
following surgery) can be up to 63%, which have been associated
with a 5 to 20-year survival rate of 67-10% (8). As CCS has been found to be resistant
to conventional soft tissue sarcoma chemotherapy regimens, for
example doxorubicin-based chemotherapy (8), therefore therapies that specifically
control metastasis are urgently required.
For the development of novel targeted therapies, a
CCS model, that exhibited similar clinicopathological features,
including metastatic potential, was established in our previous
study, by xenoplanting HS-MM CCS cells into SCID-Beige mice
(9), which was subsequently used to
investigate the pharmacological effect of a lipoate analogue,
CPI-613(9) or an anti-CD151
antibody (10).
During the establishment of the CCS murine model, in
both SCID-Beige and BALB/c nude mice, it was found that the latter
mice showed no metastasis (9). A
SCID-Beige mouse is a double-mutant mouse strain, with impaired
lymphoid development and weak NK cell activity (11). By contrast, the BALB/c nude mouse
has robust NK cell activity, which complements deficiencies in
thymus-dependent T lymphocyte function (12).
The aim of the present study was to investigate
whether NK cells impaired metastasis of CCS in murine xenoplanted
models.
Materials and methods
HS-MM clear cell sarcoma cell
line
The HS-MM CCS cell line was previously established
and characterized in our laboratory (13,14).
HS-MM cells harbor a canonical genetic background with t(12; 22)
(q13; q12) of CCS, which results in an EWSR1-ATF1 fusion
gene (1). Cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc.) containing 10% heat-inactivated fetal bovine
serum (HyClone; GE Healthcare Life Sciences). Cells were passaged
for no more than six months following resuscitation. Cells were
screened periodically for mycoplasma contamination using DAPI
staining.
Mice
The animal experiments were conducted at Gifu
University under the guidelines for animal experimentation and
followed the Japanese Law for the Humane Treatment and Management
of Animals. The experimental protocol was approved by the Animal
Care Committee of Gifu Graduate School of Gifu (approval no. 27-80
and 2020-066). A total of 3 H-2d congenic strains, namely,
SCID-Beige
(CB17.Cg-PrkdcscidLystbg-J/CrlCrlj;
weight, 18.3-21.1 g; n=19), BALB/c nude (BALB/c-nu;
CAnN.Cg-Foxn1nu/CrlCrlj; weight 17.0-18.9 g;
n=10), and BALB/c mice (weight 19.7-22.4 g; n=5), were purchased
from Charles River Laboratories, Inc. All mice were female and
8-weeks-old, and kept under specific pathogen-free conditions in
isolated and ventilated cages, with free access to food and water,
and maintained with a 12-h light/dark cycle at 23˚C. Every effort
was made to minimize suffering as previously described (9,10).
Briefly, murine behavior and body weight were monitored twice per
week. A 50 mg/ml solution of pentobarbital in sterile saline was
administered intraperitoneally to SCID-Beige and BALB/c nude mice,
at a dose of 120 mg/kg for euthanasia and were confirmed to have
died when the heart stopped beating. Time to initiation of tissue
collection was 15 min. To obtain the spleen from the BALB/c mice,
carbon dioxide inhalation was used, as the method of euthanasia
until the mice stopped breathing. Carbon dioxide (100%) was
introduced into a 5.7L cage, initially containing normal air
conditions, with the lid closed, at a low-flow rate of 30% volume
displacement per minute. Before the experiments, humane endpoints
were set as follows: i) When the body weight following tumor cell
inoculation decreased by >10% compared with the initial weight
of the mouse; ii) when the behavioral observations, such as
food/water consumption and daily activity, monitored during the
experiment, were reduced by 50%; and iii) when the fur became dull.
Finally, if ulceration was observed at the transplantation site,
the mouse was sacrificed.
However, among the 19 SCID-Beige and 10 BALB/c nu/nu
mice examined, there was no suffering in any of the mice using the
aforementioned criteria nor were any of the mice found dead. In
addition, 5 BALB/c mice, which were used to obtain the NK-cells
exhibited no suffering prior to euthanasia.
Xenoplantation and injection of NK
cell fraction
Xenoplantation of HS-MM cells was performed as
previously described (9,10). Briefly, SCID-Beige mice were
injected with 2.5x107 HS-MM cells into the aponeuroses
of the thighs, subsequently 9 mice were randomly divided to three
groups, of three mice. A total of 0.3 ml saline, with or without
1x104 murine or 1.5x106 human NK cells was
intravenously injected to SCID-Beige mice two weeks following HS-MM
xenoplantation. A total of 8 weeks following xenoplantation, the
mice were sacrificed, and the tumor size was measured using a
caliper and the following formula: Volume=(width2 x
length)/2.
NK cells were harvested from splenic cells of BALB/c
mice using a mouse NK cell isolation kit (Stemcell Technologies,
Inc.), or collected from peripheral blood samples from a healthy
volunteer using negative selection and a NK Cell Isolation kit,
MACS system (Miltenyi Biotec, Inc.) according to the manufacturer's
protocol. Written informed consent was provided from the healthy
volunteer prior to the start of the study. The study was approved
by the Committee of Gifu Graduate School of Gifu, Japan (approval
no. 27-80, 2020-066).
The isolated human NK cells were cultured in T25
flasks (Thermo Fisher Scientific, Inc.) with 10 ml KBM502 medium
(Kohjin-bio, Co., Ltd.) at 37˚C in a humidified incubator with 5%
CO2 for seven days.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted using the Qiagen RNeasy Kit
(Qiagen GmbH). cDNA synthesis from total RNA and subsequent PCR was
performed using the RT-PCR kit (Takara Bio Inc.) according to the
manufacturer's instructions and as previously described (15). qPCR was performed using the
FastStart Essential DNA SYBR® Green master mix according
to the manufacturer's instructions using a LightCycler (both Roche
Diagnostics GmbH) as previously described (16). Briefly, cells were isolated from the
buffy coat of 0.5 ml right ventricular blood, obtained from
SCID-Beige mice injected with murine NK cells (n=3) and the mice
without NK cell injection group (n=3) after euthanasia. Total RNA
was extracted and reverse transcribed, following which cDNA (1 µl
per reaction) was diluted with PCR mix, containing 0.2 pmol
primers, to a final reaction volume of 20 µl. The following qPCR
primers were used: EWSR1-ATF1 forward,
5'-CATGAGCAGAGGTGGGCG-3' and reverse,
5'-CCCCGTGTATCTTCAGAAGATAAGTC-3'; and GAPDH forward,
5'-GAAATCCCATCACCATCTTCCAGG-3' and reverse,
5'-GAGCCCCAGCCTTCTCCATG-3'. Samples were performed in triplicate,
and expression levels of each target gene were analyzed with a
LightCycler system using the 2-ΔΔCq method described by
Livak and Schmittgen (17). For
each triplicate set, the ΔCq values were normalized to GAPDH
expression in the control group. The values of the target group
were then calculated as the fold change relative to the mean values
of the control group (control; set to 1.0). An unpaired Student's
t-test was used to determine significant differences in gene
expression among the treatment groups. P<0.05 was considered to
indicate a statistically significant difference.
Anti-asialo GM1 antibody
treatment
A rabbit antibody targeting asialo-GM1 was purchased
from FUJIFILM Wako Pure Chemical Corporation (cat no. 014-09801).
BALB/c nude mice were intraperitoneally injected with or without
300 µg anti asialo-GM1 antibody at 7, 14, 21, and 28 days following
xenoplantation with 2.5x107 HS-MM cells.
Anti-CD96 antibody treatment
Rat monoclonal antibody against murine CD96 (clone
630612) was purchased from R&D Systems Inc. SCID-Beige mice
were intraperitoneally injected with or without 12.5 µg anti-CD96
antibody at 7, 14, 21, and 28 days following injection with HS-MM
cells. Intraperitoneal metastatic lymph nodes were dissected using
a stereomicroscope. Total metastatic lymph nodes were then
weighed.
Histopathological analysis
The tumor tissue sections from the primary injection
site and metastatic tumor in mice, which were transplanted with
murine NK cells, treated with anti-CD96 antibody, and control mice
were fixed for 48 h at room temperature with 10% neutral buffered
formalin, embedded in paraffin, and cut into 4-µm thick sections.
Paraffin slides were deparaffinized in xylene, twice, for 10 min
and dehydrated in a descending alcohol series (95, 90, 70%) for 2
min, each time. Following rinsing with water, tissue sections were
stained with Mayer hematoxylin solution for 8 min, then with eosin
Y solution for 1 min, both at room temperature. The tissue sections
were evaluated using light microscopy at x400 magnification.
Statistical analysis
The results are presented as the mean ± standard
deviation of triplicate experiments. Statistical analyses were
performed using EZR (v1.27) (18),
which is a graphical user interface for R (19). Statistical significance was
determined using an unpaired Student's t-test, Welch's t-test or
one-way ANOVA with the Bonferroni correction. P<0.05 was
considered to indicate a statistically significant difference.
Results
NK cells abrogate metastasis of CSS
without affecting the growth of tumors at the primary injection
site in SCID-Beige xenoplant model
The HS-MM cells were successfully transplanted into
the soft tissues of the thighs of all the SCID-Beige mice. At the
end of the experiments, the body weights of the mice in each group
were as follows: 22.5, 21.8, and 22.9 g for the control mice; 20.8,
21.0, and 22.2 g for the murine NK cell-transplanted mice; and
20.7, 20.6, and 21.2 g for the human NK cell-transplanted mice; the
weights of the control mice were significantly higher compared with
those of the human NK cell-transplanted mice (P=0.039; Fig. S1). The metastasis of HS-MM was
examined using a stereomicroscope following euthanasia.
Histopathological examination of the primary injection site, with
or without murine NK cell-transplantation or anti-CD96
antibody-injection in the tumor tissues was also performed. There
were no histopathological alternations, such as the presence of
necrosis or lymphocyte infiltration at the primary injection site
by these treatments (Fig. S2).
Therefore, the results of the present study suggest that the
activity of NK cells might abrogate the formation of the metastatic
tumor but does not affect the primary tumor.
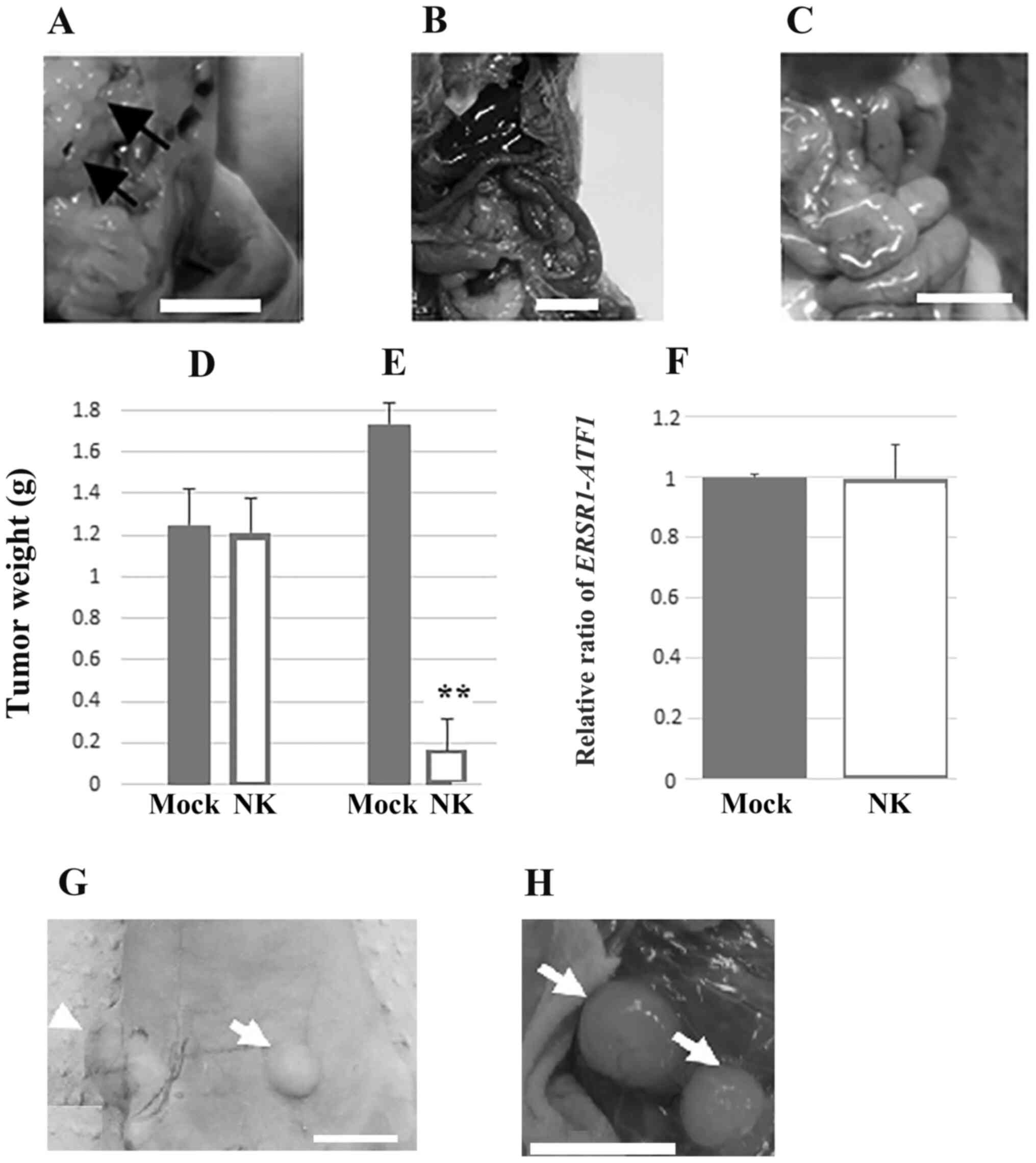
Lymph node and distant metastasis, i.e., lung
metastasis, were detected at 8 weeks, following injection in all
the inoculated mice. Representative images of mesenteric lymph node
metastasis are shown in Fig. 1A. By
contrast, murine or human NK cell transplantation inhibited
metastasis of HS-MM cells in SCID-Beige mice (Fig. 1B and C, respectively). The growth of xenoplanted
HS-MM cells at the injection sites was not altered by the adoptive
transfer of human NK cells (1.25±0.18 vs. 1.21±0.17 g for non-NK
cell transplanted and NK cell-transplanted group, respectively;
n=3; P=0.800; Fig. 1D), whereas the
growth of multiple disseminated tumors in the mesentery was
significantly inhibited (1.74±0.10 vs. 0.17±0.15 g in the non-NK
cell transplanted and NK cell-transplanted group, respectively;
n=3; P<0.001; Fig. 1E).
NK cells did not affect the mRNA
expression of EWSR1-ATF1 in circulating blood cells
NK cell transplantation did not induce significant
changes in the transcriptional expression of EWSR1-ATF1 in
the buffy coat cells from the right ventricular blood of SCID-Beige
mice (1 vs. 0.996±0.11 in the control and murine NK
cell-transplanted group; n=5; P=0.900; Fig. 1F).
Effect of anti-asialo GM1 and
anti-CD96 antibody on metastasis of CCS
At the end of the experiments, the body weights of
the mice were as follows: 20.9, 21.1, 21.2, 20.7, and 21.4 g for
the control mice and 21.2, 21.0, 21.0, 21.7, and 22.3 g for the
anti-asialo-GM1 antibody-treated mice. No significant differences
were observed in the body weights of the control and
anti-asialo-GM1 antibody-treated mice (P=0.259). Notably, treatment
with the anti-asialo GM1 antibody markedly promoted the metastasis
of xenoplanted HS-MM cells in the BALB/c nude mice (Fig. 1G and H).
Previous studies have found CD96 to be a negative
regulator of the NK-cell response in the tumor microenvironment by
using different tumor models (20,21).
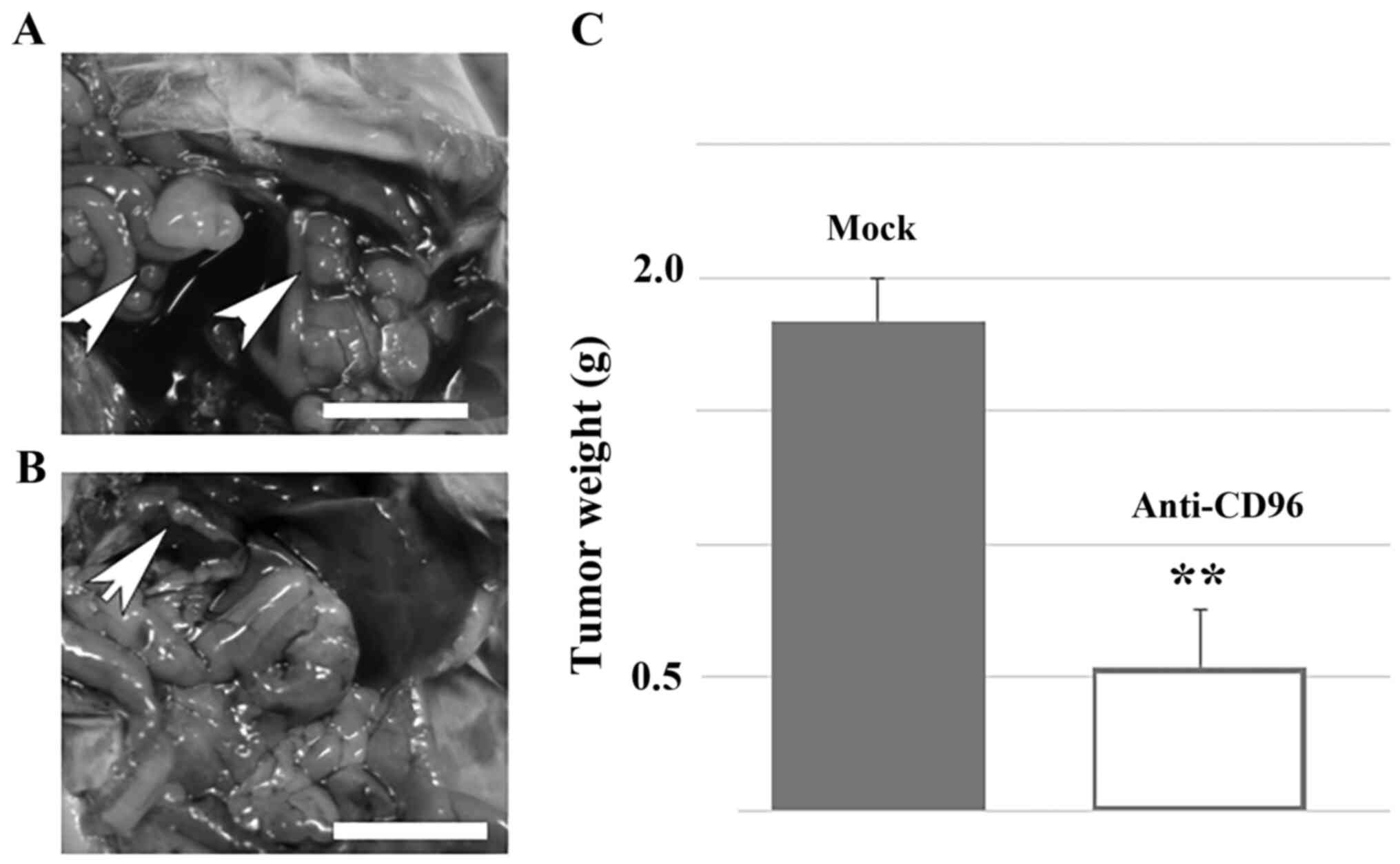
Therefore, it was investigated whether forced activation of
residual NK cells could reverse the metastasis of HS-MM cells in
SCID-Beige mice. The representative data are shown in Fig. 2. Notably, intraperitoneal injection
of anti-CD96 antibody significantly reversed the metastasis of
HS-MM cells in the present SCID-Beige mice model (1.84±0.16 vs.
0.54±0.21 g in the control and NK cell-transplanted group,
respectively; n=5; P<0.001). Furthermore, at the end of the
experiments, the body weights of the mice were as follows: 21.8,
21.0, 20.9, 21.4, and 21.6 g for the control mice and 21.4, 21.8,
21.3, 21.0, and 22.3 g for the anti-CD96 antibody-treated mice. No
significant differences were observed in the body weights of the
two groups (P=0.460).
These results demonstrated the critical role of NK
cell activity in suppressing metastasis of xenoplanted HS-MM
cells.
Discussion
The rapid growth and robust metastatic activity of
the HS-MM cells was observed during the short duration of the
experiments, which was 8 weeks following transplantation.
Therefore, the present murine model, which caused distant
metastasis, as observed in humans, was a suitable metastatic tumor
model as previously described (9,10).
Metastasis is a complex process, which proceeds in
several steps, including local invasion and intravasation,
transport through the circulation, extravasation, micrometastasis,
and subsequent development of overt metastatic tumors (22). In the present study, CCS-specific
transcription of ERSR1-ATF1 chimera gene was equally
detected in the buffy coats in cells from xenoplanted SCID-Beige
mice and in mice transplanted with murine NK cells (Fig. 1F). This result indicated the
expression of the EWSR1-ATF1 fusion gene product in
the blood was not reduced by adoptive NK cell transplantation in
the present murine model. We hypothesize that NK cells suppress the
formation of the tumor environment during the micrometastasis step,
including foreign tumor microenvironments in distant tissues. This
is consistent with the recent hypothesis that NK cells are
recruited by a subpopulation of patrolling non-classical monocytes
to the extravasating tumor cells to prevent the development of a
metastatic microenvironment (23).
CD96 has recently been found to be a negative
regulator of NK cell activity (18). The findings from the present study
also revealed that anti-CD96 antibody treatment markedly reduced
metastasis of HS-MM cells in SCID-Beige mice, in which NK cell
activity against the YAC cell line was ~50% lower compared with
that in SCID mice (11). Thus,
inoculation with the anti-CD96 antibody, which also enhances NK
cell activity (23), or targeting
other inhibitory immune checkpoint receptors for NK cell, for
example T cell immunoglobulin and immunoreceptor tyrosine-based
inhibitory motif domain (24),
could potentially be used for regulating tumor metastasis through
the inhibition of micrometastasis in the tumor environment to
improve survival in patients with CCS following surgery. In the
present study, adoptively transferred human NK cells significantly
suppressed the metastasis of human CCS cells. This suggests that
adoptive transfer of NK cells in patients with CCS could have a
potential therapeutic effect. The present study also demonstrated
that anti-asialo-GM1 antibody, which is well-known to impair the NK
cell activity (25), induced
metastasis of CCS in BALB/c nude mice. Therefore, it should be
further examined whether surgical treatment might impair NK cell
activity and attribute to the metastasis of CCS. Furthermore,
extensive studies are also required to unravel whether NK cells
directly attach to the CCS at the micrometastasis site or whether
NK cells are important to inhibit the formation of micrometastasis
environment.
In conclusion, the results of the present study
indicated that NK cells may impair the metastatic tumor
microenvironment in the mouse xenoplant model.
Supplementary Material
Body weights of SCID-Beige mice
transplanted with HS-MM cells. NK, natural killer.
Representative images of the
histopathological examination of tumors at the primary injection
site in mice (A) transplanted with natural killer cells or (B) were
treated with anti-CD96 antibody compared to the (C) control mouse.
No histopathological differences among the different groups were
observed. Scale bar, 100 μm.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
TT conceived and designed the study. YH and TT
analyzed, interpreted data and drafted the manuscript. YH, CS, and
YK performed the experiments. All authors participated in the
writing of the manuscript and have read and approved the final
manuscript.
Ethics approval and consent to
participate
The animal experiments were approved by the Animal
Care Committee of Gifu Graduate School of Gifu, Japan (approval no.
27-80). The use of human tissues was approved by the Committee of
Gifu Graduate School of Gifu, Japan (approval no. 27-80, 2020-066).
Written informed consent was provided by the healthy volunteer.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zucman J, Delattre O, Desmaze C, Epstein
AL, Stenman G, Speleman F, Fletchers CD, Aurias A and Thomas G: EWS
and ATF-1 gene fusion induced by t(12;22) translocation in
malignant melanoma of soft parts. Nat Genet. 4:341–345.
1993.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Thway K and Fisher C: Tumors with
EWSR1-CREB1 and EWSR1-ATF1 fusions: The current status. Am J Surg
Pathol. 36:e1–e11. 2012.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Enzinger FM: Clear-cell sarcoma of tendons
and aponeuroses. An analysis of 21 cases. Cancer. 18:1163–1174.
1965.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Chung EB and Enzinger FM: Malignant
melanoma of soft parts. A reassessment of clear cell sarcoma. Am J
Surg Pathol. 7:405–413. 1983.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ibrahim RM, Steenstrup Jensen S and Juel
J: Clear cell sarcoma-A review. J Orthop. 15:963–966.
2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Andreou D and Tunn PU: Sentinel node
biopsy in soft tissue sarcoma. Recent Results Cancer Res.
179:25–36. 2009.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Andreou D, Boldt H, Werner M, Hamann C,
Pink D and Tunn PU: Sentinel node biopsy in soft tissue sarcoma
subtypes with a high propensity for regional lymphatic
spread-results of a large prospective trial. Ann Oncol.
24:1400–1405. 2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Mavrogenis A, Bianchi G, Stavropoulos N,
Papagelopoulos P and Ruggieri P: Clinicopathological features,
diagnosis and treatment of clear cell sarcoma/melanoma of soft
parts. Hippokratia. 17:298–302. 2013.PubMed/NCBI
|
|
9
|
Egawa Y, Saigo C, Kito Y, Moriki T and
Takeuchi T: Therapeutic potential of CPI-613 for targeting tumorous
mitochondrial energy metabolism and inhibiting autophagy in clear
cell sarcoma. PLoS One. 13(e0198940)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kawashima K, Saigo C, Kito Y, Hanamatsu Y,
Egawa Y and Takeuchi T: CD151 confers metastatic potential to clear
cell sarcoma of the soft tissue in animal model. Oncol Lett.
17:4811–4818. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
MacDougall JR, Croy BA, Chapeau C and
Clark DA: Demonstration of a splenic cytotoxic effector cell in
mice of genotype SCID/SCID.BG/BG. Cell Immunol. 130:106–117.
1990.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Budzynski W and Radzikowski C: Cytotoxic
cells in immunodeficient athymic mice. Immunopharmacol
Immunotoxicol. 16:319–346. 1994.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Sonobe H, Furihata M, Iwata J, Ohtsuki Y,
Mizobuchi H, Yamamoto H and Kumano O: Establishment and
characterization of a new human clear-cell sarcoma cell-line,
HS-MM. J Pathol. 169:317–322. 1993.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Sonobe H, Takeuchi T, Taguchi T, Shimizu
K, Iwata J, Furihata M and Ohtsuki Y: Further characterization of
the human clear cell sarcoma cell line HS-MM demonstrating a
specific t(12;22)(q13;q12) translocation and hybrid EWSR1/ATF-1
transcript. J Pahol. 187:594–597. 1999.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Takeuchi T, Adachi Y and Nagayama T: A
WWOX-binding molecule, transmembrane protein 207, is related to the
invasiveness of gastric signet-ring cell carcinoma. Carcinogenesis.
33:548–554. 2012.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Asano Y, Takeuchi T, Okubo H, Saigo C,
Kito Y, Iwata Y, Futamura M and Yoshida K: Nuclear localization of
LDL receptor-related protein 1B in mammary gland carcinogenesis. J
Mol Med (Berl). 97:257–268. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta C(T)) method. Methods. 25:402–408. 2001.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kanda Y: Investigation of the freely
available easy-to-use software ‘EZR’ for medical statistics. Bone
Marrow Transplant. 48:452–458. 2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
R Core Team: R: A language and environment
for statistical computing. R Foundation for Statistical Computing,
Vienna, 2014.
|
|
20
|
Chan CJ, Martinet L, Gilfillan S,
Souza-Fonseca-Guimaraes F, Chow MT, Town L, Ritchie DS, Colonna M,
Andrews DM and Smyth MJ: The receptors CD96 and CD226 oppose each
other in the regulation of natural killer cell functions. Nat
Immunol. 15:431–438. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Ochoa MC, Minute L, Rodriguez I, Garasa S,
Perez-Ruiz E, Inogés S, Melero I and Berraondo P:
Antibody-dependent cell cytotoxicity: Immunotherapy strategies
enhancing effector NK cells. Immunol Cell Biol. 95:347–355.
2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Faltas B: Cornering metastases:
Therapeutic targeting of circulating tumor cells and stem cells.
Front Oncol. 2(68)2012.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Narasimhan PB, Eggert T, Zhu YP,
Marcovecchio P, Meyer MA, Wu R and Hedrick CC: Patrolling monocytes
control NK cell expression of activating and stimulatory receptors
to curtail lung metastases. J Immunol. 204:192–198. 2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Bi J and Tian Z: NK cell dysfunction and
checkpoint immunotherapy. Front Immunol. 10(1999)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
López-Soto A, Gonzalez S, Smyth MJ and
Galluzzi L: Control of metastasis by NK cells. Cancer Cell.
32:135–154. 2017.PubMed/NCBI View Article : Google Scholar
|