Epilepsy is a brain disorder characterized by
recurrent seizures, which is diagnosed in 4 to 10 out of every
1,000 individuals in developed countries and affects 75 million
individuals worldwide (1-3).
The etiology of epileptic disorders is complex and may be of
genetic, developmental or acquired origin (4,5). There
is a balance between excitatory and inhibitory synaptic mediators
[glutamate and gamma-aminobutyric acid (GABA)] in the healthy
brain, and a shift of this balance towards excitation is considered
the primary cause of epilepsy (6).
In addition, serotonergic receptors (7,8),
neuroinflammation (9-11),
nitric oxide pathway (12) and
various ion channels, such as calcium ions (13) may also play a critical role in the
mechanism of epilepsy.
There is ample evidence to indicate that the
noradrenergic system plays a key role in the regulation of
epileptogenesis and convulsions (14,15).
Norepinephrine (NE) is generally synthesized and released from
noradrenergic nerve endings in the locus coeruleus (LC) (16,17).
Abnormal NE secretion causes an increase in tonic/clonic seizures
in mice genetically prone to epileptic seizures (18). Although the LC is a small brainstem
nucleus, it is the sole source of NE in the neocortex, hippocampus
and cerebellum. NE is a potent neuromodulator involved in
regulating the excitability of large-scale brain regions. NE
concentrations have been reported to increase at seizure onset and
decrease during or shortly following the seizure (19).
The inhibition of NE release by gabapentin and
pregabalin has an anticonvulsant effect. These drugs exert their
effects by binding to the α2δ subunit of voltage-sensitive
Ca2+ channels. Similarly, gabapentin and pregabalin
cause a decrease in NE release through an increase in the
extracellular K+ concentration (20). In another study, blocking
voltage-sensitive Ca2+ channels with melatonin exerted
an anti-epileptic effect by inhibiting NE release (21). In addition, the density of adrenergic
receptors (ARs) in various brain areas decreases during seizures
(22,23). NE exerts a pronounced suppressive
effect on the development of epileptic seizures. Consistent with
this, a decrease in the NE concentration or the administration of
AR antagonists causes an increase in the frequency of seizures
(24,25). However, there is evidence to suggest
that increased NE levels under certain conditions activate
seizures, possibly via different ARs (15,26,27).
Furthermore, exposure to specific β2-adrenergic agonist
drugs poses a significant risk for epilepsy (28). Conversely, the β-AR antagonist,
propranolol, has been shown to reduce pentylenetetrazole
(PTZ)-induced tonic/clonic seizures (29).
The hippocampus plays a crucial role in the
pathogenesis of epilepsy and the activation of the
α1A-AR increases the inhibitory tone in the CA1 region
of the hippocampus (30). Selective
α1A-AR activation increases action potential firing in a
subpopulation of hippocampal CA1 interneurons. In response to this,
Na+ influx is initiated independently of second
messenger signaling. In addition, α1A-AR activation
decreases activity due to increased pre-synaptic GABA in CA1
pyramidal cells (30). Furthermore,
blockade of the α1B adrenoceptor subtype exerts both
neuroprotective and anti-epileptic effects (31).
Evidently, the activation of different ARs leads to
complex effects on epileptic seizures that have not yet been fully
elucidated. In the present review, the role of the adrenergic
system in epilepsy and the therapeutic potential of AR agonists are
discussed.
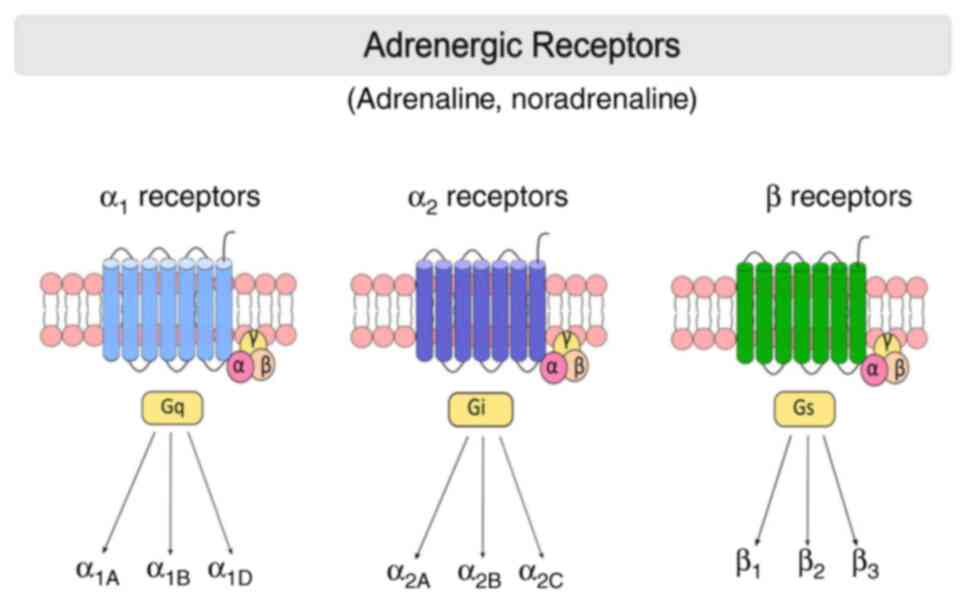
ARs are membrane-bound G protein-coupled receptors
(GPCRs) that mediate the peripheral and central effects of NE. ARs
are first divided into two major groups: α- and β-ARs (39). In recent years, the development of
new pharmacological tools has revealed nine different subtypes of
ARs: Three α1-ARs (α1A, α1B and
α1D), three α2-ARs
(α2A/D, α2B and
α2C) and three β-ARs (β1, β2 and
β3) (40) (Fig. 1).
β-ARs are essential components of the sympathetic
nervous system and belong to the superfamily of GPCRs (49). Subsequently, adenylate cyclase (AC)
activation causes an increase in cAMP, the main modulator of
intracellular events (50).
β1-AR subtypes constitute 70-80% of cardiac β-ARs
(49). β2-ARs are mostly
found in airway smooth muscle. In addition, β2-AR are
detected in alveolar type II cells, uterine muscle, mast cells,
mucous glands, skeletal muscle, epithelial cells and vascular
endothelium (51).
Increasing extracellular hippocampal dopamine and
GABA secretions plays a critical role in the anticonvulsant effect
of the NE reuptake inhibitor maprotiline. Moreover, the
anticonvulsant effect of maprotiline is potentiated by the
administration of a selective α2- and
β2-agonists. On the other hand, α1D receptor
agonists reduce the anticonvulsant effect (78). The α2-AR selective
agonist, dexmedetomidine, exerts anticonvulsant effects on
PTZ-induced seizures, whereas the α2-AR antagonist ATI
facilitates epileptic seizures in rats (66). Furthermore, dexmedetomidine
significantly reduced the number of c-Fos positive cells in the rat
brain (66). However, another study
demonstrated a pro-epileptic effect of dexmedetomidine in
spike-wave epilepsy in WAG/Rij rats (84). In previous a study on the rat
hippocampus, the α2-AR antagonist was implicated in the NE-mediated
anti-epileptic effect in the CA3 domain (85). Electrical brain stimulation in the
rat hippocampus exerts an inhibitory effect on epileptiform
activity via α1 and α2 ARs (86,87).
Moreover, the α2-AR agonist, yohimbine, and adenosine
provide an additive effect to increase the seizure threshold
induced by pentylenetetrazole in mice (36). Experimental evidence has revealed
that the specific cannabinoid CB1 agonist, ACEA, is
involved in its anticonvulsant properties by functionally
interacting with α2-adrenoceptors in PTZ-induced
seizures in mice (32).
Β-ARs are low affinity receptors for NE and are
activated during periods of intense LC activation with a high NE
release. The prolonged stimulation of β-ARs leads to a decrease in
their sensitivity (90). β-AR is
extensively distributed in the amygdala (37). Long-term antidepressant treatment
downregulates β-receptors in the amygdala and leads to an increase
in epileptic seizures in rats (24).
Similarly, reductions in the concentration of β-ARs in the amygdala
of epileptic animals may contribute to facilitating seizures
(38). The administration of
β2-AR agonists to mice also causes a reduction in
PTZ-induced seizures (82). In
addition, the β2-agonist, salbutamol, has been shown to
exhibit anti-epileptic activity in maximal electroshock-induced
seizures in mice (91).
The role of β-ARs in epileptic seizure
susceptibility is largely unclear, and there are conflicting
findings in different studies. An increase in seizures may be an
expected result in studies using β-AR blockers (92). By contrast, there are different
studies demonstrating that β-AR antagonists exert anticonvulsant
effects in various animal models of seizures (93,94). The
non-selective β-AR antagonist, propranolol, exerts an
anticonvulsant effect by blocking the sodium channel rather than
its hippocampal effects (95).
However, it is stated that a similar mechanism is responsible for
the anticonvulsant effect of clenbuterol, which is a
β2-AR agonist (1).
Moreover, the stimulation of β2-ARs reduces limbic
seizures by increasing hippocampal dopamine levels (78). The α-receptor antagonist,
phentolamine, selectively reduces anticonvulsant effects, while the
β-receptor antagonist, timolol, blocks proconvulsant activity
(96). These results suggest that
there are different mechanisms in seizure formation in various
animal models. Nevertheless, these results clearly indicate that
β2-AR activation plays a critical role in the
anticonvulsant effect of NE.
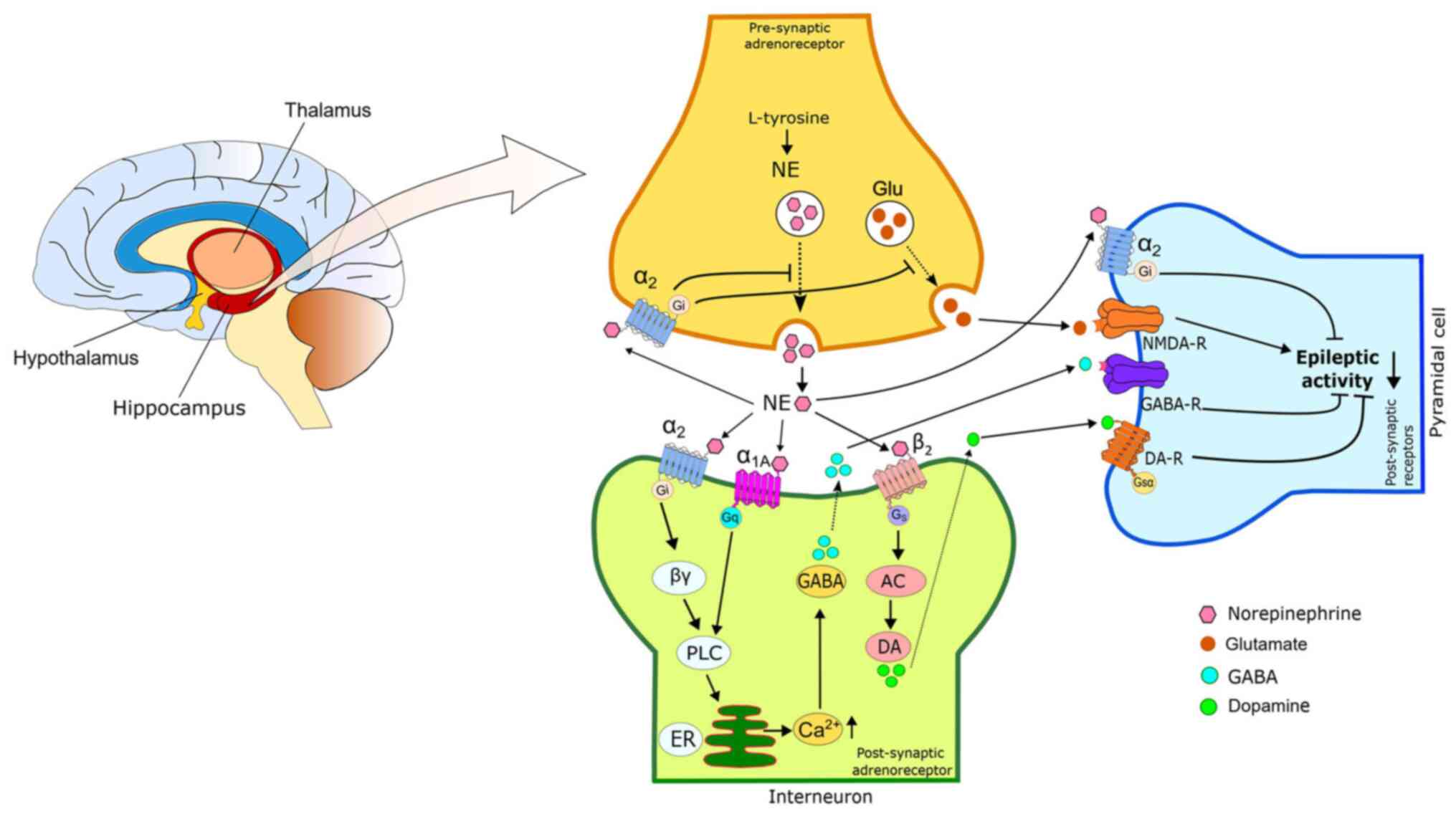
NE exerts excitatory and inhibitory effects on
neuronal excitability, depending on receptor subtypes and
locations. However, there is evidence to suggest that the dominant
effect of NE suppresses excitability in a number of brain regions
(83,97). It is a known fact that the
pathogenesis of epileptic seizures is associated with the
hyperexcitability of brain neurons. Therefore, it is important that
NE reduces excitability in its anti-epileptic effect. The effect of
NE on neuronal excitability may be via modulation of the
conductivity of ion channels or indirectly, usually through
GABAergic and glutamatergic transmission (83). Evidence has shown that activating the
noradrenergic system facilitates the presynaptic release of GABA
(68). In addition, GABA induces NE
release by activating GABAA receptors at noradrenergic
nerve terminals (98). NE has the
ability to alter the excitability of GABAergic cells in certain
brain regions (99). For example,
the chronic use of certain antidepressant drugs (e.g., citalopram
and fluoxetine) that increase NE levels causes the downregulation
of ARs and GABAA receptors (100). This regulation may be one of the
possible reasons for the proconvulsant effect of chronic
antidepressant therapy. The activation of a1-ARs can cause
epileptic seizures by increasing GABAergic transmission in various
brain limbic regions, including the hippocampus (101), piriform cortex (100) and amygdala (102). The activation of α1-ARs
through a decrease in potassium conductivity decreases epileptic
seizures in the hippocampus by depolarizing inhibitory interneurons
(30,101). In a previous study on the medial
prefrontal cortex, it was found that the stimulation of
α1-ARs with phenylephrine facilitated GABAergic
transmission to pyramidal neurons (75).
Numerous noradrenergic neurons from the LC make
synaptic connections with GABAergic interneurons in the basolateral
amygdala. Through the activation of α1-ARs, NE
depolarizes GABAergic interneurons in the amygdala and increases
GABA transmission. This causes the inhibition of pyramidal
glutamatergic cells (103). Stress
suppresses NE-mediated GABAergic transmission. Therefore, it is
suggested that this is a possible mechanism underlying the increase
in stress-induced seizure activity (102). A significant association has been
found between the decrease in the density of α2-ARs in
the amygdala of mice and epileptic seizures (64).
There is evidence to suggest strong associations
between the adrenergic and glutamatergic systems in the brain. NE
secretion also exerts prominent effects on the neuronal excitatory
glutamate system (104). NE plays a
key role in regulating the sensitivity of specific postsynaptic
glutamate receptors (105). It has
been stated that ionotropic glutamate receptors play a critical
role in the regulation of NE release, and the activation of
glutamate receptors reduces NE levels in the rat hippocampus
(104). An increase in
glutamatergic activity in the entorhinal cortex leads to the
induction of seizures. However, the administration of NE blocks
seizure activity in this area (105). NE increases epileptiform activity
in the hippocampal dentate gyrus (DG) through
N-methyl-D-aspartate (NMDA) receptor activation (106). A significant downregulation in
β1-ARs sensitivity in the DG can reduce the stimulating
effect of NE and may thus prevent seizures (105). Furthermore, the epileptic seizures
observed in transgenic mice overexpressing α1B-AR are
considered to result from an increased NMDA receptor number via
α1B-ARs (107).
There is ample evidence to suggest that the
endogenous neuromediator, NE, is involved in the modulation of
different types of epileptic seizures. Depending on the activated
AR subtype and brain region, NE sometimes has an anti-convulsant
and sometimes a convulsant effect. In addition, NE may modulate
seizures through affecting various neurotransmitter systems,
particularly GABA and glutamate, or voltage-gated Ca2+
and/or K+ channels. The seizure activity control
activity of NE may be impaired in some cases of increased
susceptibility to seizures, such as exposure to high levels of NE
due to stress. The results of various studies demonstrated that
abnormal increases or decreases in NE levels in the brain may cause
an impairment in NE-related functions, which may contribute to an
increased seizure susceptibility. In conclusion, recent data
indicate that the activation of α1-, α2- and
β2-AR subtypes with selective receptor agonists produces
anticonvulsant effects in epileptic seizures. Fully elucidating the
effects of AR subtypes on epileptic seizures may be an important
target for the pharmacological treatment of epilepsy.
Not applicable.
Funding: No funding was received.
Not applicable.
The author EO confirms being the sole contributor of
this work. EO conceived and designed the study, and wrote and
edited the manuscript. EO has read and approved the final
manuscript for publication. Data authentication is not
applicable.
Not applicable.
Not applicable.
The author declares that he has no competing
interests.
|
1
|
Fisher RS, van Emde Boas W, Blume W, Elger
C, Genton P, Lee P and Engel J Jr: Epileptic seizures and epilepsy:
Definitions proposed by the international league against epilepsy
(ILAE) and the international bureau for epilepsy (IBE). Epilepsia.
46:470–472. 2005.PubMed/NCBI View Article : Google Scholar
|
|
2
|
McHugh JC and Delanty N: Epidemiology and
classification of epilepsy: Gender comparisons. Int Rev Neurobiol.
83:11–26. 2008.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Weltha L, Reemmer J and Boison D: The role
of adenosine in epilepsy. Brain Res Bull. 151:46–54.
2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Berkovic SF and Scheffer IE: Febrile
seizures: Genetics and relationship to other epilepsy syndromes.
Curr Opin Neurol. 11:129–134. 1998.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Bence AK, Worthen DR, Stables JP and
Crooks PA: An in vivo evaluation of the antiseizure activity and
acute neurotoxicity of agmatine. Pharmacol Biochem Behav.
74:771–775. 2003.PubMed/NCBI View Article : Google Scholar
|
|
6
|
DiNuzzo M, Mangia S, Maraviglia B and
Giove F: Physiological bases of the K+ and the glutamate/GABA
hypotheses of epilepsy. Epilepsy Res. 108:995–1012. 2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Sahin B, Ozdemir E, Gumus E, Ergul M and
Taskiran AS: The 5-HT7 receptor antagonist SB-269970 alleviates
seizure activity and downregulates hippocampal c-Fos expression in
pentylenetetrazole-induced kindled rats. Neurol Res. 44:786–796.
2022.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Akyuz E, Doganyigit Z, Paudel YN, Koklu B,
Kaymak E, Villa C, Arulsamy A, Shaikh MF and Devinsky O:
Immunoreactivity of muscarinic acetylcholine M2 and serotonin
5-HT2B receptors, norepinephrine transporter and Kir channels in a
model of epilepsy. Life (Basel). 11(276)2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Chen C, Zhu T, Gong L, Hu Z, Wei H, Fan J,
Lin D, Wang X, Xu J, Dong X, et al: Trpm2 deficiency in microglia
attenuates neuroinflammation during epileptogenesis by upregulating
autophagy via the AMPK/mTOR pathway. Neurobiol Dis.
186(106273)2023.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kuang X, Chen S and Ye Q: The role of
histone deacetylases in NLRP3 inflammasomes-mediated epilepsy. Curr
Mol Med 2023: doi: 10.2174/1566524023666230731095431, 2023.
|
|
11
|
Rana A and Musto AE: The role of
inflammation in the development of epilepsy. J Neuroinflammation.
15(144)2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Gunes H, Ozdemir E and Arslan G: Coenzyme
Q10 increases absence seizures in WAG/Rij rats: The role of the
nitric oxide pathway. Epilepsy Res. 154:69–73. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Taskıran AS, Ozdemir E, Gumus E and Ergul
M: The effects of salmon calcitonin on epileptic seizures,
epileptogenesis, and postseizure hippocampal neuronal damage in
pentylenetetrazole-induced epilepsy model in rats. Epilepsy Behav.
13(107501)2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Strac DS, Pivac N, Smolders IJ, Fogel WA,
Deurwaerdere PD and Giovanni GD: Monoaminergic mechanisms in
epilepsy may offer innovative therapeutic opportunity for
monoaminergic multi-target drugs. Front Neurosci.
10(492)2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Giorgi FS, Pizzanelli C, Biagioni F, Murri
L and Fornai F: The role of norepinephrine in epilepsy: From the
bench to the bedside. Neurosci Biobehav Rev. 28:507–524.
2004.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Foote SL and Berridge CW: New developments
and future directions in understanding locus
coeruleus-Norepinephrine (LC-NE) function. Brain Res. 1709:81–84.
2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Amaral-Silva L and Santin JM: Molecular
profiling of CO2/pH-sensitive neurons in the locus coeruleus of
bullfrogs reveals overlapping noradrenergic and glutamatergic cell
identity. Comp Biochem Physiol A Mol Integr Physiol.
283(111453)2023.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Clough RW, Browning RA, Maring ML,
Statnick MA, Wang C and Jobe PC: Effects of intraventricular locus
coeruleus transplants on seizure severity in genetically
epilepsy-prone rats following depletion of brain norepinephrine. J
Neural Transplant Plast. 5:65–79. 1994.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Larsen LE, Caestecker S, Stevens L, van
Mierlo P, Carrette E, Boon P, Vonck K and Raedt R: Hippocampal
seizures differentially modulate locus coeruleus activity and
result in consistent time-locked release of noradrenaline in rat
hippocampus. Neurobiol Dis. 189(106355)2023.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Brawek B, Löffler M, Dooley DJ, Weyerbrock
A and Feuerstein TJ: Differential modulation of K(+)-evoked
(3)H-neurotransmitter release from human neocortex by gabapentin
and pregabalin. Naunyn Schmiedebergs Arch Pharmacol. 376:301–307.
2008.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Choi TY, Kwon JE, Durrance ES, Jo SH, Choi
SY and Kim KT: Melatonin inhibits voltage-sensitive Ca(2+)
channel-mediated neurotransmitter release. Brain Res. 4:34–42.
2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Briere R, Sherwin AL, Robitaille Y,
Olivier A, Quesney LF and Reader TA: Alpha-1 adrenoceptors are
decreased in human epileptic foci. Ann Neurol. 19:26–30.
1986.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Nicoletti F, Barbaccia ML, Iadarola MJ,
Pozzi O and Laird HE II: Abnormality ofalpha 1-adrenergic receptors
in the frontal cortex of epileptic rats. J Neurochem. 46:270–273.
1986.PubMed/NCBI View Article : Google Scholar
|
|
24
|
McIntyre DC and Edson N: Effect of
norepinephrine depletion on dorsal hippocampus kindling in rats.
Exp Neuron. 77:700–704. 1982.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Kokaia M, Bengzon J, Kalen P and Lindvall
O: Noradrenergic mechanisms in hippocampal kindling with rapidly
recurring seizures. Brain Res. 491:398–402. 1989.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Dailey JW and Naritoku DK: Antidepressants
and seizures: Clinical anecdotes overshadow neuroscience. Biochem
Pharmacol. 52:1323–1329. 1996.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Fitzgerald PJ: Is elevated norepinephrine
an etiological factor in some cases of epilepsy? Seizure.
19:311–318. 2010.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chen J, Liang H, Miao M, Su X, Yang F,
Thomsen RW, Yuan W and Li J: In utero beta-2-adrenergic agonists
exposure and risk of epilepsy: A Danish nationwide population-based
cohort study. Pharmacoepidemiol Drug Saf. 27:1200–1208.
2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Felippotti TT, dos Reis Ferreira CM, de
Freitas RL, de Oliveira RC, de Oliveira R, Paschoalin-Maurin T and
Coimbra NC: Paradoxical effect of noradrenaline-mediated
neurotransmission in the antinociceptive phenomenon that
accompanies tonic-clonic seizures: role of locus coeruleus neurons
and α(2)- and β-noradrenergic receptors. Epilepsy Behav. 22:165–77.
2011.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Hillman KL, Lei S, Doze VA and Porter JE:
Alpha-1A adrenergic receptor activation increases inhibitory tone
in CA1 hippocampus. Epilepsy Res. 84:97–109. 2009.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Pizzanelli C, Lazzeri G, Fulceri F, Giorgi
FS, Pasquali L, Cifelli G, Murri L and Fornai F: Lack of alpha
1b-adrenergic receptor protects against epileptic seizures.
Epilepsia. 50 (Suppl 1):S59–S64. 2009.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Shafaroodi H, Moezi L, Bahremand A and
Dehpour AR: The role of α2-adrenoceptors in the anti-convulsant
effects of cannabinoids on pentylenetetrazole-induced seizure
threshold in mice. Eur J Pharmacol. 714:1–6. 2013.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Shouse MN, Scordato JC, Farber PR and de
Lanerolle N: The alpha2 adrenoreceptor agonist clonidine suppresses
evoked and spontaneous seizures, whereas the alpha2 adrenoreceptor
antagonist idazoxan promotes seizures in amygdala-kindled kittens.
Brain Res. 1137:58–68. 2007.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Fletcher A and Forster EA: A proconvulsant
action of selective alpha 2-adrenoceptor antagonists. Eur J
Pharmacol. 151:27–34. 1988.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Payandemehr B, Bahremand A, Ebrahimi A,
Nasrabady SE, Rahimian R, Bahremand T, Sharifzadeh M and Dehpour
AR: Protective effects of lithium chloride on seizure
susceptibility: Involvement of α2-adrenoceptor. Pharmacol Biochem
Behav. 133:37–42. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Moezi L, Mansoori E, Niknahad H and
Shafaroodi H: The role of alpha-2 adrenoceptors in the
anticonvulsant effects of adenosine on pentylenetetrazole-induced
seizure threshold in mice. Pharmacol Biochem Behav. 126:36–42.
2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Abraham PA, Xing G, Zhang L, Yu EZ, Post
R, Gamble EH and Li H: beta1- and beta2-adrenoceptor induced
synaptic facilitation in rat basolateral amygdala. Brain Res.
1209:65–73. 2008.PubMed/NCBI View Article : Google Scholar
|
|
38
|
McIntyre DC and Roberts DCS: Long-term
reduction in beta-adrenergic receptor binding after amygdala
kindling in rats. Exp Neurol. 82:17–24. 1983.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Philipp M, Brede M and Hein L:
Physiological significance of alpha(2)-adrenergic receptor subtype
diversity: One receptor is not enough. Am J Physiol Regul Integr
Comp Physiol. 283:R287–R295. 2002.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wu Y, Zeng L and Zhao S: Ligands of
adrenergic receptors: A structural point of view. Biomolecules.
11(936)2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Perez DM: α1-Adrenergic receptors in
neurotransmission, synaptic plasticity, and cognition. Front
Pharmacol. 11(581098)2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Cavalli A, Lattion AL, Hummler E, Nenniger
M, Pedrazzini T, Aubert JF, Michel MC, Yang M, Lembo G, Vecchione
C, et al: Decreased blood pressure response in mice deficient of
the alpha1b adrenergic receptor. Proc Natl Acad Sci USA.
94:11589–11594. 1997.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Graham RM, Perez DM, Hwa J and Piascik MT:
alpha1-adrenergic receptor subtypes: Molecular structure, function,
and signaling. Circ Res. 78:737–749. 1996.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Perez DM and Doze VA: Cardiac and
neuroprotection regulated by α(1)-adrenergic receptor Subtypes. J
Recept Signal Transduct Res. 31:98–110. 2011.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Papay R, Gaivin R, Jha A, McCune DF,
McGrath JC, Rodrigo MC, Simpson PC, Doze VA and Perez DM:
Localization of the mouse alpha1A-adrenergic receptor (AR) in the
brain: Alpha1aar is expressed in neurons, GABAergic interneurons,
and NG2 oligodendrocyte progenitors. J Comp Neurol. 497:209–222.
2006.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Gupta MK, Papay RS, Jurgens CW, Gaivin RJ,
Shi T, Doze VA and Perez DM: Alpha1-Adrenergic receptors regulate
neurogenesis and gliogenesis. Mol Pharmacol. 76:314–326.
2009.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Trendelenburg AU, Sutej I, Wahl CA,
Molderings GJ, Rump LC and Starke K: A re-investigation of
questionable subclassifications of presynaptic α2-autoreceptors:
Rat vena cava, rat atria, human kidney and guinea-pig urethra.
Naunyn Schmiedebergs Arch Pharmacol. 356:721–737. 1997.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Rump CL, Bohmann C, Schaible U, Schöllhorn
J and Limberger N: Alpha 2C-adrenoceptor-modulated release of
noradrenaline in human right atrium. Br J Pharmacol. 116:2617–2624.
1995.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Brodde O: Beta-1 and beta-2 adrenoceptor
polymorphisms: Functional importance, impact on cardiovascular
diseases and drug responses. Pharmacol Ther. 117:1–29.
2008.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Leineweber K and Heusch G: Beta 1- and
beta 2-adrenoceptor polymorphisms and cardiovascular diseases. Br J
Pharmacol. 158:61–69. 2009.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Kume H, Nishiyama O, Isoya T, Higashimoto
Y, Tohda Y and Noda Y: Involvement of allosteric effect and
KCa channels in crosstalk between
β2-adrenergic and muscarinic M2 receptors in
airway smooth muscle. Int J Mol Sci. 19(1999)2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Sawa M and Harada H: Recent developments
in the design of orally bioavailable beta3-adrenergic receptor
agonists. Curr Med Chem. 13:25–37. 2006.PubMed/NCBI
|
|
53
|
Ferrer-Lorente R, Cabot C, Fernández-López
JA and Alemany M: Combined effects of oleoyl-estrone and a
β3-adrenergic agonist (CL316,243) on lipid stores of diet-induced
overweight male Wistar rats. Life Sci. 77:2051–2058.
2005.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Gundlach AL, Burazin TC, Jenkins TA and
Berkovic SF: Spatiotemporal alterations of central alpha
1-adrenergic receptor binding sites following amygdaloid kindling
seizures in the rat: Autoradiographic studies using (3H)prazosin.
Brain Res. 672:214–227. 1995.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Jazrawi SP and Horton RW: Brain
adrenoceptor binding sites in mice susceptible (DBA/2J) and
resistant (C57 Bl/6) to audiogenic seizures. J Neurochem.
47:173–177. 1986.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Kulik A, Haentzsch A, Lückermann M,
Reichelt W and Ballanyi K: Neuron-glia signaling via alpha(1)
adrenoceptor-mediated Ca(2+) release in Bergmann glialcells in
situ. J Neurosci. 19:8401–8408. 1999.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Terakado M: Adrenergic regulation of GABA
release from presynaptic terminals in rat cerebral cortex. J Oral
Sci. 56:49–57. 2014.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Rutecki PA: Noradrenergic modulation of
epileptiform activity in the hippocampus. Epilepsy Res. 20:125–136.
1995.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Jurgens CWD, Knudson CA, Carr PA, Perez DM
and Doze VA: a1 Adrenergic receptor regulation of interneuron
function. FASEB J. 23 (Suppl 946)(4)2009.
|
|
60
|
Knudson CA, Carr PA, Perez DM and Doze VA:
Alpha-1A adrenergic receptor overexpression protects hippocampal
interneurons. FASEB J. 21(A1209)2007.
|
|
61
|
Zuscik MJ, Sands S, Ross SA, Waugh DJ,
Gaivin RJ, Morilak D and Perez DM: Overexpression of the
alpha1B-adrenergic receptor causes apoptotic neurodegeneration:
Multiple system atrophy. Nat Med. 6:1388–1394. 2000.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Kruse SW, Dayton KG, Purnell BS, Rosner JI
and Buchanan GF: Effect of monoamine reuptake inhibition and α1
blockade on respiratory arrest and death following
electroshock-induced seizures in mice. Epilepsia. 60:495–507.
2019.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Kunieda T, Zuscik MJ, Boongird A, Perez
DM, Lüders HO and Najm IM: Systemic overexpression of the alpha
1B-adrenergic receptor in mice: An animal model of epilepsy.
Epilepsia. 43:1324–1329. 2002.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Chen CR, Qu WM, Qiu MH, Xu XH, Yao MH,
Urade Y and Huang ZL: Modafinil exerts a dose-dependent
antiepileptic effect mediated by adrenergic alpha1 and
histaminergic H1 receptors in mice. Neuropharmacology. 53:534–541.
2007.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Niitani K, Ito S, Wada S, Izumi S,
Nishitani N, Deyama S and Kaneda K: Noradrenergic stimulation of α1
adrenoceptors in the medial prefrontal cortex mediates acute
stress-induced facilitation of seizures in mice. Sci Rep.
19(8089)2023.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Ciltas AC, Ozdemir E, Gumus E, Taskiran
AS, Gunes H and Arslan G: The anticonvulsant effects of alpha-2
adrenoceptor agonist dexmedetomidine on pentylenetetrazole-induced
seizures in rats. Neurochem Res. 47:305–314. 2022.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Nissinen J, Andrade P, Natunen T, Hiltunen
M, Malm T, Kanninen K, Soares JI, Shatillo O, Sallinen J,
Ndode-Ekane XE and Pitkänen A: Disease-modifying effect of
atipamezole in a model of post-traumatic epilepsy. Epilepsy Res.
136:18–34. 2017.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Jurgens CW, Hammad HM, Lichter JA, Boese
SJ, Nelson BW, Goldenstein BL, Davis KL, Xu K, Hillman KL, Porter
JE and Doze VA: Alpha2A adrenergic receptor activation inhibits
epileptiform activity in the rat hippocampal CA3 region. Mol
Pharmacol. 71:1572–1581. 2007.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Szot P, Lester M, Laughlin ML, Palmiter
RD, Liles LC and Weinshenker D: The anticonvulsant and
proconvulsant effects of alpha2-adrenoreceptor agonists are
mediated by distinct populations of alpha2A-adrenoreceptors.
Neuroscience. 126:795–803. 2004.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Yavuz M, Aydın B, Çarçak N, Akman Ö,
Yananlı HR and Onat F: Atipamezole, a specific α2A antagonist,
suppresses spike-and-wave discharges and alters
Ca2+/calmodulin-dependent protein kinase II in the
thalamus of genetic absence epilepsy rats. Epilepsia. 61:2825–2835.
2020.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Ferraro L, Tanganelli S, Calo G, Antonelli
T, Fabrizi A, Acciarri N, Bianchi C, Beani L and Simonato M:
Noradrenergic modulation of gamma-aminobutyric acid outflow from
the human cerebral cortex. Brain Res. 629:103–108. 1993.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Louis WJ, Papanicolaou J, Summers RJ and
Vajda FJ: Role of central beta-adrenoceptors in the control of
pentylenetetrazol-induced convulsions in rats. Br J Pharmacol.
75:441–446. 1982.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Nakamura T, Oda Y, Takahashi R, Tanaka K,
Hase I and Asada A: Propranolol increases the threshold for
lidocaine-induced convulsions in awake rats: A direct effect on the
brain. Anesth Analg. 106:1450–1455. 2008.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Santana N and Artigas F: Laminar and
cellular distribution of monoamine receptors in rat medial
prefrontal cortex. Front Neuroanat. 11:1–13. 2017.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Luo F, Tang H and Cheng ZY: Stimulation of
α1-adrenoceptors facilitates GABAergic transmission onto pyramidal
neurons in the medial prefrontal cortex. Neuroscience. 300:63–74.
2015.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Hillman KL, Knudson CA, Carr PA, Doze VA
and Porter JE: Adrenergic receptor characterization of CA1
hippocampal neurons using real time single cell RT-PCR. Brain Res
Mol Brain Res. 139:267–276. 2005.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Sapa J, Zygmunt M, Kulig K, Malawska B,
Dudek M, Filipek B, Bednarski M, Kusak A and Nowak G: Evaluation of
anticonvulsant activity of novel pyrrolidin-2-one derivatives.
Pharmacol Rep. 66:708–711. 2014.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Clinckers R, Zgavc T, Vermoesen K, Meurs
A, Michotte Y and Smolders I: Pharmacological and neurochemical
characterization of the involvement of hippocampal adrenoreceptor
subtypes in the modulation of acute limbic seizures. J Neurochem.
115:1595–1607. 2010.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Gellman RL, Kallianos JA and McNamara JO:
Alpha-2 receptors mediateendogenous noradrenergic suppression of
kindling development. J Pharmacol Exp Ther. 241:891–898.
1987.PubMed/NCBI
|
|
80
|
Amabeoku GJ: The involvement of
noradrenaline, 5-hydroxytryptamine and acetylcholine in
imipramine-induced seizures in mice. Experientia. 49:859–864.
1993.PubMed/NCBI View Article : Google Scholar
|
|
81
|
MacDonald E, Kobilka BK and Scheinin M:
Gene targeting-homing in on alpha 2-adrenoceptor-subtype function.
Trends Pharmacol Sci. 18:211–219. 1997.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Weinshenker D, Szot P, Miller NS and
Palmiter RD: Alpha1 and beta2 adrenoreceptor agonists inhibit
pentylenetetrazole-induced seizures in mice lacking norepinephrine.
J Pharmacol Exp Ther. 298:1042–1048. 2001.PubMed/NCBI
|
|
83
|
Xiao Z, Deng PY, Rojanathammanee L, Yang
C, Grisanti L, Permpoonputtana K, Weinshenker D, Doze VA, Porter JE
and Lei S: Noradrenergic depression of neuronal excitability in the
entorhinal cortex via activation of TREK-2K+ channels. J Biol Chem.
284:10980–10991. 2009.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Sitnikova E, Pupikina M and Rutskova E:
Alpha2 adrenergic modulation of spike-wave epilepsy: Experimental
study of pro-epileptic and sedative effects of dexmedetomidine. Int
J Mol Sci. 24(9445)2023.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Biggane JP, Xu K, Goldenstein BL, Davis
KL, Luger EJ, Davis BA, Jurgens CWD, Perez DM, Porter JE and Doze
VA: Pharmacological characterization of the α2A-adrenergic receptor
inhibiting rat hippocampal CA3 epileptiform activity: Comparison of
ligand efficacy and potency. J Recept Signal Transduct Res.
42:580–587. 2022.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Ahmadirad N, Fathollahi Y, Janahmadi M,
Ghasemi Z, Shojaei A, Rezaei M, Barkley V and Mirnajafi-Zadeh J:
The role of α adrenergic receptors in mediating the inhibitory
effect of electrical brain stimulation on epileptiform activity in
rat hippocampal slices. Brain Res. 1765(147492)2021.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Rezaei M, Ahmadirad N, Ghasemi Z, Shojaei
A, Raoufy MR, Barkley V, Fathollahi Y and Mirnajafi-Zadeh J: Alpha
adrenergic receptors have role in the inhibitory effect of
electrical low frequency stimulation on epileptiform activity in
rats. Int J Neurosci. 133:496–504. 2023.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Wu HQ, Tullii M, Samanin R and Vezzani A:
Norepinephrine modulates seizures induced by quinolinic acid in
rats: Selective and distinct roles of alpha adrenoceptor subtypes.
Eur J Pharmacol. 138:309–318. 1987.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Eason MG, Kurose H, Holt BD, Raymond JR
and Liggett SB: Simultaneous coupling of alpha 2-adrenergic
receptors to two G-proteins with opposing effects.
Subtype-selective coupling of alpha 2C10, alpha 2C4, and alpha 2C2
adrenergic receptors to Gi and Gs. J Biol Chem. 267:15795–15801.
1992.PubMed/NCBI
|
|
90
|
Atzori M, Cuevas-Olguin R, Esquivel-Rendon
E, Garcia-Oscos F, Salgado-Delgado RC, Saderi N, Miranda-Morales M,
Treviño M, Pineda JC and Salgado H: Locus ceruleus norepinephrine
release: A central regulator of CNS spatio-temporal activation.
Front Synaptic Neurosci. 8(25)2016.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Świąder M, Zakrocka I, Świąder K, Zawadzki
A, Łuszczki JJ, Czuczwar SJ and Munir D: Influence of salbutamol on
the anticonvulsant potency of the antiepileptic drugs in the
maximal electroshock-induced seizures in mice. Pharmacol Rep.
71:466–472. 2019.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Gross RA and Ferrendelli JA: Relationships
between norepinephrine and cyclic nucleotides in brain and seizure
activity. Neuropharmacology. 21:655–661. 1982.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Anlezark G, Horton R and Meldrum B: The
anticonvulsant action of the (-)- and (+)-enantiomers of
propranolol. J Pharm Pharmacol. 31:482–483. 1979.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Levy A, Ngai SH, Finck AD, Kawashima K and
Spector S: Disposition of propranolol isomers in mice. Eur J
Pharmacol. 40:93–100. 1976.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Fischer W: Anticonvulsant profile and
mechanism of action of propranolol and its two enantiomers.
Seizure. 11:285–302. 2002.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Mueller AL and Dunwiddie TV:
Anticonvulsant and proconvulsant actions of alpha- and
beta-noradrenergic agonists on epileptiform activity in rat
hippocampus in vitro. Epilepsia. 24:57–64. 1983.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Lipski WJ and Grace AA: Activation and
inhibition of neurons in the hippocampal ventral subiculum by
norepinephrine and locus coeruleus stimulation.
Neuropsychopharmacology. 38:285–292. 2013.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Fassio A, Rossi F, Bonanno G and Raiteri
M: GABA induces norepinephrine exocytosis from hippocampal
noradrenergic axon terminals by a dual mechanism involving
different voltage-sensitive calcium channels. J Neurosci Res.
57:324–331. 1999.PubMed/NCBI
|
|
99
|
Tully K, Li Y, Tsvetkov E and Bolshakov
VY: Norepinephrine enables the induction of associative long-term
potentiation at thalamo-amygdala synapses. Proc Natl Acad Sci USA.
104:14146–14150. 2007.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Gellman RL and Aghajanian GK: Pyramidal
cells in piriform cortex receive a convergence of inputs from
monoamine activated GABAergic interneurons. Brain Res. 600:63–73.
1993.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Bergles DE, Doze VA, Madison DV and Smith
SJ: Excitatory actions of norepinephrine on multiple classes of
hippocampal CA1 interneurons. J Neurosci. 16:572–585.
1996.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Braga MF, Aroniadou-Anderjaska V, Manion
ST, Hough CJ and Li H: Stress impairs alpha(1A)
adrenoceptor-mediated noradrenergic facilitation of GABAergic
transmission in the basolateral amygdala. Neuropsychopharmacology.
29:45–58. 2004.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Prager EM, Bergstrom HC, Wynn GH and Braga
MFM: The basolateral amygdala -γ aminobutyric system in health and
disease. J Neurosci Res. 94:548–567. 2016.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Dazzi L, Matzeu A and Biggio G: Role of
ionotropic glutamate receptors in the regulation of hippocampal
norepinephrine output in vivo. Brain Res. 1386:41–49.
2011.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Stanton PK: Noradrenergic modulation of
epileptiform bursting and synaptic plasticity in the dentate gyrus.
Epilepsy Res. 7:135–150. 1992.PubMed/NCBI
|
|
106
|
Stanton PK, Jones RS, Mody I and Heinemann
U: Epileptiform activity induced by lowering extracellular (Mg2+)
in combined hippocampal-entorhinal cortex slices: Modulation by
receptors for norepinephrine and N-methyl-D-aspartate. Epilepsy
Res. 1:53–62. 1987.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Paladini CA, Fiorillo CD, Morikawa H and
Williams JT: Amphetamine selectively blocks inhibitory glutamate
transmission in dopamine neurons. Nat Neurosci. 4:275–281.
2001.PubMed/NCBI View
Article : Google Scholar
|