Introduction
One of the major prognostic determinants of cancer
patients is metastasis. The process of metastasis involves multiple
steps (1). A growing body of
evidence supports crucial roles for cell-surface carbohydrates
during the process of metastasis. Cell-surface carbohydrates
presented by glycoproteins are classified in accordance with their
linkage to proteins and comprise N-glycans
[N-acetylglucosamine (GlcNAc) to asparagine] and
O-glycans [N-acetylgalactosamine (GalNAc) to serine
(Ser) or threonine (Thr)]. It has been reported that
N-glycans are involved in several steps of the metastatic
process (2), but the roles of
O-glycans remain unclear. We have concentrated our efforts
on understanding the roles of O-glycans in tumor
metastasis.
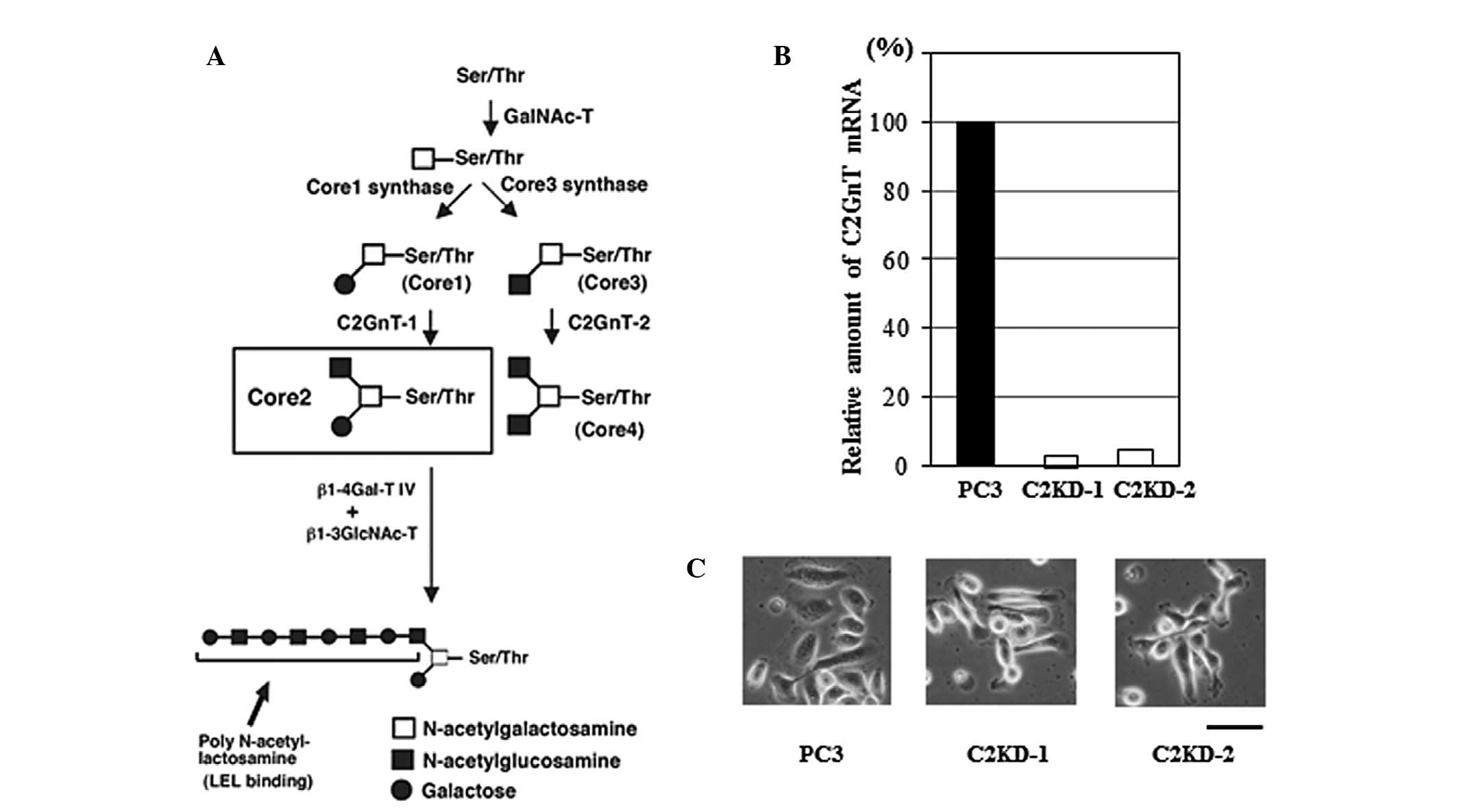
Fig. 1A shows the
biosynthesis pathway of various types of core structures of
O-glycans. GalNAc is transferred to Ser and Thr residues in
the polypeptide, and then GalNAcα1-Ser/Thr may be extended with
various carbohydrates, including galactose (Gal), GlcNAc, fucose or
sialic acid. Depending on the carbohydrates added, four common
O-glycan core structures, core1 through core4, are expressed
in mammalian tissues (Fig. 1A). A
key enzyme for the formation of O-glycans containing an
GlcNAc branch linked to GalNAc (GlcNAcβ1-6GalNAc, core2 branch) is
designated core2 β-1,6-N-acetylglucosaminyltransferase
(C2GnT; Fig. 1A). The expression
of C2GnT is closely correlated with highly metastatic phenotypes of
several types of cancer (3–7).
The core2 branch is a scaffold for the subsequent
production of lactosamine disaccharide repeats, specifically
poly-N-acetyllactosamine (Galβ1-4GlcNAc)n, on
O-glycans in a wide variety of cells (Fig. 1A) (8). Our previous studies concerning
bladder cancer revealed that C2GnT-expressing bladder cancer
cell-surface glycoproteins carrying poly-N-acetyllactosamine
on their O-glycans play an important role in bladder cancer
metastasis by facilitating the evasion of NK cell immunity by the
cancer cells (7,9,10).
Prostate cancer is the most common cancer in males
and the second leading cause of male cancer mortality in the US and
Western world, mainly due to metastatic disease (11). Our previous histological analyses
of patient specimens revealed that the expression of C2GnT is
positively correlated with poor prognosis in prostate cancer
patients (6). We hypothesized that
C2GnT-overxpressing prostate cancer cells acquire the highly
metastatic phenotype by evading NK cell immunity. In this study, we
examined how C2GnT-expressing prostate cancer cells evade NK cell
immunity.
Materials and methods
Cells, reagents and antibodies
PC3, a highly metastatic prostate cancer cell line,
was obtained from American Type Culture Collection (ATCC; Manassas,
VA, USA). Cells were maintained in RPMI-1640 medium (Sigma-Aldrich,
St. Louis, MO, USA) supplemented with 10% fetal bovine serum (FBS)
(PAA Laboratories, Morningside, Australia). All biochemical
reagents were purchased from Sigma-Aldrich, unless otherwise noted.
Recombinant human tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL) was purchased from R&D Systems, Inc.
(Minneapolis, MN, USA). The following monoclonal antibodies to
human antigens were used: anti-MUC1 (VU4H5; Cell Signaling
Technology, Danvers, MA, USA) and phycoerythrin (PE)-labeled
anti-CD56 (BD Biosciences, San Jose, CA, USA). The following
polyclonal antibodies were used: anti-lysosome-associated membrane
glycoprotein 1 (anti-LAMP1; Lifespan Biosciences, Seattle, WA,
USA), anti-death receptor 4 (anti-DR4; Millipore, Temecula, CA,
USA) and anti-actin (Sigma-Aldrich).
Stable transfectants
PC3 cells express high levels of C2GnT. PC3 cells
with reduced expression levels of C2GnT were generated using shRNA
technology as previously described (7). An shRNA expression plasmid was
constructed using pBAsi-hU6 Neo DNA (Takara Bio Inc., Shiga,
Japan). The shRNA sequence for C2GnT-1 was: GATCCGAATCCTAGTAGT
GATATTTAGTGCTCCTGGTTGAATATCACTACTAGG ATTCTTTTTTA. The siRNA
sequence is underlined. The sequence of the control shRNA
containing the scrambled siRNA sequence was: GATCCGTCTTAATCGCGTATAAGG
CTAGTGCTCCTGGTTGGCCTTATACGCGATTAAGAC TTTTTTA. The shRNA
expression plasmids (knockdown and control constructs) were
introduced into PC3 cells using Lipofectamine 2000. Drug-resistant
colonies were selected in the presence of 200 μg/ml geneticin.
Total RNA was prepared from each drug-resistant cell line using a
FastPure RNA kit (Takara Bio Inc.). Quantitative real-time PCR was
performed to quantify the C2GnT mRNA level of each drug-resistant
cell line using a Prime Script RT-PCR kit and Thermal Cycler Dice
Real Time system (Takara Bio Inc.). The sequences of the primer set
used for C2GnT-1 were: 5′-GGATGTCACCTG GAATCAGCACTA-3′ and
5′-TTATCAGAGCTGCAACGG CATC-3′. The primers specific for
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were:
5′-ATGACTCTACCCACG GCAAG-3′ and 5′-CATACTCAGCACCAGCATCAC-3′ and
were used as the internal control. Two C2GnT-deficient clones
(C2KD-1 and -2) were chosen based on their reduced mRNA expression
levels. C2KD-1, C2KD-2 and one control clone (PC3) were used for
the assays described in the present study.
Western blotting
Total lysates of cancer cells were prepared by
solubilization in 50 mM Tris-HCl buffer, pH 7.5, containing 1%
IgepalCA-630, 150 mM NaCl and proteinase inhibitors. The lysates
were resolved by SDS-PAGE on an 8–16% gradient gel (Invitrogen Life
Technologies, Carlsbad, CA, USA) and transferred to a
polyvinylidene fluoride (PVDF) membrane. Western blotting analyses
were performed using specific primary antibodies and a horseradish
peroxidase-conjugated secondary antibody. Signals were visualized
using the ECL PLUS detection system (GE Healthcare, Amersham,
UK).
Immunoprecipitation
Lectin immunopreocipitation was performed as
previously described (7). For
lectin immunoprecipitation, lysates from prostate cancer cells were
incubated with Lycoperiscon esculentum (tomato) lectin
(LEL)-agarose (Vector Laboratories, Burlingame, CA, USA). The resin
binding the immune complex was eluted with 1X Laemmli SDS-PAGE
sample buffer.
Preparation of NK cells
Human primary NK cells were purified from human
peripheral blood mononuclear cells using an NK cell isolation kit
(Myltenyi Biotech, Auburn, CA, USA). The NK cells (1×106
cells/ml) were cultured for 3 days in RPMI-1640 medium supplemented
with 10% FBS in the presence of 1000 U/ml human recombinant IL-2
(Wako, Osaka, Japan).
Conjugate formation and granzyme B
secretion assay
Heterotypic cell conjugates were quantitatively
determined by a double fluorescence assay (12) with certain modifications. Prostate
cancer target cells (2×106 cells/ml) were tranfected
with a green fluorescence protein (GFP)-expression plasmid, pmaxGFP
(Lonza Walkersville, Inc., Walkersville, MD, USA). After 1 h
incubation of the GFP-expressing tumor cells with IL-2-activated NK
cells at 37°C, the cells were stained with PE-labeled anti-CD56
antibody and then the number of conjugates were counted using a
flow cytometer, FACScant II (BD Biosciences). Double-colored (green
and red) conjugates were calculated as a percentage of the total
GFP-positive cells. For the granzyme B secretion assay, the culture
supernatant was collected from the co-culture of the target cancer
cells with NK cells. The quantity of granzyme B in the supernatant
was assayed by measuring serine proteinase activity with a
colorimetric peptide substrate, IEPD-p-nitroanilide (Biomol
International, Plymouth Meeting, PA, USA) as previously described
(7). Specific granzyme B secretion
was expressed as a percentage of the total cellular enzyme activity
after subtracting the spontaneous release.
NK cytotoxicity assay
Cytotoxicity was measured using the Cytotox 96
Non-radioactive Cytotoxicity Assay kit (Promega Corporation,
Madison, MI, USA). The target cells (tumor cells) were incubated
with IL-2-activated NK cells for 4 h at 37°C. The release of
lactate dehydrogenase from the lysed target cells was measured. All
assays were performed in quadruplicate. Percentage cytotoxicity =
(experimental lactate dehydrogenase release - effector spontaneous
release - target spontaneous release)/(target maximum release -
target spontaneous release) × 100.
Cell viability assay
For the cell viability assay, the cells
(1×104 cells/well in 96-well plates) were incubated with
the indicated concentration of recombinant TRAIL at 37°C for the
indicated time. The cell viability was assayed using the Cell
Counting Kit-8 (Dojindo Laboratories, Kumamoto, Japan) according to
the manufacturer’s instructions.
Statistical analysis
We used the statistical program SPSS 12.0 (SPSS,
Chicago, IL, USA). Statistically significant differences were
determined using the Student’s t-test. P<0.05 was considered to
indicate a statistically significant result.
Results
Establishment of C2GnT-deficient prostate
cancer cells
C2GnT is responsible for the formation of core2
O-glycans (Fig. 1A). To
test our hypothesis, we investigated the role of core2
O-glycans in the evasion of NK cell immunity by
C2GnT-expressing prostate cancer. We used a prostate cancer cell
line, PC3, which is derived from a malignant and metastatic
prostate cancer and expresses C2GnT at a high level (13). We established two C2GnT-deficient
cell lines (designated C2KD-1 and C2KD-2) and one control cell line
(PC3) using PC3 cells. RT-PCR analysis of the C2GnT expression
showed that the C2GnT expression levels were markedly reduced in
the C2KD-1 and C2KD-2 cells compared with those in the PC3 cells
(Fig. 1B). These cells exhibited
no marked morphological differences (Fig. 1C). The results obtained for C2KD-1
and PC3 only are shown, since the two C2GnT-deficient clones
yielded almost identical results in all assays.
Core2 O-glycosylation of MUC1
We have previously demonstrated that patients with
C2GnT-expressing prostate cancer have a significantly shorter
survival time than patients with C2GnT-non-expressing prostate
cancer (6), suggesting that
C2GnT-expressing prostate cancer is highly metastatic. However, a
growing body of evidence indicates that O-glycans in
cell-surface mucins are important in numerous biological processes,
including the protection of epithelial cell surfaces, immune
response, cell adhesion, inflammation, tumorigenesis and tumor
metastasis (14). These
observations led us to postulate that core2 O-glycans
carried by mucins play an important role in prostate cancer
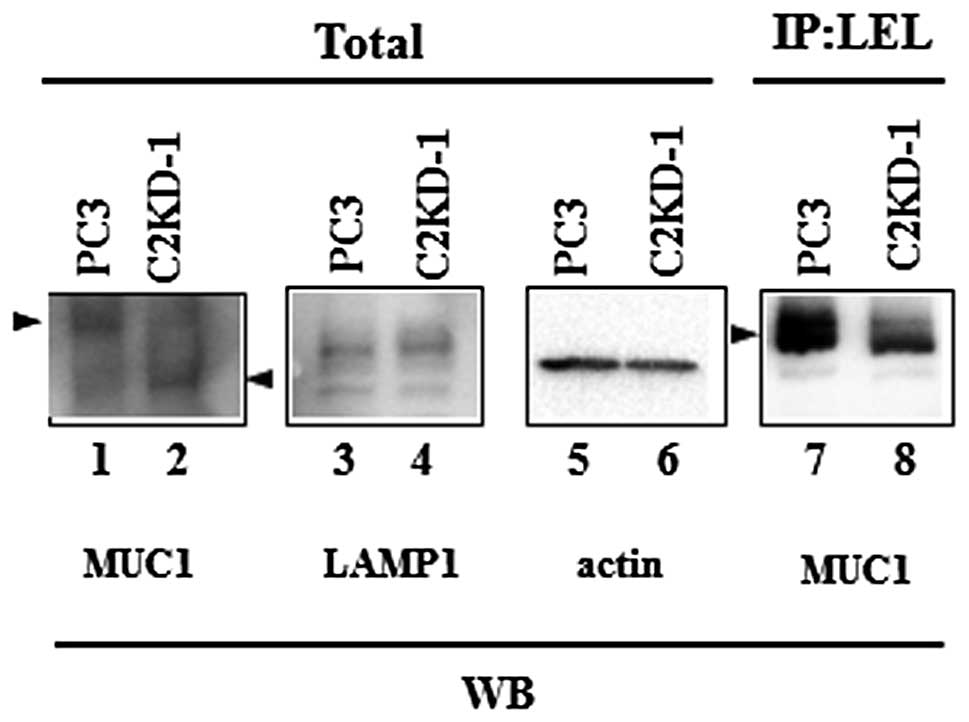
metastasis. We analyzed the MUC1 glycoproteins of the prostate
cancer cells for O-glycosylation by western blotting. The
MUC1 of the control PC3 cells exhibited a higher molecular weight
than the MUC1 from C2GnT-deficient cells (C2KD-1; Fig. 2, lanes 1 and 2). By contrast, we
observed no significant differences in the molecular weights of the
non-O-glycosylated cell-surface protein, LAMP1, between the
PC3 and C2KD-1 cells (Fig. 2,
lanes 3 and 4). These results indicate that MUC1 glycoproteins from
PC3 cells carry higher levels of core2 O-glycans than those
from C2KD-1 cells due to their higher expression levels of
C2GnT.
The core2 branch is a scaffold for the subsequent
production of lactosamine disaccharide repeats, specifically
poly-N-acetyllactosamine (Galβ1-4GlcNAc)n (Fig. 1A) (8). To determine whether MUC1 from PC3
cells carries poly-N-acetyllactosamine on its
O-glycans, we first excluded N-glycans from prostate
cancer cells by treatment with tunicamycin, an
N-glycosylation inhibitor, and then analyzed the cell
lysates by immunoprecipitation using LEL. LEL binds specifically to
poly-N-acetyllactosamines with at least three lactosamine
unit repeats. The LEL immunoprecipitates were subjected to western
blotting with anti-MUC1 antibody. MUC1 was detected in the LEL
immunoprecipitates from the PC3 cells, but the amount of MUC1
detected in the LEL immunoprecipitates from the C2KD-1 cells was
markedly reduced (Fig. 2, lanes 7
and 8). This result indicates that MUC1 from C2GnT-expressing PC3
cells carries a larger amount of poly-N-acetyllactosamine on
its core2 O-glycans than that from C2KD-1 cells.
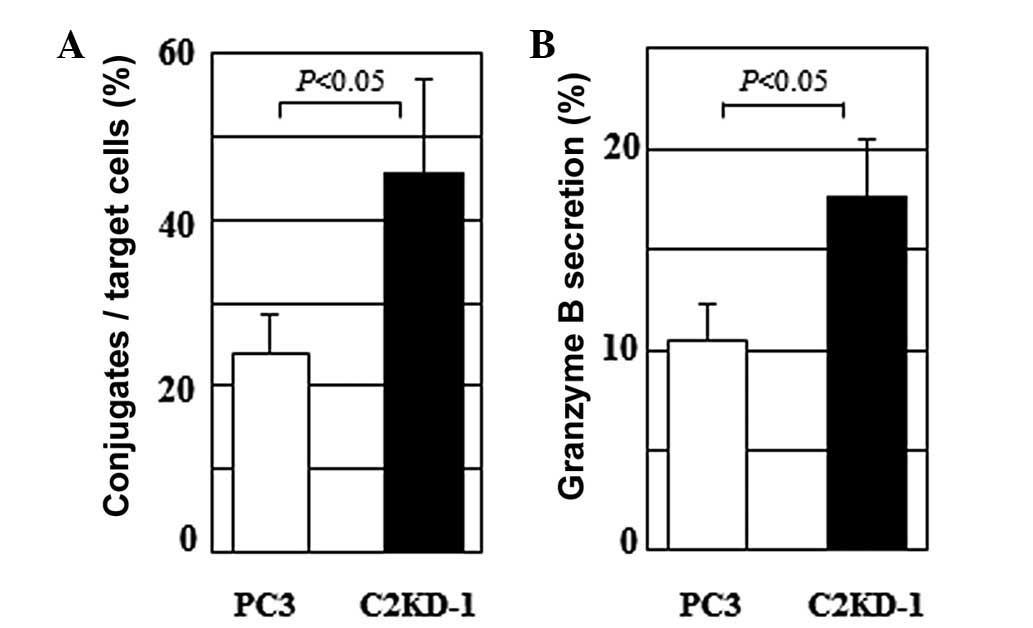
Effects of poly-N-acetyllactosamine on NK
cell functions
NK cells play a critical role in tumor rejection
responses in the host blood circulation. The attack on cancer cells
by NK cells is initiated by the NK cell-cancer cell interaction
mediated through the NK receptor-ligand interaction (15). To determine whether C2GnT
expression affects the NK cell-prostate cancer cell interaction, we
compared the conjugate formation of NK cells with PC3 and C2KD-1
cells. Following the incubation of the target cells with NK cells
for 1 h, the NK cells formed a significantly higher number of
conjugates with the C2KD-1 cells than with the PC3 cells (Fig. 3A). This result, taken together with
Fig. 2, suggests that in the
C2GnT-expressing cells (PC3), the poly-N-acetyllactosamine
moieties carried by the MUC1 core2 O-glycans reduce the
adhesive properties of MUC1 due to their bulkiness, resulting in
reduced conjugate formation between the PC3 and NK cells (Fig. 3A).
NK cells are activated through the NK cell-target
cell interaction and release their granular contents to kill the
target cells. The granular contents include perforin and granzyme B
which induce apoptosis of the target cells. We measured the
secretion of granzyme B stimulated by the NK cell-prostate cancer
cell interaction using an assay of the protease activity of
granzyme B in the co-culture supernatant. The C2GnT-expressing PC3
cells induced the secretion of significantly lower levels of
granzyme B than the C2GnT-deficient C2KD-1 cells (Fig. 3B). These results suggest that MUC1
glycoproteins carrying poly-N-acetyllactosamine attenuate
the NK cell-prostate cancer cell interaction, resulting in
decreased secretion of granzyme B (Fig. 3).
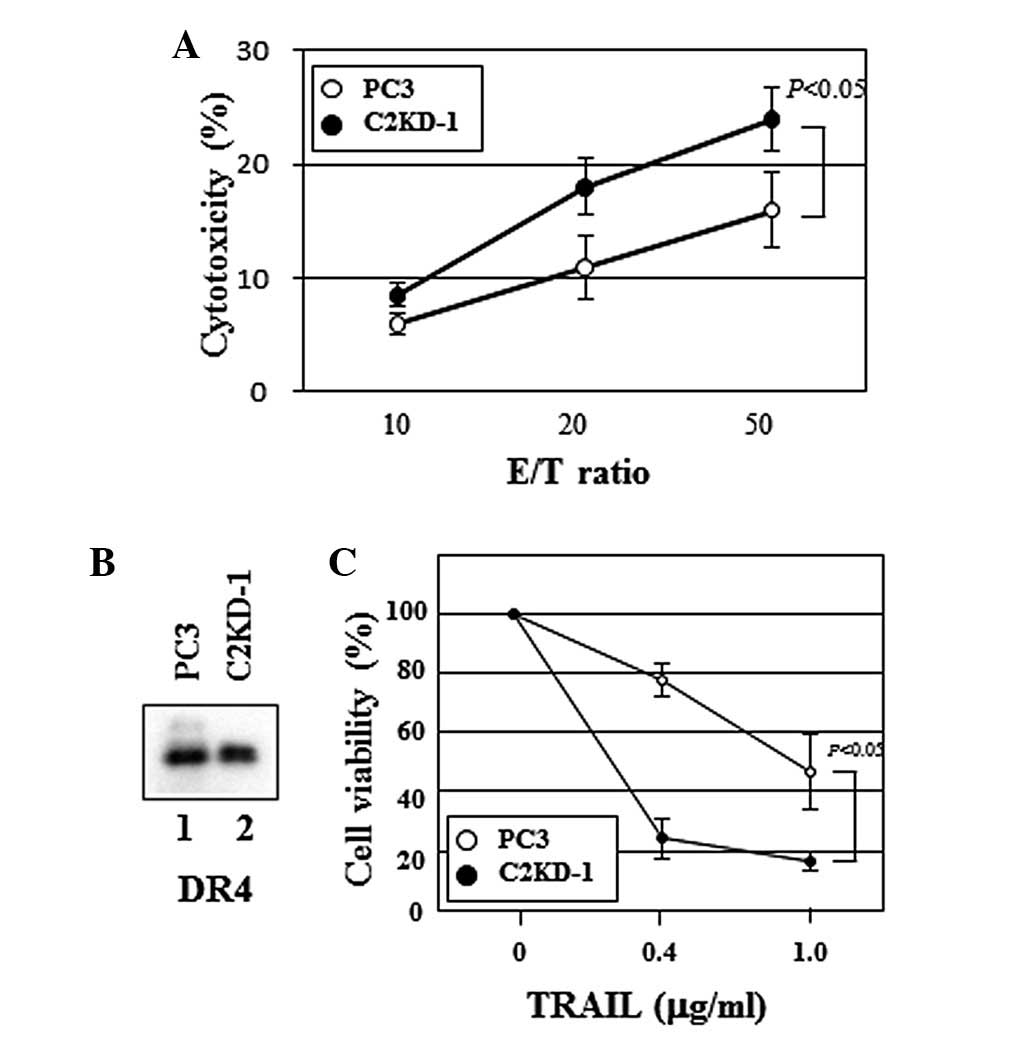
Effect of poly-N-acetyllactosamine on NK
cell cytotoxicity
We then questioned whether C2GnT expression by
prostate cancer cells affects the cytotoxic activity of NK cells,
since the interaction of NK cells with C2GnT-expressing prostate
cancer cells was impaired and the secretion of the target cell
apoptosis-inducing substance, granzyme B was reduced (Fig. 3). To address this, we assayed the
cytotoxicity of NK cells against prostate cancer cells. Fig. 4A shows that NK cells killed C2KD-1
cells more efficiently than PC3 cells (Fig. 4A), indicating that C2GnT-expressing
prostate cancer cells (PC3) were more resistant to NK cell
cytotoxicity than were C2GnT-deficient cells (C2KD-1).
NK cells use two main mechanisms to kill tumor
cells. One is that NK cells are activated through the NK cell
receptor-tumor ligand interaction to release cytotoxic granules,
including perforin and granzymes. The other is that NK cells kill
tumor cells using death ligands, including TRAIL (16). TRAIL induces target cell apoptosis
through its interaction with death receptors, including DR4, on the
surface of the target cell. We evaluated the expression of DR4 on
the prostate cancer cells by western blotting. We observed no
significant differences in the DR4 expression levels between PC3
and C2KD-1 (Fig. 4B, lanes 1 and
2). Unlike MUC1, no significant differences in the
O-glycosylation levels of DR4 between PC3 and C2KD-1 cells
were observed on western blots (Figs.
2 and 4B). To evaluate the
effect of C2GnT expression on the sensitivity of prostate cancer
cells to TRAIL, we measured TRAIL-induced cell death using soluble
recombinant TRAIL. In the presence of TRAIL, the viability of the
PC3 cells was significantly higher than that of the C2KD-1 cells
(Fig. 4C), indicating that
C2GnT-expressing prostate cancer cells are more resistant to TRAIL
than are C2GnT-deficient cells.
Discussion
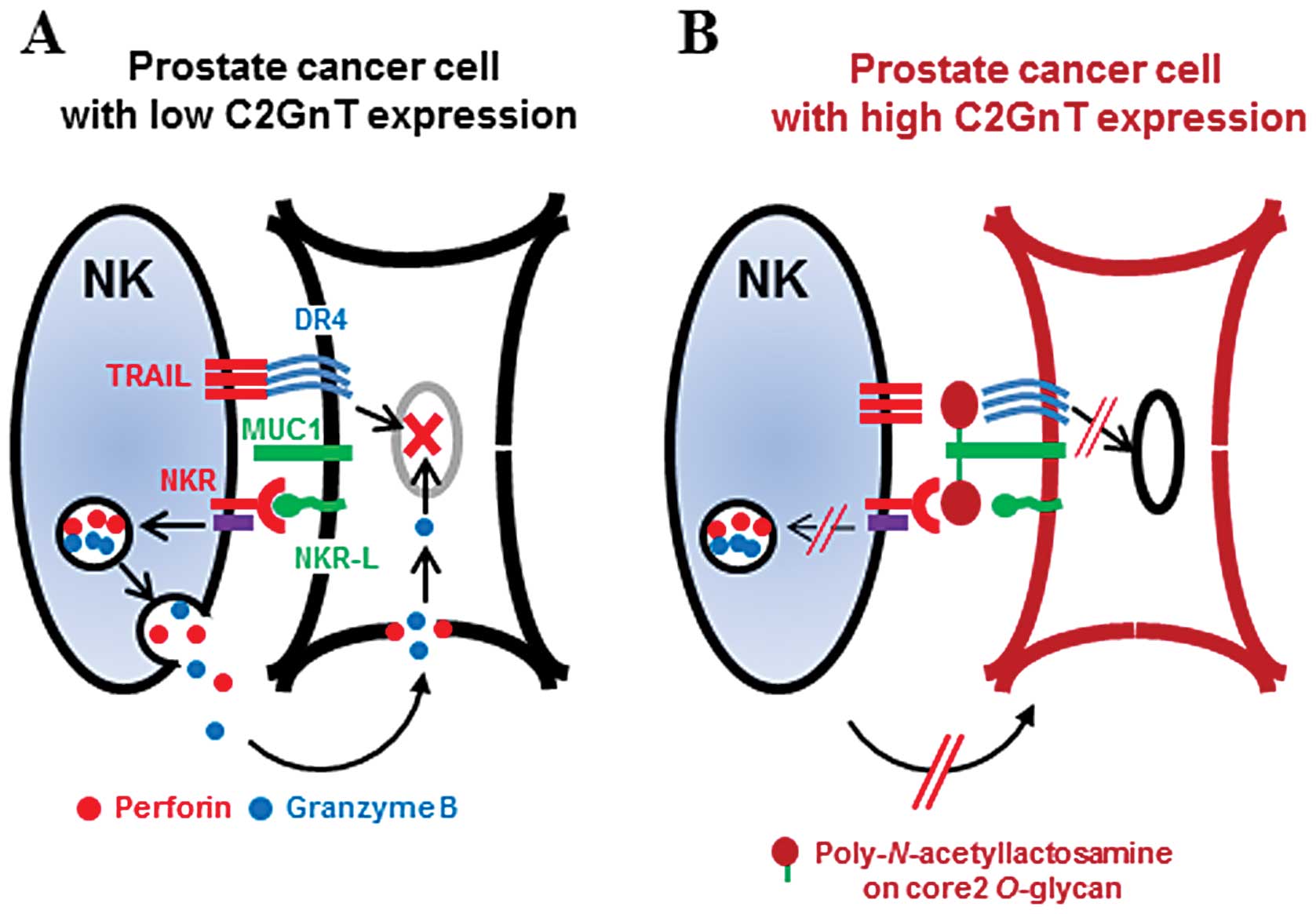
We have previously shown that MUC1 is modified by
poly-N-acetyllactosamine on its O-glycan residues in
C2GnT-expressing prostate cancer cells. MUC1 is significant in
adhesion as it is one of the molecules that extends most highly
above the cell surface (14). Our
results taken together with this observation suggest that the
modification of MUC1 in C2GnT-expressing prostate cancer cells with
bulky poly-N-acetyllactosamine moieties reduces their
adhesiveness, thereby attenuating the NK cell-cancer cell
interaction. The attenuated interaction results in decreased
degranulation by NK cells and reduction of the accessibility of
TRAIL to the death receptors (Fig.
5). These effects allow C2GnT-expressing prostate cancer cells
to evade NK cell immunity in the circulation. In our previous
study, when bladder tumor cells were intravenously injected into
nude mice, C2GnT-expressing tumor cells produced a greater number
of metastatic foci in the lungs than were produced by
C2GnT-non-expressing tumor cells (7). Our present data taken together with
the previous observations strongly suggest that this immune evasion
results in longer survival of C2GnT-expressing prostate cancer
cells in the host blood circulation, resulting in the promotion of
prostate cancer metastasis.
Wagner et al reported that tumor-cell
sensitivity to TRAIL was controlled by O-glycosylation of
death receptors (DR4 and DR5). In 22 out of 28 TRAIL-sensitive
cancer cell lines, the expression levels of a peptidyl
O-glycosyltransferase which catalyzes the initial step of
O-glycosylation were elevated (17). It was shown that
O-glycosylation of death receptors promoted TRAIL-stimulated
clustering of the receptors, mediating recruitment and activation
of the apoptosis-initiating protease, caspase-8. Although their
O-glycan structures were not analyzed,
O-glycosylation increased the TRAIL-sensitivity of the
cancer cells. The authors observed O-glycosylation of death
receptors on the western blots of 22 of the 28 TRAIL-sensitive cell
lines. However, no significant O-glycosylation of DR4 was
observed in the present study, suggesting that C2GnT-expressing
prostate cancer cells use a different mechanism to control cancer
cell-sensitivity to TRAIL from that reported by Wagner et
al.
Our investigation provides a new insight into the
roles of the carbohydrates carried by tumor cell-surface mucins in
tumor metastasis. Tumor cells take advantage of the functions of
mucins to maintain homeostasis and promote their survival in
variable conditions. Further analyses of the roles of tumor
cell-surface carbohydrates are likely to contribute to a better
understanding of the tumor metastasis process.
Acknowledgements
This study was supported by the grants-in-aid for
Scientific Research from the Japanese Society for the Promotion of
Science (22570131) (to S.T.) and (B22390301) (to C.O.), the
Ministry of Education, Culture, Sports, Science and Technology of
Japan (21791483) (to C.O.) and (21791484) (to T.K.), Japan Science
and Technology Agency (CREST) (to C.O.) and NIH grants P01CA71932
and R01CA3000 (to M.F.). This study was also supported by the
grant-in-aid from the Mizutani Foundation for Glycoscience (to
S.T.).
References
|
1
|
Gupta GP and Massagué J: Cancer
metastasis: building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fuster MM and Esko JD: The sweet and sour
of cancer: glycans as novel therapeutic targets. Nat Rev Cancer.
5:526–542. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yousefi S, Higgins E, Daoling Z,
Pollex-Krüger A, Hindsgaul O and Dennis JW: Increased
UDP-GlcNAc:Gal beta 1–3GaLNAc-R (GlcNAc to GaLNAc) beta-1,
6-N-acetylglucosaminyltransferase activity in metastatic
murine tumor cell lines. Control of polylactosamine synthesis. J
Biol Chem. 266:1772–1782. 1991.
|
|
4
|
Shimodaira K, Nakayama J, Nakamura N,
Hasebe O, Katsuyama T and Fukuda M: Carcinoma-associated expression
of core 2 beta-1,6-N-acetylglucosaminyltransferase gene in
human colorectal cancer: role of O-glycans in tumor
progression. Cancer Res. 57:5201–5206. 1997.PubMed/NCBI
|
|
5
|
Machida E, Nakayama J, Amano J and Fukuda
M: Clinicopathological significance of core 2
beta1,6-N-acetylglucosaminyltransferase messenger RNA
expressed in the pulmonary adenocarcinoma determined by in situ
hybridization. Cancer Res. 61:2226–2231. 2001.PubMed/NCBI
|
|
6
|
Hagisawa S, Ohyama C, Takahashi T, et al:
Expression of core 2 beta1,6-N-acetylglucosaminyltransferase
facilitates prostate cancer progression. Glycobiology.
15:1016–1024. 2005.PubMed/NCBI
|
|
7
|
Tsuboi S, Sutoh M, Hatakeyama S, et al: A
novel strategy for evasion of NK cell immunity by tumours
expressing core2 O-glycans. EMBO J. 30:3173–3185. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Maemura K and Fukuda M:
Poly-N-acetyllactosaminyl O-glycans attached to
leukosialin. The presence of sialyl Le(x) structures in
O-glycans. J Biol Chem. 267:24379–24386. 1992.PubMed/NCBI
|
|
9
|
Tsuboi S, Hatakeyama S, Ohyama C and
Fukuda M: Two opposing roles of O-glycans in tumor
metastasis. Trends Mol Med. 18:224–232. 2012. View Article : Google Scholar
|
|
10
|
Suzuki Y, Sutoh M, Hatakeyama S, et al:
MUC1 carrying core 2 O-glycans functions as a molecular
shield against NK cell attack, promoting bladder tumor metastasis.
Int J Oncol. 40:1831–1838. 2012.PubMed/NCBI
|
|
11
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar
|
|
12
|
van de Wiel-van Kemenade E, Ligtenberg MJ,
de Boer AJ, et al: Episialin (MUC1) inhibits cytotoxic
lymphocyte-target cell interaction. J Immunol. 151:767–776.
1993.PubMed/NCBI
|
|
13
|
Kaighn ME, Narayan KS, Ohnuki Y, Lechner
JF and Jones LW: Establishment and characterization of a human
prostatic carcinoma cell line (PC-3). Invest Urol. 17:16–23.
1979.PubMed/NCBI
|
|
14
|
Hollingsworth MA and Swanson BJ: Mucins in
cancer: protection and control of the cell surface. Nat Rev Cancer.
4:45–60. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lanier LL: Up on the tightrope: natural
killer cell activation and inhibition. Nature Immunology.
9:495–502. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cretney E, Shanker A, Yagita H, Smyth MJ
and Sayers TJ: TNF-related apoptosis-inducing ligand as a
therapeutic agent in autoimmunity and cancer. Immunol Cell Biol.
84:87–98. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wagner KW, Punnoose EA, Januario T, et al:
Death-receptor O-glycosylation controls tumor-cell
sensitivity to the proapoptotic ligand Apo2L/TRAIL. Nat Med.
13:1070–1077. 2007.PubMed/NCBI
|