Introduction
The morbidity and mortality of lung cancer has been
rapidly increasing over the past 50 years. The incidence of lung
cancer has accounted for 12% of all new cancer cases, and lung
cancers are the second most common malignancies in individuals
(1). Surgical intervention is the
most effective method to treat lung cancer; however, the recurrence
following surgery remains high. Moreover, the effect of traditional
radiation and chemical treatment on pulmonary carcinoma is
extremely limited. It is therefore necessary to urgently find a
novel method to treat lung cancers more efficiently.
Exosomes are lipid bilayer vesicles, 30–90 nm in
diameter, formed within late endocytic compartments and
physicochemically distinct from other secreted vesicles (2–4).
Exosomes may be released from various living cells, including tumor
cells, dendritic cells (DCs), platelets, cytotoxic T lymphocytes
(CTLs) and reticulocytes (2,5).
Exosomes derived from tumor cells (TEX) contain tumor antigens, and
a number of studies have indicated that exosomes may be used as an
effective vaccine to initiate antitumor immunity (6,7).
However, the immunogenicity of the tumor antigen in TEX is poor and
the efficiency of TEX to induce antitumor immunity requires further
improvement. One of the most significant characteristics of
exosomes is their suitability for artificial modification. It has
been reported that TEX from heat-stressed tumor cells or
super-antigen-anchored TEX are more immunogenic and may induce more
efficient tumor antigen-specific CTL responses (8,9). It
may be assumed that TEX is likely to be an attractive vaccine for
the prevention of tumors. TEX from heat-stressed 3LL lung tumor
cells are effective in the regression of the established lung
cancer in mice, which indicates that TEX are also effective in the
treatment of lung cancers.
CD40, a member of the tumor necrosis factor receptor
superfamily, is expressed on B cells, monocytes and DCs. CD40 is
significant in the improvement of immune responses following
ligation with its natural ligand, CD40 ligand (CD40)/CD154
(10). DCs are unique professional
antigen-presenting cells that prime naïve T cells. DCs are
considered to be the most attractive candidates in the activation
of immunity against cancer (11).
The activation of CD40 signaling in DCs may promote their
differentiation and maturation, and the secretion of
immunostimulatory cytokines, including interleukin (IL)-12, which
is significant in the activation of the antitumor T cell response
(12,13). However, CD40L is predominantly
expressed on activated T cells, but not on resting T cells
(14,15), which possibly restricts the
activation of DCs. It was hypothesized that TEX from CD40L
gene-modified 3LL cells are likely to be more efficient in the
activation of DCs and the subsequent antitumor T cell immunity.
In the present study, the antitumor effect of
exosomes derived from CD40L gene-modified 3LL tumor cells was
investigated. Recombinant murine CD40L adenovirus (Ad-CD40L) was
constructed, and then CD40L gene-modified 3LL tumor cells
(CD40L-EXO) from Ad-CD40L-infected 3LL lung cancer were isolated.
The CD40L-EXO were demonstrated to be more efficient in the
induction of mature DCs and IL-12 secretion than TEX from Ad-Lac
Z-infected (Lac Z-EXO) and uninfected 3LL tumor cells
(Control-EXO). CD40L-EXO were more efficient in improving the
ability of DCs to present antigens than Lac Z-EXO and Control-EXO.
Moreover, DCs pulsed with CD40L-EXO significantly increased tumor
antigen-specific CD4+ T cell proliferation and CTL
responses. Notably, CD40L-EXO revealed a more powerful antitumor
effect than Lac Z-EXO and Control-EXO in vivo.
Materials and methods
Reagents
CD4+ T cell isolation kit II was
purchased from Milteny Biotec (Bergisch Gladbach, Germany) and the
ELISA kit with IL-2, IL-12p70, TNF-α and interferon (INF)-γ and
phycoerythrin (PE)-conjugated antibodies against CD40 (1c10), MHCII
(M5/114.15.2), CD80 (16-10A1) and CD86 (D03.1) was obtained from
eBioscience (San Diego, CA, USA). The PE-conjugated antibodies
against CD40L (MR1) were obtained from BD Biosciences (San Diego,
CA, USA). The antibodies against HSP70 (F-3), CD63 (Y-8), CD9
(KMC8.8) and TSG101 (c-2) were purchased from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA). The lactose
dehydrogenase (LDH) assay kit was purchased from Promega
Corporation (Wisconsin, CA, USA). The aldehyde/sulfate latex beads
and the alamarBlue cell proliferation assay kit were from
Invitrogen Life Technologies (Carlsbad, CA, USA).
Cell lines and mice
The 3LL Lewis lung cancer and B16 melanoma cell
lines were obtained from the American Type Culture Collection
(Manassas, VA, USA). The 3LL and B16 cells were maintained in
RPMI-1640 media and Dulbecco’s modified Eagle’s medium (DMEM;
Hyclone, Rockford, IL, USA), respectively, supplemented with 10%
fetal calf serum (FCS; Hyclone). C57BL/6 mice were obtained from
Joint Ventures Sipper BK Experiment Animals Co. (Shanghai, China).
Female mice, 6–8 weeks old, were housed in a specific pathogen-free
facility. The experimental protocols were approved by the Animal
Care and Use Committee of the School of Medicine, Zhejiang
University (Hangzhou, China).
Construction of recombinant mouse
Ad-CD40L
Ad-CD40L was constructed using the AdMax™ system
(Microbix Biosystems, Mississauga, OR, Canada). Briefly, the DNA
fragment for murine CD40L was amplified from splenocytes of the
C57BL/6 mice by polymerase chain reaction using the following
specific primers: Sense: 5′-GCCGAATTCATGATA GAAACATACAGCCAA-3′ and
antisense: 5′-GACGTCGAC GAGTTTGAGTAAGCCAAAAGA-3′, synthesized by
Sangong Biotech Shanghai Co. Ltd. (Shanghai, China). The CD40L
sequence was then inserted into a pDC315 shuttle vector and
co-transfected with pBHGloxΔE1, 3Cre Ad backbone plasmid
(transfected with pBHGloxΔE1 and backbone plasmid) into HEK293
cells to generate Ad-CD40L. The generated Ad-CD40L was purified and
stored at −80°C.
Ad infection and exosome
purification
For Ad infection, the 3LL cells were infected with
Ad-CD40L or Ad-Lac Z for 24 h. The cells were then washed
intensively five times and incubated with fresh medium for 48 h.
The cell-culture supernatants were centrifuged at a series of
speeds (300 × g for 10 min, 1,200 × g for 20 min and 10,000 × g for
20 min) to remove the cell debris, the centrifuged supernatant was
aggregated through ultracentrifugation (100,000 × g for 1 h) and
the pellets were run on a 30 (1.13) to 45% (1.19 g/ml)
discontinuous sucrose density gradient (100,000 × g for 2 h) to
purify the exosomes. The middle layer was collected and subjected
to another ultracentrifugation to obtain the purified exosomes.
Electron microscopy
Exosomes were fixed in 4% paraformaldehyde, loaded
onto formvar carbon-coated electron microscopy grids (Phillips
Electronic Instruments, Mahwah, NJ, USA), contrasted and embedded
in a mixture of uranyl acetate and methylcellulose. Sections were
observed with a Philips Tecnai-10 transmission electron microscope
operating at 80 kV (Phillips Electronic Instruments).
Western blot analysis
In total, 10 μg exosomal proteins were separated by
a 10% SDS-polyacrylamide gel and transferred onto a polyvinylidene
difluoride membrane. Following incubation with anti-HSP70 (F-3),
CD63 (Y-8), CD9 (KMC8.8) and TSG101 (c-2) antibody and
corresponding horseradish peroxidase-coupled secondary antibody
sequentially, the specific band on the membrane was visualized with
a chemiluminescence kit (ECL detection kit, Amersham Bioscience,
Piscataway, NJ, USA).
DC generation and fluorescence-activated
cell sorting (FACS) analysis
Bone marrow cells from the C57BL/6 mice were
cultured in RPMI-1640 medium containing 10% FCS, 10 ng/ml
granulocyte macrophage-colony stimulating factor and 1 ng/ml IL-4
to generate bone marrow-derived DCs (BMDCs). Following culture
growth for a period of five days, 1×106/ml BMDCs were
stimulated with 1 or 10 μg/ml CD40L-EXO, LacZ-EXO and Control-EXO
or 1 μg/ml lipopolysaccharide (LPS) for 48 h. For FACS analysis of
the BMDCs, 1×106 BMDCs were incubated with the
corresponding PE-conjugated antibodies for 20 min.
For the FACS analysis of the exosomes, 20 μg
exosomes were incubated with 5 μl 4-μm diameter aldehyde/sulfate
latex beads for 15 min at room temperature in a 20-μl final volume,
followed by gentle agitation for 1 h in 1 ml phosphate-buffered
saline (PBS) and centrifugation at 3,000 × g for 5 min. The pellet
was blocked by incubation with 20 μl FCS for 30 min. Exosome-coated
beads were washed three times in PBS and then resuspended in 50 μl
PBS. The beads were incubated for 1 h with the corresponding
fluorescent antibodies. The cells and beads were analyzed by flow
cytometry using a FACS Calibur flow cytometer and CellQuest
software (Becton-Dickinson, Mountain View, CA, USA).
Cytokine assay
The levels of IL-12p70 and tumor necrosis factor
(TNF)-α from the exosome-stimulated BMDCs and the levels of IFN-γ
and IL-2 from the splenocytes of the exosome-immunized C57BL/6
tumor mice ex vivo were detected by ELISA kits according to
the manufacturer’s instructions.
Mixed lymphocyte reaction (MLR)
The BMDCs from the C57BL/6 mice were stimulated with
10 μg/ml CD40L-EXO, LacZ-EXO or Control-EXO, or 1 μg/ml LPS for 48
h. The BMDCs were then collected and inactivated by 50 μg/ml
mitomycin C for 30 min at 37°C. CD4+ T cells from the
splenocytes of the BALB/c mice were isolated using a
CD4+ T cell isolation kit II (Miltenyi Biotec, Surrey,
UK). BMDCs and CD4+ T cells were co-cultured at the
indicated ratio for 72 h. AlamarBlue was then added per well for an
additional culture for 24 h. The fluorescence intensities were
detected to determine the CD4+ T cell proliferation.
Antigen presentation assay
The C57BL/6 mice were immunized four times at an
interval of seven days by 100 μg 3LL cell lysates. At 35 days
post-immunization, the CD4+ T cells derived from the
splenocytes were isolated and co-cultured with syngeneic BMDCs that
had been pre-treated with 1 or 10 μg/ml exosomes for 48 h.
Following co-culture for 72 h, the proliferation of the
CD4+ T cells was also detected by the alamarBlue
assay.
Immunization and tumor challenge
The C57BL/6 mice were immunized subcutaneously
(s.c.) into the left flank by CD40L-EXO, LacZ-EXO, Control-EXO (10
μg/100 μl PBS/mouse) or 100 μl PBS 13 days before 3LL cell
challenge. The immunization was boosted three times on days seven,
nine and 11, respectively. On day 0, the immunized mice were
challenged with 1×106 3LL cells s.c. The tumor size was
measured and the survival rate of the tumor mice was recorded on
the indicated days. To induce IL-2 and IFN-γ, 5×106/ml
splenocytes from mice seven days after the last immunization were
stimulated with inactivated 3LL cells at a ratio of 10:1 for 48
h.
To investigate the therapeutic effect of CD40L-EXO,
the 3LL tumor model was established with 1×106 3LL cells
per mouse. Exosomes were then injected (10 μg/mouse) with on days
+7, +9, +11 and +13. The tumor size was measured and the survival
rate of the tumor mice was also recorded on the indicated days.
Cytotoxic assay of CTL
For the cytotoxic assay, splenocytes were obtained
from the immunized mice. The splenocytes were stimulated with
inactivated 3LL cells and with IL-2 (30 U/ml) for seven days. The
3LL cells were used as the specific target cells, while B16 cells
were used as the control target cells and pulsed splenocytes were
used as the effector cells. The ratios of effector to target were
12.5:1, 25:1 and 50:1. The cells were co-cultured for 4 h and then
the cytotoxic activity of the pulsed splenocytes was measured by
the LDH assay.
Statistical analysis
The statistical analysis was performed using one-way
analysis of variance. P<0.05 was used to indicate a
statistically significant difference.
Results
Characterization of CD40L-EXO
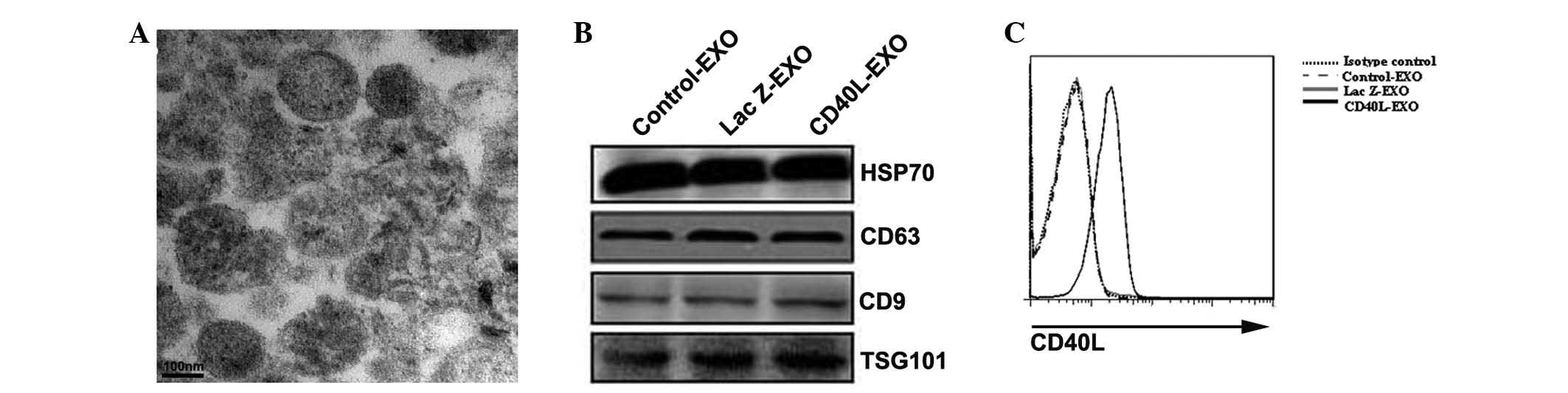
The morphology of the exosomes that were obtained
was examined first. Under electron microscopy, the exosomes
revealed the typical characteristics of a round morphology, with a
diameter of 30–90 nm (Fig. 1A).
Western blot analysis demonstrated that all the types of exosomes
contained the exosome-associated proteins, HSP70, TSG101, CD63 and
CD9 (Fig. 1B). In total, ~10 μg
exosomal proteins was routinely obtained from supernatants
containing 1×107 3LL cells. There was no marked
difference in the production of exosomes from the Ad-CD40L and
Ad-Lac Z-infected and uninfected 3LL cells. The CD40L was then
detected on each type of exosome by FACS. CD40L was only detected
on CD40L-EXO, but not Lac Z-EXO or Control-EXO (Fig. 1C). These results indicate that the
exosomes carrying CD40L are likely to possess a potent ability to
induce the activation of DCs were successfully isolated.
CD40L-EXO induce activation of BMDCs more
efficiently
Exosomes from heat-stressed and IL-18- or IL-2
gene-modified tumor cells were more efficient in promoting DC
maturation (8,16,17).
Considering the significance of CD40 signaling in the immune
function of DCs, the phenotype of DCs treated with CD40L-EXO was
examined first. Following stimulation with 10 μg of all the types
of exosomes for 48 h, the expression of MHC II, CD80, CD86 and CD40
on the BMDCs was markedly increased. However, the effect of
CD40L-EXO was strongest, which was comparable with the effect of
LPS (Fig. 2A). IL-12p70 and TNF-α
production in the culture supernatants of the BMDCs was markedly
enhanced by CD40L-EXO and was significantly higher than that of Lac
Z-EXO and Control-EXO (Fig. 2B).
In the MLR, the CD40L-EXO-, Lac Z-EXO- and Control-EXO-treated DCs
all induced T cell proliferation. However, DCs stimulated with
CD40-EXO were more effective in stimulating a proliferative
response than those with Lac Z-EXO and Control-EXO (Fig. 2C). Furthermore, it was also
determined whether DCs pulsed with CD40L-EXO, Lac Z-EXO and
Control-EXO had differential abilities to induce the activation of
tumor antigen-specific CD4+ T cells from mice immunized
with 3LL cell lysates. The proliferation of the primed
CD4+ T cells induced by the DCs treated with CD40L-EXO
was greater than those treated with Lac Z-EXO and Control-EXO.
Notably, TEX from the CD40 gene-modified B16 tumor cells hardly
induced the proliferation of the primed CD4+ T cells
(Fig. 2D). These data reveal that
CD40L-EXO are more effective in inducing the phenotypic and
functional maturation of DCs than Lac Z-EXO and Control-EXO, and
CD40L-EXO may also induce a more efficient tumor antigen-specific
CD4+ T cell immunity.
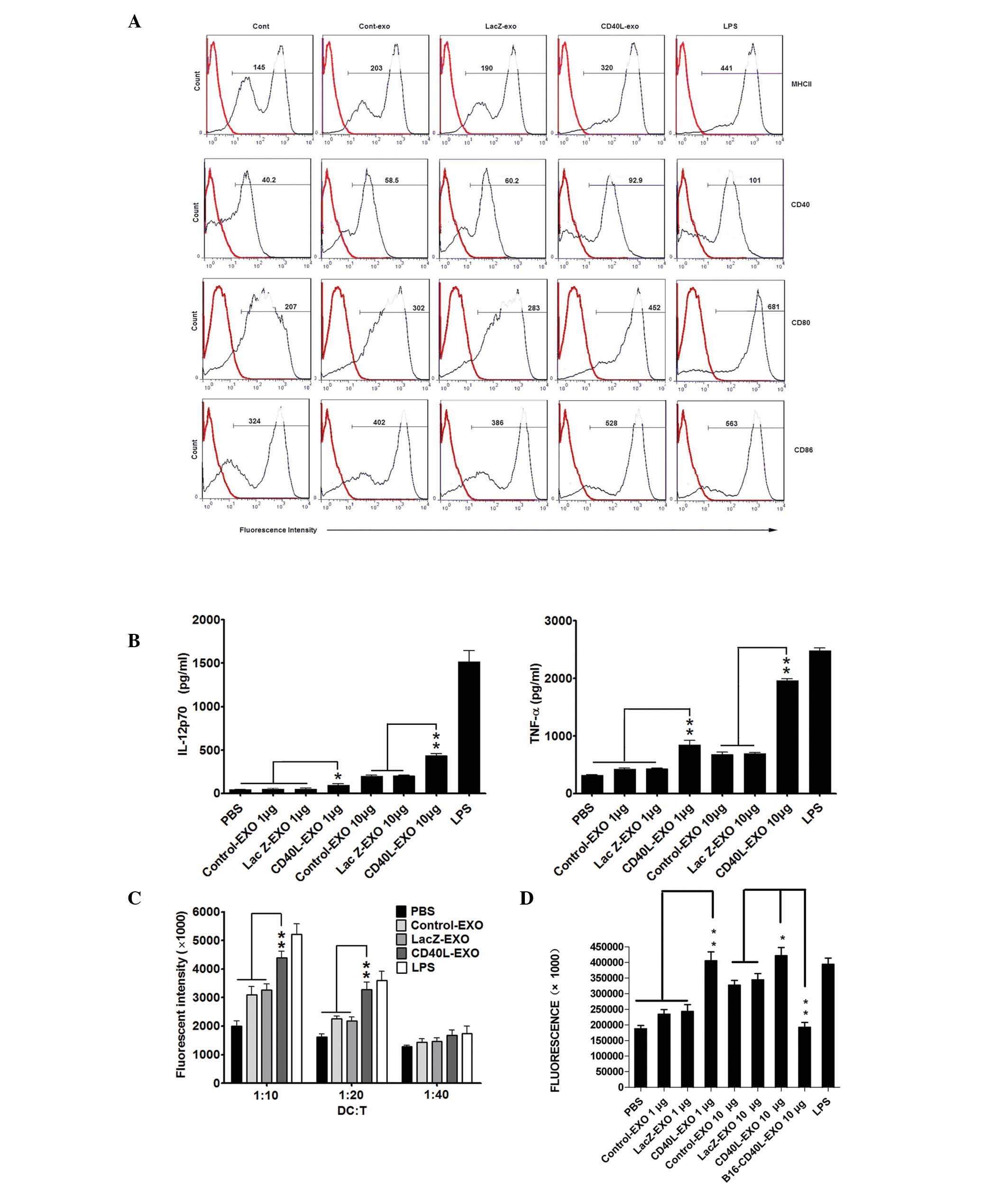 | Figure 2CD40L-EXO induce activation of BMDCs
more efficiently. (A) FACS analysis of the BMDC phenotype. BMDCs
were stimulated with each type of exosome (1 or 10 μg/ml) or LPS (1
μg/ml). Following 48 h of incubation, the cells were collected and
stained with isotype (broken line) or PE-labeled anti-MHC-II, CD40,
CD80 and CD86 (solid line) monoclonal antibody, washed and analyzed
by FACS. The numbers indicate the mean fluorescence intensity. The
data are representative of three independent experiments. (B) In
parallel, cytokines in the supernatants were detected by ELISA. (C)
CD40L-EXO increased the ability of DCs to stimulate allogeneic
splenic CD4+ T cell proliferation in MLR. Following
co-culture with each type of exosome for 48 h, BMDCs were collected
and inactivated by mitomycin C. The splenic CD4+ T cells
from BALB/c mice were cultured with the pre-treated C57BL/6 DCs at
the indicated ratios for 96 h. The proliferation of T cells was
determined by the alamarBlue assay (n=5). (D) Tumor
antigen-specific CD4+ T cell proliferation induced by
CD40L-EXO. C57BL/c mice were immunized with 3LL cell lysates.
CD4+ T cells were purified from the spleen of immunized
mice on day seven and co-cultured with syngeneic BMDCs that were
pretreated with each type of exosome (1 or 10 μg/ml). Following a
total 96 h of culture, the proliferation of CD4+ T cells
was also determined by the alamarBlue assay (n=5). Data are
representative of three independent experiments.
*P<0.05 and **P<0.01. CD40L-EXO, CD40
ligand gene-modified 3LL tumor cells; BMDC, bone marrow-derived
dendritic cells; FACS, fluorescence activated cell sorting; MLR,
mixed lymphocyte reaction; DC, dendritic cells; LPS,
lipopolysaccharide; PE, phycoerythrin.. |
CD40L-EXO induce a stronger protective
antitumor immunity
To determine the antitumor immunity induced by
CD40L-EXO, the mice were immunized with each type of exosome and
challenged s.c. with 1×106 3LL cells. The immunization
of CD40L-EXO resulted in a significantly enhanced antitumor effect
when compared with the Lac Z-EXO-, Control-EXO- or PBS-treated mice
(Fig. 3A). The survival rate of
the tumor mice immunized with CD40L-EXO was ~80% on day 90, but
only 40 and 50% in the tumor mice immunized with Lac Z-EXO and
Control-EXO, respectively (Fig.
3B). As predicted, treatment with PBS revealed no significant
protection against the inoculated 3LL cells (Fig. 3A and B). These results indicate
that CD40L-EXO induce a stronger protective antitumor immunity than
unmanipulated exosomes.
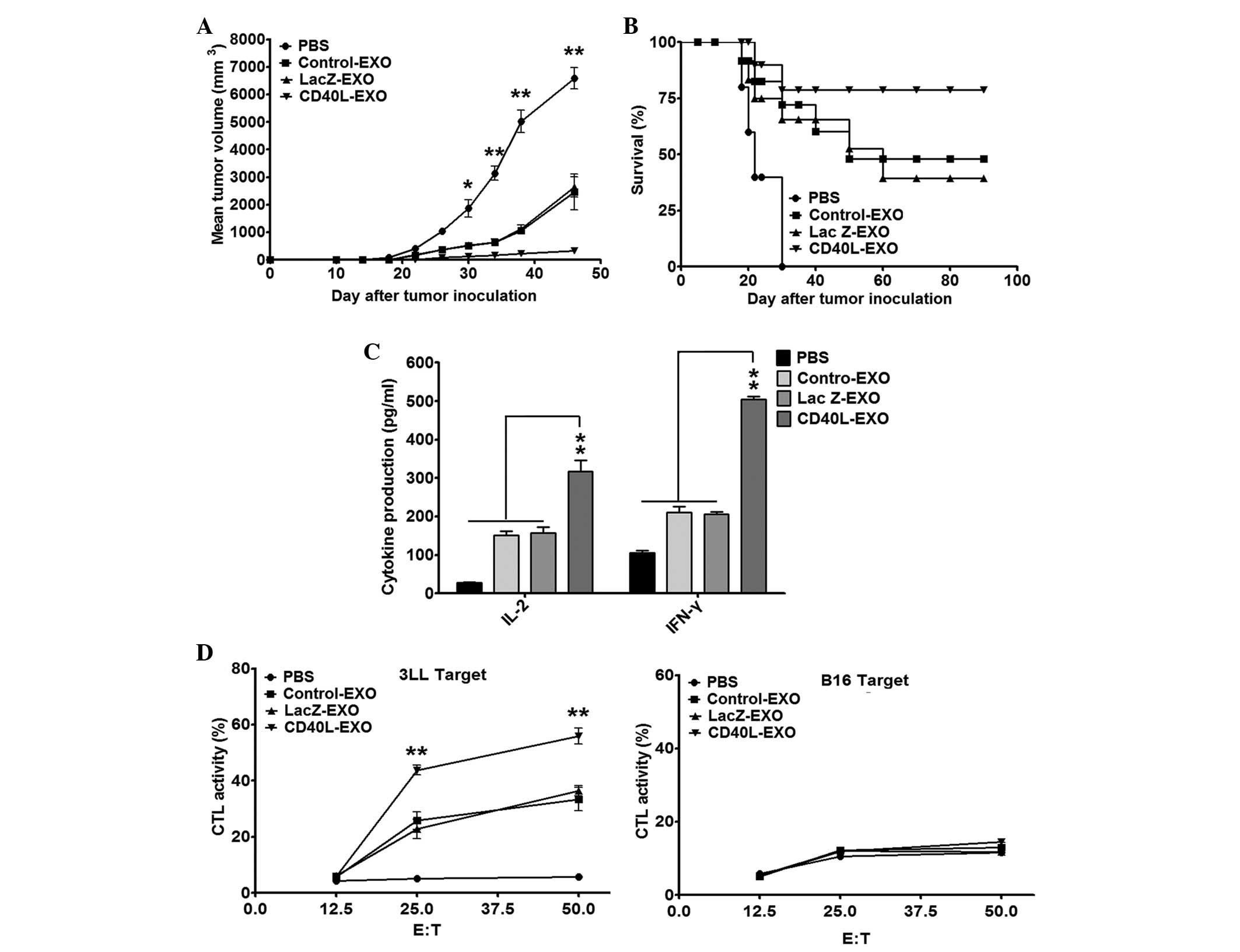 | Figure 3CD40L-EXO induce a stronger protective
antitumor immunity. Tumor development in CD40L-EXO-immunized mice.
(A) Tumors were established in C57BL/6 mice (n=10/group) by
immunization four times by s.c. injection with PBS and 10 μg of
each type of exosome on days -13, -11, -9 and -7. On day 0, the
mice received a s.c. injection of 1×106 3LL cells. The
tumor size was monitored with a caliper on the indicated days. (B)
The tumor-free mice were also recorded. (C) The levels of IFN-γ and
IL-2 secreted from splenocytes of C57BL/6 mice immunized with
CD40L-EXO. Mice (n=10) of different groups were sacrificed 7 days
following the last immunization. Splenocytes (5×106/ml)
were stimulated with inactivated 3LL cells at a ratio of 10:1 for
48 h. The levels of IFN-γ and IL-2 in the culture supernatants were
measured by ELISA. (D) CTL activity induced by CD40L-EXO
immunization. Splenocytes from immunized mice were stimulated with
inactivated 3LL cells and mIL-2 (30 U/ml) for 7 days in
vitro, and then the cytotoxicity of pulsed splenocytes to 3LL
target cells or syngeneic B16 cells that served as a specificity
control were determined by the LDH assay (n=5). Data are
representative of three independent experiments.
*P<0.05 and **P<0.01 vs. PBS,
Control-EXO or Lac Z-EXO. CD40L-EXO, CD40L gene-modified 3LL tumor
cells; CTL, cytotoxic T lymphocytes; s.c., subcutaneously; E:T,
effect cells : target cells; mIL-2, mouse interleukin 2. |
CTLs are important in the eradication of
tumor cells
Effective cytokines, including IFN-γ and IL-2, are
extremely important in the antitumor immunity of CTLs. The levels
of IFN-γ and IL-2 were detected from the splenocytes of the
exosomes from the immunized mice following restimulation with 3LL
cells ex vivo. Immunization with all exosomes significantly
increased the production of IFN-γ and IL-2 from the splenocytes.
However, the effect of CD40L-EXO was the strongest (Fig. 3C). Furthermore, the cytotoxic
activity of the splenocytes from the CD40L-immunized mice to 3LL
targets was more potent than that of the splenocytes from Lac
Z-EXO-, Control-EXO- or PBS-immunized mice (Fig. 3D). These results indicate that
immunization with CD40L-EXO induces more robust Th1 and
antigen-specific CTL responses.
CD40L-EXO induce a more powerful
therapeutic antitumor immunity
To further confirm the antitumor potential of
CD40L-EXO, the present study examined whether CD40L-EXO were
effective in eradicating the established tumor in mice. Mice were
inoculated with 3LL cells on day 0. Following the formation of
visible tumors, as observed by the naked eye, the mice were treated
with each type of exosomes (10 μg/mouse) by s.c. injection on days
+7, +9, +11 and +13. As predicted, tumor growth in the mice treated
with CD40L-EXO was effectively inhibited, with 50% of the mice
surviving up to day 90 (Fig. 4).
These results demonstrate that CD40L-EXO have a superior antitumor
effect compared with Lac Z-EXO and Control-EXO.
Discussion
With recent scientific and technological progress,
the development of tumor immunotherapy has significantly improved
the prognosis of cancer patients, but the reality of the ‘war on
cancer’ remains ongoing (18). A
number of technical challenges restrict the effect of tumor
immunotherapy in the clinic. For example, tumor antigens often
exhibit poor immunogenicity. Tumor cells frequently escape the
immune response via variation. Exosomes, as potential therapeutic
agents for tumors, have attracted the intensive attention of
oncologists (19,20). However, the efficacy of
traditionally prepared exosomes in the induction of antitumor
responses remains to be optimized. In the present study, exosomes
from CD40L gene-modified 3LL lung tumor cells were isolated, and it
was revealed that CD40L-EXO were more efficient in the induction of
antitumor T cell immunity.
Exosomes have been tested in three Phase I clinical
studies in melanoma, lung and colorectal cancer patients (19–21).
These studies demonstrated that the safety of administering
exosomes in humans was satisfactory. Clinical observations
indicated that exosomes may stimulate the adaptive and innate
cellular immune responses. Regardless of the observation of
positive antitumor responses, the effect of exosomes in tumor
therapy is not yet ideal. It has been reported that TEX from
immune-activated gene-modified tumor cells and heat-stressed tumor
cells or super-antigen anchored TEX demonstrated potent antitumor
effects (8,9,16).
Similarly, it was hypothesized that TEX from CD40L gene-modified
tumor cells also have enhanced the immunostimulatory potential.
This hypothesis was suggested to be valid by the findings of the
present study, which showed that CD40L-EXO more efficiently induced
DC activation and the subsequent priming of antitumor T cell
immunity.
DCs are important in the initiation of antitumor T
cell immunity, which requires the full activation of DCs. CD40
molecules are expressed on the immature DCs and are important in
their activation. Following maturation mediated by CD40 signaling,
DCs may effectively prime antigen-specific CD4+ and
CD8+ T cell responses (14). However, CD40L is predominantly
expressed on activated T cells, but not on resting T cells, which
is contradictory and inefficient for the activation of DCs.
CD40L-EXO-containing tumor antigens were rich in CD40L, which may
have efficiently activated the DCs, resulting in the activation of
tumor antigen-specific T cells. The activated T cells upregulated
the expression of CD40L and further activated the DCs, which
possibly led to a positive-feedback for tumor antigen-specific T
cell activation.
It has been reported that TEX from heat-stressed 3LL
lung tumor cells induce strong antitumor effects. These exosomes
have been demonstrated to effectively chemoattract DCs to the tumor
site (23). The accumulation of
DCs in the lungs is beneficial to prime a robust antitumor T cell
immunity. It also has been reported that DCs infiltrating human
non-small cell lung cancer are blocked at an immature stage
(22). In lung cancer, the
function of DCs is converted from immune activation to tolerance.
CD40L-EXO have the potent ability to activate DCs, which possibly
turns over the tolerance of DCs. It may be supposed that TEX from
heat-stressed and CD40L-gene modified 3LL lung tumor cells is
likely to increase the numbers of DCs in the tumor site and enhance
their activation. Therefore, TEX prepared from heat-stressed and
CD40L-gene modified lung tumor cells may be more promising for the
treatment of lung cancers.
Altogether, in the present study, CD40L-EXO were
isolated from 3LL tumor cells infected by Ad-CD40L and it was
demonstrated that CD40L-EXO were more efficient in DC activation
and antitumor T cell immunity induction. As cell-free vesicles,
exosomes are stable for a long period of time when preserved at
−80°C. The production of TEX is extremely large-scale and the cost
for TEX preparation is economical. Moreover, due to TEX often
carrying a low number of or no MHC molecules, the biological safety
of TEX is ideal. All these characteristics render CD40L-EXO a
prospective universal agent for the therapy of lung cancer.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 81200014), the Natural
Science Foundation of Zhejiang Province (grant no. LY12H13003) and
the Medicine and Health Foundation of the Health Bureau of Zhejiang
Province (grant nos. 2012KYA152 and 2010KYA104).
Abbreviations:
|
DCs
|
dendritic cells
|
|
CD40L
|
CD40 ligand
|
|
MLR
|
mixed lymphocyte reaction
|
|
CTLs
|
cytotoxic T lymphocytes
|
|
TEX
|
exosomes derived from tumor cells
|
|
Ad-CD40L
|
recombinant murine CD40L
adenovirus
|
|
CD40L-EXO
|
exosomes from Ad-CD40L-infected 3LL
lung cancer cells
|
|
Lac Z-EXO
|
exosomes from Ad-Lac Z-infected 3LL
lung cancer cells
|
|
Control-EXO
|
exosomes from uninfected 3LL lung
cancer cells
|
|
Ad
|
adenovirus
|
|
BMDCs
|
bone marrow-derived DCs
|
References
|
1
|
Sugimura H, Nichols FC, Yang P, et al:
Survival after recurrent nonsmall-cell lung cancer after complete
pulmonary resection. Ann Thorac Surg. 83:409–418. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Théry C, Zitvogel L and Amigorena S:
Exosomes: composition, biogenesis and function. Nat Rev Immunol.
2:569–579. 2002.
|
|
3
|
Lamparski HG, Metha-Damani A, Yao JY, et
al: Production and characterization of clinical grade exosomes
derived from dendritic cells. J Immunol Methods. 270:211–226. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chaput N and Théry C: Exosomes: immune
properties and potential clinical implementations. Semin
Immunopathol. 33:419–440. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ohshima K, Inoue K, Fujiwara A, et al:
Let-7 microRNA family is selectively secreted into the
extracellular environment via exosomes in a metastatic gastric
cancer cell line. PLoS One. 5:e132472010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang J, Zhang Y, Luo C, Xia Y, Chen H and
Wu X: Glycosyl-phosphatidylinositol-anchored interleukin-2
expressed on tumor-derived exosomes induces antitumor immune
response in vitro. Tumori. 96:452–459. 2010.
|
|
7
|
Zhang Y, Wu XH, Luo CL, Zhang JM, He BC
and Chen G: Interleukin-12-anchored exosomes increase cytotoxicity
of T lymphocytes by reversing the JAK/STAT pathway impaired by
tumor-derived exosomes. Int J Mol Med. 25:695–700. 2010.PubMed/NCBI
|
|
8
|
Dai S, Wan T, Wang B, et al: More
efficient induction of HLA-A*0201-restricted and carcinoembryonic
antigen (CEA)-specific CTL response by immunization with exosomes
prepared from heat-stressed CEA-positive tumor cells. Clin Cancer
Res. 11:7554–7563. 2005.
|
|
9
|
Xiu F, Cai Z, Yang Y, Wang X, Wang J and
Cao X: Surface anchorage of superantigen SEA promotes induction of
specific antitumor immune response by tumor-derived exosomes. J Mol
Med (Berl). 85:511–521. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Armitage RJ, Fanslow WC, Strockbine L, et
al: Molecular and biological characterization of a murine ligand
for CD40. Nature. 357:80–82. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Melief CJ: Cancer immunotherapy by
dendritic cells. Immunity. 29:372–383. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dohnal AM, Luger R, Paul P, Fuchs D and
Felzmann T: CD40 ligation restores type 1 polarizing capacity in
TLR4-activated dendritic cells that have ceased interleukin-12
expression. J Cell Mol Med. 13:1741–1750. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Elmetwali T, Searle PF, McNeish I, Young
LS and Palmer DH: CD40 ligand induced cytotoxicity in carcinoma
cells is enhanced by inhibition of metalloproteinase cleavage and
delivery via a conditionally-replicating adenovirus. Mol Cancer.
9:522010. View Article : Google Scholar
|
|
14
|
van Kooten C and Banchereau J: CD40-CD40
ligand. J Leukoc Biol. 67:2–17. 2000.
|
|
15
|
Van Nuffel AM, Corthals J, Neyns B,
Heirman C, Thielemans K and Bonehill A: Immunotherapy of cancer
with dendritic cells loaded with tumor antigens and activated
through mRNA electroporation. Methods Mol Biol. 629:405–452.
2010.PubMed/NCBI
|
|
16
|
Dai S, Zhou X, Wang B, et al: Enhanced
induction of dendritic cell maturation and HLA-A*0201-restricted
CEA-specific CD8(+) CTL response by exosomes derived from IL-18
gene-modified CEA-positive tumor cells. J Mol Med (Berl).
84:1067–1076. 2006.
|
|
17
|
Chen W, Wang J, Shao C, et al: Efficient
induction of antitumor T cell immunity by exosomes derived from
heat-shocked lymphoma cells. Eur J Immunol. 36:1598–1607. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Miller MJ, Foy KC and Kaumaya PT: Cancer
immunotherapy: present status, future perspective, and a new
paradigm of peptide immunotherapeutics. Discov Med. 15:166–176.
2013.PubMed/NCBI
|
|
19
|
Escudier B, Dorval T, Chaput N, et al:
Vaccination of metastatic melanoma patients with autologous
dendritic cell (DC) derived-exosomes: results of thefirst phase I
clinical trial. J Transl Med. 3:102005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Morse MA, Garst J, Osada T, et al: A phase
I study of dexosome immunotherapy in patients with advanced
non-small cell lung cancer. J Transl Med. 3:92005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wei H, Wang H, Lu B, et al: Cancer
immunotherapy using in vitro genetically modified targeted
dendritic cells. Cancer Res. 68:3854–3862. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Perrot I, Blanchard D, Freymond N, et al:
Dendritic cells infiltrating human non-small cell lung cancer are
blocked at immature stage. J Immunol. 178:2763–2769. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen T, Guo J, Yang M, Zhu X and Cao X:
Chemokine-containing exosomes are released from heat-stressed tumor
cells via lipid raft-dependent pathway and act as efficient tumor
vaccine. J Immunol. 186:2219–2228. 2011. View Article : Google Scholar : PubMed/NCBI
|