Introduction
Acute pancreatitis (AP) is a commonly occurring
disease of considerable severity. AP involves a complicated cascade
of events initiated by the injury of pancreatic acinar cells
(1), resulting in local and
systemic inflammation (2).
Although the exact mechanism underlying acinar cell injury in AP is
complex and remains to be defined, it has been hypothesized that
calcium overload is the trigger of acinar cell injury in AP
(3). Previous studies have
indicated that excess insult agents, including bile salts (4), ethanol metabolites and fatty acids
(5,6), induce marked and global calcium
release from the endoplasmic reticulum (ER) to the cytoplasm, which
further induces premature intracellular digestive enzyme
activation, vacuolization, secretory blockade, disorganization of
the cytoskeleton and cellular necrosis. Furthermore, calcium
depletion of the ER stores has been observed to induce the
activation of ER stress responses, which is associated with acinar
injury and AP (7). In rats with
arginine-induced AP (8), specific
key ER stress sensors, including RNA-activated protein kinase-like
ER kinase (PERK), activation transcription factor 6 (ATF6) and
inositol requiring protein 1 (IRE1), are highly activated and these
signaling molecules lead to complex responses, including
translation attenuation, inflammatory and immune responses and
induction of apoptosis. Similarly, in the cerulein model of acute
pancreatitis, ER stress responses are observed early, paralleling
trypsinogen activation (9,10). It has also been observed that
stimulation of isolated pancreatic acini by secretagogues generates
distinct ER stress responses and eventually cell death (11). Moreover, reducing ER stress by
tauroursodeoxycholic acid, a chemical chaperone or by genetic
manipulation of ER chaperone immunoglobulin-binding protein (Bip),
induces protective effects against acute pancreatitis (12,13).
All these observations indicate that ER stress is one of the
mechanisms underlying pancreatic acinar cell injury and may be a
potential target for preventing and treating this disease.
Emodin (6-methyl-1,3,8-tanthragallol), the
predominant effective component of rhubarb, has been used
clinically to treat AP for a number of years in China (14). Several in vivo
investigations have also shown that emodin has significant
therapeutic effects on AP in rats by correcting intestinal flora
disturbances, promoting intestinal peristalsis (15) and inhibiting inflammatory cytokine
release and pancreatin activity (16). Furthermore, studies have shown that
emodin ameliorates pancreatic acinar cell injury by suppressing
toll-like receptor 4 expression, inhibiting pancreatin activity,
scavenging oxygen free radicals, increasing occludin expression and
promoting apoptosis in the pathogenesis of AP. Our previous studies
have shown that emodin markedly ameliorates pancreatic injury via
attenuating ER stress responses in the pancreas of rats with AP
induced by sodium taurocholate (17). In the present study, the effects of
emodin on the activation of ER stress sensor proteins were further
evaluated using an in vitro model of AP.
Materials and methods
Materials
Rat pancreatic acinar AR42J cells (CRL 1492) were
obtained from the American Type Culture Collection (Manassas, VA,
USA). Emodin was purchased from China National Institutes for Food
and Drug Control (Beijing, China). Ham's F12 medium, cerulein,
lipoplysaccharide (LPS) and dexamethasone were purchased from
Sigma-Aldrich, (St. Louis, MO, USA). Fetal bovine serum (FBS),
glutamine, penicillin/streptomycin, fluo-3 acetoxymethyl ester were
obtained from Invitrogen Life Technologies (Carlsbad, CA, USA). The
amylase assay kit was purchased from Nanjing Jiancheng
Bioengineering Institute (Nanjing, China). RNase-free DNase set and
RNA safe were purchased from Tiangen Biotech. Co. Ltd. (Beijing,
China). An RNA extraction kit, Oligo(dT)15 and dNTP were
obtained from Servier Pharmaceutical Research & Development Co.
Ltd. (Beijing, China). M-MLV reverse transcriptase was purchased
from Promega Corporation (Madison, WI, USA). Primers were designed
and synthesized by Invitrogen Life Technologies.
Cell culture
AR42J is the only cell line that exhibits numerous
characteristics of normal pancreatic acinar cells and has been used
to study the secretion, growth and proliferation of exocrine
pancreatic cells (18). In the
present study, this cell line was used as an in vitro model
to evaluate the effects of emodin against pancreatic acinar cell
injury and the activation of ER stress sensor proteins. AR42J cells
were cultured in Ham's F12 medium supplemented with 2 mM glutamine,
20% FBS and antibiotics (100 U/ml penicillin and 100 μg/ml
streptomycin) at 37°C in a humidified atmosphere of 5%
CO2 and 95% air. Treatment with dexamethasone has been
observed to convert these cells into exocrine cells (19). AR42J cells were plated at a density
of 2×104 cells/cm2 in 6-well culture dishes,
incubated for 48 h with 100 nM dexamethasone and washed with
phosphate-buffered saline (PBS) prior to incubation in fresh
medium.
Emodin treatment protocol
In preliminary experiments, the toxic effects of
emodin on AR42J cells were detected by examining the cellular
viability and amylase activity in culture media. AR42J cells were
incubated in the absence of stimulus or in the presence of 5, 10,
20, 40 and 80 μM emodin. Following incubation for 3, 6, 12, 24 and
48 h, the cell viability and amylase activity in the culture media
were evaluated. The results showed that the cellular viability and
amylase activity in the culture media did not vary in incubation
with 5–40 μM emodin for 48 h; however, the viability of AR42J cells
was reduced when treated with emodin at 40 μM for 24 or 48 h
(P<0.01) (data not shown). Therefore, in this study, the cells
were treated with emodin at 10 and 20 μM.
AR42J cells were divided into control, model
(treated with cerulein plus LPS) and emodin treatment (treated with
cerulein, LPS and emodin at 10 or 20 μM) groups. For the treatment
groups, cells were pre-treated with emodin at various
concentrations for 10 min prior to the addition of cerulein and
LPS. After 3 h the culture media and cells were harvested for
further analysis.
Measurement of amylase activity
Culture media was collected and the cells were
washed twice with Krebs-HEPES buffer [140 mM NaCl, 5.9 mM KCl, 1.2
mM MgCl2, 15 mM HEPES, 10 mM glucose, 2.5 mM
CaCl2, 0.1 mM PMSF and 2.5 mM aprotinin (pH 7.4)]. Cells
were then lysed by being forced to pass through a 26G syringe
followed by brief sonication. The amylase activity in the culture
media and cellular lysates was measured using an enzyme assay kit
(Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Amylase release was determined as the percentage of total amylase
secreted into the culture media.
Measurement of cytosolic calcium
Fluorescence imaging was performed to evaluate the
cytosolic calcium concentration. AR42J cells were plated at a
density of 2×104 cells/cm2 in 35-mm glass
bottom dishes. Cells were loaded with 5 μM membrane-permeable
calcium indicator, Fluo-3AM ester, for 30 min at 37°C in Hank's
buffer. Following loading with the fluo-3 dye, cells were washed
with D-Hank's solution that lacked calcium and contained 5 mM EGTA.
Fluorescence measurements were performed using an Olympus
Fluoview-500 confocal system equipped with a 40× objective (Olympus
Corporations, Tokyo, Japan). Fluo-3 was excited by argon laser
light at 488 nm wavelength and fluorescence was observed at 515 nm
wavelength. Image processing and analysis were performed using
Olympus Fluoview software. Intracellular calcium were expressed as
average fluorescence intensity (FI).
Reverse transcription-polymerase chain
reaction (RT-PCR)
Gene expression of Bip, PERK, ATF6 and IRE1α in
AR42J cells was assessed using RT-PCR standardized by co-amplifying
with GAPDH, which served as an internal control. Total RNAs
isolated from the cells were reversely transcribed into cDNAs and
used for PCR with rat specific primers for Bip, PERK, ATF6, IRE1α
and GAPDH. The sequences of primers used in the PCR were as
follows: Forward: ATTCCTGCGTC GGTGTATT and reverse:
TCGGCAGTTTCCTTCATTT for Bip; forward: TACAGTGGACGGCGATGATGAG and
reverse: CTTAGGGTGGTTGGCCTGGTAG for PERK; forward:
TCCCTCCACCTCCATGTCA and reverse: CTTCCAGGCGAAGCGTAAT for ATF6;
forward: GCTGTGGAGACCCTACGCTAT and reverse: TCGATGTTTGGGAAGATTGTTAG
for IRE1α; and forward, AAGGTCGGTGTGAACGGATTT and reverse,
AGATGATGACCCTTTTGGCCC for GAPDH. The predicted sizes of the PCR
products were: Bip, 358 bp; PERK, 270 bp; ATF6, 265 bp; IRE1α, 426
bp; and GAPDH, 352 bp. Following co-amplification with 32–35
cycles, the PCR products were separated on 1.5% agarose gels and
visualized by UV transillumination.
Flow cytometry
AR42J cells were collected to detect apoptosis and
necrosis using Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) staining according to the
manufacturer's instructions. Briefly, 1×105 cells were
digested, washed twice with cold PBS and re-suspended in 500 μl 1X
binding buffer. Next, 5 μl annexin V and 10 μl PI were added. Cells
were gently vortexed and incubated for 15 min at 25°C in the dark
followed by analysis by flow cytometry within 1 h.
Statistical analysis
Data are presented as the mean ± SD. The comparisons
between two groups were performed by a one-way analysis of variance
test. All statistical analyses were performed using SPSS 15.0
(SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Effects of emodin on the expression and
release of amylase in AR42J cells treated with cerulein and
LPS
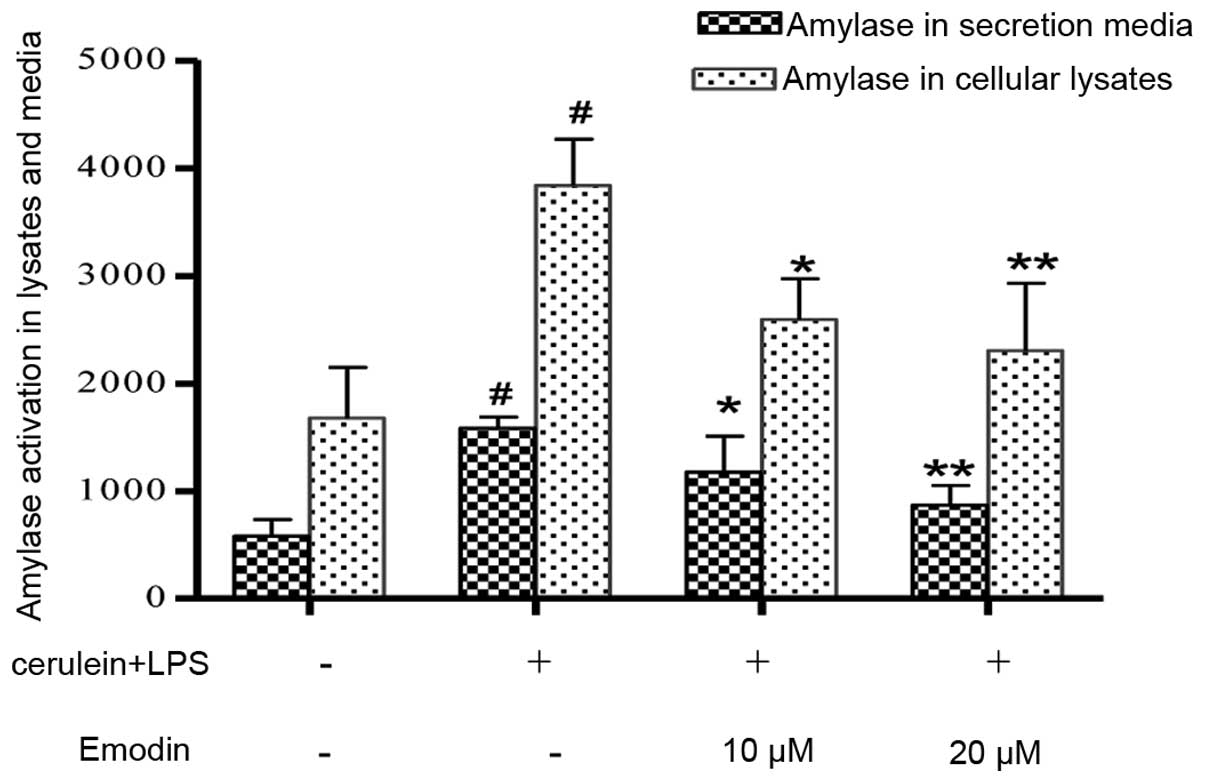
Effects of emodin on the expression and release of
amylase are shown in Fig. 1.
Compared with the control cells, the levels of amylase in cellular
lysates and culture media were significantly elevated in the model
group (P<0.01). Pretreatment with emodin markedly decreased the
amylase activity in AR42J cell lysates and culture media.
Effects of emodin on cytosolic calcium
concentration in AR42J cells treated with cerulein and LPS
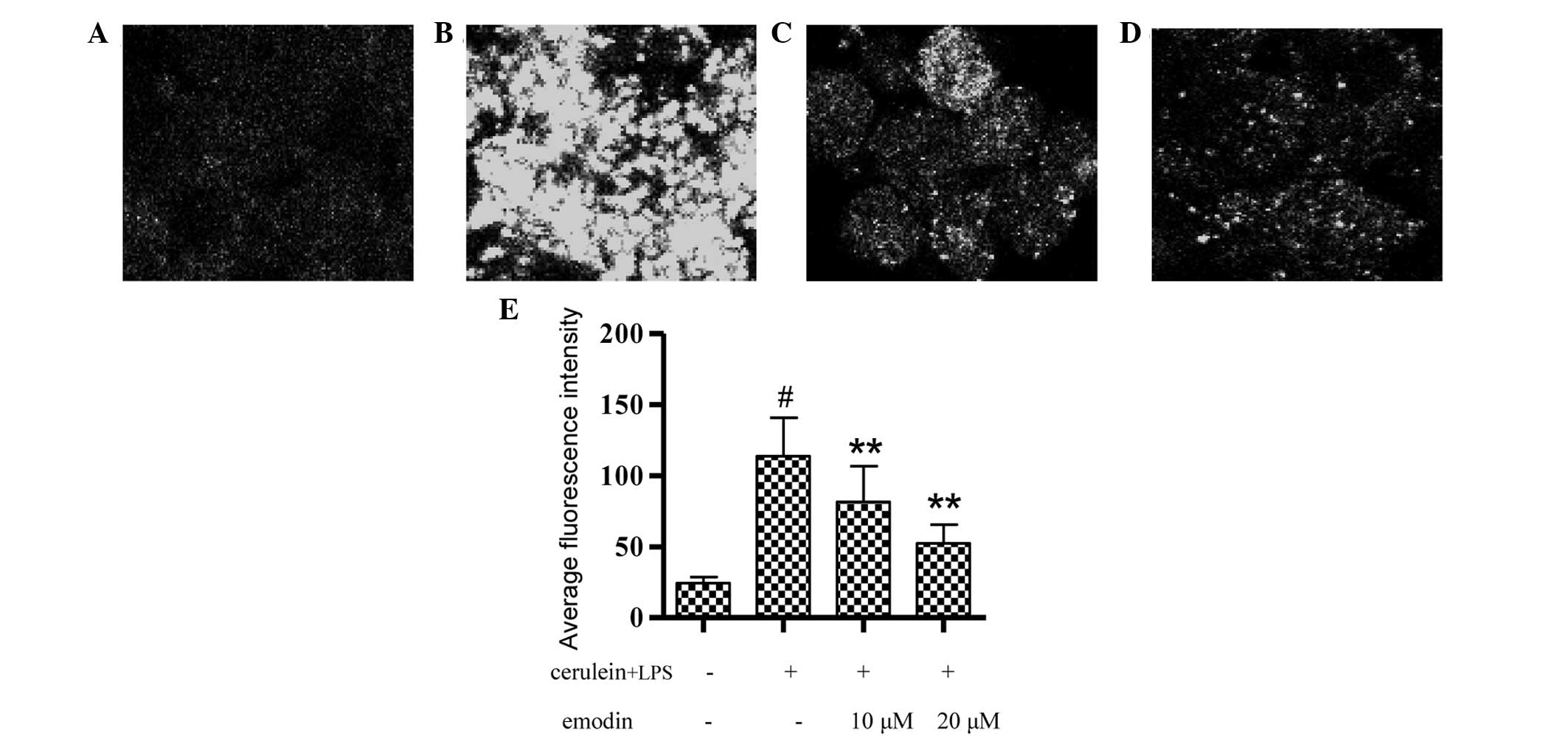
The cytosolic calcium concentrations of AR42J cells
are shown in Fig. 2. Stimulation
of AR42J cells with cerulein plus LPS for 3 h led to calcium
overload in the cytoplasm (Fig.
2B) and the average FI was higher than that of the control
group (Fig. 2E; P<0.01).
Pretreatment with emodin at 10 or 20 μM markedly alleviated calcium
overload in AR42J cells (Fig. 2C and
D) and also decreased the average FI value (Fig. 2F; P<0.01).
Effects of emodin on the mRNA expression
of Bip, PERK, ATF6 and IRE1 in AR42J cells treated with cerulein
plus LPS
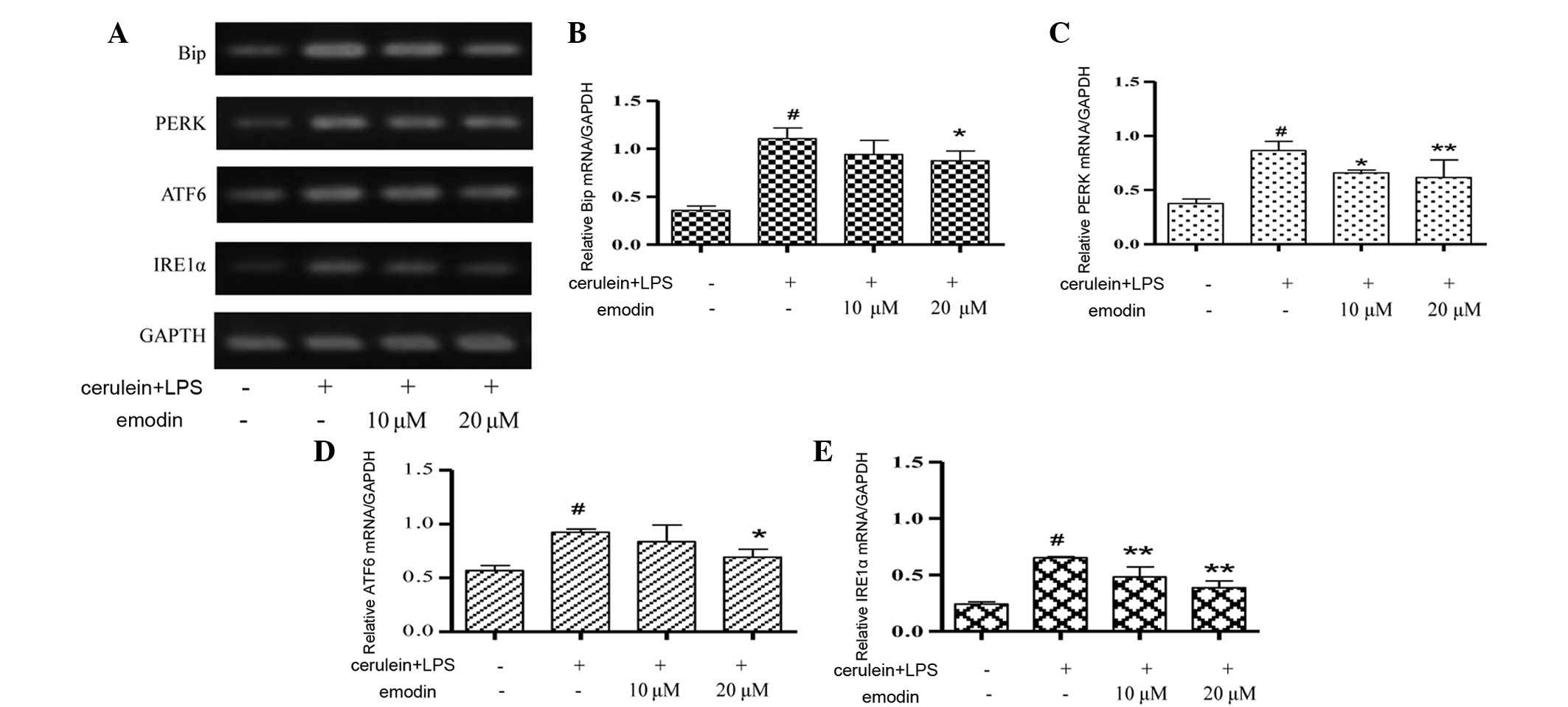
UPR is a cellular response associated with ER
stress. The mRNA expression of UPR transducers, including Bip,
PERK, ATF6 and IRE1 in AR42J cells is shown in Fig. 3. In the model group, several UPR
transducers were activated, as shown by the significantly increased
mRNA expression of Bip, PERK, IRE1 and ATF6 compared with the
control cells (P<0.01). Pretreatment with emodin at 10 μM
significantly reduced the mRNA expression of PERK (P<0.05) and
IRE1 (P<0.01) and in the 20 μM group, emodin significantly
attenuated the upregulated mRNA levels of ER stress transducers,
Bip (P<0.05), PERK (P<0.01), IRE1 (P<0.01) and ATF6
(P<0.05).
Effects of emodin on apoptosis and
necrosis of AR42J cells treated with cerulein plus LPS
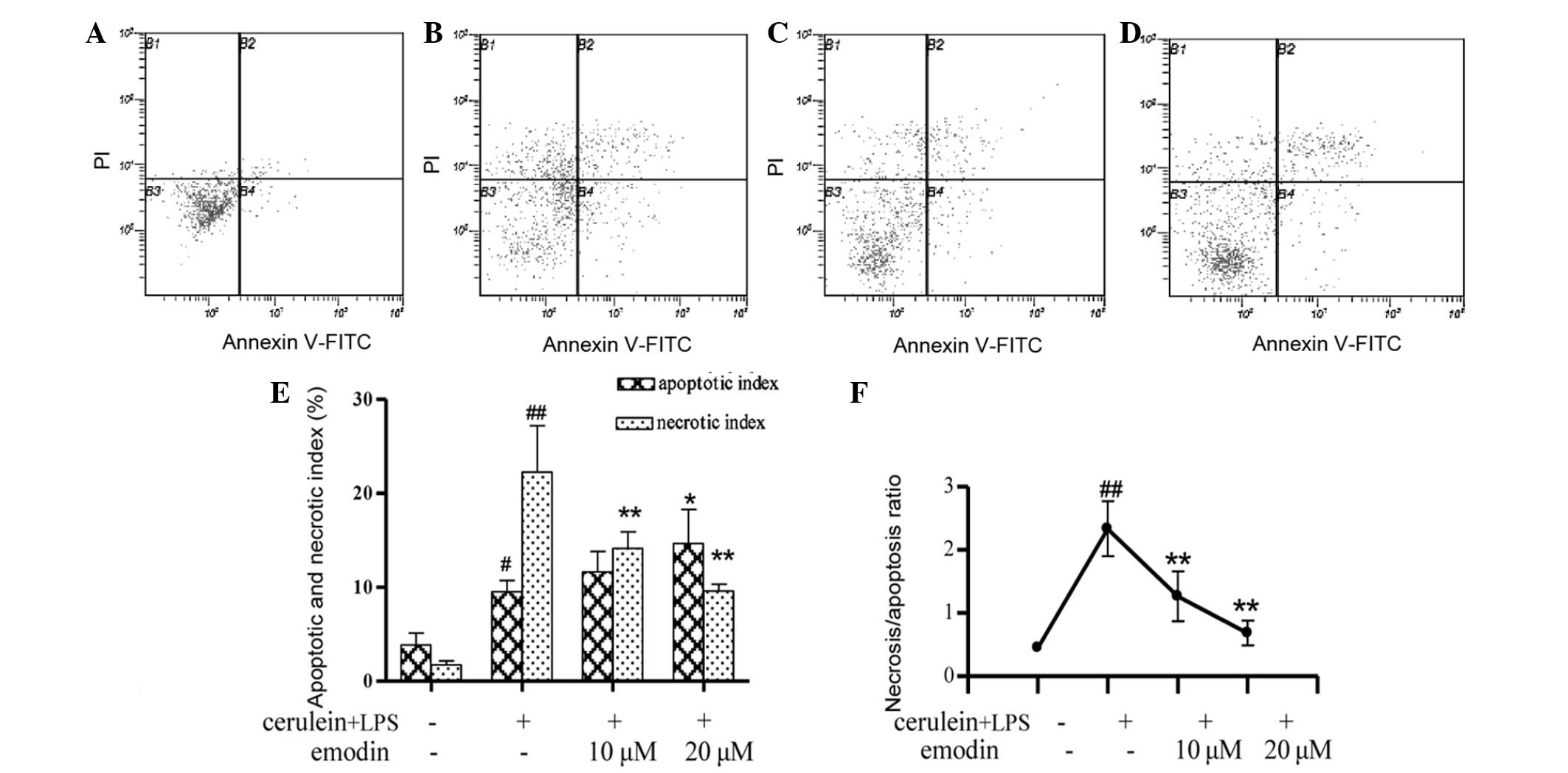
The occurrence of apoptosis and necrosis of AR42J
cells is shown in Fig. 4. Compared
with the control group, cerulein plus LPS caused a significant
increase in apoptosis and necrosis of AR42J cells (Fig. 4A and B). The apoptotic (P<0.05)
and necrotic indices (P<0.01) were markedly increased (Fig. 4E). In addition, the
necrosis/apoptosis ratio in the cerulein plus LPS group was 5-fold
higher than that in the control group (Fig. 4F). However, pretreatment with
emodin at 10 or 20 μM significantly attenuated the necrotic indices
(Fig. 4C–E; P<0.01) and in the
20 μM dose group, the apoptotic indices were increased (Fig. 4D and E; P<0.05). Accordingly,
the necrosis/apoptosis ratio in AR42J cells treated with emodin was
significantly lower than that in the model group (Fig. 4F; P<0.01).
Discussion
The pancreatic acinar cell is the functional unit of
the exocrine pancreas and synthesizes, stores and secretes
digestive enzymes. Under physiological conditions, secretagogue
substances, including cholecystokinin and acetylcholine, induce
instantaneous increases in the concentration of cytosolic calcium,
which passes through the plasma membrane and is released from the
ER, in acinar cells. These processes are tightly controlled in a
spatiotemporal manner and coupled to the physiological dose of
zymogen expression (20). However,
under pathological conditions, cytosolic calcium homeostasis is
disrupted, as calcium stores (ER and apical stores) become depleted
and the level of calcium in the cytoplasm remains elevated. To
date, cytosolic calcium overload has been hypothesized to be
pivotal in inducing pancreatic acinar cell injury (21), which was found to impair cellular
defense mechanisms and initiate the activation of premature
digestive enzymes within the cell. These processes led to the death
of acinar cells (22,23) with spillage of activated enzymes
into the interstitial space, affecting the surrounding acinar cells
and initiating a vicious circle that ends in AP and systemic
inflammatory response syndrome. Thus, inhibition of calcium
overload in the early stages of the disease is a valid avenue of
therapeutic intervention for acinar cell injury and
pancreatitis.
As an effective laxative, antiphlogistic and
homeostatic traditional Chinese medicine, rhubarb is widely used in
the treatment of digestive system diseases, including constipation,
diarrhea, jaundice, pancreatitis and colitis (24). Emodin, a major ingredient isolated
from rhubarb, has shown therapeutic promise for pancreatitis in
vivo and in vitro, as it was found to suppress
inflammation, promote apoptosis, inhibit pancreatin activity and
scavenge oxygen free radicals. However, to date, few studies have
determined whether emodin affects intracellular calcium homeostasis
in pancreatic acinar cells during the development of pancreatitis,
although specific studies have demonstrated that the
anti-inflammatory and analgesic activities of emodin are associated
with the regulation of intracellular calcium homeostasis.
Kuo et al(25) showed that emodin reduces the local
inflammatory response by blocking calcium mobilization and
diminishing inflammatory T cell activation and inflammatory
cytokine production in rats with arthritis. Sui et
al(26) reported that emodin
inhibits inflammatory hyperalgesia by significantly downregulating
the intracellular calcium fluorescence intensity in the dorsal root
ganglion neurons and TRPV1 mRNA expression. In the present study,
the effects of emodin on calcium overload were evaluated in AR42J
cells treated with cerulein plus LPS, an in vitro model
mimicking severe AP (27). The
results showed that the levels of calcium in the cytoplasm remained
elevated, accompanied by marked apoptosis and necrosis. However,
pretreatment with emodin was found to markedly alleviate calcium
overload in AR42J cells. Furthermore, emodin significantly
ameliorated pancreatic cell injury by inducing apoptosis and
decreasing necrosis.
The association between calcium overload and
organelle dysfunction is being increasingly studied. As the most
important reservoir of calcium, the ER participates in calcium
overload during cell injury. Pathological stimuli, including fatty
acid ethyl esters, cholecystokinin and cerulein, have been shown to
induce massive calcium release via inositol triphosphate receptors
from ER calcium stores, which is sustained by calcium entry
resulting in mitochondrial depolarization, failure of ATP
production and cellular necrosis (28). By contrast, calcium depletion of
the ER may induce the perturbation of ER homeostasis leading to the
activation of UPR that is associated with acinar cell injury, as
well as death. As part of the ER stress responses, unfolded protein
response contains at least three distinct signaling components that
are activated, PERK, ATF6 and IRE1. These stress sensors and their
downstream signaling partners participate in apoptosis and the
inflammatory reaction when severe ER stress occurs (29). Severe stress also induces the
release of ER calcium, which may stimulate mitochondrial generation
of reactive oxygen species and lead to the activation of nuclear
factor-κB, subsequently inducing inflammatory responses. Thus,
sustained calcium overload and severe ER stress responses lead to a
vicious cycle of cell injury and death.
In the present study, the effects of emodin on mRNA
expression of the key stress sensors were investigated in AR42J
cells using RT-PCR. Cerulein plus LPS-induced AR42J cell injury was
accompanied by a significant increase in the mRNA expression of
Bip, PERK, ATF6 and IRE1. Pretreatment with emodin reduced the mRNA
expression of these molecules, particularly IRE1, whose mRNA level
was significantly lowered by emodin at 10 and 20 μM compared with
the model cells, indicating that emodin attenuates ER stress
responses in AR42J cells. IRE1 has been hypothesized to be an ER
stress sensor protein and is important for the transduction of
stress signals from the ER to the cytoplasm and nucleus (30). It has been shown that activated
IRE1α on the ER membrane recruits tumor necrosis factor
receptor-associated factor 2 (TRAF2) and apoptosis
signal-regulating kinase 1 (31).
In addition, the activation of the IRE1α/TRAF2 complex may result
in the phosphorylation of c-Jun N-terminal kinase and p38
mitogen-activated protein kinases and promote pro-inflammatory
cytokine activation, subsequently inducing inflammatory responses,
cell injury and death (32,33).
In conclusion, the results of the present study
showed that calcium overload and ER stress are early molecular
events of specific forms of cell injury, apoptosis and necrosis in
AR42J cells. Emodin may alleviate cell injury and decrease the
number of apoptotic and necrosis cells accompanied by increasing
apoptosis and decreasing necrosis. These effects are likely to be
associated with reduced calcium overload and suppressed ER stress
responses.
Acknowledgements
This work was supported by Priority Academic Program
Development of Jiangsu Higher Education Institutions (ysxk-2010),
Open Project Program of National First-Class Key Discipline for
Traditional Chinese Medicine of Nanjing University of Chinese
Medicine (2011ZYX4-006), National Natural Science Foundation of
China (81073022), Science and Technology Foundation of Jiangsu
Traditional Chinese Medicine Administration (LZ11190).
References
|
1
|
Talukdar R and Swaroop Vege S: Early
management of severe acute pancreatitis. Curr Gastroenterol Rep.
13:123–130. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kylänpää ML, Repo H and Puolakkainen PA:
Inflammation and immunosuppression in severe acute pancreatitis.
World J Gastroenterol. 16:2867–2872. 2010.
|
|
3
|
Gorelick FS and Thrower E: The acinar cell
and early pancreatitis responses. Clin Gastroenterol Hepatol. 7(11
Suppl): S10–S14. 2009. View Article : Google Scholar
|
|
4
|
Kim JY, Kim KH, Lee JA, Namkung W, Sun AQ,
Ananthanarayanan M, Suchy FJ, Shin DM, Muallem S and Lee MG:
Transporter-mediated bile acid uptake causes
Ca2+-dependent cell death in rat pancreatic acinar
cells. Gastroenterology. 122:1941–1953. 2002. View Article : Google Scholar
|
|
5
|
Criddle DN, Raraty MG, Neoptolemos JP,
Tepikin AV, Petersen OH and Sutton R: Ethanol toxicity in
pancreatic acinar cells: mediation by nonoxidative fatty acid
metabolites. Proc Natl Acad Sci USA. 101:10738–10743. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Criddle DN, Murphy J, Fistetto G, Barrow
S, Tepikin AV, Neoptolemos JP, Sutton R and Petersen OH: Fatty acid
ethyl esters cause pancreatic calcium toxicity via inositol
trisphosphate receptors and loss of ATP synthesis.
Gastroenterology. 130:781–793. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kubisch CH and Logsdon CD: Endoplasmic
reticulum stress and the pancreatic acinar cell. Expert Rev
Gastroenterol Hepatol. 2:249–260. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kubisch CH, Sans MD, Arumugam T, Ernst SA,
Williams JA and Logsdon CD: Early activation of endoplasmic
reticulum stress is associated with arginine-induced acute
pancreatitis. Am J Physiol Gastrointest Liver Physiol.
291:G238–G245. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kubisch CH and Logsdon CD: Secretagogues
differentially activate endoplasmic reticulum stress responses in
pancreatic acinar cells. Am J Physiol Gastrointest Liver Physiol.
292:G1804–G1812. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Seyhun E, Malo A, Schäfer C, Moskaluk CA,
Hoffmann RT, Göke B and Kubisch CH: Tauroursodeoxycholic acid
reduces endoplasmic reticulum stress, acinar cell damage, and
systemic inflammation in acute pancreatitis. Am J Physiol
Gastrointest Liver Physiol. 301:G773–G782. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Szmola R and Sahin-Tóth M:
Pancreatitis-associated chymotrypsinogen C (CTRC) mutant elicits
endoplasmic reticulum stress in pancreatic acinar cells. Gut.
59:365–372. 2010. View Article : Google Scholar
|
|
12
|
Malo A, Krüger B, Seyhun E, Schäfer C,
Hoffmann RT, Göke B and Kubisch CH: Tauroursodeoxycholic acid
reduces endoplasmic reticulum stress, trypsin activation, and
acinar cell apoptosis while increasing secretion in rat pancreatic
acini. Am J Physiol Gastrointest Liver Physiol. 299:G877–G886.
2010. View Article : Google Scholar
|
|
13
|
Ye R, Mareninova OA, Barron E, Wang M,
Hinton DR, Pandol SJ and Lee AS: Grp78 heterozygosity regulates
chaperone balance in exocrine pancreas with differential response
to cerulein-induced acute pancreatitis. Am J Pathol. 177:2827–2836.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang G, Sun B, Gao Y, Meng QH and Jiang
HC: The effect of emodin-assisted early enteral nutrition on severe
acute pancreatitis and secondary hepatic injury. Mediators Inflamm.
2007:296382007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Z, Xia X, Zhang S, Zhang A, Bo W and
Zhou R: Up-regulation of Toll-like receptor 4 was suppressed by
emodin and baicalin in the setting of acute pancreatitis. Biomed
Pharmacother. 63:120–128. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia XM, Li BK, Xing SM and Ruan HL: Emodin
promoted pancreatic claudin-5 and occluding expression in
experimental acute pancreatitis rats. World J Gastroenterol.
18:2132–2139. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu JH, Lim JW, Kim KH, Morio T and Kim H:
NADPH oxidase and apoptosis in cerulein-stimulated pancreatic
acinar AR42J cells. Free Radic Biol Med. 39:590–602. 2005.
View Article : Google Scholar
|
|
18
|
Wu L, Cai B, Zheng S, Liu X, Cai H and Li
H: Effect of emodin on endoplasmic reticulum stress in rats with
severe acute pancreatitis. Inflammation. 36:1020–1029. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Eum WS, Li MZ, Sin GS, Choi SY, Park JB,
Lee JY and Kwon HY: Dexamethasone-induced differentiation of
pancreatic AR42J cell involves p21(waf1/cip1) and MAP kinase
pathway. Exp Mol Med. 35:379–384. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Low JT, Shukla A and Thorn P: Pancreatic
acinar cell: new insights into the control of secretion. Int J
Biochem Cell Biol. 42:1586–1589. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Frick TW: The role of calcium in acute
pancreatitis. Surgery. 152(Suppl 1): S157–S163. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Criddle DN, Gerasimenko JV, Baumgartner
HK, Jaffar M, Voronina S, Sutton R, Petersen OH and Gerasimenko OV:
Calcium signalling and pancreatic cell death: apoptosis or
necrosis? Cell Death Differ. 14:1285–1294. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Criddle DN, Sutton R and Petersen OH: Role
of Ca2+ in pancreatic cell death induced by alcohol
metabolites. J Gastroenterol Hepatol. 21(Suppl 3): S14–S17.
2006.
|
|
24
|
Chen DC and Wang L: Mechanisms of
therapeutic effects of rhubarb on gut origin sepsis. Chin J
Traumatol. 12:365–369. 2009.PubMed/NCBI
|
|
25
|
Kuo YC, Meng HC and Tsai WJ: Regulation of
cell proliferation, inflammatory cytokine production and calcium
mobilization in primary human T lymphocytes by emodin from
Polygonum hypoleucum Ohwi. Inflamm Res. 50:73–82. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sui F, Huo HR, Zhang CB, Yang N, Guo JY,
Du XL, Zhao BS, Liu HB, Li LF, Guo SY and Jiang TL: Emodin
down-regulates expression of TRPV1 mRNA and its function in DRG
neurons in vitro. Am J Chin Med. 38:789–800. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu Y, Zhou ZG, Chen KL, Zhou B, Yang L,
Yan H and Li Y: The ER chaperone GRP78 is associated with the
severity of cerulein-induced pancreatic inflammation via regulating
apoptosis of pancreatic acinar cells. Hepatogastroenterology.
59:1670–1676. 2012.PubMed/NCBI
|
|
28
|
Petersen OH and Sutton R: Ca2+
signalling and pancreatitis: effects of alcohol, bile and coffee.
Trends Pharmacol Sci. 27:113–120. 2006.
|
|
29
|
Kubisch CH and Logsdon CD: Secretagogues
differentially activate endoplasmic reticulum stress responses in
pancreatic acinar cells. Am J Physiol Gastrointest Liver Physiol.
292:G1804–G1812. 2007. View Article : Google Scholar
|
|
30
|
Pavitt GD and Ron D: New insights into
translational regulation in the endoplasmic reticulum unfolded
protein response. Cold Spring Harb Perspect Biol. 4:a0122782012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sendler M, Dummer A, Weiss FU, Krüger B,
Wartmann T, Scharffetter-Kochanek K, van Rooijen N, Malla SR,
Aghdassi A, Halangk W, Lerch MM and Mayerle J: Tumour necrosis
factor α secretion induces protease activation and acinar cell
necrosis in acute experimental pancreatitis in mice. Gut.
62:430–439. 2013.
|
|
32
|
Maguire JA, Mulugeta S and Beers MF:
Endoplasmic reticulum stress induced by surfactant protein C
Brichos mutants promotes proinflammatory signaling by epithelial
cells. Am J Respir Cell Mol Biol. 44:404–414. 2011. View Article : Google Scholar
|
|
33
|
Kim SM, Chung MJ, Ha TJ, Choi HN, Jang SJ,
Kim SO, Chun MH, Do SI, Choo YK and Park YI: Neuroprotective
effects of black soybean anthocyanins via inactivation of
ASK1-JNK/p38 pathways and mobilization of cellular sialic acids.
Life Sciences. 90:874–882. 2012. View Article : Google Scholar : PubMed/NCBI
|