Introduction
Acute lymphoblastic leukaemia (ALL) is a malignant
disorder of lymphoid progenitor cells. ALL is the most common type
of childhood malignancy and accounts for almost 20% of cases of
acute leukaemia in adults. B-cell precursor ALL (BCP-ALL) is the
most common type of childhood ALL (1). Survival rates have markedly improved
with combination chemotherapy and treatment intensification;
however, the occurrence of minimal residual disease has prompted
further examination of the molecular mechanisms underlying ALL
(2).
BCP-ALL is a malignancy characterised by the
progressive accumulation of immature clonal B-cell precursors in
the bone marrow. Numerous transcriptional regulators have critical
functions in this malignant process and may regulate the expression
of genes, whose products affect the fate and function of lymphoid
cells (3). The Ikaros family of
proteins is an example of these factors (4), which encode zinc-finger DNA-binding
proteins (5–7). Aiolos, the second Ikaros family
member identified, is of particular interest in the investigation
of BCP-ALL (6). Aiolos is an
important regulator of B-cell differentiation, proliferation and
maturation to an effector state. Disruption of Aiolos increases the
number of pre-B and immature B cells, and significantly reduces the
number of circulating B cells (8).
In addition, an Aiolos null mutation in mice causes B-cell
hyperproliferation, elevates serum antibody levels, promotes
auto-antibody formation and triggers lymphoma development, thereby
demonstrating tumour suppressor function (8). The deregulated expression of Aiolos
has been associated with adult B-cell ALL and chronic lymphocytic
leukaemia (CLL) in human patients (9–11)
and aberrant expression levels of Aiolos levels been reported in
cases of lymphoma (12).
However, the molecular mechanisms through which
Aiolos exerts its growth inhibitory effect remain to be elucidated.
The Aiolos gene is crucial in lymphoid development and apoptosis;
therefore, the function of Aiolos in childhood BCP-ALL and the
downstream regulatory mechanisms of Aiolos require investigation.
In the present study, a lentiviral system was used to achieve
stable overexpression of the Aiolos gene in the Nalm-6 BCP-ALL cell
line. Subsequently, the effects of Aiolos upregulation on the
proliferation, apoptosis and cell cycle distribution of Nalm-6
cells in vitro was examined. Further experiments indicated
that the Akt, protein kinase, signalling pathways were regulated by
loss of phosphatase and tensin homologue deleted on chromosome ten
(PTEN) caused by overexpression of Aiolos.
Materials and methods
Recombinant lentiviral vector
production
Aiolos cDNA (NM_012481; YRgene, Changsha, China) was
cloned into a pWPT-PURO-green fluorescent protein (GFP) plasmid
(Shanghai Telebio Biomedical Co. Ltd., Shanghai, China), which was
co-transfected with three packaging plasmids (pRsv-REV, pMD1g-pRRE
and pMD2G; Shanghai Telebio Biomedical Co. Ltd.) into human
embryonic kidney (HEK)293T cells (Shanghai Telebio Biomedical Co.
Ltd.). The HEK293T cells were plated at 7×105 on
six-well plates (Corning Costar; Corning, Inc., New York, NY, USA)
in 2 ml of DMEM (Invitrogen Life Technologies, Carlsbad, CA, USA)
with 10% foetal bovine serum (FBS; Gibco-BRL, Grand Island, NY,
USA) and no antibiotics, and incubated at 37°C with 5%
CO2, overnight. Subsequently, 10 μg pRsv-REV, 15 μg
pMDlg-pRRE and 7.5 μg pMD2G packaging plasmids diluted in 1600 μl
of Opti-MEM-I (Invitrogen Life Technologies) were combined, and 20
μg pWPT-PURO-GFP-Aiolos lentiviral DNA diluted in 200 μl of
Opti-MEM-I was added to the tube. Finally 200 μl CaCl2
(2.5 mol/l) was added to the tube, directly into the media to bring
the final reaction volume to 2000 μl. The tube was agitated and
then incubated at room temperature for 30 min. The reaction mixture
(300 μl) was added to each well of the HEK293T cells plated the
previous day, and incubated at 37°C, 5% CO2, overnight.
The following day the media was aspirated and replaced with 2 ml
fresh DMEM containing 10% FBS and 1% PenStrep (Invitrogen Life
Technologies), and incubated overnight. The virions, released into
the media, were collected 48 and 72 h after transfection. To
concentrate the viral particles, the media were ultracentrifuged
twice at 106,000 × g (1.5 h/round) and the final pellet was
dissolved in RPMI 1640 (Invitrogen Life Technologies), 10% FBS and
1% glutamine (Invitrogen Life Technologies). Lenti-GFP was
constructed similarly, but without the Aiolos insert.
Cell culture and transfection
The BCP-ALL cell line Nalm-6 was purchased from the
American Type Culture Collection. (Manassas, VA, USA) and cultured
in standard RPMI 1640 medium (Gibco-BRL) containing 10% foetal
bovine serum (Gibco-BRL) and 1% penicillin-streptomycin (Gibco-BRL)
at 37°C in 5% CO2. The Nalm-6 cells were divided into
the following three groups: Untransfected control (UT), Lenti-GFP
group and Aiolos-transfected (Lenti-Aiolos) group. The viral
concentrate was diluted in polybrene (5 μg/ml; Sigma-Aldrich, St.
Louis, MO, USA) to infect the Nalm-6 cells at a multiplicity of
infection of 100. Successful transduction was confirmed by
visualisation of enhanced GFP after 4 days using a fluorescence
microscope (IX71; Olympus Corp., Tokyo, Japan) and Phenix
micro-image analysis software version 2.2 (Phenix Optical Holding
Stock Co., Ltd, Jiangxi, China). The cells were maintained and
allowed to grow at 37°C for an additional 6 days, following which
the expression levels of Aiolos were confirmed by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blot analysis. The virus-infected cells were selected with
8 μg/ml puromycin (Invitrogen Life Technologies, Carlsbad, CA,
USA). The antibiotic-resistant clones were pooled and used for
subsequent assays.
Treatment with Akt inhibitor
Akti-1/2, previously known as Akt-I-1/2, was
purchased from Sigma-Aldrich and reconstituted in dimethyl
sulfoxide (DMSO; Sigma-Aldrich) to a stock concentration of 50 mM.
The final concentration of DMSO in the cultures was maintained at
0.1%. For all the experiments, the cells (2×105/ml) were
treated with 0.5 μM Akti-1/2 at 37°C for 48 h and treatment was
continued for subsequent experiments. The control cells were
treated with equal quantities of the solvent. All experiments were
performed in triplicate.
Cell growth curve
The Nalm-6 cells of the UT, Lenti-GFP and
Lenti-Aiolos groups were seeded into 24-well plates (Corning
Costar) in triplicate at a density of 2×105 per well at
37°C seven days after transfection. The number of live cells was
determined by trypan blue staining (Sigma-Aldrich) and was measured
daily for 5 days using a haemocytometer (Jingmai Biotech., Ji’nan,
China) under an inverted microscope (IX71; Olympus Corp., Tokyo,
Japan).
Cell cycle analysis
For cell cycle analysis, a total of 1×106
cells were first fixed with1 ml ice-cold 70% ethanol and then
incubated with RNase A at 37°C for 30 min. The cells were stained
with propidium iodide (PI; 50 μg/ml PI in 0.1% sodium citrate and
0.1% Nonidet P-40; Sigma-Aldrich) for 30 min at 4°C. The cells were
then analysed using a fluorescence-activated cell sorting
(FACS)Calibur instrument equipped with Cell Quest software version
5.0 (BD Biosciences, Franklin Lakes, NJ, USA). The results are
expressed as the percentage of the cells in each cell cycle phase
and error bars indicate the standard deviation of the mean
(SEM).
Detection of apoptosis
Annexin V/PI labelling was performed using the
Vybrant Apoptosis Assay kit #3 (Invitrogen Life Technologies),
according to the manufacturer’s instructions, followed by flow
cytometry. The Nalm-6 cells were scanned in fluorescein
isothiocyanate (FL1-H), vs. PI (FL2-H) channels using a FACSCalibur
instrument and Cell Quest software version 5.0.
Isolation of RNA and RT-qPCR
analysis
Total RNA was extracted from each group of Nalm-6
cells using TRIzol reagent (Invitrogen Life Technologies),
according to the manufacturer’s instructions. Purified total RNA (1
μg) was reverse transcribed to cDNA using the Omniscript cDNA
synthesis kit (Qiagen, Hamburg, Germany) using random primers. The
relative gene expression levels were measured by qPCR using
gene-specific primers and SYBR Green supermix (Promega Corp.,
Madison, WI, USA). β-actin was used as an endogenous control. The
primers used are listed in Table
I.
 | Table IPrimers used in reverse
transcription-quantitative polymerase chain reaction. |
Table I
Primers used in reverse
transcription-quantitative polymerase chain reaction.
| Gene |
Forward/reverse | Sequence | Product (bp) |
|---|
| PTEN | Forward |
5′-GTGGCGGAACTTGCAATCCT-′3 | 119 |
| Reverse |
5′-CGGCTGAGGGAACTCAAAGT-′3 |
| Akt1 | Forward |
5′-ATGGACAGGGAGAGCAAACG-′3 | 134 |
| Reverse |
5′-CTGGCCACAGCCTCTGATG-′3 |
| Akt2 | Forward |
5′-GGTGACAGACTGTGCCCTG-′3 | 81 |
| Reverse |
5′-GGTACGCTGTCACCTAGCTC-′3 |
| Akt3 | Forward |
5′-TTGGTTCGAGAGAAGGCAAGT-′3 | 87 |
| Reverse |
5′-GTGTGCCACTTCATCCTTTGC-′3 |
| GAPDH | Forward |
5′-ATAAATTGAGCCCGCAGCC-′3 | 140 |
| Reverse |
5′-ACCAAATCCGTTGACTCCGA-′3 |
Western blot analysis
The proteins were extracted using
radioimmunoprecipitation assay buffer containing 1×
phosphate-buffered saline, 1% Nonidet P-40, 0.5% sodium
dexoycholate, 0.1% SDS and protease inhibitor mixture tablet (Roche
Diagnostics GmbH, Mannheim, Germany). At least 20 μg/lane proteins
were subjected to 12% SDS-PAGE and subsequently transferred to
polyvinylidene fluoride hydrophobic membranes (EMD Millipore,
Bedford, MA, USA). Membranes were blocked with 5% nonfat dry milk
(Shifeng Biotech) in Tris-buffered saline containing 0.05% Tween-20
(TBST; Sigma-Aldrich) at 4°C for 2 h. The membranes were rinsed
with TBST and incubated with rabbit monoclonal Aiolos antibody
(1/20;000; cat. 39408; Abcam, Cambridge, MA, USA), rabbit
monoclonal PTEN antibody (1/500; cat. 32199; Abcam), rabbit
monoclonal Akt (1/10,000; cat. 32505; Abcam), rabbit polyclonal
phosphorylated-Akt antibody (Thr-308, 1/2,000; cat. 66134; Abcam)
and rabbit monoclonal β-actin (1/20,000; cat. 79467; Abcam).
Horseradish peroxidase-conjugated secondary antibodies (goat
anti-rabbit immunoglobulin G H&L-HRP; 1/10,000; cat. 16721;
Abcam) and an Enhanced Chemiluminescence (ECL) Plus Immunoblotting
Detection system (Beyotime, Shanghai, China) were used to detect
specific binding, and images of the signals were captured using
X-ray films (Kodak, Rochester, NY, USA) and WD-9413C
Electrophoresis image analysis system (Liuyi, Beijing, China).
Statistical analysis
All data are presented as the mean ± standard
deviation and analysed via analysis of variance, which was followed
by Student’s t-test. P<0.05 was considered to indicate a
statistically significant difference. All statistical analyses were
conducted using the SPSS 13.0 software program (SPSS Inc., Chicago,
IL, USA).
Results
Aiolos overexpression by stable
transfection in Nalm-6 cells
The Nalm-6 cells were infected with the
pWPT-PURO-GFP-Aiolos lentivirus vector, which was inserted with the
entire Aiolos coding sequence, to induce the upregulation of
Aiolos. As a control, Nalm-6 cells were either infected with a
lentivirus vector expressing GFP or were not infected. At 4 days
after infection, the infection efficiency was determined based on
the expression level of GFP using a fluorescence microscope. A
substantial number of cells emitted bright green fluorescence,
suggesting high infection efficiency (Fig. 1A–F). The cells were maintained in
culture and grown for an additional 3 days. Subsequently, the mRNA
and protein expression levels of Aiolos in the Nalm-6 cells of the
three groups were determined by RT-qPCR and western blot assays.
The RT-qPCR results demonstrated that the mRNA expression of
Aiolos in the Nalm-6 cells of the Lenti-Aiolos group was
1.5-fold higher compared with that in the Nalm-6 cells of the UT
group (P<0.01; Fig. 1G). This
result is consistent with the increase in the protein expression of
Aiolos (Fig. 1H). No significant
difference was observed in the distribution of expression of Aiolos
between the Lenti-GFP group and UT group.
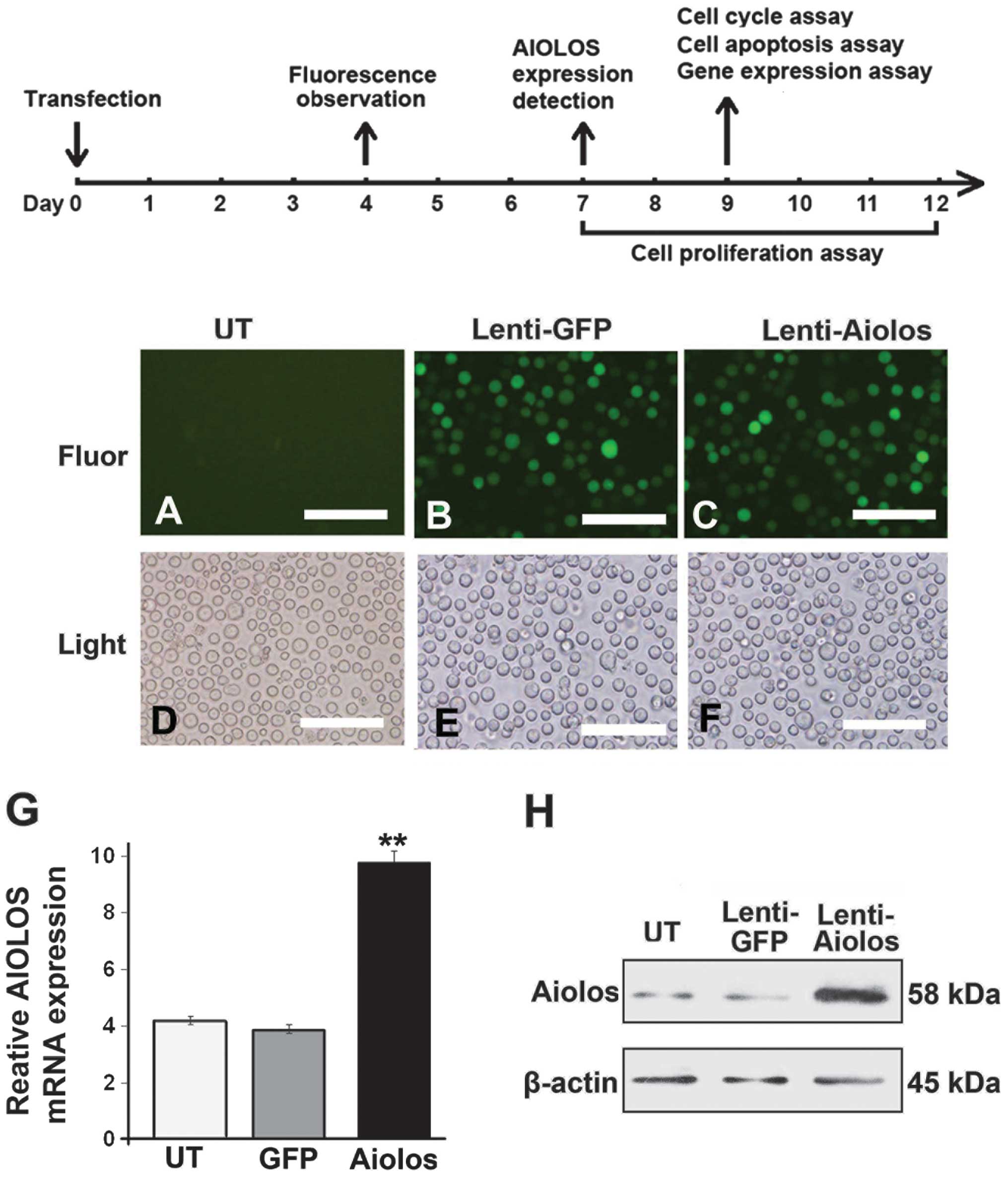 | Figure 1Timeline of the treatment of Nalk-6
cells involving the overexpression of Aiolos. (A–F) Lentiviral
transduction efficiency in the Nalm-6 cells. Transduction
efficiency was estimated 4 days after infection at a multiplicity
of infection of100. The expression of GFP was observed under (A, B
and C) fluorescence or (D, E and F) light microscopy. Scale bar:
200 μm (magnification, ×200). (G) Reverse
transcription-quantitative polymerase chain reaction of the
relative transcription levels of Aiolos in the UT, Lenti-GFP, and
Lenti-Aiolos groups. β-actin was used as an internal control. (H)
Western blotting results revealed that protein expression of Aiolos
was present in the Nalm-6 cells of the three groups. β-actin was
used as a loading control. Error bars indicate the standard
deviation of the mean of three experiments. **P<0.01
Lenti-Aiolos group vs. Lenti-GFP group. GFP, green fluorescent
protein; UT, untransfected control; Lenti-GFP, lentiviral vector
control; Lenti-Aiolos, Aiolos-transfected; Fluor, fluorescence. |
Overexpression of Aiolos inhibits the
growth of Nalm-6 cells and leads to cell cycle arrest at the G0/G1
phase
The growth properties of the Lenti-Aiolos group were
measured and compared with those of the control Lenti-GFP
transfected group to determine whether overexpression of Aiolos
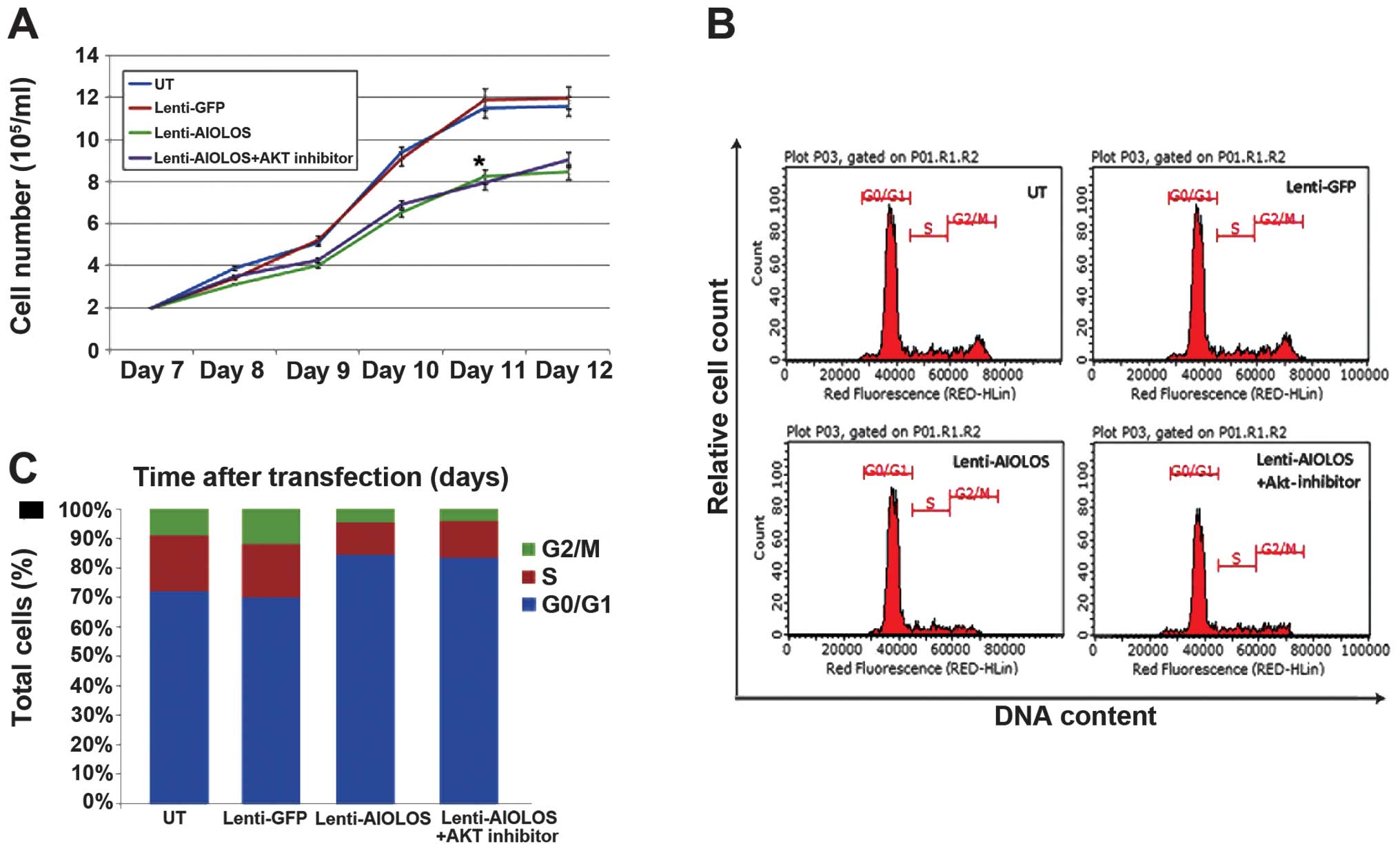
affects the proliferation of Nalm-6 cells. As shown in Fig. 2A, the growth rate of the
Lenti-Aiolos group was significantly lower compared with that of
the Lenti-GFP or UT group. This result suggested that the
upregulation of Aiolos may inhibit the growth of Nalm-6 cells.
The resulting inhibition of the proliferation of
Nalm-6 cells infected with Lenti-Aiolos compared with control
Nalm-6 cells may be, in part, caused by differences in cell cycle
regulation. Thus, the cell cycles of Nalm-6 cells in different
groups were characterised by FACS analysis 9 days after
transfection. No significant difference was detected between the
Lenti-GFP and Nalm-6 cells (Fig.
2B; P>0.05). However, the percentage of Nalm-6 cells in the
G0/G1 phase increased between 72% (UT) and 84.5% (Lenti-Aiolos;
P<0.01), and the percentage of Nalm-6 cells in the S-phase cells
decreased between 19% (UT) and 10.9% (Lenti-Aiolos; P<0.01). A
significant difference in the percentage of Nalm-6 cells in the
G2/M phase was found between the Lenti-Aiolos group and the UT
group (4.5. vs. 9%, respectively; P<0.01). These data indicated
that Aiolos upregulation arrested an increased number of Nalm-6
cells at the G0/G1 phase, which possibly contributed to the growth
inhibition observed.
Aiolos overexpression suppresses the
apoptosis of Nalm-6 cells
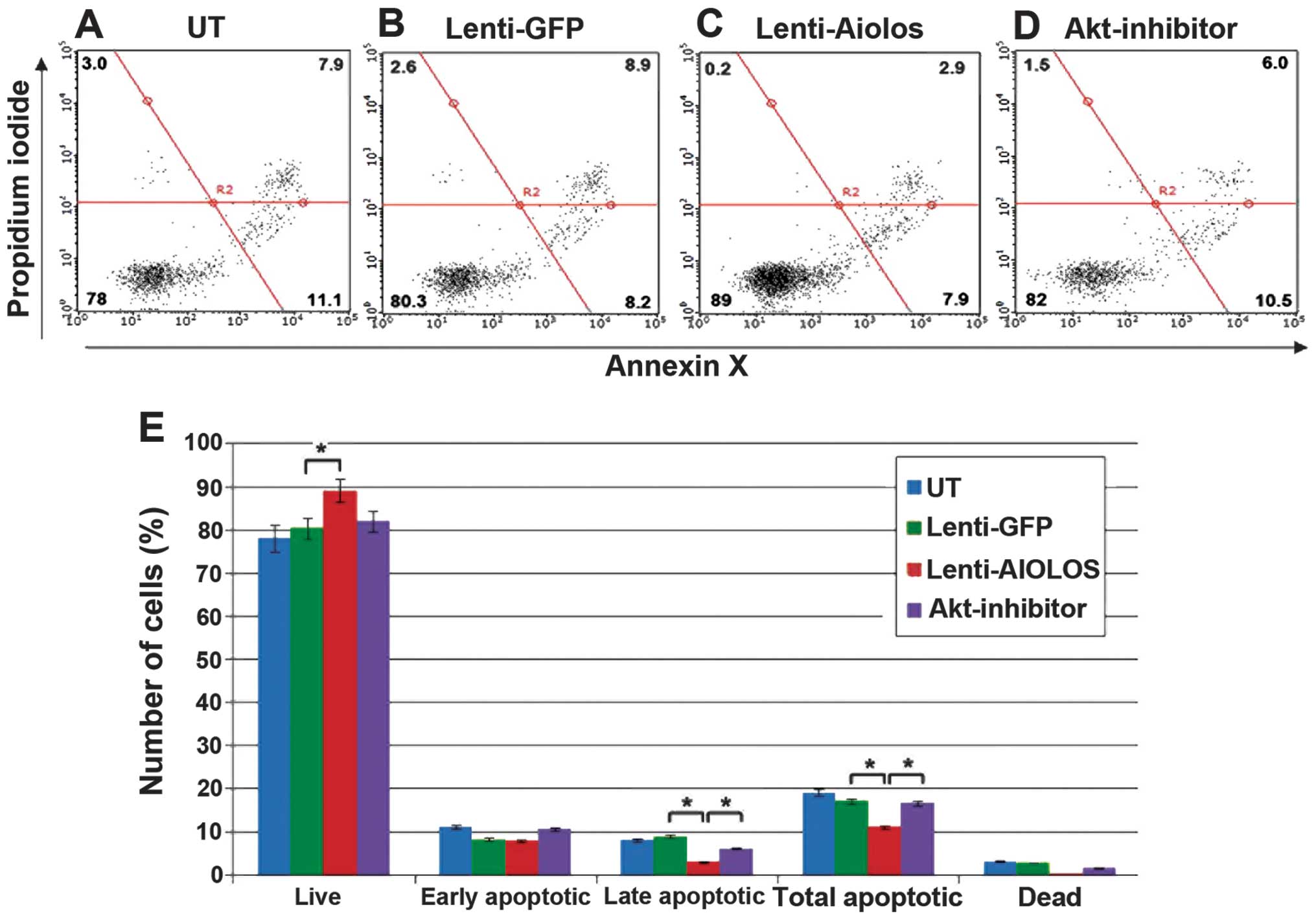
An apoptosis assay kit was used to measure cell
apoptosis 9 days after transfection to determine whether
overexpression of Aiolos affects the apoptosis of Nalm-6 cells. As
shown in Fig. 3, the total number
of apoptotic cells was significantly lower in the
Aiolos-transfected Nalm-6 cells (11%) compared with the cells in
the Lenti-GFP (16.5%) or UT groups (18.9%; P<0.05). A minimal
difference in the percentage of early apoptotic cells was found
between the Aiolos-transfected and UT Nalm-6 cells (8.05, vs.
11.2%; P>0.05), whereas a significant difference was observed in
the percentage of late apoptotic cells between the cell groups
(2.95, vs. 7.7%; P<0.05). The obtained data suggested that
overexpression of Aiolos may suppress apoptosis in Nalm-6
cells.
Upregulated Aiolos suppresses the
expression of PTEN, but activates certain
phosphatidylinositol-3-kinase (PI3K)/Akt signalling
pathway-associated genes in Nalm-6 cells
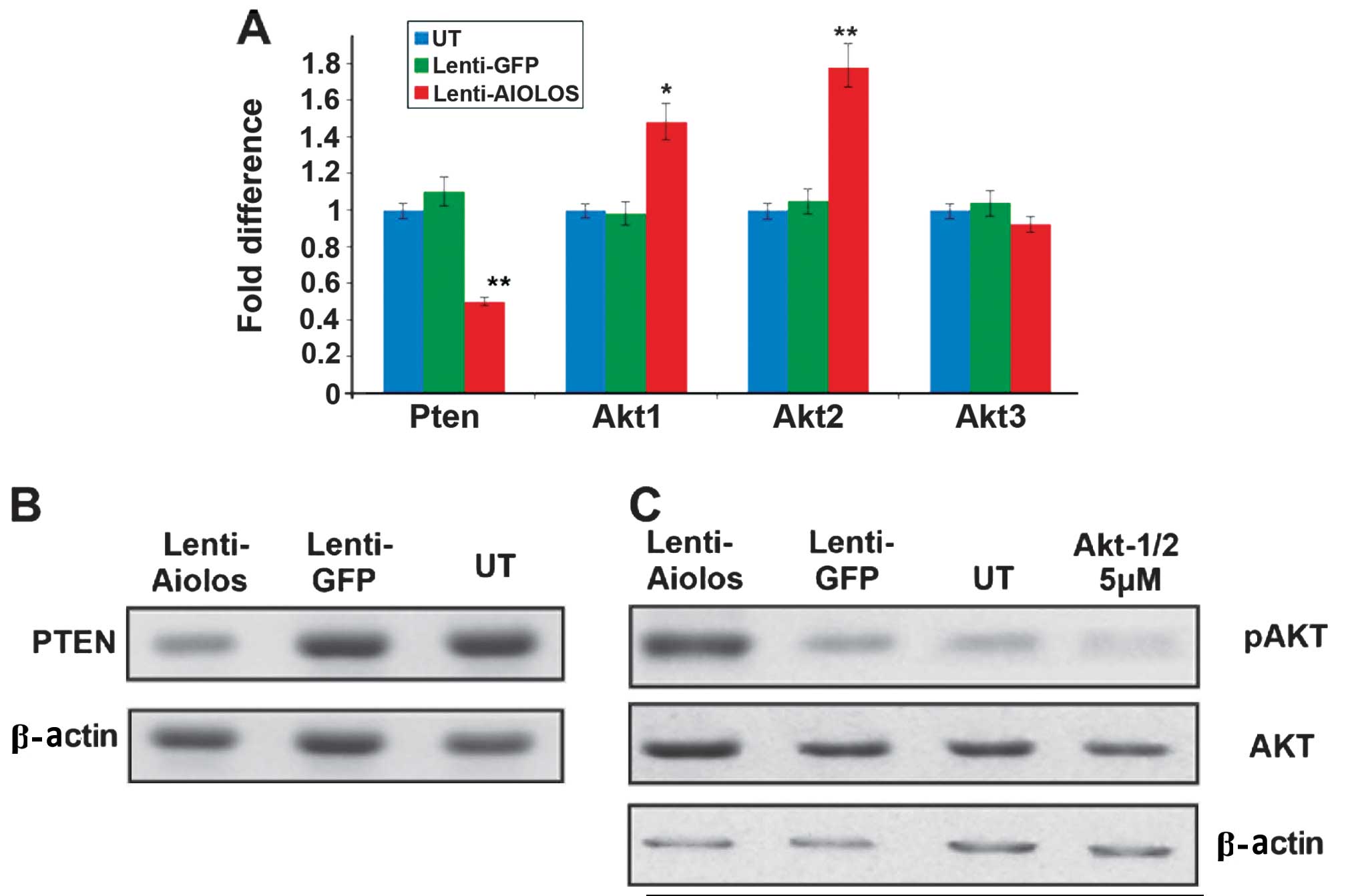
Microarrays were used to analyse the gene expression
of Nalm-6 cells transfected with Aiolos and to further examine the
underlying mechanism responsible for the aforementioned changes in
biological behaviour (data not shown). Among the deregulated genes,
several key genes are of note. The expression levels of PTEN and
certain Akt family genes were further detected using RT-qPCR and
western blot analysis. The RT-qPCR results revealed that the
expression level of PTEN in the Nalm-6 cells was significantly
lower in the Lenti-Aiolos group compared with the control group
(P<0.05). In addition, upregulation of Akt1 (P<0.05) and Akt2
(P<0.01) was confirmed in the Lenti-Aiolos group. No significant
change was observed in the expression of Akt3. Consistent with the
RT-qPCR results, decreased expression levels of PTEN was observed
by western blot analysis. The protein expression of Akt did not
change considerably, whereas the levels of phosphorylated Akt were
significantly higher in the Lenti-Aiolos group compared with the
control group (Fig. 4).
Akt inhibitors reverse Aiolos
overexpression-induced apoptosis, but do not affect proliferation
arrest
The cells were incubated with varying concentrations
of the inhibitor, Akti-1/2, (0.1–10 μM) for 48 h and cell viability
was measured to verify whether the activation of Akt was
principally responsible for the decreased apoptosis in the
Aiolos-overexpressed Nalm-6 cells. The results of the western
blotting revealed that Akti-1/2, at a concentration of 5 μM,
effectively inhibited Akt phosphorylation. In addition, Akti-1/2
effectively increased the percentage of apoptotic cells in the
Aiolos-transfected Nalm-6 cells (Fig.
3). However, in the cell proliferation experiments, Akti-1/2
treatment did not improve the growth of the Nalm-6 cells in the
Lenti-Aiolos group (Fig. 2A).
Similarly, Akti-1/2 did not affect the cell cycle of the
Lenti-Aiolos group (Fig. 2B and
C).
Discussion
In the present study, the Nalm-6 BCP-ALL cell line
was used for a series of functional investigations to understand
the function of Aiolos in the pathogenesis of BCP-ALL. The results
demonstrated that lentivirus-mediated overexpression of Aiolos
suppressed cell apoptosis and arrested the cell cycle at the G0/G1
phase, which possibly contributed to growth inhibition of the
Nalm-6 cells. Furthermore, these changes were possibly associated
with downregulation of PTEN and phosphorylation of Akt.
At least 16 different isoforms of Aiolos exist,
resulting from alternate splicing, have different cellular
localisations and have the ability to alter the localisation of
other Ikaros members (13).
However, the AIO1 transcript comprises >80% of the Aiolos
isoforms in B-cells (14). To
mimic the isoforms and cellular localizations of Aiolos in B-cells,
the pWPT-PURO-GFP-Aiolos plasmid, containing the entire Aiolos
coding sequence, was constructed and lentivirus-mediated
transduction of Nalm-6 cells was performed to create a stable
transfection cell line. The expression of the Aiolos isoform has
been previously investigated in lymphoid development, however, its
expression in unaffected Nalm-6 cells remains to be elucidated. In
the present study, untransfected Nalm-6 cells expressed low levels
of Aiolos at the RNA and protein levels. The results indicated that
Nalm-6 cells were successfully transduced with the lentivirus and
that Aiolos was successfully overexpressed in the Nalm-6 cells.
Aiolos is undoubtedly important in the control of
mature B-lymphocyte differentiation and proliferation (15). Aiolos silences the expression of
the surrogate light-chain gene λ5, thereby downregulating the
pre-B-cell receptor (pre-BCR) (16). In addition, Aiolos directly binds
to the c-Myc promoter and represses the expression of c-Myc in
pre-B cells. Repression of c-Myc by Aiolos subsequently leads to
the expression of p27 and the downregulation of cyclin D3 (17). P27 is a type of cyclin-dependent
kinase inhibitor protein, which inhibits G1/S conversion and
negatively regulates the cell cycle by inhibiting the functions of
cell cycle regulators, including cyclin D3 (18,19).
By contrast, pre-BCR, cyclin D3 and c-Myc are necessary for the
proliferation of pre-B-cells (20–22).
Consistent with results of previous studies, the present study
demonstrated that overexpression of Aiolos suppressed proliferation
of Nalm-6 cells. In addition, the results of cell cycle analysis
indicated that upregulation of Aiolos arrested the Nalm-6 cells at
the G0/G1 phase.
Previous tudies have confirmed that overexpression
of Aiolos in Nalm-6 cells inhibits cell apoptosis, which is
consistent with previous studies demonstrating that disruption of
Aiolos accelerates premature B cell apoptosis, mediated by BCR
signalling through elevation in the release of cytochrome c
(23,24). However, different findings have
been reported regarding the function of cyclin D3 in the apoptotic
process of B cells. Wang et al (25) reported that downregulation of
cyclin D3 increases the number of B-cell CLL cells undergoing
apoptosis and another study suggested that repression of c-Myc and
cyclin D3 is necessary to arrest human leukaemia cells at the G1
phase of the cell cycle, but neither is required for apoptosis
(26). Therefore, in addition to
c-Myc and cyclin D3, Aiolos potentially acts through other
mechanisms to regulate the apoptotic process.
The present results demonstrate the crucial function
of PTEN. The expression of PTEN was significantly reduced in
Aiolos-overexpressed Nalm-6 cells. Previous studies have identified
the functions of PTEN in leukaemogenesis (27–29),
however, the functions of PTEN in association with Aiolos remain to
be elucidated. PTEN was initially identified as a tumour
suppressor, and loss or mutation of PTEN is reported to be
associated with the development of various types of cancer
(30,31). PTEN may exert this regulatory role
by altering its own expression and activity in response to external
stimuli, including cigarette smoking or cytokines (32–35).
PTEN primarily acts downstream of various other pathways as a
negative regulator of the PI3K pathway due to its lipid phosphatase
activity. Loss of PTEN increases the level of PI3-phosphate,
thereby mimicking the effect of constitutive PI3K activation
(36–38). Accumulation of PI3-phosphate
activates various protein kinases, including Akt kinases (39). In addition to the reduction in PTEN
levels, the results of the present study demonstrated that the
expression and phosphorylation of Akt increased. This was
consistent with the findings of the previously mentioned studies
(40). Akt is the central node in
the PTEN-regulated pathway, and activated Akt can act on multiple
downstream targets, which are involved in proliferation, cell
metabolism and apoptosis (40).
Thus, improved cell survival may be attributed to the activated Akt
pathway in the Aiolos-overexpressed Nalm-6 cells.
Previous studies have demonstrated that, in addition
to inhibiting apoptosis, activated Akt can promotes cell cycle
progression and cell growth (41–43).
However, the present study revealed that overexpression of Aiolos
inhibited the growth of Nalm-6 cells and arrested the cells at the
G0/G1 phase. Thus, the loss of PTEN may be insufficient to provide
a proliferative advantage in BCP-ALL cells. Downregulation of c-Myc
and cyclin D3 was sufficient for growth inhibition in the
Aiolos-overexpressed Nalm-6 cells, despite proliferation signals,
driven by the loss of PTEN. Subsequently, chemical inhibitors of
Akt (Akti-1/2) were used to verify whether Akt is a key regulatory
factor of apoptosis or growth regulation. The findings indicated
that inhibiting the phosphorylation of Akt promoted the apoptosis
of Aiolos-overexpressed Nalm-6 cells. Akti-1/2 inhibited Akt,
however, it did not induce changes in proliferation or the cell
cycle. The results further demonstrated that overexpression of
Aiolos induced changes in PTEN, and that Akt affected BCP-ALL cell
apoptosis. Certain previous studies have indicated that loss of
PTEN, which positively regulates Akt signalling, leads to the
expansion of leukaemic stem cell subpopulations (44). However, loss of PTEN does not exert
a positive effect in promoting a short-term increase in the total
number of cells.
In conclusion, the results of the present study
indicated that upregulation of the expression of Aiolos in Nalm-6
cells inhibited cell proliferation, suppressed apoptosis and
arrested the cell cycle at the G0/G1 phase. These findings revealed
two critical functions of Aiolos in Nalm-6 cells; to limit
pre-B-cell expansion via c-Myc and cyclin D3-dependent pathways and
to improve pre-B cell survival via PTEN- and Akt-dependent
processes. Characterising these potential genetic interactions may
provide a foundation for further investigation of the pathogenesis
of BCP-ALL.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (no. 81102710), the Science and
Technology Plan of Shandong Province (nos. 2009GG10002041 and
2009GG2HZ02003) and the Independent Innovation Foundation of
Shandong University (no. 2012TS149).
References
|
1
|
Kaatsch P: Epidemiology of childhood
cancer. Cancer Treat Rev. 36:277–285. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pui CH, Robison LL and Look AT: Acute
lymphoblastic leukaemia. Lancet. 371:1030–1043. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cobaleda C and Sánchez-Garcia I: B-cell
acute lymphoblastic. leukaemia: towards understanding its cellular
origin. Bioessays. 31:600–609. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Georgopoulos K, Moore DD and Derfler B:
Ikaros, an early lymphoid-specific transcription factor and a
putative mediator for T cell commitment. Science. 258:808–812.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kelley CM, Ikeda T, Koipally J, et al:
Helios, a novel dimerization partner of Ikaros expressed in the
earliest hematopoietic progenitors. Curr Biol. 8:508–515. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Morgan B, Sun L, Avitahl N, et al: Aiolos,
a lymphoid restricted transcription factor that interacts with
Ikaros to regulate lymphocyte differentiation. EMBO J.
16:2004–2013. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Georgopoulos K, Winandy S and Avitahl N:
The role of the Ikaros gene in lymphocyte development and
homeostasis. Annu Rev Immunol. 15:155–176. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang JH, Avitahl NA, Cariappa C, et al:
Aiolos regulates B cell activation and maturation to effector
state. Immunity. 9:543–553. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakase K, Ishimaru F, Avitahl N, et al:
Dominant negative isoform of the Ikaros gene in patients with adult
B-cell acute lymphoblastic leukemia. Cancer Res. 60:4062–4065.
2000.PubMed/NCBI
|
|
10
|
Nuckel H, Frey UH, Sellmann L, et al: The
IKZF3 (Aiolos) transcription factor is highly upregulated and
inversely correlated with clinical progression in chronic
lymphocytic leukaemia. Br J Haematol. 144:268–270. 2009. View Article : Google Scholar
|
|
11
|
Billot K, Soeur J, Chereau F, et al:
Deregulation of Aiolos expression in chronic lymphocytic leukemia
is associated with epigenetic modifications. Blood. 117:1917–1927.
2011. View Article : Google Scholar
|
|
12
|
Antica M, Cicin-Sain L, Kapitanovic S, et
al: Aberrant Ikaros, Aiolos and Helios expression in Hodgkin and
non-Hodgkin lymphoma. Blood. 111:3296–3297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Caballero R, Setien F, Lopez-Serra L, et
al: Combinatorial effects of splice variants modulate function of
Aiolos. J Cell Sci. 120:2619–2630. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Duhamel M, Arrouss I, Merle-Béral H, et
al: The Aiolos transcription factor is up-regulated in chronic
lymphocytic leukemia. Blood. 111:3225–3228. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cobb BS and Smale ST: Ikaros-family
proteins: in search of molecular functions during lymphocyte
development. Curr Top Microbiol Immunol. 290:29–47. 2005.
|
|
16
|
Thompson EC, Cobb BS, Sabbattini P, et al:
Ikaros DNA-binding proteins as integral components of B-cell
developmental-stage-specific regulatory circuits. Immunity.
26:335–344. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma S, Pathak S, Mandal M, et al: Ikaros
and Aiolos inhibit pre-B-cell proliferation by directly suppressing
c-Myc expression. Mol Cell Biol. 30:4149–4158. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Myung DS, Park YL, Chung CY, et al:
Expression of livin in colorectal cancer and its relationship to
tumor cell behavior and prognosis. PLoS One. 8:e732622013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Toyoshima H and Hunter T: P27, a novel
inhibitor of G1 cyclin-Cdk protein kinase activity, is related to
p21. Cell. 78:67–74. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Herzog S, Reth M and Jumaa H: Regulation
of B-cell proliferation and differentiation by pre-B-cell receptor
signaling. Nat Rev Immunol. 9:195–205. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cooper AB, Sawai CM, Sicinska E, et al: A
unique function for cyclin D3 in early B cell development. Nat
Immunol. 7:489–497. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Habib T, Park H, Tsang M, et al: Myc
stimulates B lymphocyte differentiation and amplifies calcium
signaling. J Cell Biol. 179:717–731. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Narvi E, Nera KP, Terho P, et al: Aiolos
controls gene conversion and cell death in DT40 B cells. Scand J
Immunol. 65:503–513. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kikuchi H, Yamashita K, Nakayama M, et al:
Lacking of Aiolos accelerates pre-mature B cell apoptosis mediated
by BCR signaling through elevation in cytochrome c release. Biochim
Biophys Acta. 1793:1304–1314. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang P, Pavletic ZS and Joshi SS:
Increased apoptosis in B-chronic lymphocytic leukemia cells as a
result of cyclin D3 down regulation. Leuk Lymphoma. 43:1827–1835.
2002. View Article : Google Scholar
|
|
26
|
Ausserlechner MJ, Obexer P, Böck G, et al:
Cyclin D3 and c-MYC control glucocorticoid-induced cell cycle
arrest but not apoptosis in lymphoblastic leukemia cells. Cell
Death Differ. 11:165–174. 2004. View Article : Google Scholar
|
|
27
|
Shehata M, Schnabl S, Demirtas D, et al:
Reconstitution of PTEN activity by CK2 inhibitors and interference
with the PI3-K/Akt cascade counteract the antiapoptotic effect of
human stromal cells in chronic lymphocytic leukemia. Blood.
116:2513–2521. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhiyong C, Wentong L, Xiaoyang Y, et al:
PTEN Regulates VEGF, VEGFR1 Expression and Its Clinical
Significance in Myeloid Leukemia. Blood (ASH Annual Meeting
Abstracts). 114:Abstract 1001. 2009.
|
|
29
|
Gutierrez A, Sanda T, Grebliunaite R, et
al: High frequency of PTEN, PI3 K and AKT abnormalities in T-cell
acute lymphoblastic leukemia. Blood. 114:647–650. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Karoui M, Tresallet C, Julie C, et al:
Loss of heterozygosity on 10q and mutational status of PTEN and BM
PR1A in colorectal primary tumours and metastases. Br J Cancer.
90:1230–1234. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Poetsch M, Lorenz G and Kleist B:
Detection of new PTEN/MM AC 1 mutations in head and neck squamous
cell carcinomas with loss of chromosome 10. Cancer Genet Cytogenet.
132:20–24. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hou R, Zhang J, Yin T, et al: Upregulation
of PTEN by peroxynitrite contributes to cytokine-induced apoptosis
in pancreatic beta-cells. Apoptosis. 15:877–886. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Koul D, Yao Y, Abbruzzese JL, et al: Tumor
suppressor MMAC/PTEN inhibits cytokine-induced NFkappa B activation
without interfering with the IkappaB degradation pathway. J Biol
Chem. 276:11402–11408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee SY, Kang EJ, Hur GY, et al: Peroxisome
proliferator-activated receptor-gamma inhibits cigarette smoke
solution-induced mucin production in human airway epithelial
(NCI-H292) cells. Am J Physiol Lung Cell Mol Physiol. 291:L84–L90.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim S, Domon-Dell C, Kang J, et al:
Down-regulation of the tumor suppressor PTEN by the tumor necrosis
factor-alpha/nuclear factor-kappaB (NF-kappaB)-inducing
kinase/NF-kappaB pathway is linked to a default IkappaB-alpha
autoregulatory loop. J Biol Chem. 279:4285–4291. 2004. View Article : Google Scholar
|
|
36
|
Maehama T and Dixon JE: The tumor
suppressor, PTEN/MMAC1, dephosphorylates the lipid second
messenger, phosphatidylinositol 3,4,5-trisphosphate. J Biol Chem.
273:13375–13378. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Stambolic V, Suzuki A, de la Pompa JL, et
al: Negative regulation of PKB/Akt-dependent cell survival by the
tumor suppressor PTEN. Cell. 95:29–39. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun H, Lesche R, Li DM, et al: PTEN
modulates cell cycle progression and cell survival by regulating
phosphatidylinositol 3,4,5,-trisphosphate and Akt/protein kinase B
signaling pathway. Proc Natl Acad Sci USA. 96:6199–6204. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gold MR, Scheid MP, Santos L, et al: The B
cell antigen receptor activates the Akt (protein kinase B)/glycogen
synthase kinase-3 signaling pathway via phosphatidylinositol
3-kinase. J Immunol. 163:1894–1905. 1999.PubMed/NCBI
|
|
40
|
Leslie NR and Downes CP: PTEN function:
how normal cells control it and tumour cells lose it. Biochem J.
382:1–11. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Datta SR, Dudek H, Tao X, et al: Akt
phosphorylation of BAD couples survival signals to the
cell-intrinsic death machinery. Cell. 91:231–241. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Salmena L, Carracedo A and Pandolfi PP:
Tenets of PTEN tumor suppression. Cell. 133:403–414. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Perry JM, He XC, Sugimura R, et al:
Cooperation between both Wnt/{beta}-catenin and PTEN/PI3 K/Akt
signaling promotes primitive hematopoietic stem cell self-renewal
and expansion. Genes Dev. 25:1928–1942. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tothova Z and Gilliland DG: FoxO
transcription factors and stem cell homeostasis: Insights from the
hematopoietic system. Cell Stem Cell. 16:140–152. 2007. View Article : Google Scholar
|