Introduction
The concept that ‘tumor growth is
angiogenesis-dependent’ was initially proposed by Folkman in 1971
(1), and suggests that the growth
and metastasis of tumors depend on the development of blood
vessels. In the early stages of cancer, the uncontrolled
proliferation of cancer cells leads to a shortage of nutrients and
oxygen, which causes a large degree of cell death (2,3). In
response to the abnormal microenvironment, angiogenic molecules
secreted by the tumor stimulate the formation of new functional
vessels, from the pre-existing vasculature (4). Once the tumor cells acquire the
ability to induce angiogenesis, tumor expansion is initiated, and
tends to be malignant (5).
Therefore, inhibiting the angiogenic process may be used as a
potential therapy to prevent tumor growth and metastasis. Among the
angiogenic factors, vascular endothelial growth factor-A165
(VEGF-A165, commonly referred to as VEGF) is positively associated
with the proliferation, migration and tube formation of endothelial
cells (6,7).
Upon VEGF binding to VEGF receptor 2 (R2), the main
functional receptor tyrosine kinases of VEGF are activated, and a
cascade of events are initiated, including the phosphoinositide
3-kinase (PI3K)/protein kinase B (Akt) and RAF/MEK/ERK signaling
pathways, which stimulate endothelial cell proliferation, migration
and tube formation (8). Numerous
studies have demonstrated that focal adhesion kinase (FAK), and its
substrate paxillin, are involved in the signal transduction that
participates in focal adhesion during cell migration (9–11).
Albizia julibrissin (AJ) (Leguminosae)
is a Traditional Chinese Medicine that was recorded as an
anti-inflammatory and sedative drug to treat swelling and pain in
the lungs, shin ulcers, wounds and for the removal of carbuncles
(12). In modern pharmacology, AJ
exhibits marked inhibitory activity against certain cancer cell
lines in vitro (13,14),
which suggested that it may be used as an anti-tumor agent. In our
previous study it was demonstrated that the crude extract of AJ
exhibits anti-angiogenic effects on 3B11 and HMEC-1 cells (15). Further studies focused on the
anti-angiogenic effects of AJ revealed that the active ingredients
are the saponins. The total saponins from AJ (TSAJ), whose major
constituents include julibroside J, julibroside A, julibroside B1,
julibroside C1, julibroside I, julibroside II and julibroside III
(16–20), have been shown to possess
anti-angiogenic and anti-tumor activities; however, the underlying
mechanisms of action remain to be elucidated. The present study
investigated the anti-angiogenic effects of TSAJ on VEGF-induced
angiogenesis in vitro and in vivo, in order to
explore the potential for TSAJ as an anti-tumor drug targeting the
VEGF/VEGFR2 signaling pathway.
Materials and methods
Preparation of TSAJ
Dried stem barks from AJ (lot no. 20100154) were
obtained from Zhejiang (China) and were identified by Professor
Jian-wei Chen (Chinese Herbal Pharmacy, Nanjing University of
Traditional Chinese Medicine, Nanjing, China). The voucher
specimens were maintained in the herbarium stock room at the
Laboratory of Natural Medicine, School of Pharmaceutical Science,
Jiangnan University (Wuxi, China).
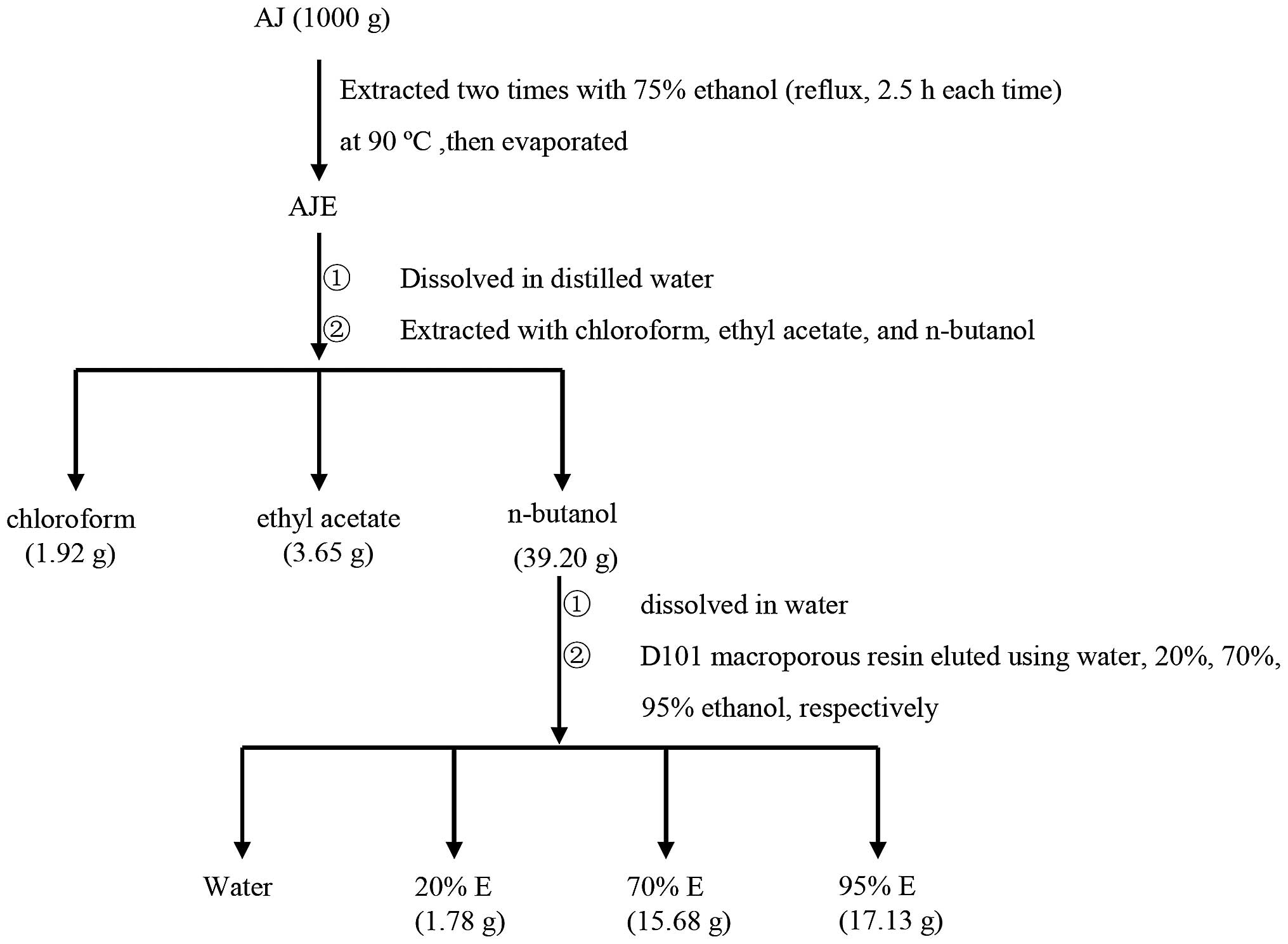
The TSAJ sample used in the present study was
originally isolated from the AJ specimens. Briefly, the powder of
AJ dried stem barks was extracted twice with 75% ethanol (reflux,
2.5 h each time) at 90°C. The ethanol extract was then evaporated,
dissolved in deionized water and sequentially extracted with
chloroform, ethyl acetate and n-butanol. The
n-butanol fraction, which contained the most bioactive
components, was dissolved in water and subjected to D101
macroporous resin (Tianjin Bohong Resin Technology, Tianjin, China)
to yield 20% ethanol (20% E), 70% ethanol (70% E), 95% ethanol (95%
E), and water components (Fig.
1).
Preliminary phytochemical screening of
total saponins
The presence of saponins in the four fractions was
assessed using foam and hemolytic tests.
Foam test
The four dried fractions (10 mg) were placed in a
graduated cylinder with 10 ml distilled water. The suspension was
shaken for 30 sec and a 2–3 cm layer of foam indicated the presence
of saponins.
Hemolytic test
Once the hemaleucin in the anticoagulant whole blood
of rabbit was discarded, the blood was washed three times with 0.9%
normal sodium solution. Subsequently, the prepared erythrocytes
were suspended in 0.9% saline solution to a final concentration of
2% (v/v). The solutions of the four fractions (1 ml) were
separately added to the various erythrocyte suspensions (3 ml), and
incubated at 37°C for 1 h. Normal sodium solution (0.9%) was used
as a control. The hemolytic degree of the four fractions was
assayed by observation. Briefly, following incubation, the mixtures
were then centrifuged at room temperature for 5 min at 495 × g to
separate the supernatant (hemoglobin) and the precipitation
(complete erythrocytes and cell debris). If the solution in the
tube was transparent and red, significant hemolysis phenomena was
shown, thus it indicated the presence of saponins. If the
supernatant was transparent and colorless, and all erythrocytes
were sunk, thus it indicated the absence of saponins (21).
Antibodies and other materials
Human VEGF-A165 was obtained from
PeproTech, Inc. (Rocky Hill, NJ, USA). Matrigel™ was obtained from
BD Biosciences (Franklin Lakes, NJ, USA). Rabbit polyclonal
antibodies targeting β-tubulin (2148S), pTyr1175-VEGFR2
(2478S), pTyr576/Tyr577-Fak (3281S),
pSer473-Akt (8200S) and
pThr202/Thr204-extracellular signal-regulated kinase
(Erk)1/2 (4370S) were purchased from Cell Signaling Technology
(Danvers, MA, USA). Cluster of differentiation 31 (CD31) was
purchased from Sangon Biotech Co., Ltd (Shanghai, China). Goat
anti-rabbit immunoglobulin G (IgG) (H+L) horseradish peroxidase
(HRP)-conjugated antibodies (21621) were purchased from EMD
Millipore (Billerica, MA, USA). Reagents, including ethanol,
chloroform, ethyl acetate, n-butanol, formalin and paraffin
were purchased from China National Medicines Corporation (Beijing,
China). Cell Counting kit-8 (CCK-8) was purchased from Beyotime
Institute of Biotechnology (Haimen, China).
Animals
Female BALB/c mice (17–20 g) were purchased from the
Research Center of Laboratory Animals (Hangzhou, China; grade
specific-pathogen free and certificate no. SCXK (Zhe) 2008-0033).
The mice were maintained in a barrier facility, in a temperature-
and humidity-controlled environment, fed sterilized food and were
given ad libitum access to water. Prior to the experiment,
all of the mice were allowed to acclimate for one week. All
experiments were conducted according to the Guides for the Care and
Use of Laboratory Animals (Ministry of Science and Technology of
China, 2006), and were approved by the Animal Ethics Committees of
Jiangnan University (JN NO 20130327-0702).
Cell culture
The Ea.hy926 human endothelial cell line, generated
from fusion of the A549 epithelial cell line with HUVEC, was
provided by Professor Quan-sheng Zhou (Soochow University, Suzhou,
China). The cells were cultured in cell medium consisting of
Dulbecco’s Modified Eagle Medium (DMEM; Hyclone Laboratories, Inc.,
Logan, UT, USA) supplemented with 10% (v/v) fetal bovine serum
(FBS; Gibco Life Technologies, Carlsbad, CA, USA), in 25
cm2 culture flasks at 37°C in an atmosphere containing
5% CO2.
Determination of anti-angiogenic effects
in vitro
Cell proliferation assay
The viability of the Ea.hy926 cells was assessed
using a CCK-8. Briefly, the Ea.hy926 cells (6×103
cells/well) were plated in 96-well plates (Corning, Inc., Corning,
NY, USA) and cultured in normal growth medium for 24 h. The culture
medium was then replaced with normal growth medium containing
various concentrations of TSAJ (0, 0.78125, 1.5625, 3.125, 6.25,
12.5, 25, and 50 μg/ml), for 12, 24 and 48 h. Subsequently, the
medium was replaced with DMEM containing 10% CCK-8. Following a 2 h
incubation at 37°C, the absorbance of the resulting product was
measured at a wavelength of 450 nm, using an ELISA microplate
reader (Thermo Labsystems, Waltham, MA, USA). The percentage
viability of the cells was then calculated using the following
formula: viability (% of control) =
(ODcontrol-ODtreated)/ODcontrol
ODcontrol and ODtreated represent the average OD450 value of cells
in the control and TSAJ-treated groups, respectively. Three
independent experiments were performed.
The effects of TSAJ on VEGF-induced cell viability
were determined as described by previous methods (22). Briefly, the Ea.hy926 cells
(6×103 cells/well) were seeded in 96-well cell plates
and cultured in normal growth medium for 24 h. Subsequently, the
cells were exposed to various concentrations of TSAJ (0, 0.78125,
1.5625, 3.125, 6.25, 12.5, 25, and 50 μg/ml), with or without VEGF
(10 ng/ml), for 48 h in DMEM supplemented with 5% FBS. The medium
was then replaced with DMEM containing 10% CCK-8. Following a 2 h
incubation at 37°C, the absorbance of the resulting product was
measured at 450 nm, using an ELISA microplate reader (Labsystem,
USA). The percentage viability of the cells was then calculated. At
least three independent experiments were performed. The control
group, which did not receive VEGF or TSAJ treatment, was set at
100%.
Migration activity assay
The migratory ability of the cells was assessed
using a scratch-wound directional assay. Briefly, the Ea.hy926
cells were seeded at a cell density of 8×104 cells/well
in a 24-well cell plate (Corning, Inc.), and grown overnight into a
confluent monolayer. A sterile 20–200 μl micropipette tip
(Axygen®; Corning Life Sciences, Tewksbury, MA, USA) was
then used to create a ‘wound field’ of ±1000 μm width. The cells
were washed twice with phosphate-buffered saline (Boster, Wuhan,
China) and replaced in fresh medium containing the indicated
concentrations of TSAJ, supplemented with 0.5% FBS and VEGF (10
ng/ml).
Following a 14 h incubation, the migrated cells were
photographed using an inverted microscope (TE2000-S; Nikon
Corporation, Tokyo, Japan) with NIS-Elements software. Cell
migration was estimated by measuring the endothelial cells that had
migrated from the edge of the wounded monolayer (23). The percentage of migration was the
mean calculated from five replicates of each experiment. Three
independent experiments were performed. The control group, which
did not receive VEGF or TSAJ treatment, was set at 100%.
Tube formation assay
The effects of TSAJ on the morphogenesis of
endothelial cells were investigated using a capillary tube
formation assay on Matrigel™. Briefly, the Matrigel™ was thawed at
4°C overnight, and 80 μl was then added to a 96-well plate.
Following a 45 min incubation at 37°C, a cell density of
4×104 cells/well was seeded onto the
Matrigel™-pre-coated 96-well plate. The cells were treated with
TSAJ at 4.5 and 9 μg/ml, with or without VEGF (10 ng/ml). Following
6 h, tube formation was visualized and images were captured using
an inverted microscope (TE2000-S; Nikon Corporation). Tube
formation was quantified by counting the number of tubular
structures within the capillary networks of five random fields
(control group was set at 100%) using Image-Pro Plus 6.0 software
(Media Cybernetics, Inc., Rockville, MD, USA). The results were the
means calculated from five replicates of each experiment.
Western blot analysis
Protein expression levels were analyzed by western
blot analysis, as previously described (24). Briefly, 25 μg of total protein/well
was loaded, following denaturing in loading buffer at 100°C for 5
min. The protein extracts were then separated by 8–12% SDS-PAGE and
electrophoretically transferred to nitrocellulose membranes (EMD
Millipore). Subsequently, the membranes were blocked at room
temperature for 2 h in 5% nonfat dry milk/Tris-buffered saline with
Tween® (TBST). The membranes were then incubated at 4°C
overnight with various primary antibodies. The primary antibodies
used in the present study were as follows: p-VEGFR2 (1:2,000),
p-Fak (1:2,000), p-Akt (1:1,000), p-Erk (1:2,000) and β-tubulin
(1:1,000). The following day, the membranes were washed three times
with TBST for 5 min at room temperature, and subsequently incubated
with HRP-conjugated anti-rabbit IgG secondary antibodies, for 2 h
at room temperature. Following the incubation, the membranes were
washed with TBST, and the proteins bands were visualized using the
Diaminobenzidine Detection system (Boster). β-tubulin was used as
the protein loading control.
Anti-angiogenic effects in vivo
In vivo Matrigel™ plug assay
A Matrigel™ plug assay was performed in BALB/c mice,
as described previously with some modifications (25). Briefly, female BALB/c mice (5–6
weeks old; weighing 17–20 g), were subcutaneously injected with 500
μl Matrigel™ containing heparin (130 U; Sigma-Aldrich) and mouse
VEGF (250 ng). Mice injected with Matrigel™ and heparin alone were
included as a vehicle control. The following day, the mice were
orally administrated a single dose of saline, or TSAJ at 1.8 or 3.6
mg/kg/d. Each group contained six mice and each dose was
administered daily by oral gavage. After 14 days, the Matrigel™
plugs were carefully removed, photographed, formalin-fixed and
paraffin-embedded.
To assay the microvascular density (MVD) of each
group, Masson’s Trichrome (M-T; Nanjing Jiancheng Bioengineering
Institute, Nanjing, China) staining was performed to visualize
endothelial infiltration. Briefly, 3 μm sections were stained with
M-T solution. The number of blood vessels in a high power field
were quantified by counting vessel infiltration into the plugs.
In vivo anti-tumor effects
H22 hepatoma cells [2×106 cells in 0.2 ml
normal saline (NS)] were subcutaneously injected into the armpits
of the right forearms of the mice. Once the tumor model had been
established, the mice were randomly divided into three groups each
containing 10 mice. The TSAJ-treated groups were orally
administrated 1.8 mg/kg/d or 3.6 mg/kg/d of TSAJ. The vehicle
control group was orally treated with the equivalent of 0.9% normal
saline. Each dose was administered daily by oral gavage and the
treatment lasted for 14 days. At the end of the experiment all the
mice were sacrificed with an overdose of 10% chloral hydrate by
intraperitoneal injection (400 mg/kg; Sigma-Aldrich), and the tumor
tissue was segregated, weighed and photographed. The tumor
inhibitory ratio was calculated using the following formula: Tumor
inhibitory rate (%)=(W control-W
treated)/W control × 100%. W
control and W treated represent the
average weights of the tumor in the vehicle control and
TSAJ-treated groups, respectively.
The tumor samples were then formalin-fixed and
paraffin-embedded. The sections (4 μm) were stained with
hematoxylin and eosin (KeyGen BioTech, Nanjng, China), or
immunostained with antibodies targeting mouse CD31 (1:100), p-ERK
(1:200), p-AKT (1:200), and p-VEGFR2 (1:200). The immunostaining
procedures were performed according to the manufacturer’s
instructions. MVD was calculated using Image-Pro Plus 6.0
software.
Statistical analysis
All experiments were repeated three times. Data
represent the mean ± standard error of the mean. Statistical
significance was analyzed by unpaired Student’s t-test or one-way
analysis of variance, using GraphPad v5.0 software (GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Preliminary phytochemical screening of
total saponins
In order to determine the existence of saponins
among the four fractions of the A. julibrissin extract,
traditional preliminary phytochemical screening assays (foam and
hemolytic tests) were conducted. The majority of saponins were
contained within 70% E (named TSAJ).
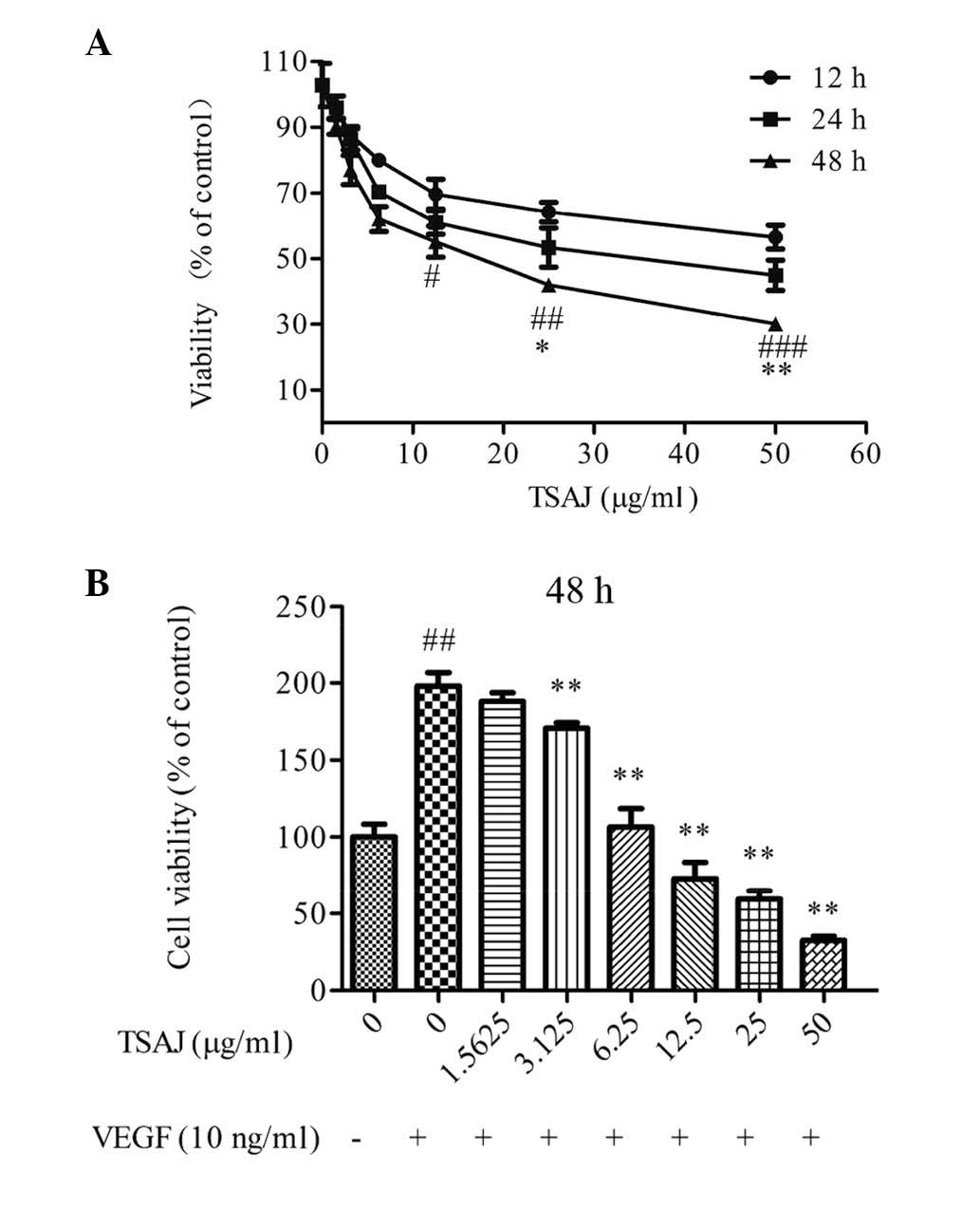
TSAJ inhibits VEGF-induced viability of
endothelial cells
The inhibitory effects of TSAJ on cell viability in
normal growth medium (containing 10% FBS) was initially evaluated
using a CCK-8 assay. TSAJ inhibited cell viability in a dose- and
time-dependent manner (Fig. 2A).
When the Ea.hy926 cells were treated with 18 μg/ml TSAJ for 48 h, a
significant inhibitory effect on viability was observed.
The present study also determined whether TSAJ
inhibited VEGF-induced endothelial cell viability, at the same dose
range. Stimulation with VEGF for 48 h increased the number of
Ea.hy926 cells by ~2 fold (Fig.
2B). Treatment with TSAJ significantly suppressed the
VEGF-induced increase in cell viability at doses from 9 μg/ml, in a
dose-dependent manner. These results demonstrate that
VEGF-activated endothelial cells were more sensitive to TSAJ, as
compared with those cultured in normal growth medium, and that TSAJ
may be a potent inhibitor of VEGF-induced increases in endothelial
cell viability.
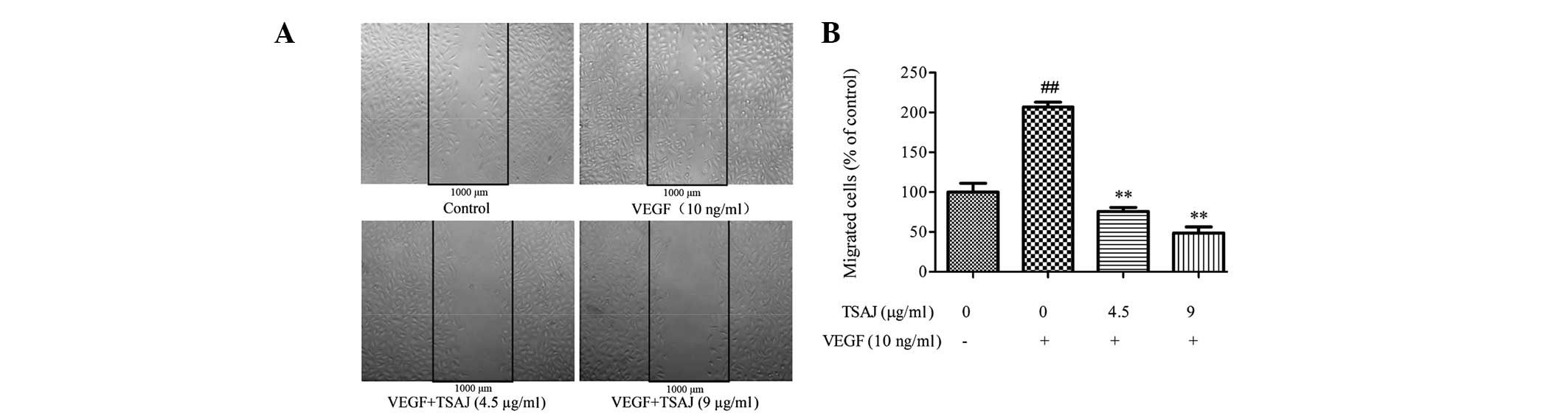
TSAJ inhibits VEGF-induced migration of
endothelial cells
During angiogenesis, endothelial cell migration is
an essential process (26).
Therefore, the effects of TSAJ on the migratory ability of Ea.hy926
cells was determined using a wound-healing assay. VEGF
significantly stimulated the directional migration of endothelial
cells (Fig. 3A), and the number of
migratory cells in the VEGF-treated group was >2 times more, as
compared with the control group (Fig.
3B). The migratory rate of the TSAJ-treated endothelial cells
was markedly suppressed, as compared with the VEGF-treated cells,
and the inhibitory effects were more obvious at 9 μg/ml, as
compared with at 4.5 μg/ml. These results suggest that TSAJ
significantly suppressed VEGF-induced migration of endothelial
cells in a dose-dependent manner.
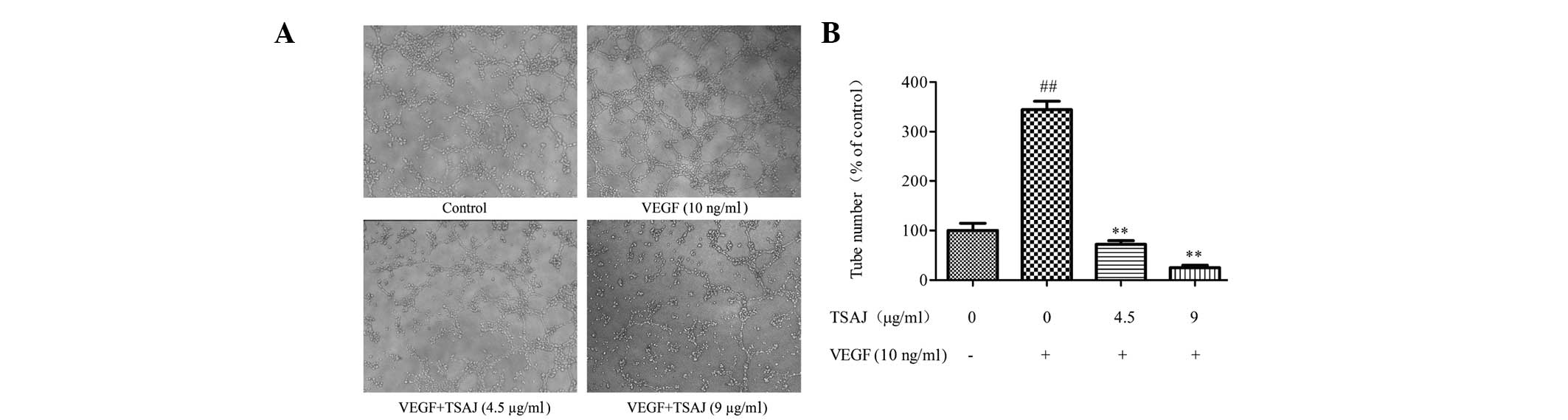
TSAJ inhibits VEGF-induced endothelial
cell tube formation
Angiogenesis is the formation of blood vessels from
pre-existing vasculature, and is considered to be a complex
process. During angiogenesis, the maturation of migrated
endothelial cells into a capillary tube is a critical early step
(27). Therefore, the present
study investigated how TSAJ regulates capillary tube formation.
Treatment with VEGF alone stimulated complete and robust tubular
capillary structures (Fig. 4A).
However, treatment with TSAJ massively disrupted the capillary tube
network, and the tubes formed in the TSAJ-treated group were rather
incomplete. In addition, the number of tubes formed was quantified
using Image-Pro Plus software in low power field, and tube
formation was shown to be negatively interrupted in response to
treatment with TSAJ (Fig. 4B).
These results demonstrate that TSAJ effectively inhibits
VEGF-induced Ea.hy926 cell tube formation on Matrigel™ in a
dose-dependent manner.
TSAJ inhibits activation of the
VEGFR2-mediated signaling pathway
It is well known that the VEGF signaling pathway has
a vital role in angiogenesis. VEGF binding to VEGFR2 results in the
autophosphorylation of VEGFR2, which subsequently activates various
downstream signaling molecules that are responsible for endothelial
cell migration, proliferation and survival (28). To further elaborate the underlying
mechanisms of the anti-angiogenic effects of TSAJ, the present
study evaluated some key signaling molecules involved in the
VEGFR2-mediated signaling pathway. VEGF induces survival of
endothelial cells mainly through activation of Akt, whereas
activation of Erk1/2 mitogen-activated protein kinases (MAPKs) is
thought to be essential for VEGF-induced proliferation.
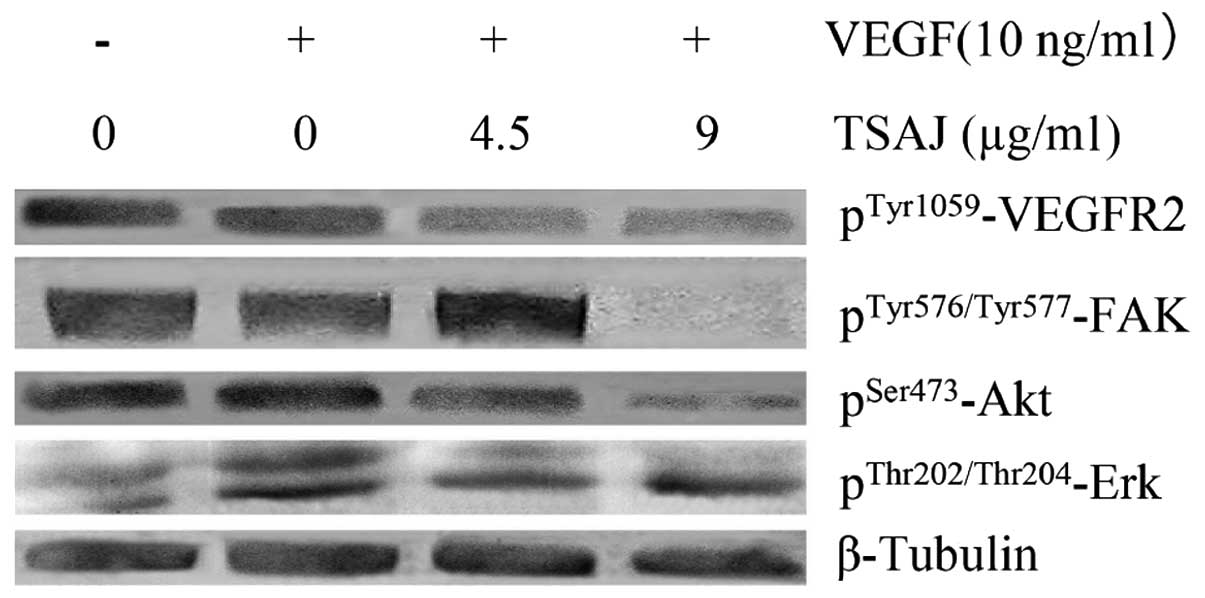
VEGF-induced phosphorylation of VEGFR2 at Tyr-1175 was suppressed
by treatment with TSAJ in a dose-dependent manner (Fig. 5). In addition, TSAJ significantly
suppressed the VEGF-induced phosphorylations of Akt
(Ser473) and Erk (Thr202/Thr204).
The effects of TSAJ on the phosphorylation of FAK were also
examined, and TSAJ was shown to inhibit VEGF-induced
phosphorylation of FAK, when administered at a dose of 9 μg/ml.
These results indicate that TSAJ exerts its anti-angiogenic effects
by selectively targeting certain signaling events downstream of
VEGFR2.
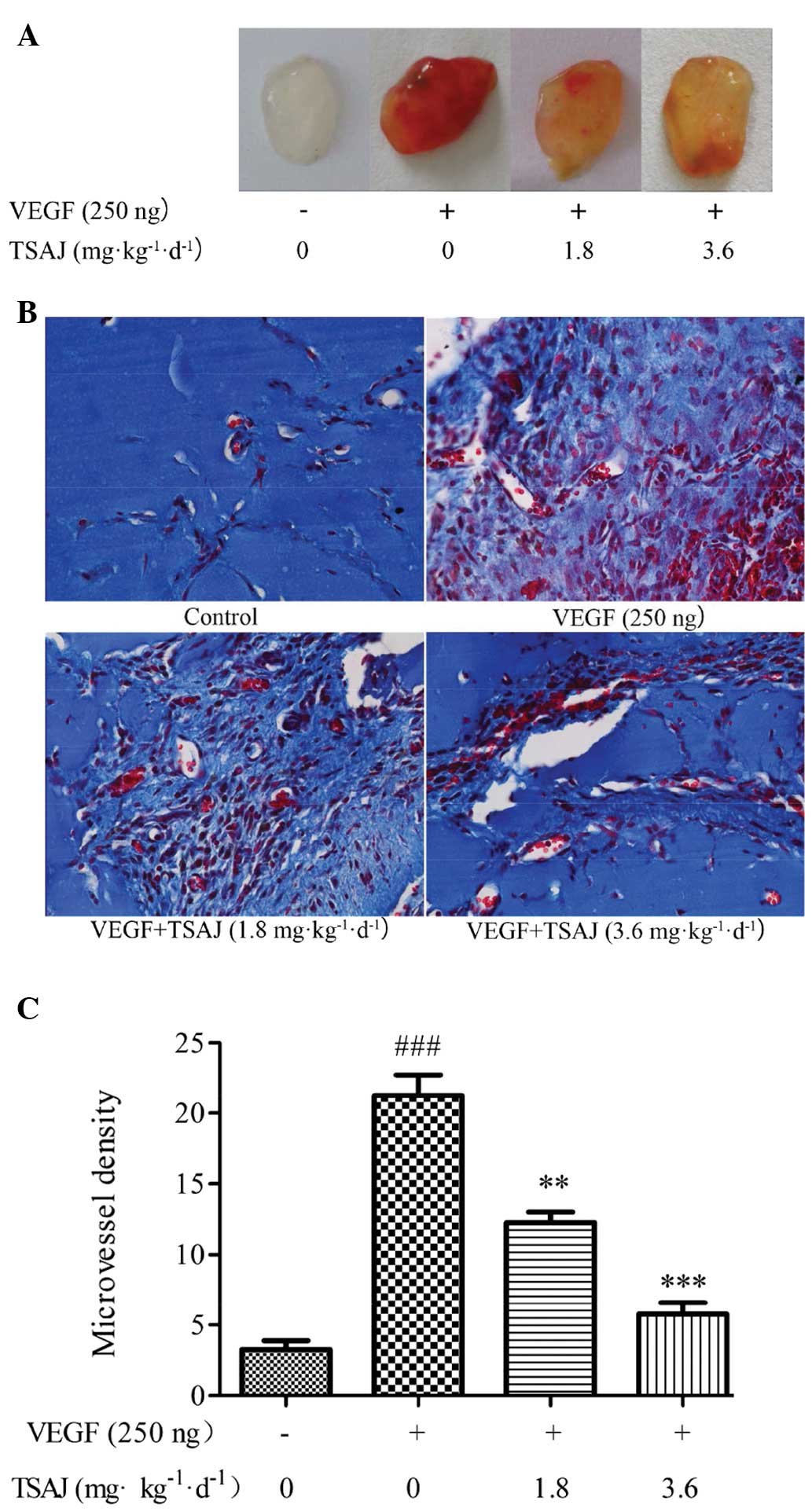
TSAJ inhibits VEGF-induced blood vessel
formation in mice
The results of the present study demonstrated that
TSAJ effectively inhibited VEGF-induced angiogenesis in
vitro, therefore the anti-angiogenic effects of TSAJ were
subsequently validated in vivo. The anti-angiogenic effects
of TSAJ were evaluated in a mouse Matrigel™ plug model, which is a
powerful in vivo angiogenesis assay (29). Matrigel™ plugs containing VEGF
alone were dark red, whereas the Matrigel™ plugs of the
TSAJ-treated group were light red (Fig. 6A). Conversely, the Matrigel™ plugs
of the vehicle group were avascular and pale white. These
superficial phenomena suggest that abundant functional vasculatures
had formed within the VEGF group, whereas fewer blood vessels had
formed within the Matrigel™ plugs of the TSAJ-treated group. To
provide further evidence for this finding, the number of functional
vessels within the Matrigel™ plugs were compared by M-T staining,
which stained the Matrigel™ blue and the endothelial cells/vessels
red (Fig. 6B). Fewer vessels were
observed in the Matrigel™ plugs of the mice treated with VEGF and
TSAJ, as compared with those in the mice treated with VEGF alone
(Fig. 6C). These results
demonstrate that TSAJ strongly suppresses VEGF-induced angiogenesis
14 days after Matrigel™ implantation. These findings were
concordant with the results of the in vitro tube formation
assay.
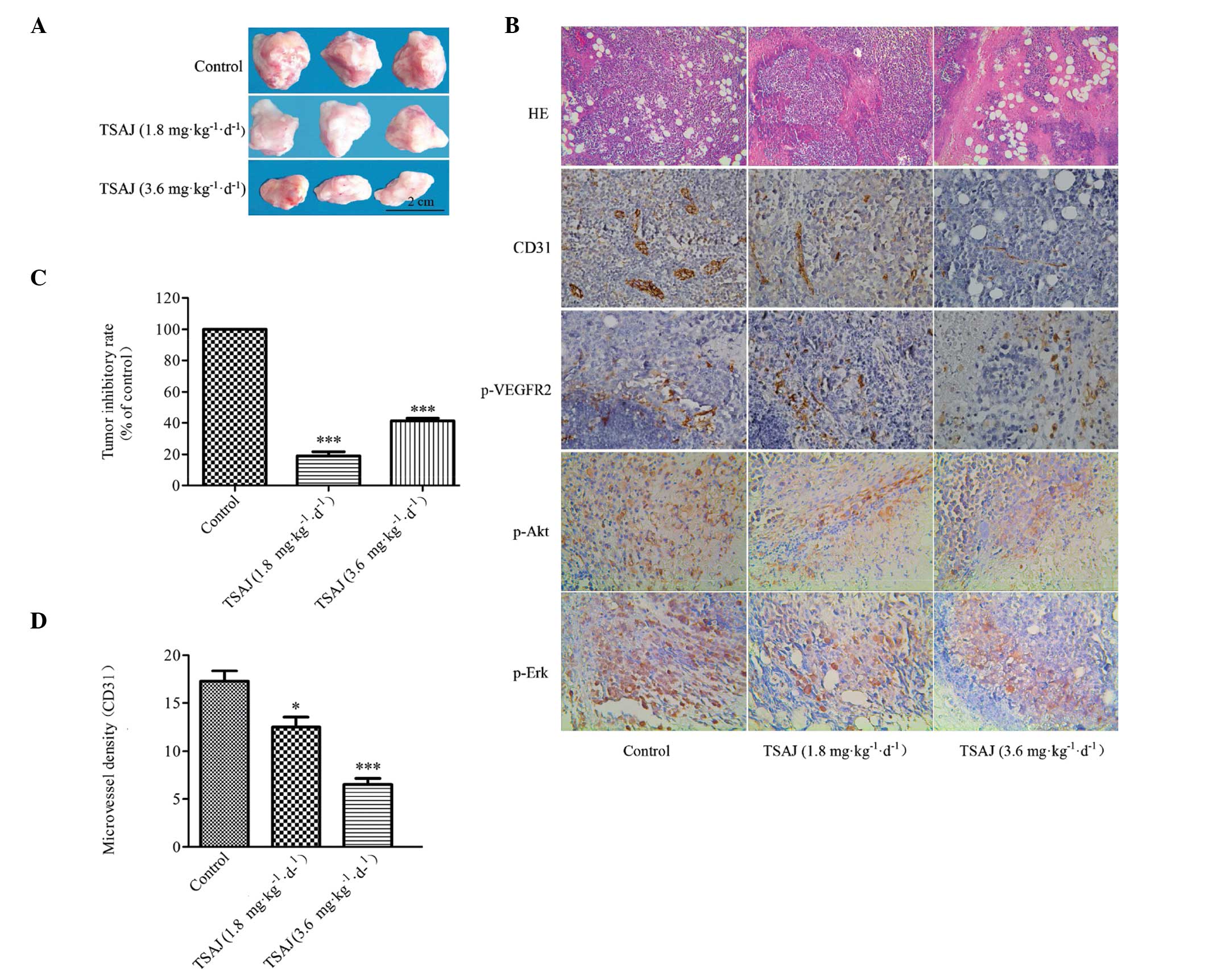
Anti-tumor effects of TSAJ on a H22
hepatoma cell transplantation model
Angiogenesis is the pivotal step in tumor growth and
metastasis, which provides necessary oxygen and nutrients for the
tumor (30). To investigate the
anti-tumor effects of TSAJ, a H22 hepatoma tumor model was
generated.
Tumor growth was significantly suppressed by
treatment with TSAJ, at the dose of 3.6 mg/kg/d, after 14 days of
treatment (Fig. 7A). The tumors of
the TSAJ-treated group were significantly smaller, as compared with
those in the control group. The average tumor weight in the control
group was 2.023±0.119 g, whereas the average tumor weight in the
3.6 and 1.8 mg/kg/d TSAJ-treated groups were 1.189±0.106 g and
1.637±0.167 g, respectively. The pathological results showed that
extensive densely vesicular nuclei and sparse cytoplasm was
observed in the TSAJ-treated groups. Furthermore, as the
concentration of TSAJ increased, large areas of necrosis appeared
in the tumor tissue, and the region of necrosis expanded (Fig. 7B). The inhibitory rate of tumors in
the 3.6 mg/kg/d treatment group was 41.245% (Fig. 7C). These results indicate that the
rate of proliferation of tumor cells was inhibited by TSAJ, in a
dose-dependent manner.
Although tumor growth was indeed suppressed in
response to treatment with TSAJ, the anti-tumor mechanisms of TSAJ
still required further validation. To investigate new blood vessel
formation in solid tumors, immunostaining for CD31 was performed
(Fig. 7B). Microvessels were
stained brown by CD31, which is a specific-endothelial marker. The
MVD values in the TSAJ-treated groups were significantly reduced,
as compared with the control group (Fig. 7D). Furthermore, treatment with TSAJ
significantly decreased the expression levels of p-VEGFR-2, p-Erk
and p-Akt (brown-stained field), as compared with the control group
(Fig. 7B). These results suggest
that TSAJ is a potent angiogenic inhibitor, which inhibits tumor
neovascularization and suppresses tumor growth through inhibition
of the VEGFR-2 signaling pathway.
Discussion
In recent years, numerous efforts have been made to
identify potential anti-angiogenic agents from traditional Chinese
medicinal herbs. AJ has been used to treat various diseases for
thousands of years (31); however,
little is currently known with regard to the inhibitory effects of
AJ on angiogenesis. Previous studies have reported that the
chemical composition of AJ includes saponins, flavonoids, alkaloids
and polysaccharides (32), and
saponins are considered to be the main anti-tumor component
(33). The present study explored
the anti-angiogenic mechanisms of TSAJ. Angiogenesis depends on the
proliferation, migration and tube formation of endothelial cells
(34), and VEGF is a major
proangiogenenic factor that mediates these processes (35). The results of the present study
demonstrated that VEGF significantly promoted the proliferation of
Ea.hy926 cells. However, treatment with TSAJ resulted in a marked
decrease in the VEGF-induced proliferation of Ea.hy926 cells, in a
dose-dependent manner. Migration of endothelial cells as a response
to pro-angiogenic factors is an integral feature of angiogenesis.
The inhibitory effects of TSAJ in vitro were evaluated using
a wound healing assay and a growth factor-reduced two-dimensional
Matrigel™ model. In the two models, TSAJ inhibited the VEGF-induced
cell migration and capillary tube formation of Ea.hy926 cells, thus
suggesting that treatment with TSAJ suppressed VEGF-induced
angiogenesis by restraining the cellular connection.
Angiogenesis stimulated by VEGF is a complex
process. Upon binding to VEGF, VEGFR2 undergoes autophosphorylation
and transmits the signal to downstream molecules associated with
angiogenic processes. The PI3K/Akt and RAF/MEK-ERK pathways are
involved in VEGF-induced angiogenesis. The phosphorylation of
VEGFR2 at Tyr-1175 is required for activation of Akt, which
contributes to the proliferation and survival of endothelial cells
(36). In addition,
phosphorylation of VEGFR2 at Tyr-1175 activates the MAPK/ERK
cascades, which also promote the proliferation of endothelial cells
(36). FAK and its substrate
paxillin are signaling molecules associated with VEGF-induced
migration, which participate in focal adhesion during cell
migration (37–39).
To elucidate the underlying anti-angiogenic
mechanisms of TSAJ, the expression of signaling molecules and
pathways were analyzed by western blotting. Treatment with TSAJ
suppressed the phosphorylation of VEGFR2 at the Tyr1175 site in
endothelial cells, suggesting that TSAJ may inhibit the kinase
activity of VEGFR2 by downregulating the expression of VEGFR2.
Furthermore, TSAJ significantly suppressed the VEGF-induced
phosphorylation of Akt (Ser473) and Erk (Thr202/Thr204), in a
dose-dependent manner. However, the phosphorylation of FAK was only
markedly inhibited when the cells were treated with TSAJ at a dose
of 9 μg/ml. These results suggest that TSAJ modulated VEGF-induced
vascular permeability and angiogenesis by inhibiting the
phosphorylation of VEGFR2, FAK, AKT, and ERK in endothelial
cells.
The results of the present study demonstrated that
TSAJ exhibits highly anti-angiogenic effects in vitro. In
order to confirm the anti-angiogenic efficacy of TSAJ in an animal
model, a mouse Matrigel™ plug assay was used. The Matrigel™ plug
assay is a relatively quick and easy method to evaluate angiogenic
and anti-angiogenic compounds (25). TSAJ markedly suppressed
VEGF-induced neovascularization in the Matrigel™ plug assay, by
inhibiting endothelial cell infiltration into the plug. The number
of functional capillaries containing erythrocytes were markedly
decreased in the TSAJ-treated group, as compared with the
VEGF-induced group. These results suggest that TSAJ inhibited
VEGF-induced neovascularization in the Matrigel™ plug assay, by
inhibiting the invasion of endothelial cells.
Since the anti-angiogenic effects of TSAJ in
vivo were validated using the Matrigel™ plug model, the H22
hepatoma tumor model was used to provide further evidence regarding
the inhibitory effects. The hepatoma tumor model is widely used in
the field of anti-tumor research (40). In the primary stages of tumor
growth, tumor cells rapidly proliferate, and an abundance of
nutrients and oxygen is required (41). Therefore, oxygen and nutrients,
which are provided by tumor blood vessels, are considered to be
critical factors to limit the growth of tumors. Once the formation
of blood vessels is suppressed, tumor cells will undergo necrosis,
due to ischemia and hypoxia (42,43).
In response to hypoxia, vasculogenesis is induced by VEGF-A165,
which is secreted by tumor cells and can improve the tumor
microenvironment (7). Based on
this concept, inhibition of angiogenesis may be an effective
approach to prevent and treat cancer. In the present study, tumor
growth was significantly inhibited in the TSAJ-treated group.
Furthermore, the percentage of necrosis in the TSAJ-treated group
was higher, as compared with that of the control group.
Immunohistochemical staining with antibodies targeting CD31
demonstrated that the MVD was significantly decreased in the
TSAJ-treated group, which limited the blood supply to the tumor. In
addition, immunohistochemical staining showed that the expression
of p-VEGFR2, p-Akt and p-Erk were significantly suppressed in the
TSAJ-treated group. These signaling molecules are the main factors
associated with the VEGF/VEGFR2 signaling pathway in angiogenesis
of vascular endothelial cells. Therefore, downregulation of
p-VEGFR2, p-Akt and p-Erk expression may be the underlying
mechanism for the anti-angiogenic effects of TSAJ.
In conclusion, the present study was the first, to
the best of our knowledge, to demonstrate that TSAJ is able to
inhibit VEGF-induced endothelial cell proliferation, migration, and
tube formation, by suppressing the activation of VEGFR2 and its
downstream signaling of Fak, Akt and Erk in vitro and in
vivo. Furthermore, the anti-angiogenic effects of TSAJ in
vivo demonstrated that TSAJ may be considered as a potential
oral anti-angiogenic drug, which targets the VEGF/VEGFR2 signal
pathway.
Acknowledgements
The present study was supported by Fundamental
Research Funds for the Central Universities (grant no.
JUSRP51412B).
Abbreviations:
|
AJ
|
Albizia julibrissin
|
|
TSAJ
|
total saponins from Albizia
julibrissin
|
|
FBS
|
fetal bovine serum
|
|
HE
|
hematoxylin and eosin
|
|
M-T
|
Masson’s Trichrome
|
|
IHC
|
immunohistochemistry
|
|
TBST
|
Tris-buffered saline with Tween
|
|
VEGF
|
vascular endothelial growth factor
|
|
p-VEGFR2
|
phosphorylated-vascular endothelial
growth factor receptor 2
|
|
p-Erk
|
phosphorylated-extracellular
signal-regulated kinase
|
|
p-Akt
|
phosphorylated-Akt
|
|
p-Fak
|
phosphorylated focal adhesion
kinase
|
References
|
1
|
Folkman J: Tumor angiogenesis: therapeutic
implications. N Engl J Med. 285:11821971. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wei A, Zhou D, Ruan J, Cai Y, Xiong C and
Wu G: Anti-tumor and anti-angiogenic effects of Macrothelypteris
viridifrons and its constituents by HPLC-DAD/MS analysis. J
Ethnopharmacol. 139:373–380. 2012. View Article : Google Scholar
|
|
3
|
Rodriguez-Brenes IA, Komarova NL and
Wodarz D: Tumor growth dynamics: insights into evolutionary
processes. Trends Ecol Evol. 28:597–604. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gacche RN and Meshram RJ: Targeting tumor
micro-environment for design and development of novel
anti-angiogenic agents arresting tumor growth. Prog Biophys Mol
Biol. 1132:333–354. 2013. View Article : Google Scholar
|
|
5
|
Makrilia N, Lappa T, Xyla V, Nikolaidis I
and Syrigos K: The role of angiogenesis in solid tumours: an
overview. Eur J Intern Med. 20:663–671. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ellis LM and Hicklin DJ: VEGF-targeted
therapy: mechanisms of anti-tumour activity. Nat Rev Cancer.
8:579–591. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Saharinen P, Eklund L, Pulkki K, Bono P
and Alitalo K: VEGF and angiopoietin signaling in tumor
angiogenesis and metastasis. Trends Mol Med. 17:347–362. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kang Z, Jiang W, Luan H, Zhao F and Zhang
S: Cornin induces angiogenesis through PI3K-Akt-eNOS-VEGF signaling
pathway. Food Chem Toxicol. 58:340–346. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoon H, Choi YL, Song JY, et al: Targeted
inhibition of FAK, PYK2 and BCL-XL synergistically enhances
apoptosis in ovarian clear cell carcinoma cell lines. PLoS One.
9:e885872014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin SH and Shih YW: Antitumor effects of
the flavone chalcone: inhibition of invasion and migration through
the FAK/JNK signaling pathway in human gastric adenocarcinoma AGS
cells. Mol Cell Biochem. 391:47–58. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu J, Wang YS, Zhang J, et al: Focal
adhesion kinase signaling pathway participates in the formation of
choroidal neovascularization and regulates the proliferation and
migration of choroidal microvascular endothelial cells by acting
through HIF-1 and VEGF expression in RPE cells. Exp Eye Res.
88:910–918. 2009. View Article : Google Scholar
|
|
12
|
China Pharmacopoeia Committee, . Chinese
Pharmacopoeia. 1. Chemical Industry Press; Beijing: pp. p972005
|
|
13
|
Zou K, Zhao YY and Zhang RY: A cytotoxic
saponin from Albizia julibrissin. Chem Pharm Bull (Tokyo).
54:1211–1212. 2006. View Article : Google Scholar
|
|
14
|
Won HJ, Han CH, Kim YH, et al: Induction
of apoptosis in human acute leukemia Jurkat T cells by Albizzia
julibrissin extract is mediated via mitochondria-dependent
caspase-3 activation. J Ethnopharmacol. 106:383–389. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu LY, Du FF, Feng L and Qiu LY: Effect
of different components from Albizia julibrissin on HMEC cells and
3B11 cells. Lishizhen Med Mater Med Res. 22:762–764. 2011.
|
|
16
|
Liang H, Tong WY, Zhao YY, Cui JR and Tu
GZ: An antitumor compound julibroside J28 from Albizia julibrissin.
Bioorg Med Chem Lett. 15:4493–4495. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zou K, Zhao YY and Zhang RY: Triterpenoid
saponins from Cortex Albiziae. Journal of Practical Training of
Medicine. 36:77–87. 2008.
|
|
18
|
Hua H, Feng L, Zhang XP, Zhang LF and Jin
J: Anti-angiogenic activity of julibroside J8, a natural product
isolated from Albizia julibrissin. Phytomedicine. 16:703–711. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zheng L, Zheng J, Zhang Q, Wang B, Zhao Y
and Wu L: Three new oleanane triterpenoid saponins acetylated with
monoterpenoid acid from Albizia julibrissin. Fitoterapia.
81:859–863. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jung YH, Ha RR, Kwon SH, et al: Anxiolytic
effects of Julibroside C1 isolated from Albizia julibrissin in
mice. Prog Neuropsychopharmacol Biol Psychiatry. 44:184–192. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou MH, Li JG, Wang RL, Liang CQ, Lu YL
and Yang G: Experimental study on hemocytolysis of Rhizoma paridis
total saponins. Chinese Journal of Cerebrovascular Diseases.
18:1611–1612. 2007.
|
|
22
|
Angulo J, Peir C, Romacho T, et al:
Inhibition of vascular endothelial growth factor (VEGF)-induced
endothelial proliferation, arterial relaxation, vascular
permeability and angiogenesis by dobesilate. Eur J Pharmacol.
667:153–159. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pinkaew D, Cho SG, Hui DY, et al:
Morelloflavone blocks injury-induced neointimal formation by
inhibiting vascular smooth muscle cell migration. Biochim Biophys
Acta. 1790:31–39. 2009. View Article : Google Scholar
|
|
24
|
Kim GD, Cheong OJ, Bae SY, Shin J and Lee
SK: 6′-Debromohamacanthin A, a bis (indole) alkaloid, inhibits
angiogenesis by targeting the VEGFR2-mediated PI3K/AKT/mTOR
signaling pathways. Mar Drugs. 11:1087–1103. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Malinda KM: In vivo matrigel migration and
angiogenesis assay. Methods Mol Biol. 467:287–294. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sadanandam A, Rosenbaugh EG, Singh S,
Varney M and Singh RK: Semaphorin 5A promotes angiogenesis by
increasing endothelial cell proliferation, migration and decreasing
apoptosis. Microvasc Res. 79:1–9. 2010. View Article : Google Scholar
|
|
27
|
Lai SL, Cheah SC, Wong PF, Noor SM and
Mustafa MR: In vitro and in vivo anti-angiogenic activities of
Panduration A. PLoS One. 7:e381032012. View Article : Google Scholar
|
|
28
|
Pober JS and Sessa WC: Evolving functions
of endothelial cells in inflammation. Nat Rev Immunol. 7:803–815.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Adini A, Fainaru O, Udagawa T, Connor KM,
Folkman J and D’Amato RJ: Matrigel cytometry: a novel method for
quantifying angiogenesis in vivo. J Immunol Methods. 342:78–81.
2009. View Article : Google Scholar
|
|
30
|
Gomes FG, Nedel F, Alves AM, Nr JE and
Tarquinio SB: Tumor angiogenesis and lymphangiogenesis:
tumor/endothelial crosstalk and cellular/microenvironmental
signaling mechanisms. Life Sci. 92:101–107. 2013. View Article : Google Scholar :
|
|
31
|
Tian WY, Ma CL, Liu F, Xia LH, Wang L and
Wang JX: Research on the antineoplastic activity of the extract of
alcoholized Albizia cortex in vivo of tumor-bearing mice. College.
22:5–6. 2000.
|
|
32
|
Yu DH, Qiao SY and Zhao YM: Advance in
study on bark of Albizzia julibrissin. 29:619–624. 2004.
|
|
33
|
Li Q, Feng L, Shi JJ, Liu LY, Du B and Qiu
LY: Screening of active substances of anti-angiogenesis induced by
tumour cell in Albizia. Chin Tradit Pat Med. 34:744–747. 2012.
|
|
34
|
Goodwin AM: In vitro assays of
angiogenesis for assessment of angiogenic and anti-angiogenic
agents. Microvasc Res. 74:172–183. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao T, Zhao W, Chen Y, Ahokas RA and Sun
Y: Vascular endothelial growth factor (VEGF)-A: role on cardiac
angiogenesis following myocardial infarction. Microvasc Res.
80:188–194. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Holmqvist K, Cross MJ, Rolny C, et al: The
adaptor protein shb binds to tyrosine 1175 in vascular endothelial
growth factor (VEGF) receptor-2 and regulates VEGF-dependent
cellular migration. J Biol Chem. 279:22267–22275. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao X and Guan JL: Focal adhesion kinase
and its signaling pathways in cell migration and angiogenesis. Adv
Drug Deliv Rev. 63:610–615. 2011. View Article : Google Scholar :
|
|
38
|
Chen XL, Nam JO, Jean C, et al:
VEGF-induced vascular permeability is mediated by FAK. Dev Cell.
22:146–157. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee J, Ku T, Yu H, et al: Blockade of
VEGF-A suppresses tumor growth via inhibition of autocrine
signaling through FAK and AKT. Cancer Lett. 318:221–225. 2012.
View Article : Google Scholar
|
|
40
|
Wu, Gao, Chen, et al: Anti-tumor effects
of a novel chimeric peptide on S180 and H22 xenografts bearing nude
mice. Peptides. 31:850–864. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li Q, Fu GB, Zheng JT, et al: NADPH
oxidase subunit p22 (phox)-mediated reactive oxygen species
contribute to angiogenesis and tumor growth through AKT and ERK1/2
signaling pathways in prostate cancer. Biochim Biophys Acta.
1833.3375–3385. 2013.
|
|
42
|
Burkovetskaya ME, Levin SG and Godukhin
OV: Neuroprotective effects of interleukin-10 and tumor necrosis
factor-alpha against hypoxia-induced hyperexcitability in
hippocampal slice neurons. Neurosci Lett. 416:236–240. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Carloni S, Carnevali A, Cimino M and
Balduini W: Extended role of necrotic cell death after
hypoxia-ischemia-induced neurodegeneration in the neonatal rat.
Neurobiol Dis. 27:354–361. 2007. View Article : Google Scholar : PubMed/NCBI
|