Introduction
The hedgehog (Hh) genes, including Sonic hedgehog
(Shh), Indian hedgehog (Ihh) and Desert hedgehog are evolutionarily
conserved and serve important roles in controlling the
differentiation of progenitor cells into either osteoblasts or
chondrocytes (1). Dysfunction of
Hh signaling leads to disruption of bone development and
homeostasis (2).
Osteoblasts originate from pluripotent mesenchymal
stem cells. The most important function of osteoblasts is to form
mineralized bones. Osteoblasts express various phenotypic markers,
including high alkaline phosphatase (ALP) activity, and synthesize
collagenous and noncollagenous bone matrix proteins, including
osteocalcin (OCN) (3). Osteoblasts
express receptors for various hormones, including parathyroid
hormone-related protein (PTHrP), runt-related transcription factor
2 (RUNX2) and tumor necrosis factor ligand superfamily member 11
(RANKL), which are involved in the regulation of osteoblast
differentiation (4,5).
It has been suggested that Ihh may induce adjacent
perichondrial cells to differentiate into bone-forming osteoblasts
(6). Ihh null mutant mice
exhibited a failure of osteoblast development in endochondral bones
(7). Ihh overexpression increased
ALP activity and the level of OCN mRNA expression in MC3T3-E1 cells
(8). In addition, recombinant Shh
synergistically stimulated bone morphogenetic protein-induced ALP
activity and the expression level of OCN mRNA in C3H10T1/2 cells
(9). Although various studies have
demonstrated that Hh signaling is a potent local factor that
regulates osteoblast differentiation, the specific transcription
factors that determine osteoblast differentiation remain unclear.
Further studies are required to determine the precise mechanism
through which Ihh regulates osteoblast differentiation.
In the present study, Ihh was knocked down in
osteoblast MC3T3-E1 cells using short hairpin (sh)RNA, in order to
investigate the function of Ihh in osteoblast proliferation and
differentiation and to examine the potential mechanism through
which Ihh may induce osteoblast apoptosis and cell cycle
arrest.
Materials and methods
Cell line and cell transfection
The mouse osteoblast MC3T3-E1 cells were purchased
from the American Type Culture Collection (Manassas, VA, USA).
Cells were cultured routinely in Dulbecco's modified Eagle's
medium/F-12 (Invitrogen; Thermo Fisher Scientific, Inc., Waltham,
MA, USA), supplemented with 10% fetal bovine serum (Gibco; Thermo
Fisher Scientific, Inc.), and cultured in a 37°C humidified
atmosphere with 5% CO2.
Knockdown of Ihh in cells was achieved by infection
with a lentivirus containing Ihh shRNA sequences (Ihh-shRNA;
Shanghai GenePharma Co., Ltd., Shanghai, China). Cells transfected
with empty lentivirus were used as the negative control (NC), and
untreated cells were used as the blank control. Cells
(5×105 cells/ml) were plated in 6-well clusters (1 ml)
or 96-well plates (0.2 ml) and transfected with lentivirus
(108 U/ml) for 48 h by using Lipofectamine 3000 (Thermo
Fisher Scientific, Inc.). Transfected cells were used in further
assays or RNA/protein extraction.
RNA extraction and SYBR green reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). RevertAid
First Strand cDNA Synthesis kit (Thermo Fisher Scientific, Inc.)
was used to reverse transcribe the mRNA to cDNA according to the
manufacturer's protocol. The mRNA expression of Ihh, PTHrP,
transforming growth factor (TGF)-β, OCN, RUNX2, tumor necrosis
factor receptor superfamily member 11B (OPG), mothers against
decapentaplegic homolog (Smad)2, Smad3, collagen α-1 (X) chain
(COL10A) and RANKL mRNA expression was measured using a SYBR Green
qPCR assay (Takara Biotechnology Co., Ltd., Dalian, China) under
the following conditions: 95°C for 5 min, followed by 40 cycles of
95°C for 15 sec and 60°C for 30 sec. The primers used in this study
were as follows: Ihh, forward: 5′-CTCAGCCTGCTCTCACTACG-3′, reverse:
5′-AAGCACATCCAACCCACCTC-3′; PTHrP, forward:
5′-CGAGGTTCAAAGGTTTGCCTC-3′, reverse: 5′-GGCCAGAGAAGCCTGTTACC-3′;
TGF-β, forward: 5′-AGGGCTACCATGCCAACTTC-3′, reverse:
5′-TGACACAGAGATCCGCAGTC-3′; OCN, forward:
5′-TCCTTTGGGGTTTGGCCTAC-3′, reverse: 5′-CTTGGACACAAAGGCTGCAC-3′;
RUNX2, forward: 5′-CGCCTCACAAACAACCACAG-3′, reverse:
5′-TCACTGTGCTGAAGAGGCTG-3′; OPG, forward:
5′-CTGGAACCCCAGAGCGAAAT-3′, reverse: 5′-GCGTTTACTTTGGTGCCAGG-3′;
Smad2, forward: 5′-GTTCCTTTCCTCCTCCGCTC-3′, reverse:
5′-AGTCTCTTCACAACTGGCGG-3′; Smad3, forward:
5′-TTCACTGGTGCTGGGGTTAG-3′, reverse: 5′-GGTAGGGATTCACGCAGACC-3′;
COL10A, forward: 5′-CTCCCAGCACGCAGAATC-3′, reverse:
5′-TTCCCTACAGCTGATGGTCC-3′; RANKL, forward:
5′-GGAGTTGGCCGCAGACAAGA-3′, reverse: 5′-TGATGTGCTGTGATCCAACGA-3′
and β-actin, forward: 5′-GCAGGAGTATGACGAGTCCG-3′, reverse:
5′-AACAACGCATCTCATATTTGGAA-3′. The expression of β-actin was used
as an endogenous control. Data were processed using the
2−ΔΔCq method (10).
Cell Counting Kit-8 (CCK-8) cell
proliferation assay
Cell proliferation rates were measured using a CCK-8
assay (Beyotime Institute of Biotechnology, Haimen, China). A total
of 0.5×104 cells/well were seeded into a 96-well plate
for 24 h, transfected with the indicated lentiviruses, and further
incubated for 24, 48 and 72 h, respectively. A total of 10 µl CCK-8
reagents were added to each well at 1 h prior to the endpoint of
incubation. The optical density at 490 nm for each well was
determined using a microplate reader.
Flow cytometric analysis of apoptosis
with Annexin-V/PI double staining
An annexin V apoptosis detection kit (Thermo Fisher
Scientific, Inc.) was used for the analysis of apoptosis. Following
transfection, the cells were trypsinized, harvested and
resuspended. A total of 2×105 cells were harvested and
washed twice with cold PBS, and subsequently resuspended in 500 µl
binding buffer. A total of 10 µl annexin V-fluorescein
isothiocyanate and 10 µl propidium iodide was added to the solution
and mixed well. Following 15 min of incubation, the cells were
analyzed by flow cytometric analysis (BD Biosciences, San Jose, CA,
USA) with FCS Express 6 flow cytometry software (FCS Express
6.04.0015, De Novo Software, Los Angeles, CA, USA).
Cell cycle analysis
Cell cycle analysis was performed using flow
cytometry. A total of 1×104 cells were seeded into each
well of a 6-well plate. Following 48 h of transfection with
Ihh-shRNA or empty lentivirus, the cells were harvested and fixed
in 70% ice-cold ethanol for 24 h, followed by staining with
propidium iodide at 37°C for 15 min. The different phases of the
cell cycle were analyzed using a FACSCalibur instrument (BD
Biosciences) FCS Express 6 software.
ALP assay
An ALP Assay kit (Beyotime Institute of
Biotechnology) was used to detect ALP activity, according to the
manufacturer's protocol. The positive signals were stained with
black CoS. Cells were counterstained with nuclear fast red at room
temperature for 5 min to label the nucleus and cytoplasm of cells.
Images were obtained using an inverted microscope (Nikon Eclipse TC
100; Nikon Corporation, Tokyo, Japan) at ×100 magnification.
Determination of mineralization using
von Kossa staining
Von Kossa staining (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was used to determine the extent of minerals
deposited on osteoblast cells. Silver nitrate solution (0.5%) was
added onto the cells and the plate was exposed to ultraviolet light
for 30 min. Subsequently, the cells were washed with saline and the
reaction was stopped with the addition of 500 µl 5% sodium
thiosulfate (Sigma-Aldrich; Merck KGaA). Positive mineral
depositions were stained in black. Cells were counterstained with
nuclear fast red at room temperature for 5 min to label the nucleus
and cytoplasm of cells. Images were obtained using an inverted
microscope (Nikon Eclipse TC 100) at ×100 magnification.
Western blot analysis
Immunoblotting was performed to detect the
expression of Ihh, PTHrP, TGF-β, OCN, RUNX2, OPG, Smad2, Smad3,
COL10A and RANKL in chondrocyte cells. Cultured or transfected
cells were lysed in radioimmunoprecipitation assay buffer (Boster
Biological Technology, Pleasanton, CA, USA) with 1%
phenylmethylsulfonyl fluoride. The concentration of protein was
determined using a BCA Protein Assay kit (cat. no. P0011, Beyotime
Institute of Biotechnology) according to the manufacturer's
protocol. Protein (60 µg) was loaded onto a 12% SDS-PAGE minigel
and transferred onto a polyvinylidene fluoride membrane. The
membranes were blocked in 10% non-fat milk for 2 h at 37°C.
Following probing with primary antibodies (anti-Ihh, cat. no.
SAB1403965, dilution, 1:1,000; anti-PTHrP, cat. no. SAB5300029,
dilution, 1:1,000; anti-TGF-β, cat. no. SAB4502958, dilution,
1:2,000 and anti-OCN, cat. no. SAB1306277, dilution, 1:2,000 from
Sigma Aldrich; Merck KGaA; anti-RUNX2, cat. no. 12556, dilution,
1:1,000; anti-OPG, cat. no. 4816, dilution, 1:1,000; anti-Smad2,
cat. no. 5339, dilution, 1:1,000 and anti-Smad3, cat. no. 9523,
dilution, 1:1,000 from Cell Signaling Technology, Inc., Danvers,
MA, USA; anti-COL10A, cat. no. ab58632, dilution, 1:1,000;
anti-RANKL, cat. no. ab9957, dilution, 1:1,000 and anti-GAPDH, cat.
no. ab9485, dilution, 1:3,000 from Abcam, Cambridge, UK) at 4°C
overnight, the blots were subsequently incubated with horseradish
peroxidase-conjugated secondary antibody (cat. nos. ab7090 and
ab97040; 1:5,000, Abcam) for 1 h at 37°C. Signals were visualized
using enhanced chemiluminescence substrates (EMD Millipore,
Billerica, MA, USA). GAPDH was used as an endogenous protein for
normalization. The densitometry of bands was analyzed using
Quantity One software (version 4.6.9; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Statistical analysis
All data from three independent experiments are
expressed as the mean ± standard deviation and were processed using
SPSS 17.0 statistical software (SPSS, Inc., Chicago, IL, USA). The
difference among the groups was determined using Student's t-test
or one-way analysis of variance with Tukey's post hoc test,
depending on the conditions. P<0.05 was considered to indicate a
statistically significant difference.
Results
Knockdown of Ihh inhibits osteoblast
cell proliferation, induces apoptosis and arrests the cell cycle at
the G1 to S phase
In order to investigate the biological role of Ihh
in osteoblast cell growth, osteoblast MC3T3-E1 cells transfected
with lentivirus containing Ihh shRNA were used for further analysis
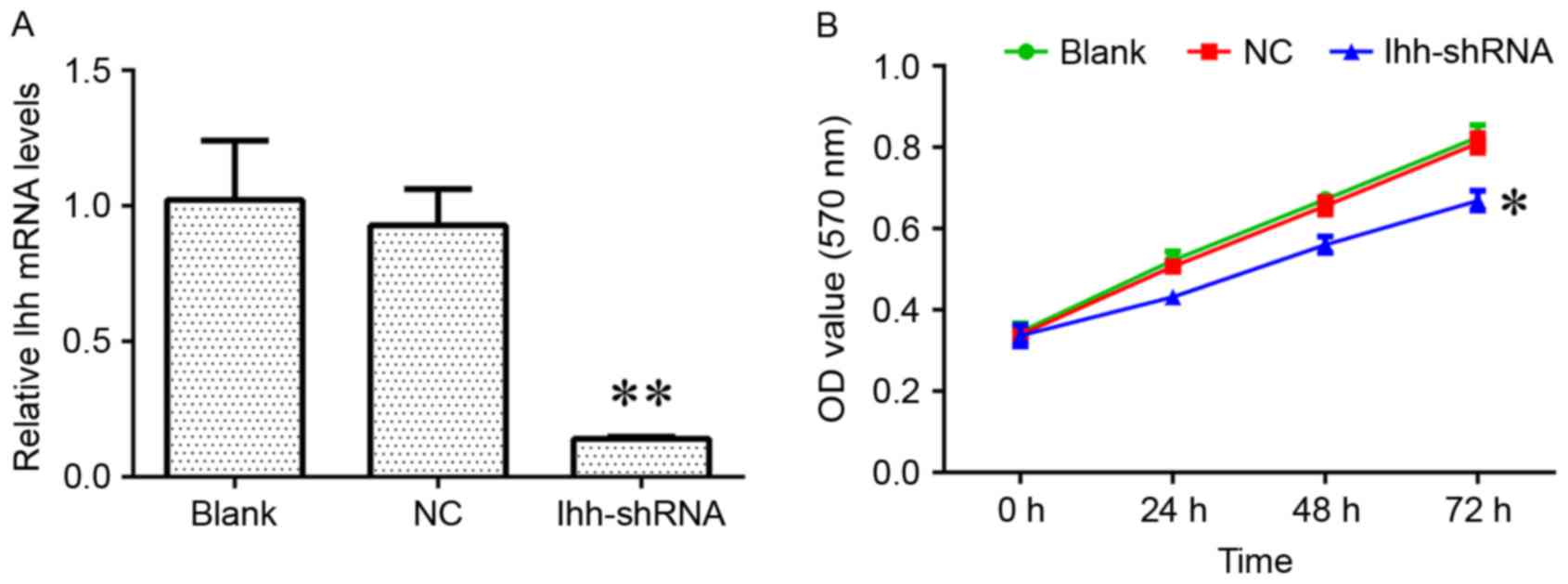
(Fig. 1A). The results of the
present study demonstrated that the decreased expression of Ihh
induced a significant inhibition of cell growth and markedly
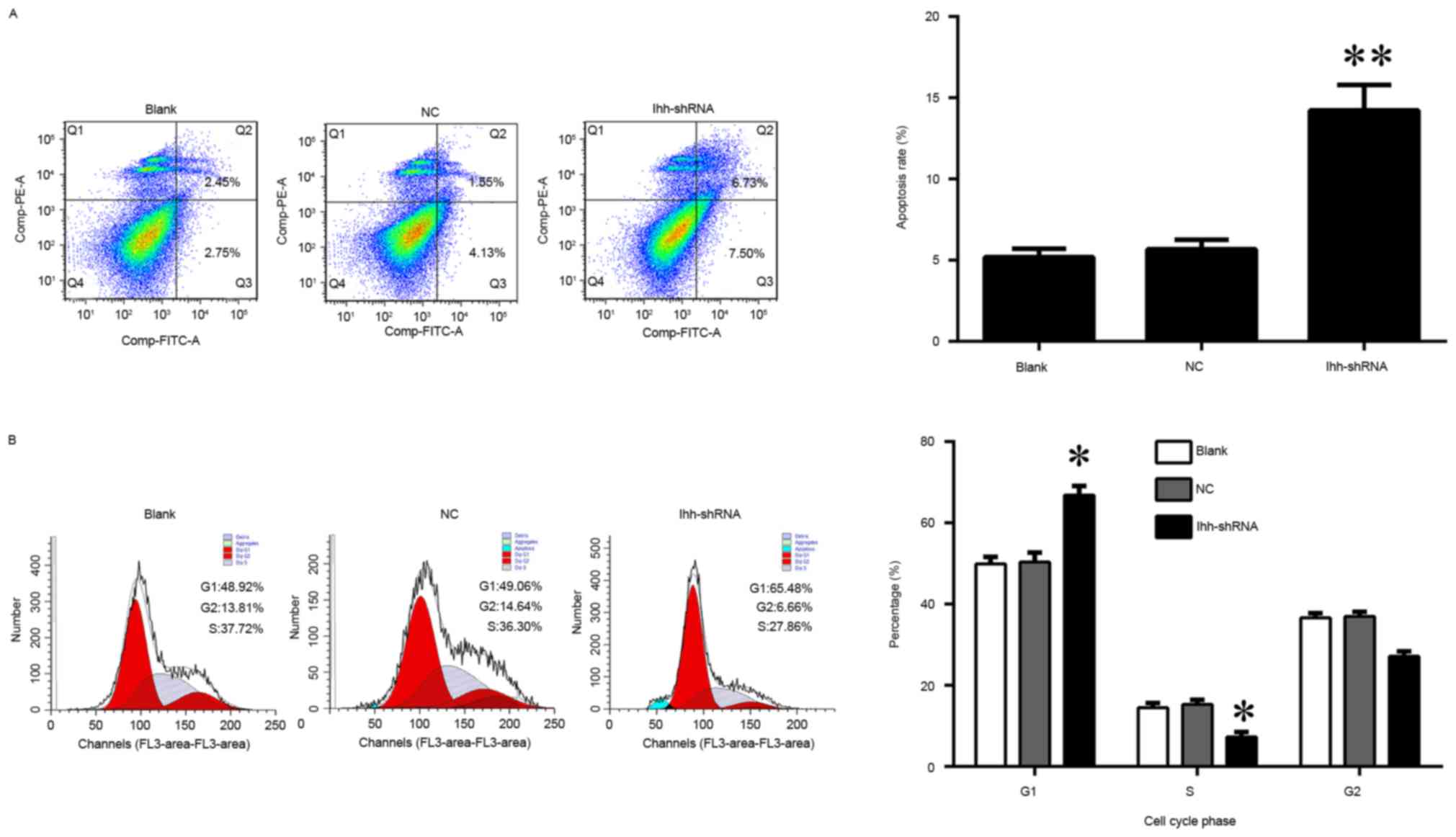
increased the apoptosis rate, compared with the NC group (Figs. 1B and 2A). In addition, the knockdown of Ihh
significantly decreased the percentage of cells in the S phase,
while increasing the percentage in the G1 phase, indicating a cell
cycle arrest at G1 to S phase (Fig.
2B).
Knockdown of Ihh suppresses osteoblast
cell differentiation
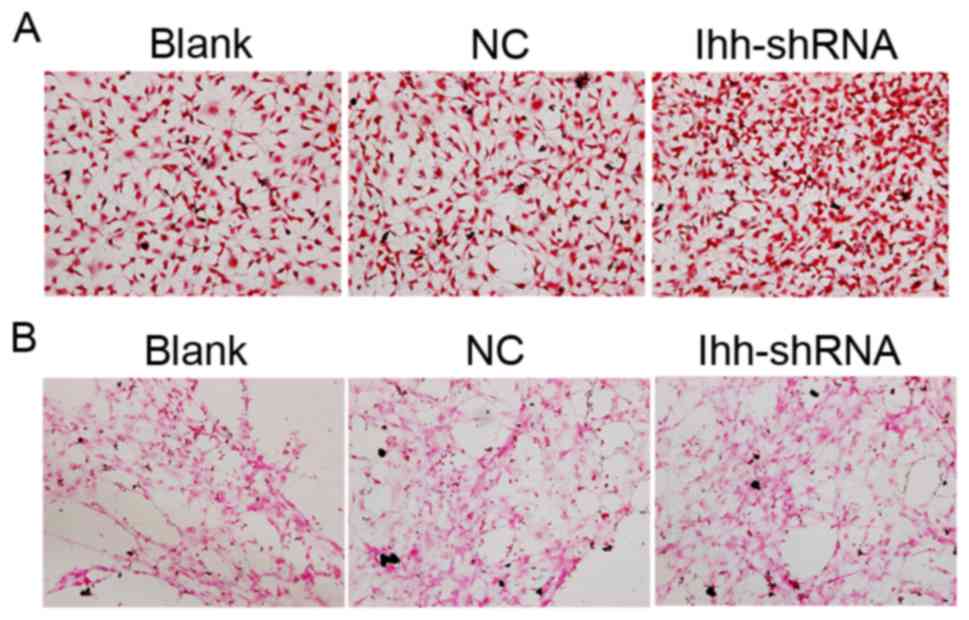
ALP is an enzyme which serves a role in bone
mineralization. ALP activity was determined in the present study.
Compared with the NC group, the ALP activity was decreased in
chondrocytes with knocked-down Ihh (Fig. 3A). Calcification and mineralization
of the bone matrix is essential for the strength and rigidity of
the spinal skeletal system. In order to estimate the degree of
mineralization and calcification, von Kossa staining following Ihh
knockdown was performed. Representative images of von Kossa
staining were obtained by brightfield microscopy, as presented in
Fig. 3B. Compared with the NC
group, the mineral deposition of osteoblasts lacking Ihh was
decreased.
Effects of Ihh on TGF-β/Smad and
OPG/RANKL signaling
The present study further determined the effects of
Ihh on TGF-β/Smad and OPG/RANKL signaling, which is associated with
bone differentiation and generation. As exhibited in Fig. 4, the mRNA expression of COL10A,
RANKL, Smad2 and Smad3 was significantly upregulated, although the
levels of OCN, OPG, PTHrP, RUNX2 and TGF-β were significantly
decreased, following knockdown of Ihh in osteoblast cells. The
protein expression of these genes was analyzed using western
blotting. As presented in Fig. 5,
the protein expression of COL10A, RANKL, Smad2 and Smad3 was
significantly upregulated, while the levels of OCN, OPG, PTHrP,
RUNX2 and TGF-β were significantly decreased, following knockdown
of Ihh in osteoblast cells. The results of the present study
suggested that the inhibitory effects of Ihh knockdown on bone
differentiation and generation are associated with the TGF-β/Smad
and OPG/RANKL signaling pathways.
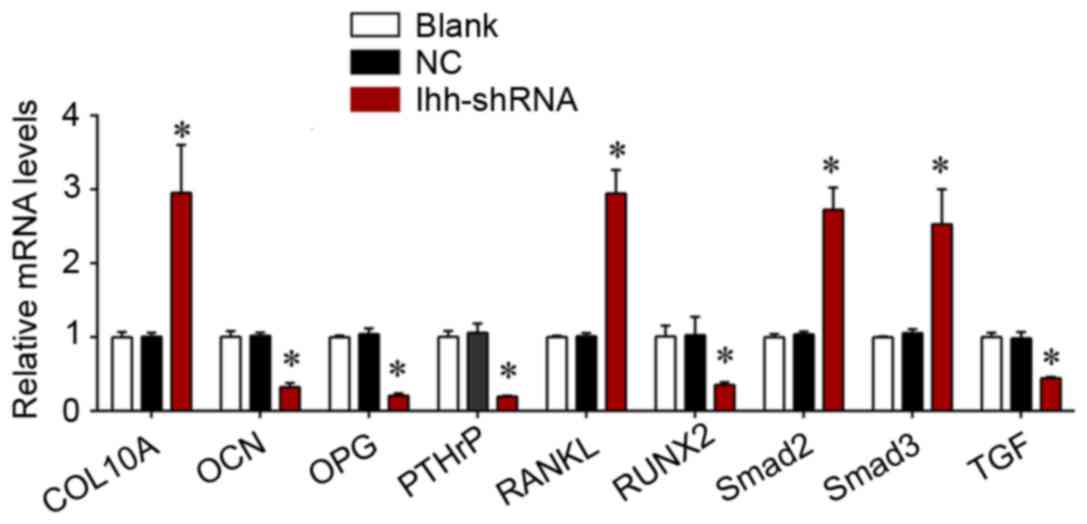 | Figure 4.Reverse transcription-quantitative
polymerase chain reaction analysis was used for detecting the mRNA
expression of COL10A, RANKL, Smad2, Smad3, OCN, OPG, PTHrP, RUNX2
and TGF-β in MC3T3-E1 cells following transfection with Ihh shRNA
lentivirus or empty lentivirus. *P<0.05 vs. NC. COL10A, collagen
α-1 (X) chain; RANKL, tumor necrosis factor ligand superfamily
member 11; Smad, mothers against decapentaplegic homolog; OCN,
osteocalcin; OPG, tumor necrosis factor receptor superfamily member
11B; PTHrP, parathyroid hormone-related protein; RUNX2,
runt-related transcription factor 2; TGF-β, transforming growth
factor-β; Ihh, Indian hedgehog protein; shRNA, short hairpin RNA;
NC, negative control. |
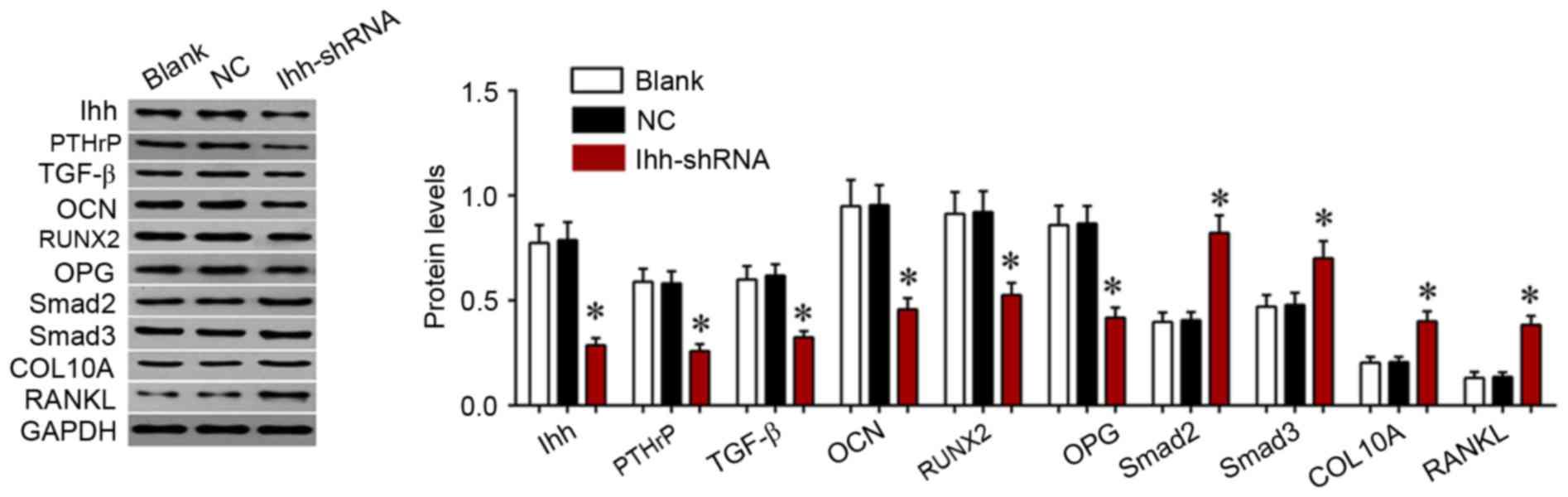 | Figure 5.Western blot analysis of the
expression of COL10A, RANKL, Smad2, Smad3, OCN, OPG, PTHrP, RUNX2
and TGF-β in MC3T3-E1 cells following transfection with Ihh shRNA
lentivirus or empty lentivirus. *P<0.05 vs. NC. COL10A, collagen
α-1 (X) chain; RANKL, tumor necrosis factor ligand superfamily
member 11; Smad, mothers against decapentaplegic homolog; OCN,
osteocalcin; OPG, tumor necrosis factor receptor superfamily member
11B; PTHrP, parathyroid hormone-related protein; RUNX2,
runt-related transcription factor 2; TGF-β, transforming growth
factor-β; Ihh, Indian hedgehog protein; shRNA, short hairpin RNA;
NC, negative control |
Discussion
In the present study, it was observed that the
knockdown of Ihh induced a significant inhibition of cell growth
and markedly increased the apoptosis rate, compared with the NC
group, in osteoblasts. Downregulation of Ihh resulted in a cell
cycle arrest at the G1 to S phase in osteoblasts. In addition,
knockdown of Ihh decreased the ALP activity and mineral deposition
in osteoblasts. The inhibitory roles of Ihh downregulation in
osteoblast growth and differentiation may be associated with the
TGF-β/Smad and OPG/RANKL signaling pathways.
Various hormones and factors, including Hh proteins
and transcription factors, regulate osteoblast differentiation. Ihh
is an essential transcription factor for osteoblast differentiation
and bone formation, which acts by interacting with bone
morphogenetic proteins (11).
Osteoblast MC3T3-E1 cells expressing Ihh exhibited increased ALP
activity, which was synergistically enhanced by bone morphogenetic
protein-2, indicating that Hh proteins directly act on osteogenic
precursor cells and osteoblasts to stimulate osteogenic
differentiation (12). The removal
of smoothened homolog, which triggers Hh signaling, from
perichondrial cells prevents the formation of a normal bone collar
and abolishes the development of the primary spongiosa (13,14).
Ihh knockdown results in markedly foreshortened limbs and a
complete absence of mature osteoblasts, and acts as an important
positive regulator of cranial bone ossification (15). Ectopic Ihh promotes the progression
of the bone collar towards the epiphysis, and Hh signaling is found
to be required in BMP-induced osteogenesis in cultures of a
limb-bud cell line, suggesting that Ihh signaling is directly
required for the osteoblast lineage in the developing long bones
and that Ihh functions in conjunction with other factors such as
BMPs to induce osteoblast differentiation (14). In addition, Hh proteins promote
osteoblast recruitment and differentiation, possibly by stimulating
PTHrP expression and secretion (16). RUNX2 is a transcription factor
which is required for osteoblast differentiation. RUNX2
overexpression has been demonstrated to stimulate chondrocyte
maturation, as indicated by increases in ALP activity,
mineralization, and COL10A and matrix metalloproteinase-13
expression, and by maintenance of Ihh expression, whereas PTHrP
treatment blocked RUNX2 expression and its promaturation effects
(17). However, forced expression
of RUNX2 in skeletogenic cells restores bone formation in the
Runx2-null, although not in the Ihh-null, embryo, indicating that
in addition to Runx2, Ihh requires other effectors to induce
osteoblast differentiation, including zinc finger proteins Gli2 and
Gli3 (8,18,19).
Wnt signaling acts downstream of Ihh signaling and is required in
osteoblast maturation during endochondral bone formation; to
enhance fracture repair or bone formation, Ihh signaling requires
enhancement at the early stages by controlling osteoblast
proliferation and differentiation, whereas Wnt signaling may be
upregulated later in differentiated osteoblasts (1,20).
Notably, osteoblast differentiation and bone formation may occur in
conditions of specific ablation of Ihh in the limb mesenchyme
(2,21).
In the present study, it was observed that the mRNA
and protein expression of osteoblastic and osteocytic markers,
including COL10A, RANKL, Smad2 and Smad3 was notably upregulated,
while the levels of OCN, OPG, PTHrP, RUNX2 and TGF-β were
significantly decreased following knockdown of Ihh in osteoblasts.
TGF-β signaling exerts important roles in embryonic skeletal
development and postnatal bone homeostasis, by transducing signals
in the canonical Smad-dependent signaling pathway to regulate
mesenchymal stem cell differentiation during skeletal development,
bone formation and bone homeostasis (22). The coordinated activity of RUNX2
and TGF-β-activated Smad proteins is important for the formation of
the skeleton (23). Knockout or
mutation of TGF-β signaling-associated genes in mice leads to bone
abnormalities of varying severity (24). For example, AlCl3 has been
demonstrated to inhibit osteoblast mineralization via the
TGF-β1/Smad signaling pathway in rat osteoblasts (25).
In conclusion, the knockdown of Ihh suppressed
osteoblast growth and differentiation via a mechanism which may be
associated with the TGF-β/Smad and OPG/RANKL signaling pathways.
The results of the present study suggested that osteoblast-derived
Ihh is essential for maintaining the growth plate. The manipulation
of Ihh expression or its signaling components may be of benefit in
the treatment of skeletal diseases.
Acknowledgements
The present study was supported by the Natural
Science Foundation of Hunan Province, China (grant no. 2016JJ3160)
and the National Natural Science Foundation of China (grant no.
81472145).
References
|
1
|
Day TF and Yang Y: Wnt and hedgehog
signaling pathways in bone development. J Bone Joint Surg Am. 90
Suppl 1:S19–S24. 2008. View Article : Google Scholar
|
|
2
|
Yang J, Andre P, Ye L and Yang YZ: The
Hedgehog signalling pathway in bone formation. Int J Oral Sci.
7:73–79. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xing W, Cheng S, Wergedal J and Mohan S:
Epiphyseal chondrocyte secondary ossification centers require
thyroid hormone activation of Indian hedgehog and osterix
signaling. J Bone Miner Res. 29:2262–2275. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Komori T: Signaling networks in
RUNX2-dependent bone development. J Cell Biochem. 112:750–755.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hart GT, Burton EL and Mincer HH: Use of
screening blood studies in dental schools. J Dent Educ. 55:735–737.
1991.PubMed/NCBI
|
|
6
|
Kronenberg HM and Chung U: The parathyroid
hormone-related protein and Indian hedgehog feedback loop in the
growth plate. Novartis Found Symp. 232:144–157. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Colnot C, de la Fuente L, Huang S, Hu D,
Lu C, St-Jacques B and Helms JA: Indian hedgehog synchronizes
skeletal angiogenesis and perichondrial maturation with cartilage
development. Development. 132:1057–1067. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shimoyama A, Wada M, Ikeda F, Hata K,
Matsubara T, Nifuji A, Noda M, Amano K, Yamaguchi A, Nishimura R
and Yoneda T: Ihh/Gli2 signaling promotes osteoblast
differentiation by regulating Runx2 expression and function. Mol
Biol Cell. 18:2411–2418. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cai JQ, Huang YZ, Chen XH, Xie HL, Zhu HM,
Tang L, Yang ZM, Huang YC and Deng L: Sonic hedgehog enhances the
proliferation and osteogenic differentiation of bone marrow-derived
mesenchymal stem cells. Cell Biol Int. 36:349–355. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yamaguchi A, Komori T and Suda T:
Regulation of osteoblast differentiation mediated by bone
morphogenetic proteins, hedgehogs, and Cbfa1. Endocr Rev.
21:393–411. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakamura T, Aikawa T, Iwamoto-Enomoto M,
Iwamoto M, Higuchi Y, Pacifici M, Kinto N, Yamaguchi A, Noji S,
Kurisu K and Matsuya T: Induction of osteogenic differentiation by
hedgehog proteins. Biochem Biophys Res Commun. 237:465–469. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pacheco M, Valencia M, Caparros-Martin JA,
Mulero F, Goodship JA and Ruiz-Perez VL: Evc works in chondrocytes
and osteoblasts to regulate multiple aspects of growth plate
development in the appendicular skeleton and cranial base. Bone.
50:28–41. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Long F, Chung UI, Ohba S, McMahon J,
Kronenberg HM and McMahon AP: Ihh signaling is directly required
for the osteoblast lineage in the endochondral skeleton.
Development. 131:1309–1318. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lenton K, James AW, Manu A, Brugmann SA,
Birker D, Nelson ER, Leucht P, Helms JA and Longaker MT: Indian
hedgehog positively regulates calvarial ossification and modulates
bone morphogenetic protein signaling. Genesis. 49:784–796. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jemtland R, Divieti P, Lee K and Segre GV:
Hedgehog promotes primary osteoblast differentiation and increases
PTHrP mRNA expression and iPTHrP secretion. Bone. 32:611–620. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Iwamoto M, Kitagaki J, Tamamura Y, Gentili
C, Koyama E, Enomoto H, Komori T, Pacifici M and Enomoto-Iwamoto M:
Runx2 expression and action in chondrocytes are regulated by
retinoid signaling and parathyroid hormone-related peptide (PTHrP).
Osteoarthritis Cartilage. 11:6–15. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tu X, Joeng KS and Long F: Indian hedgehog
requires additional effectors besides Runx2 to induce osteoblast
differentiation. Dev Biol. 362:76–82. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hilton MJ, Tu X, Cook J, Hu H and Long F:
Ihh controls cartilage development by antagonizing Gli3, but
requires additional effectors to regulate osteoblast and vascular
development. Development. 132:4339–4351. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Maeda Y, Nakamura E, Nguyen MT, Suva LJ,
Swain FL, Razzaque MS, Mackem S and Lanske B: Indian Hedgehog
produced by postnatal chondrocytes is essential for maintaining a
growth plate and trabecular bone. Proc Natl Acad Sci USA. 104:pp.
6382–6387. 2007; View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Amano K, Densmore MJ and Lanske B:
Conditional deletion of Indian hedgehog in limb mesenchyme results
in complete loss of growth plate formation but allows mature
osteoblast differentiation. J Bone Miner Res. 30:2262–2272. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hendy GN, Kaji H, Sowa H, Lebrun JJ and
Canaff L: Menin and TGF-beta superfamily member signaling via the
Smad pathway in pituitary, parathyroid and osteoblast. Horm Metab
Res. 37:375–379. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen G, Deng C and Li YP: TGF-β and BMP
signaling in osteoblast differentiation and bone formation. Int J
Biol Sci. 8:272–288. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu M, Chen G and Li YP: TGF-β and BMP
signaling in osteoblast, skeletal development, and bone formation,
homeostasis and disease. Bone Res. 4:160092016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun X, Cao Z, Zhang Q, Li M, Han L and Li
Y: Aluminum trichloride inhibits osteoblast mineralization via
TGF-β1/Smad signaling pathway. Chem Biol Interact. 244:9–15. 2016.
View Article : Google Scholar : PubMed/NCBI
|