Introduction
Angiogenesis, the formation of new blood vessels
initiated by endothelial cell proliferation and migration, is
necessary for tumors to provide nutrients and remove metabolic
wastes continuously (1). Tumors
cannot grow beyond a certain size without concomitant vascular bed
expansion. Therefore, angiogenesis is a key step in tumor growth,
and anti-angiogenesis has become an attractive therapeutic strategy
in cancer treatment (2–4). Angiogenesis is regulated by multiple
growth factors and cytokines. Among these factors, vascular
endothelial growth factor (VEGF), generated by most cancer cells
and endothelial cells as well, is one of the most potent angiogenic
factors involved in tumor growth (5). VEGF stimulates endothelial cell
proliferation, migration and tube formation by binding its receptor
tyrosine kinases expressed on endothelial cells. In these receptor
tyrosine kinases, VEGF receptor 2 (VEGFR2) mediates the major
function (6). Activation of VEGFR2
leads to activation of various downstream signal transduction
proteins, including focal adhesion kinase (FAK), phosphoinositide
3-kinase (PI3K)/protein kinase B (AKT) (7). Inhibition of VEGF activity or/and
disabling the function of VEGFR2 are both accepted as potential
strategies for antiangiogenesis intervention in tumor
treatment.
Natural products present promising application in
cancer therapy due to their special pharmacological activities and
low toxicity. Cynanchum auriculatum (C. auriculatum) is a
traditional herb medicine mainly distributed in China and other
Asian countries (8). C-21
steroidal glycoside, the major active component of C. auriculatum,
showed multiple bioactivities, including anti-proliferation and
invasion of tumor cells (9–12).
Caudatin, isolated from the root of C. auriculatum, is a species of
C-21 steroidal glycosides (13).
The chemical structure of caudatin can be found in our previous
publication (8). It is reported
that caudatin could inhibit tumor growth via inducing cell cycle
arrest in several cell lines (10,13)
or inducing apoptosis through various signal pathway in several
cell lines such as human hepatoma cell line SMMC7721 (10,14,15)
and HepG2 cells (10,16,17),
human alveolar basal epithelial cell line A549 (18) and gastric cancer cells (19). In our previous studies, caudatin
was demonstrated effective in inducing cell cycle arrest (8) and inducing apoptosis (13) in human glioma cells in vitro
and in vivo. However, the anti-angiogenesis effect of
caudatin on human glioma has not been explored yet, and the
underlying mechanism remains elusive.
Herein, the antiangiogenesis properties and the
underlying mechanism of caudatin on human glioma cells were
investigated, and the results suggested that caudatin significantly
inhibited endothelial cell proliferation and blocked the human
umbilical vein endothelial cell (HUVEC) migration, invasion and
capillary-like tube formation by disturbing the VEGF-VEGFR2-AKT/FAK
signal axis, and eventually resulted in the inhibition of glioma
growth in vivo.
Materials and methods
Materials
Caudatin was purchased from Sigma-Aldrich (St.
Louis, MO, USA), and dissolved with dimethyl sulfoxide (DMSO). Cell
culture reagents and fetal bovine serum (FBS) were procured from
Invitrogen (Carlsbad, CA, USA). The second antibody IgG (cat. no.
3452) and all of the primary antibodies used in this study were
both purchased from Cell Signaling Technology (Beverly, MA, USA),
including p-FAK (cat. no. 3281), VEGF (cat. no. 2463), VEGFR2 (cat.
no. 9698), p-VEGFR2 (cat. no. 2478S), AKT (cat. no. 9272), p-AKT
(cat. no. 4060), Ki67 (cat. no. 9027) and CD34 (cat. no. 3569).
LY294002 was obtained from Calbiochem (San Diego, CA, USA).
PF-562271 was obtained from Selleck Chemicals (Houston, TX, USA).
All solvents used were of high-performance liquid chromatography
(HPLC) grade. Water used in this study was provided by a Milli-Q
water purification system from Merck Millipore (Billerica, MA,
USA).
Cell culture
HUVECs were purchased from the KeyGen Biotech
(Nanjing, China) and cultured in DMEM-F12 medium supplemented with
10% FBS, 100 U/ml penicillin and 50 U/ml streptomycin. Cells were
maintained in a humidified incubator of 5% CO2 at
37°C.
Measurement of cell viability
Cell viability was measured with
3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazoliumbromide (MTT)
assay. Briefly, HUVECs were cultured in a 96-well plate
(6×103 cells/well) for 24 h and then were treated with
indicated concentrations of caudatin for 48 h. After that, 20 µl
MTT (5 mg/ml) solution/well was added and maintained for 4 h. Then,
the supernatant was removed, and 200 µl DMSO/well was added,
followed by 20 min shake to dissolve the formazan crystals.
Subsequently, the absorbance at 570 nm was measured with a
microplate reader (Molecular Device, USA). Cell viability was
expressed as percentage of control (as 100%). Cell morphology was
observed by inverted microscope (magnification, ×200).
Migration assay
Wound healing assay was used to evaluate cell
migration. HUVECs seeded in 6-well culture plate were incubated to
full monolayer. Monolayer HUVECs were wounded by scratching with a
pipette tip and washed twice with phosphate buffer solution (PBS).
Fresh medium containing 1% FBS was then added together with
caudatin or other reagents as designed. Cells were photographed
under a Nikon Ti-S inverted microscope at the beginning and after
incubation for 48 h (magnification, ×100; Nikon Corporation, Tokyo,
Japan). Cell migration distance was measured by image plus
software, and the migrated rate was expressed as percentage of
control. Three independent experiments were performed.
Invasion assay
Transwell assay was used to determine the effect of
caudatin on HUVECs invasion. Transwell chambers (BD Biosciences,
Bedford, MA, USA) containing a 6.5-mm-diameter polycarbonate filter
with a pore size of 8 µm were used. Briefly, each filter was coated
with 60 µl Matrigel (1 mg/ml; BD Biosciences) diluted with DMEM-F12
and incubated at 37°C for 45 min. HUVECs (4×104 cells)
suspended in 100 µl DMEM-F12 (FBS-free) were seeded in the filter,
and treated with or without 100 µM caudatin for 24 h. Complete
DMEM-F12 (600 µl, 10% FBS) was added into the chamber below. VEGF
(200 ng/ml) and cisplatin (5 µM) were employed as the positive and
negative control, respectively. After incubation, the cells on the
filter were wiped away, and the invaded cells below the filter were
washed with PBS, fixed with 90% ethanol and stained with 0.1%
crystal violet. The invaded cells were calculated by manual
counting with a Nikon Ti-S inverted microscope (magnification,
×200). The invaded rate was expressed as percentage of the control.
Three independent experiments were performed.
Tube formation
Capillary-like tube formation assay was used to
further detect the antiangiogenesis effect of caudatin. Matrigel
was pipetted into 48-well plate (60 µl/well) and polymerized at
37°C for 45 min. HUVECs (100 µl, 1×104 cells/well) mixed
with 100 µM caudatin were seeded into Matrigel pre-coated wells and
incubated at 37°C with 5% CO2 for 24 h. Cells were
photographed and the tube number was calculated by manual counting
using a Nikon Ti-S inverted microscope (magnification, ×100). Tube
formation was scored as follows: A three branch point event was
defined as one tubular structure. Eight random fields per well were
quantified by manual counting. Three independent experiments were
performed.
Western blotting
Western blot analysis was performed as described
before (8). HUVECs were cultured
in dishes (10 cm) and treated with indicated concentrations of
caudatin or other reagents for 48 h. Then, cells were washed twice
by PBS and lysed in RIPA lysis buffer (1xPBS, 1% NP40, 0.1% SDS, 5
mM EDTA, 0.5% sodium deoxycholate and 1 mM sodium orthovanadate)
for 20 min in the ice environment. Lysates were centrifuged and
supernatants were kept as total cell protein. The concentration of
total cell protein was quantified by bicinchoninic acid assay (BCA;
Beyontime, Beijing, China) according to the manufacturer's
instructions. Total protein was added with sample loading buffer,
boiled for 10 min to be denatured, and stored at −80°C environment
for subsequent SDS-polyacrylamide gel electrophoresis (SDS-PAGE)
analysis. Briefly, total protein (40 µg/lane) was separated by
SDS-PAGE and then transferred onto polyvinylidene difluoride (PVDF)
membranes (Merck Millipore). After that, the membranes were blocked
with 5% bovine serum albumin (BSA) at room temperature for 2 h.
Then, the membranes were incubated with specific primary antibodies
overnight at 4°C and secondary antibodies for 2 h at room
temperature sequentially. At last, target protein bands were
visualized with ECL under an Imaging System (ChemiDoc MP, Bio-Rad).
β-actin plays as a positive control.
In vivo antiangiogenesis
U251 human glioma cells xenograft model was
constructed as described previously (13). Briefly, 1×107 U251 cells
in 100 µl serum-free medium were subcutaneously injected into the
right oxter of male nude mice. When tumors grew to 60
mm3 on average after 8 days, mice were randomly divided
into control group and the caudatin-treated groups (25 and 100
mg/kg). Drugs were given through caudal vein injection every other
day for 16 days. At the termination of the experiment, tumors were
collected and used for western blotting and immunohistochemical
(IHC) assay (magnification, ×200). All animal experiments were
approved by the Animal Experimentation Ethics Committee.
Results
Caudatin inhibits cell proliferation
of HUVECs
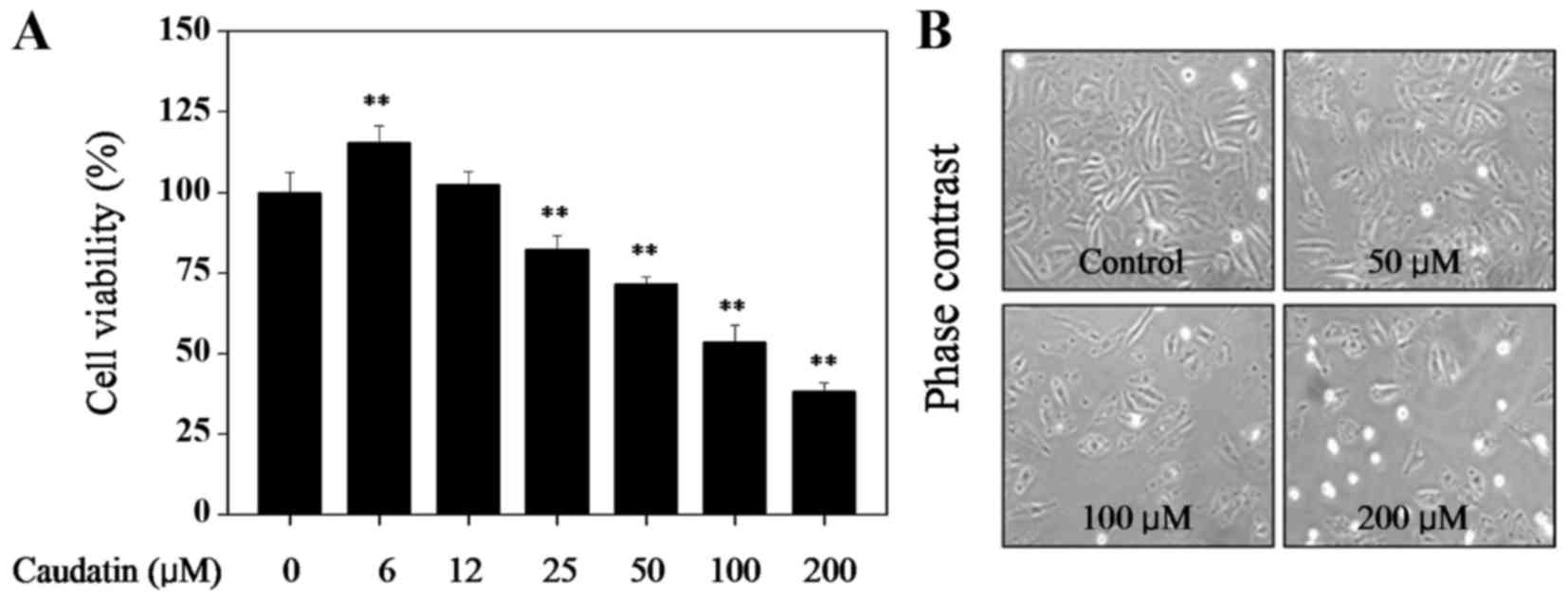
Firstly, MTT assay was employed to examine the
anti-proliferation activity of caudatin against HUVECs. As shown in
Fig. 1, treatment of HUVECs with
caudatin (below 12 µM) showed no cytotoxicity. Treatment of cells
with 6 µM caudatin slightly promoted U251 cells growth. However,
caudatin (25–200 µM) significantly inhibited U251 cells growth in a
dose-dependent manner. For instance, cells exposed to 50, 100 and
200 µM of caudatin for 48 h markedly inhibited the HUVECs viability
to 71.5, 53.6 and 38.1%, respectively. This result suggests that
caudatin may act as a potential cytostatic agent in hunting HUVECs
growth.
Caudatin blocks HUVECs migration,
invasion and capillary-like tube formation
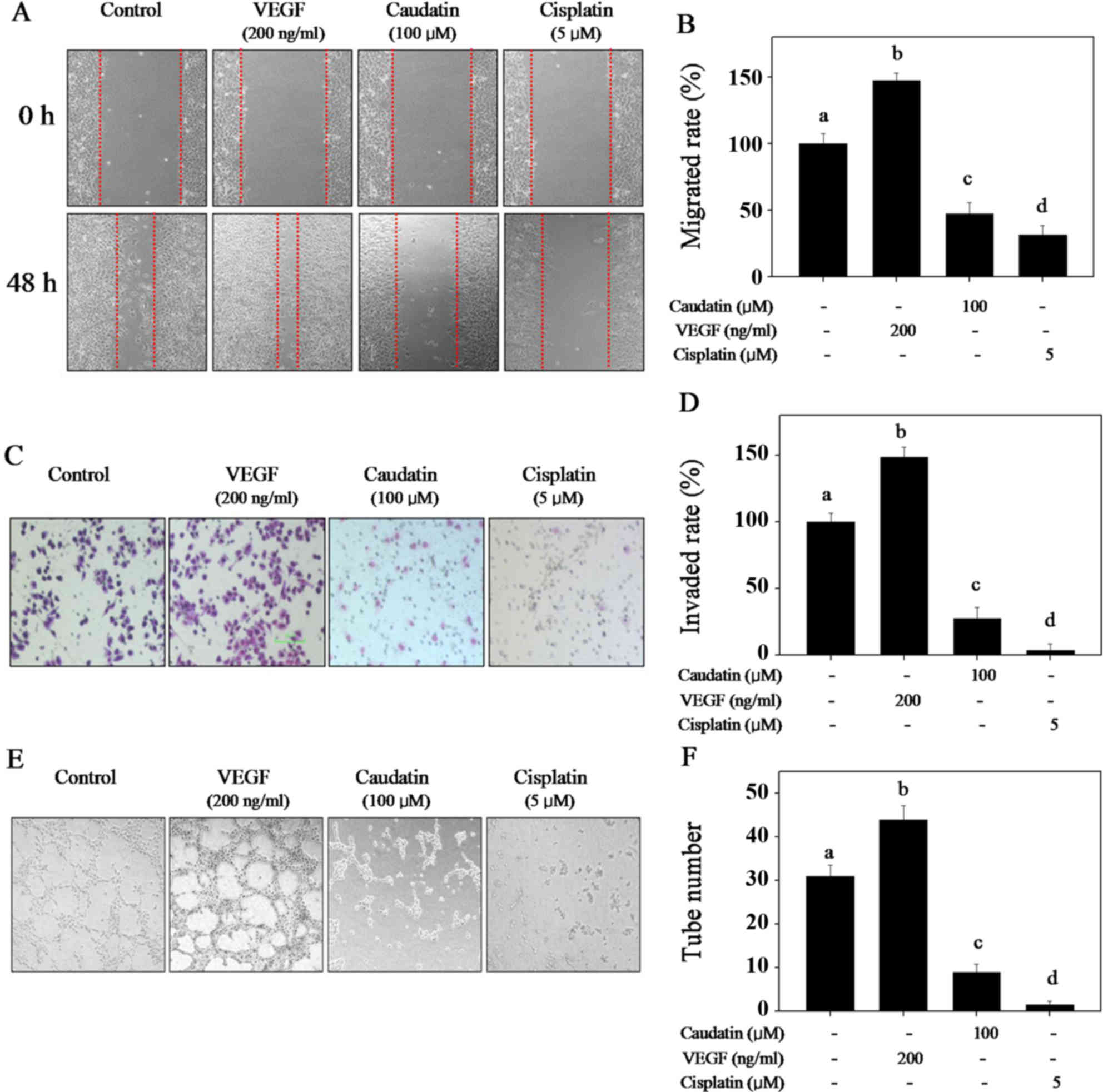
Wound-healing assay, Transwell assay and
capillary-like tube formation were used to evaluate the inhibition
effect of caudatin on cell migration, invasion and tube formation,
respectively. VEGF and cisplatin were employed as positive and
negative control respectively in these assays to evaluate
caudatin's potential. Cell migration and invasion is critical for
endothelial cells to form blood vessels during angiogenesis. As
shown in Fig. 2, VEGF obviously
promoted cell migration (Fig. 2A and
B), invasion (Fig. 2C and D)
and capillary-like tube formation (Fig. 2E and F). However, caudatin (100 µM)
treatment effectively blocked HUVECs migration, invasion and
capillary-like tube formation (Fig.
2). The statistical data of migrated rate, invaded rate and the
tube formation further confirmed caudatin's inhibitory effects,
which is similar with that of cisplatin. These results showed that
caudatin has the potential to block the capillary-like tube
formation in vitro by inhibiting the HUVECs migration and
invasion.
Caudatin suppresses FAK
phosphorylation in HUVECs
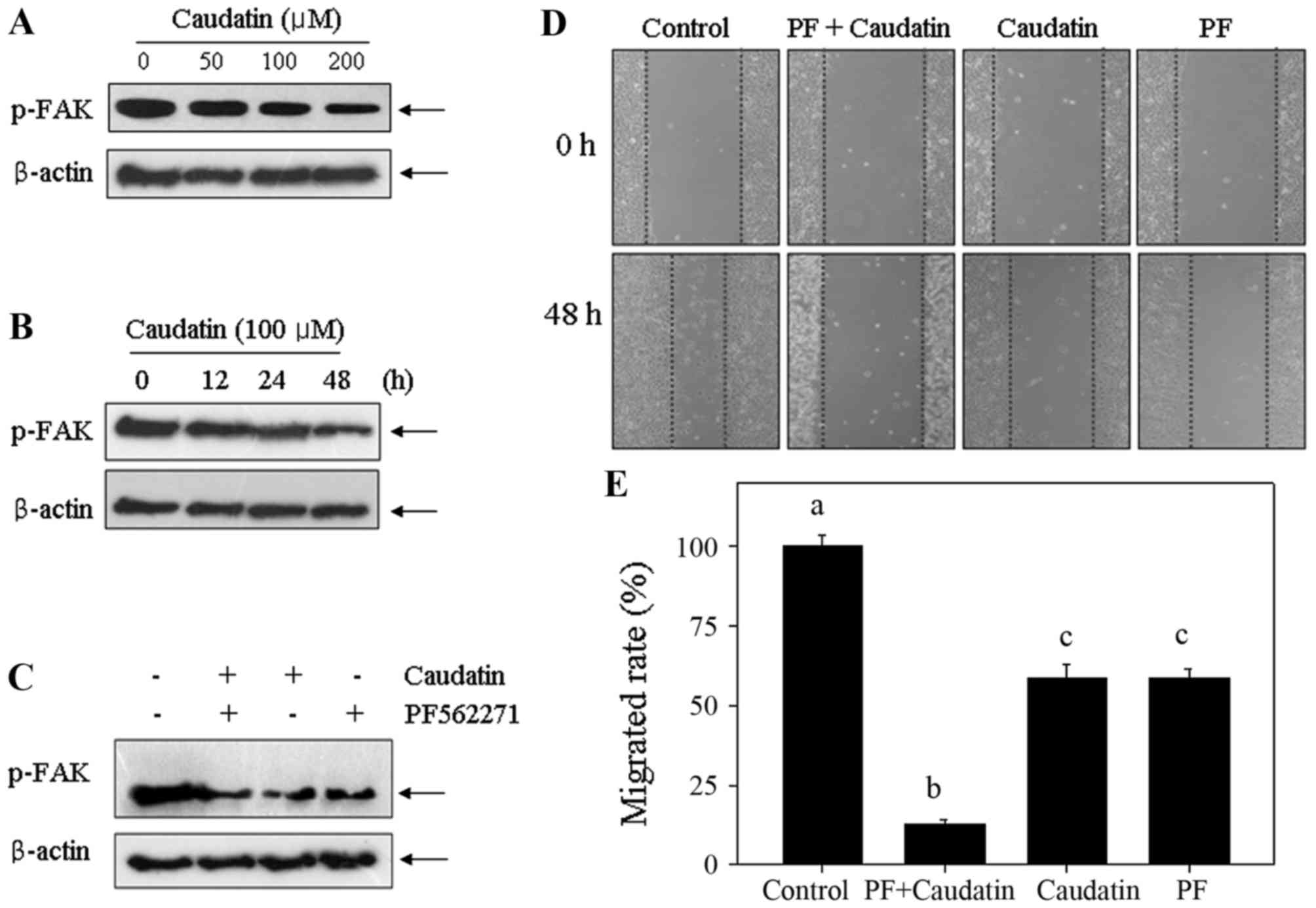
FAK is an upstream kinase that has a key role in the
regulation of cell migration and invasion. Therefore, it was
interested to investigate whether FAK was involved in the
caudatin-induced inhibition of HUVEC migration and invasion. A
specific antibody against the phosphorylated (activated) form of
FAK was used (Fig. 3). The results
showed that caudatin treatment caused significant dephosphorylation
of FAK in a dose- and time-dependent manner (Fig. 3A and B). To further confirm the
role of FAK, wound healing assay was operated using the specific
FAK inhibitor (PF562271). As shown in Fig. 3D and E, pretreatment of HUVECs with
PF562271 (10 nM) for 1 h markedly enhanced caudatin-induced
inhibition against cell migration, which confirmed the effect of
inactivation FAK on cell migration inhibition. Results in protein
levels further confirmed this conclusion. Treatment of HUVECs with
caudatin caused significant inactivation of FAK, and this effect
was enhanced at the presence of PF562271 (Fig. 3C). Taken together, the results
above suggest that caudatin can inhibit cell migration of HUVECs
through FAK dephosphorylation.
Caudatin disturbs the VEGF-VEGFR2-AKT
signal axis
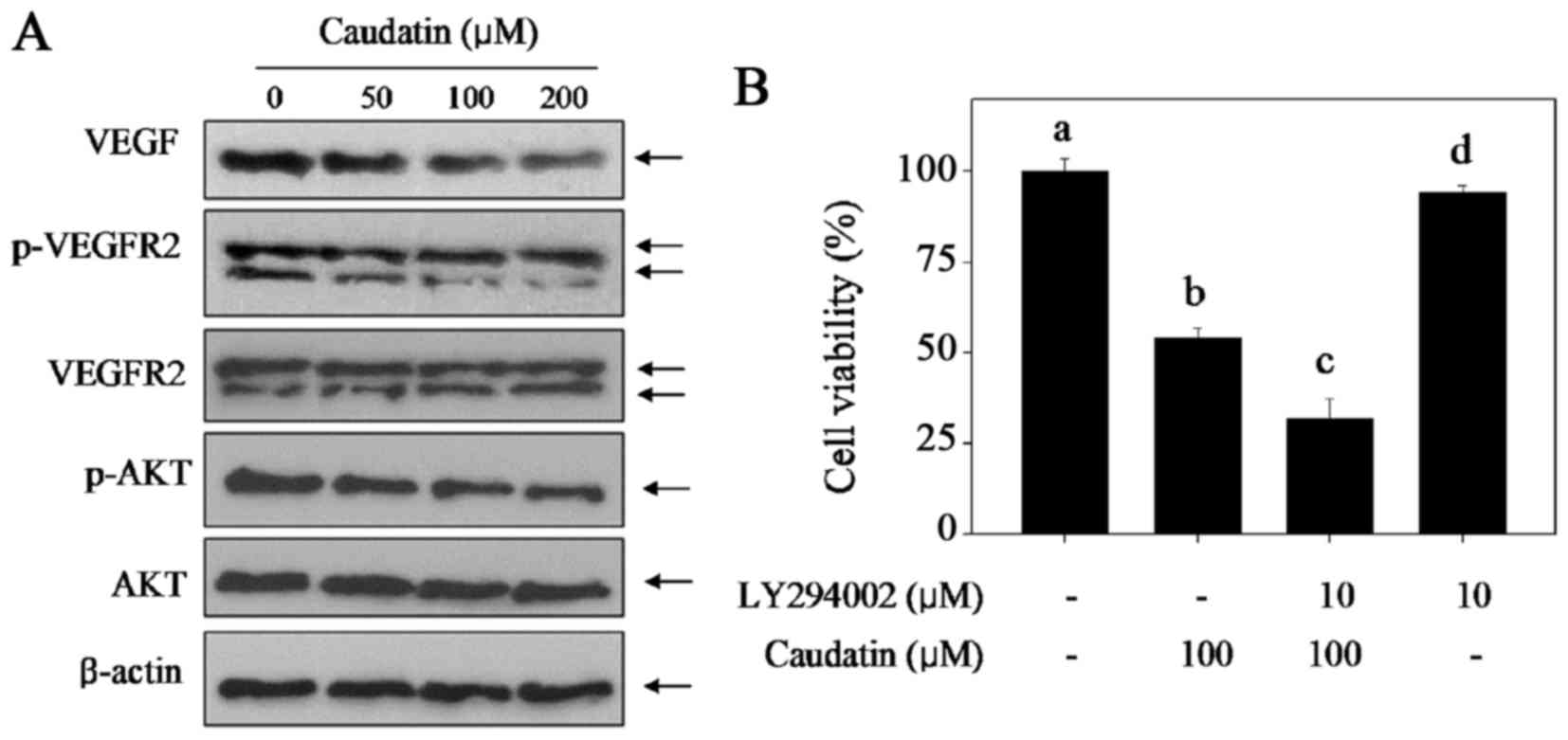
The VEGF-VEGFR2- AKT signal axis has key roles in
regulation of cell proliferation, cell growth, survival and
angiogenesis. Therefore, it was of interest to investigate whether
the VEGF-VEGFR2-AKT signal axis was involved in the
caudatin-induced inhibition against HUVECs proliferation and
angiogenesis. Firstly, western blotting assay was employed and
different concentrations of caudatin on the VEGF-VEGFR2-AKT signal
axis expression was assayed, and the results showed that caudatin
treatment significantly suppressed the expression of VEGF and
p-VEGFR2 in a dose-dependent manner (Fig. 4A), but caused no significant
changes in the expression of total-VEGFR2 and total-AKT. To further
confirm the role of the VEGF-VEGFR2- AKT signal axis, cell
viability was detected by MTT assay using a specific AKT inhibitor
(LY294002). As shown in Fig. 4B,
pre-treatment of HUVECs with LY294002 (an AKT-upstream inhibitor)
for 1 h markedly enhanced caudatin-induced cell growth inhibition
against HUVECs, indicating that caudatin inhibited HUVEC
proliferation with AKT-dependent manner. These results indicated
that the disturbance of VEGF-VEGFR2-AKT signal axis contributed to
caudatin-induced inhibition against proliferation and
angiogenesis.
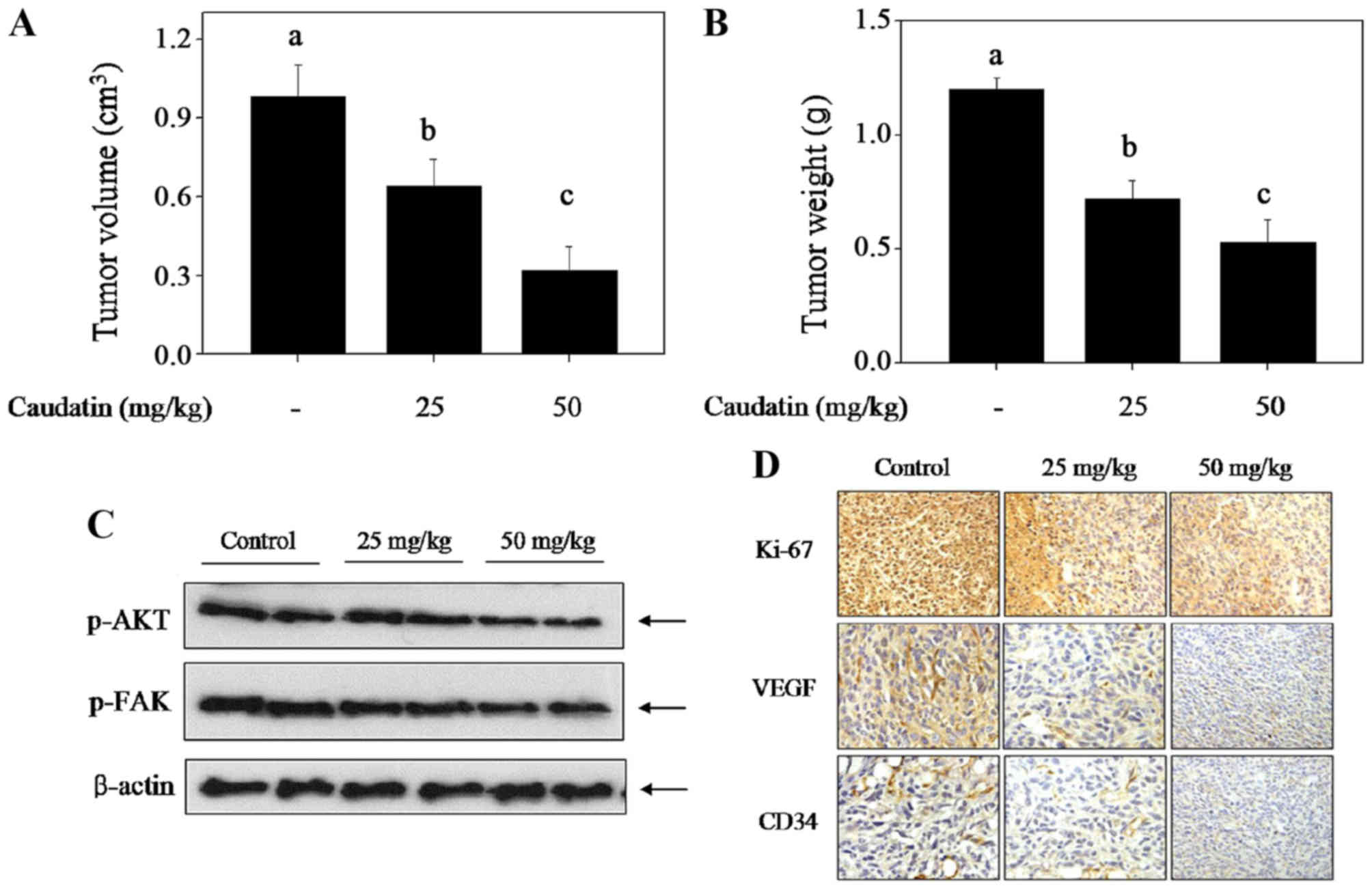
Caudatin hinders the glioma growth by
inhibiting in vivo angiogenesis
To investigate the antiangiogenesis effect of
caudatin in vivo, immune-deficient nude mice bearing U251
xenograft tumors were employed (Fig.
5). The results demonstrated that caudatin treatment in
vivo significantly inhibited U251 cell xenograft tumor growth,
as convinced by the decrease of tumour volume (Fig. 5A) and tumor weight (Fig. 5B) and ki-67 expression (Fig. 5D). Moreover, CD-34, one of the
endothelial cell markers, was detected in xenograft tumors by IHC
method, and the results clearly demonstrated that caudatin
treatment obviously abolished the tumor angiogenesis in
vivo. VEGF expression as one of the most important
pro-angiogenic factors also showed dramatically decrease (Fig. 5D). Furthermore, caudatin treatment
caused the dephosphorylation of AKT and FAK detected by western
blotting, which further affirmed the inhibitory effect of caudatin
on glioma angiogenesis. Taken together, these results above suggest
that caudatin can inhibit glioma cells growth by suppression of the
in vivo angiogenesis involving the FAK and AKT
dephosphorylation and inhibition of VEGF expression.
Discussion
Recently, great efforts have been made to identify
anticancer agents which can block insurgence of primary tumors or
recurrence. Current interest is focusing on the beneficial effects
of phytochemicals (20), which
have been found to influence some steps in cancer angiogenesis
besides their traditional application (21). Angiogenesis is essential to the
growth of solid tumors and inhibition of angiogenesis is an
effective and promising target for anticancer therapy. Caudatin is
a bioactive C-21 steroidal glycoside extracted from the root of C.
auriculatum. Our previous studies have revealed that caudatin had
the potential to inhibit human glioma cells growth in vitro
and in vivo through triggering cell cycle arrest or/and
apoptosis (8,13). However, little information about
the antiangiogenic effect of caudatin was available. Therefore, in
the present study, we evaluated the antiangiogenic effect and
mechanism of caudatin in human glioma in vitro and in
vivo.
Rapid proliferation of cancer cells usually causes
the intracellular hypoxia environment, which in turn accelerates
the development of new blood vessels (22). VEGF as the most important
pro-angiogenic factors was involved in almost every stage of tumor
angiogenesis (23–25). Increasing studies have indicated
that VEGF was capable of stimulating endothelial cell
proliferation, migration and formation of new capillaries from
pre-existing vasculature, which are all key steps in the process of
angiogenesis (26,27). VEGF can supply pro-survival and
pro-angiogenic signals to normal and tumor-derived endothelial
cells, which is mainly mediated by VEGF receptors, including
VEGFR1, VEGFR2 and VEGFR3 (28).
VEGFR2, a receptor tyrosine kinase, can be activated by
autophosphorylation through binging of VEGF, which subsequently
activated the secondary messengers including AKT, ERK and FAK to
regulate endothelial cell proliferation, migration and angiogenesis
(29–34). In the present study, exposure of
HUVECs to caudatin resulted in the suppression of VEGF and p-VEGFR2
expression. The results indicated that caudatin can act as the
inhibitor of VEGF and VEGFR2 to inhibit HUVECs migration, invasion
and angiogenesis.
It is reported that overexpression of FAK in
endothelial cells can promote the endothelial cell migration,
invasion and angiogenesis (31,35).
Inactivation of FAK by conditional knockout mice could disrupt
angiogenesis in late-stage embryos (36). Therefore, FAK inhibition may result
in an antiangiogenic effect and was considered as novel anticancer
strategy (32). Our data showed
that caudatin treatment time- and dose-dependent suppressed FAK
phosphorylation. FAK inhibitor (PF562271) effectively enhanced
caudatin-induced inhibition against HUVECs migration. The results
in vivo also that caudatin inhibited the glioma growth
involving the suppression of FAK phosphorylation. These results all
validated the contribution of FAK dephosphorylation to
caudatin-induced inhibition against HUVECs migration.
VEGF and VEGFR2 regulate endothelial cell survival
and proliferation through activating the PI3K/AKT signal (37). AKT is recruited during signal
transduction from growth factor receptors and intracellular
pathways (37). AKT activation not
only promotes endothelial cell survival but also regulates
vasomotor responses (37,38). Overexpression of PI3K/AKT has been
shown to induce angiogenesis and regulate tumor growth (39–41).
In the present study, caudatin treatment causes obvious inhibition
of AKT both in vitro and in vivo. PI3K inhibitor
further confirmed the AKT-dependent effect of caudatin-induced cell
killing.
In conclusion
In the present study, the antiangiogenic properties
and mechanism of caudatin in vitro and in vivo was
investigated, and the results suggested that caudatin had the
potential to inhibit HUVECs proliferation, migration, invasion and
capillary-like tube formation in vitro and in vivo
through suppression of the VEGF-VEGFR2-AKT/FAK signal axis. Our
findings validated the antiangiogenic potential of caudatin in the
tumor chemotherapy.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China grant no. 81571284 to G. Li.
Key Research and Development Program of Shandong no. 2016GSF202036
to CD. Fan.
References
|
1
|
Carmeliet P and Baes M: Metabolism and
therapeutic angiogenesis. N Engl J Med. 358:2511–2512. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Berz D and Wanebo H: Targeting the growth
factors and angiogenesis pathways: Small molecules in solid tumors.
J Surg Oncol. 103:574–586. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Burstein HJ and Schwartz RS: Molecular
origins of cancer. N Engl J Med. 358:5272008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kerbel RS: Tumor angiogenesis. N Engl J
Med. 358:2039–2049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ferrara N: Vascular endothelial growth
factor: Basic science and clinical progress. Endocr Rev.
25:581–611. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pyun BJ, Choi S, Lee Y, Kim TW, Min JK,
Kim Y, Kim BD, Kim JH, Kim TY, Kim YM and Kwon YG: Capsiate, a
nonpungent capsaicin-like compound, inhibits angiogenesis and
vascular permeability via a direct inhibition of Src kinase
activity. Cancer Res. 68:227–235. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fu XY, Zhang S, Wang K, Yang MF, Fan CD
and Sun BL: Caudatin inhibits human glioma cells growth through
triggering DNA damage-mediated cell cycle arrest. Cell Mol
Neurobiol. 35:953–959. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li X, Zhang M, Xiang C, Qin Y, He J, Li BC
and Li P: C21 steroids from roots of Cynanchum otophyllum. Zhongguo
Zhong Yao Za Zhi. 39:1450–1456. 2014.(In Chinese). PubMed/NCBI
|
|
10
|
Peng Y and Ding Y: Pharmacokinetics and
tissue distribution study of caudatin in normal and
diethylnitrosamine-induced hepatocellular carcinoma model rats.
Molecules. 20:4225–4237. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang YQ, Yang B, Zhang RS and Wei EQ:
Inhibitive effect of C-21 steroidal glycosides of Cynanchum
auriculatum on rat glioma cells in vitro. Zhejiang Da Xue Xue Bao
Yi Xue Ban. 40:402–407. 2011.(In Chinese). PubMed/NCBI
|
|
12
|
Wang YQ, Zhang SJ, Lu H, Yang B, Ye LF and
Zhang RS: A C 21 -steroidal glycoside isolated from the roots of
cynanchum auriculatum induces cell cycle arrest and apoptosis in
human gastric cancer SGC-7901 cells. Evid Based Complement Alternat
Med. 2013:1808392013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu LZ, Hou YJ, Zhao M, Yang MF, Fu XT,
Sun JY, Fu XY, Shao LR, Zhang HF, Fan CD, et al: Caudatin induces
caspase-dependent apoptosis in human glioma cells with involvement
of mitochondrial dysfunction and reactive oxygen species
generation. Cell Biol Toxicol. 32:333–345. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo Y, Sun Z, Li Y, Liu L, Cai X and Li Z:
Caudatin inhibits human hepatoma cell growth and metastasis through
modulation of the Wnt/β-catenin pathway. Oncol Rep. 30:2923–2928.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Peng YR, Ding YF, Wei YJ, Shu B, Li YB and
Liu XD: Caudatin-2,6-dideoxy-3-O-methy-β-D-cymaropyranoside 1
induced apoptosis through caspase 3-dependent pathway in human
hepatoma cell line SMMC7721. Phytother Res. 25:631–637. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fei HR, Chen HL, Xiao T, Chen G and Wang
FZ: Caudatin induces cell cycle arrest and caspase-dependent
apoptosis in HepG2 cell. Mol Biol Rep. 39:131–138. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang LJ, Chen H, Ma YB, Huang XY, Geng CA,
Zhang XM and Chen JJ: Design, synthesis and biological evaluation
of caudatin analogs as potent hepatitis B virus inhibitors. Med
Chem. 11:165–179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fei HR, Cui LY, Zhang ZR, Zhao Y and Wang
FZ: Caudatin inhibits carcinomic human alveolar basal epithelial
cell growth and angiogenesis through modulating GSK3β/β-catenin
pathway. J Cell Biochem. 113:3403–3410. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li X, Zhang X, Liu X, Tan Z, Yang C, Ding
X, Hu X, Zhou J, Xiang S, Zhou C and Zhang J: Caudatin induces cell
apoptosis in gastric cancer cells through modulation of
Wnt/β-catenin signaling. Oncol Rep. 30:677–684. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Aggarwal BB and Shishodia S: Molecular
targets of dietary agents for prevention and therapy of cancer.
Biochem Pharmacol. 71:1397–1421. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ren W, Qiao Z, Wang H, Zhu L and Zhang L:
Flavonoids: Promising anticancer agents. Med Res Rev. 23:519–534.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Carmeliet P, Dor Y, Herbert JM, Fukumura
D, Brusselmans K, Dewerchin M, Neeman M, Bono F, Abramovitch R,
Maxwell P, et al: Role of HIF-1alpha in hypoxia-mediated apoptosis,
cell proliferation and tumour angiogenesis. Nature. 394:485–490.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bolat F, Haberal N, Tunali N, Aslan E, Bal
N and Tuncer I: Expression of vascular endothelial growth factor
(VEGF), hypoxia inducible factor 1 alpha (HIF-1alpha), and
transforming growth factors beta1 (TGFbeta1) and beta3 (TGFbeta3)
in gestational trophoblastic disease. Pathol Res Pract. 206:19–23.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Carmeliet P: Angiogenesis in life, disease
and medicine. Nature. 438:932–936. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ferrara N and Kerbel RS: Angiogenesis as a
therapeutic target. Nature. 438:967–974. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Holopainen T, Bry M, Alitalo K and
Saaristo A: Perspectives on lymphangiogenesis and angiogenesis in
cancer. J Surg Oncol. 103:484–488. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nussenbaum F and Herman IM: Tumor
angiogenesis: Insights and innovations. J Oncol. 2010:1326412010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ferrara N: VEGF and the quest for tumour
angiogenesis factors. Nat Rev Cancer. 2:795–803. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Le Boeuf F, Houle F and Huot J: Regulation
of vascular endothelial growth factor receptor 2-mediated
phosphorylation of focal adhesion kinase by heat shock protein 90
and Src kinase activities. J Biol Chem. 279:39175–39185. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jośko J and Mazurek M: Transcription
factors having impact on vascular endothelial growth factor (VEGF)
gene expression in angiogenesis. Med Sci Monit. 10:RA89–RA98.
2004.PubMed/NCBI
|
|
31
|
Parsons JT: Focal adhesion kinase: The
first ten years. J Cell Sci. 116:1409–1416. 2013. View Article : Google Scholar
|
|
32
|
Braren R, Hu H, Kim YH, Beggs HE,
Reichardt LF and Wang R: Endothelial FAK is essential for vascular
network stability, cell survival, and lamellipodial formation. J
Cell Biol. 172:151–162. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Holmqvist K, Cross MJ, Rolny C, Hägerkvist
R, Rahimi N, Matsumoto T, Claesson-Welsh L and Welsh M: The adaptor
protein shb binds to tyrosine 1175 in vascular endothelial growth
factor (VEGF) receptor-2 and regulates VEGF-dependent cellular
migration. J Biol Chem. 279:22267–22275. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qi JH and Claesson-Welsh L: VEGF-induced
activation of phosphoinositide 3-kinase is dependent on focal
adhesion kinase. Exp Cell Res. 263:173–182. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mitra SK, Mikolon D, Molina JE, Hsia DA,
Hanson DA, Chi A, Lim ST, Bernard-Trifilo JA, Ilic D, Stupack DG,
et al: Intrinsic FAK activity and Y925 phosphorylation facilitate
an angiogenic switch in tumors. Oncogene. 25:5969–5984. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shen TL, Park AY, Alcaraz A, Peng X, Jang
I, Koni P, Flavell RA, Gu H and Guan JL: Conditional knockout of
focal adhesion kinase in endothelial cells reveals its role in
angiogenesis and vascular development in late embryogenesis. J Cell
Biol. 169:941–952. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shiojima I and Walsh K: Role of Akt
signaling in vascular homeostasis and angiogenesis. Circ Res.
90:1243–1250. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vara JA Fresno, Casado E, de Castro J,
Cejas P, Belda-Iniesta C and González-Barón M: PI3K/Akt signalling
pathway and cancer. Cancer Treat Rev. 30:193–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jiang BH, Zheng JZ, Aoki M and Vogt PK:
Phosphatidylinositol 3-kinase signaling mediates angiogenesis and
expression of vascular endothelial growth factor in endothelial
cells. Proc Natl Acad Sci USA. 97:pp. 1749–1753. 2000; View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fan C, Zheng W, Fu X, Li X, Wong YS and
Chen T: Enhancement of auranofin-induced lung cancer cell apoptosis
by selenocystine, a natural inhibitor of TrxR1 in vitro and in
vivo. Cell Death Dis. 5:e11912014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fan C, Zheng W, Fu X, Li X, Wong YS and
Chen T: Strategy to enhance the therapeutic effect of doxorubicin
in human hepatocellular carcinoma by selenocystine, a synergistic
agent that regulates the ROS-mediated signaling. Oncotarget.
5:2853–2563. 2014. View Article : Google Scholar : PubMed/NCBI
|