Introduction
In the world, hepatocellular carcinoma (HCC) is a
common type of tumor and the third most frequent to cause death
(1). Women aged 50 or older is the
high-risk population of HCC (2).
As so far, the most radical therapy of HCC is hepatectomy which can
improve the function of liver and patients' quality of life
immediately. And the other treatments such as orthotopic liver
transplantation (OLT), radiofrequency ablation (RFA) are effective
methods. However, the one of features of HCC is have no obvious
early symptoms which mean some patients missed the best time of
hepatectomy when they were diagnosed as HCC at first. So it is
important to search new therapies for getting more effective
curative effect. In recent years, the chemotherapy becomes the
viable option for the HCC treatment or post-hepatectomy. Sorafenib
and the multikinase inhibitor have shown effective activity against
HCC and has been approved in part of the world (2). However, the drug resistance hampers
the long-term effectiveness of these chemotherapeutic drugs.
Formerly, aspirin is a nonsteroidal
anti-inflammatory drug which uses in treatment of tumor
complication frequently. In recent years, aspirin was found against
cancer. The drug has been demonstrated that it can suppress cancer
growth and induce apoptosis and differentiation in some cancers
(3–7). Now, the concentration of aspirin is a
tissue in clinical application. Aspirin is associated with
potential side effects including gastrointestinal and
cardiovascular effects. There is a contradiction between the
demanded dose and the concentration of occurring adverse effects.
It is interest to investigate the union use effect from two drugs
together.
Previous studies showed that the different dose of
Aspirin can impact the postoperative survival in subsets of
post-esophagectomy patients (8).
Patients who had received a low-dose of aspirin daily for more than
5 years showed significant anti-cancer effect compared with
treatment without aspirin (9,10).
The clear mechanisms of antitumor of Aspirin are inhibition of the
cyclooxygenase (COX)-independent mechanisms and inhibition of
platelet. A high dose of aspirin can inhibit COX-2 expression which
be found abnormal high expression in various cancers (11–14).
It also inhibit the phosphatidylinositol 3-kinase-related pathways
(PIK3CA) pathway via reduce COX-2 expression to enhance the tumor
cells senescence in colorectal cancer and breast cancer (15). The other downstream transcriptional
targets interact with the targeted molecules of aspirin including
ERK, MAPK and NF-κB and so on. Aspirin may induce COX-2 expression
through activate inactivation of platelets in epithelial cells of
colorectal adenomas and cancers (16). In addition, it is gradually
accepted that aspirin can acetylate lysine residues in serum
proteins. A report in human breast cancer cells showed Aspirin
induced p21CIP1 protein levels in a transient fashion
via acetylating the p53 protein markedly.
Nutlin-3 is an MDM2 antagonist which can protect P53
from binding with MDM2 and activate P53 (17). There are a number of studies
involving the mechanism of nutlin-3 function for a various of
cancers in which nutlin-3 can induce growth inhibition, cell
migration and/or apoptosis (18–23).
Nutlin-3 can bind with the three hydrophobic amino acid (-Leu26,
Trp23, Phe19) of P53 specifically and efficiently. Then it breaks
the combination of P53 and MDM2 and gather p53 again to exert
antitumor effect including the arrest of tumor cell cycle and the
cell apoptosis. Nutlin-3 has been used as a probe compound in
preclinical (24). However,
because of the dose-dependent toxicities and drug-resistance,
Nutlin-3 not be used to cure disease clinically. There is still a
long way to use Nutlin-3 in clinical extensively.
Although the intrinsic mechanisms of aspirin on COX
and platelet have been well-studied, the certified pathways of
Aspirin in P53 network is little. Herein, we hypothesized that the
antitumor effects of aspirin may be enhanced by nutlin-3 in HCC
cells. Its mechanism involve the P53-dependent pathway. So the aim
of the present study was to evaluate the antitumor effect of
combined treatment of aspirin and nutlin-3 in human HCC cell
lines.
However, whether aspirin combined with nutlin-3 has
a synergistic effect on HCC cell lines' proliferation and apoptosis
is still unknown. Based on these findings, in the present study, we
aim to evaluate the synergistic antitumor effect of aspirin and
nutlin-3 combination in human HCC cell lines.
Materials and methods
Cell culture
Human HCC line (HepG2) was obtained from Shanghai
Institute of Biochemistry and Cell Biology, Chinese Academy of
Sciences (Shanghai, China). HepG2 cells were cultured in DMEM
culture medium (HyClone; GE Healthcare Life Sciences, Logan, UT,
USA) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), 100 U/ml penicillin and 100 U/ml
streptomycin. The phenomena of culturing HepG2 cells is a
humidified atmosphere containing 5% carbon dioxide at 37°C.
Drugs and reagents
Aspirin and Nutlin-3 were purchased by Sigma-Aldrich
(Merck KGaA, Darmstadt, Germany). Nutlin-3 was dissolved in
dimethyl sulfoxide (20 mg/ml) and diluted with H2O.
Aspirin was dissolved in dimethyl sulfoxide (20 mg/ml) and diluted
with PBS medium. The final concentration of Aspirin and Nutlin-3 is
1 mmol/l and 5 µmol/l, respectively.
MTT assay for cell viability
HepG2 cells were seeded into 96-well plates at a
density of 5×104 cells per well, and incubated for 24 h
to make sure all cells attached to the plate sufficiently. Then we
would starve the cells by change culture medium without FBS for 12
h. The cells were incubated with aspirin and/or nutlin-3 at varying
doses of single drug or its combination for 24, 48 or 72 h. After
treatments, supernatants were discarded and washed with PBS. The
cells were incubated with MTT (5 mg/ml in PBS) at 37°C for another
4 h. Discard the medium, MTT formazan crystals were dissolved in
200 µl DMSO. In the end, it was measured using a POLAR star OPTIMA
microplate reader (BMG Labtechnologies, Ortenberg, Germany) at
absorbance at 570 nm. Cell viability is equal with the percentage
of viable cells in total cell population. Each experiment was
performed three replicates.
Colony formation assay
One thousand cells were plated into 6-well plates
and incubated in DMEM with 10% FBS at 37°C. Seven days later, the
cells were fixed and stained with 0.1% crystal violet. The number
of colonies, defined as >50 cells/colony, were counted.
Cell migration assays
The invasive potential of cells was measured in 6.5
micrometers Transwell with 8.0 micrometers Pore Polycarbonate
Membrane Insert (Corning Incorporated, Corning, NY, USA) according
to the manufacturer's instructions. Then HepG2 cells were seeded
into the upper chamber with DMEM. The lower chambers were filled
with 600 µl of DMEM medium with 10% FBS as chemoattractant. After
48 h, the invasive cells across membrane and adhere to the lower
surface of the membrane which were fixed with 4% paraformaldehyde
for 30 min, permeabilized with 0.2% Triton X-100 at room
temperature for 15 min, and then stained with 0.1% crystal violet
for 5 min. Light microscope was used to measure the number of
invading cells in five random fields at a magnification of
×100.
Apoptosis assay
Annexin V: PE Apoptosis Detection kit (BD
Biosciences, Franklin Lakes, NJ, USA) was used to measure the
cells' apoptosis induced by nutlin-3 or/and Aspirin. After
treatment for 48 h, cells were digested by pancreatin with EDTA and
washed with PBS at 4°C. Then it were resuspended in 500 µl binding
buffer (1×106 cells per ml) containing 5 µl of Annexin
V-APC and 5 µl of 7-AAD. The cells were incubated for 15 min at
room temperature without light and analyzed by flow cytometry (FACS
Calibur; BD Biosciences). Apoptotic cells were analyzed by quadrant
statistics of the propidium iodide-negative and Annexin V-positive
cells. The apoptotic index was calculated by the percentage of the
propidium iodide-negative and Annexin V-positive cells in total
cells.
Protein extraction and western
blotting
HepG2 cells were cultured by treating with nutlin-3
and/or aspirin for 48 h in 6-well plates. Then it was digested by
pancreatin with EDTA to collect total cells. The cells were
sonicated and centrifuged to collect cell extracts at 12,000 × g
for 10 min at 4°C. Protein concentrations were determined with a
BCA assay. Equal amounts of total proteins were transferred to
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA). After blocking in 5% non-fat dry milk, the membrane was
subsequently immunoblotted with primary antibodies following:
Anti-β-actin, anti-B-cell lymphoma 2 (Bcl-2), anti-Bax,
anti-caspase-3, anti-caspase-8 and anti-caspase-9 overnight at 4°C.
Next day, immunoreactivity was incubated with HRP conjugated
anti-goat or anti-rabbit antibody (Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA). The signals were visualized by the ECL kit
(Pierce; Thermo Fisher Scientific, Inc.). The primary antibodies
were all purchased from Santa Cruz Biotechnology, Inc..
Xenograft mouse model
BALB/c nude mice (5 weeks old, 20–25 g) were
purchased from Animal Feeding Center of Xi'an Jiaotong University
Health Science Center. Animal experimental protocols was approved
by the Institutional Animal Care and Use Committee, and the mice
were fed in compliance with criteria outlined in the Guide for the
Care and Use of Laboratory Animals established by the US National
Institutes of Health, strictly. HepG2 cells were harvested and
re-suspended sterile, serum-free PBS. The mice were randomized into
four groups (N=4 per group). Then 5×106 HepG2 cells were
inoculated into each mice on posterior hind flank region
subcutaneously. After the injection of HepG2 cells on 3 days, the
treatment of Aspirin (13 mg/kg/day, p.o.) and/or Nutlin-3 (200
mg/kg, twice a day, p.o.) was started after the injection of HepG2
cells (25). As a controls group,
the mice were administrated by PBS (0.2 ml/kg/day). 3 weeks after
daily administration, mice were killed. The tumor was then removed
and underwent the immunohistochemistry. Tumor growth was monitored
twice weekly. The length (L) and the width (W) of the tumors were
measured and the volume (V) of each tumor was calculated with the
formula: V=Lx W2/2 (26). The study was approved by the Animal
Research Committee of Xi'an Jiaotong University Health Science
Center.
Statistical analysis
Experiments were performed with values expressed as
mean ± standard deviation. All statistical analyses were conducted
using the Graphpad Prism 6.0 software. The correlation differences
between the respective groups were evaluated by either ANOVA or
nonparametric test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Aspirin and nutlin-3 inhibits the
proliferation and migration of HepG2 cells
As we all know, nutlin-3 inhibits the proliferation
in HCC cells which were confirmed by previous studies (27,28).
To study the function of aspirin in anti-proliferative and the
effect of cooperating with nutlin-3, aspirin and nutlin-3 were
detected by concentration gradient in HepG2 cell line separately.
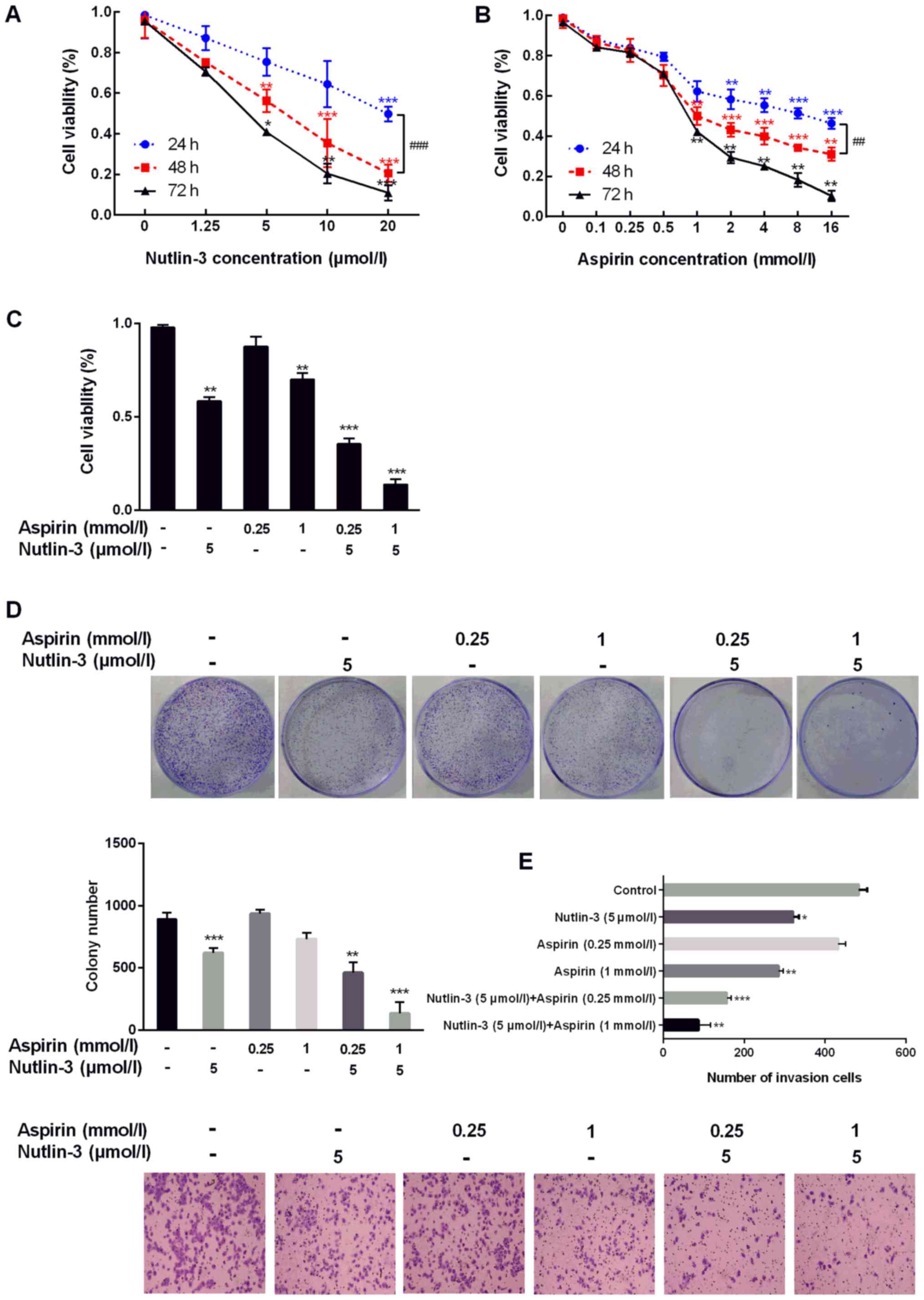
Then the MTT assay were performed for 24, 48, 72 h. We can easily
found that both aspirin (Fig. 1A)
and nutlin-3 (Fig. 1B) would
inhibit the growth of viable HCC cells in a dose-dependent manner
significantly. After 48 h treatment for aspirin or nutlin-3, the
anti-proliferation effect of the two drugs showed statistical
significance. Based on the IC50 dose, the experimental
concentration of nutlin-3 in HepG2 cells was determined as 5 µmol/l
at 48 h. The experimental concentration of aspirin was determined
as 1 µmol/l at 48 h. Moreover, the low dose of aspirin identified
as 0.25 mmol/l were used to investigate the side effect.
Next, aspirin (1 or 0.25 mmol/l) and nutlin-3 (5
µmol/l) were used in HepG2 cells simultaneously for 48 h, the MTT
assay showed a remarkable decrease in HepG2 viable cell numbers and
the effect of growth inhibitory is remarkly stronger compared with
control or drug treatment alone (Fig.
1C). Colony formation assays revealed that aspirin (1 mmol/l)
and nutlin-3 (5 µmol/l) formed significantly less colonies than
control in HepG2 cells. While the low dose of aspirin (0.25 mmol/l)
formed a similar colonies as control. When combined aspirin and
nutlin-3, the colonies of HepG2 cells is higher than drug
administration alone and control (Fig.
1D). Furthermore, the high aggressive potency is one of the key
feature of malignant cells. The migration assays were performed to
evaluate the migration capabilities of HepG2 cells treated with
aspirin and nutlin-3. Co-treatment of aspirin (l mmol/l) and
nutlin-3 (5 µmol/l) suppressed cells' migration (Fig. 1E) compared with control or drug
treatment alone.
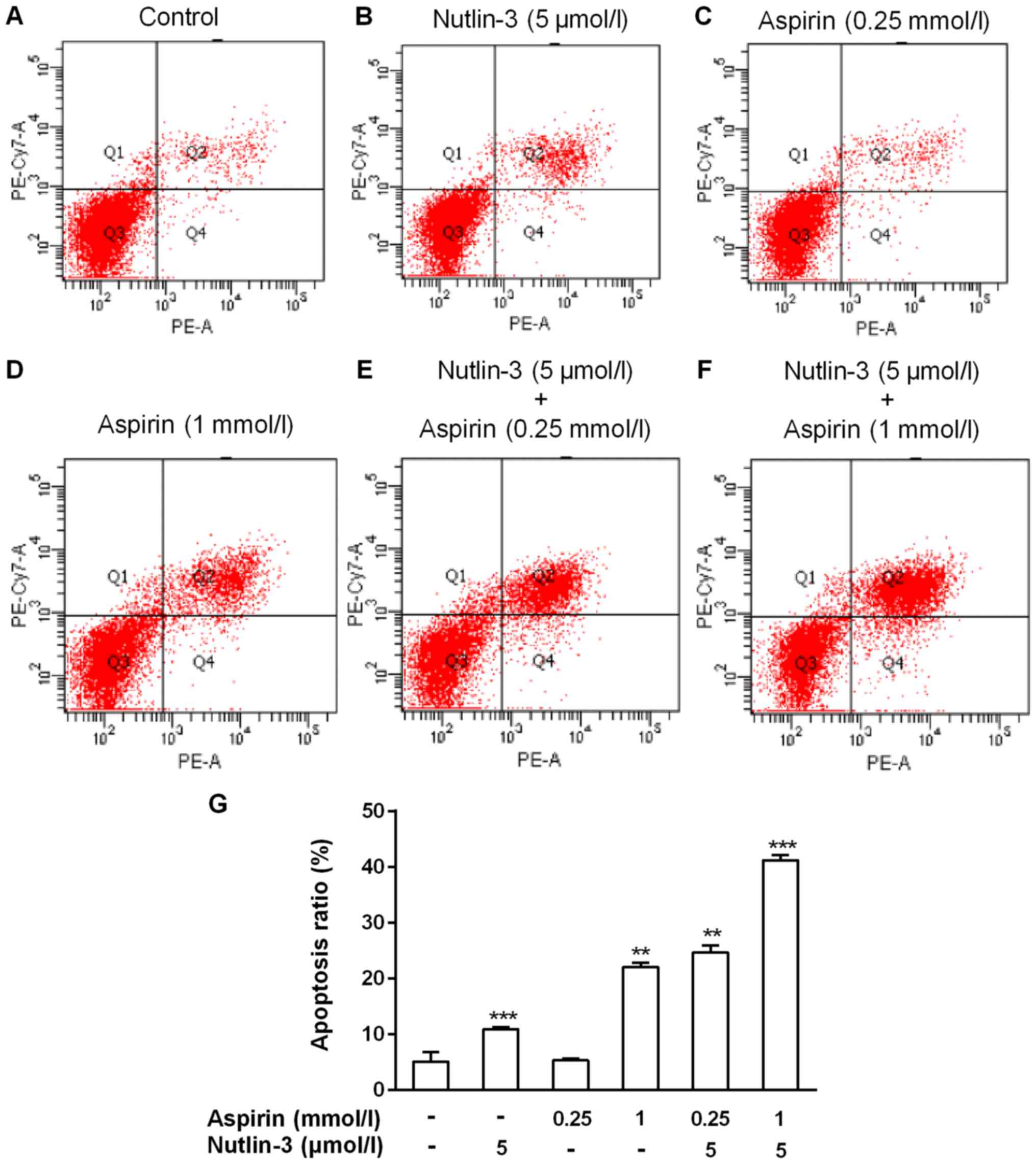
Aspirin enhances nutlin-3-induced
apoptosis of HCC cell
Aspirin can induce tumor cells apoptosis by signal
pathways. We speculated that aspirin and nutlin-3 can regulate the
tumor cells proliferation via affects apoptosis in HepG2 cells. To
study the potential synergistic effect of apoptosis for aspirin and
nutlin-3, HepG2 cells were treated for 48 h in six groups. Then
Annexin V-APC/7-AAD Apoptosis Detection kit were used to stain the
apoptotic cells in six groups respectively. Compared with control
group, the two drugs are all increase the apoptosis ratio of HepG2
cells, low dose of aspirin showed no statistical difference. But
compared with either aspirin or nutlin-3 alone, the combination of
aspirin (1 or 0.25 mmol/l) and nutlin-3 (5 µmol/l) both induced a
significant increase in HepG2 cells (Fig. 2). These results suggested that
nutlin-3 can enhance the apoptotic effect of aspirin and nutlin-3
can enhance the apoptot ic effect of a low dose of aspirin in HCC
HepG2 cells.
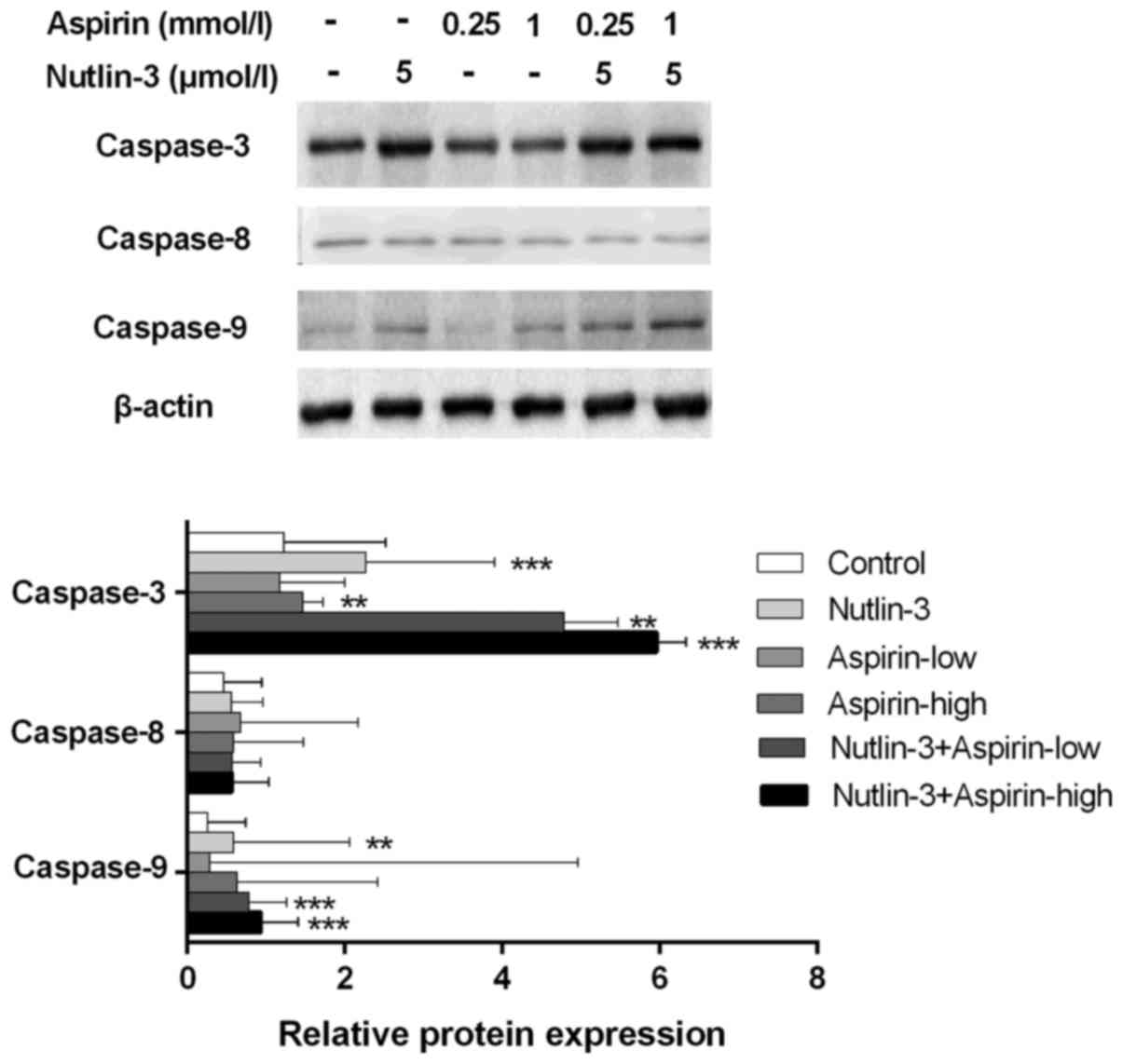
Combination of aspirin and nutlin-3
enhances apoptosis via caspase cascade proteins
In previous combination treatment of nutlin-3 and
aspirin, significant decline of cell numbers was observed. To
confirm the mechanism induced apoptosis by the co-treatment, we
extract the total protein from HepG2 cells treated with the two
drugs alone or in combination respectively to detect the change of
apoptotic protein biomarkers. The results showed that the
administration of nutlin-3 can increase the levels of caspase-3 and
caspase-9. While there is no change on caspase-8 in HepG2 cells
treated with nutlin-3. A low dose of aspirin (0.25 mmol/l) did not
change the level of caspase-9 and caspase-3. However, the result of
the combined treatment is the expression of caspase-9 and caspase-3
go up which was significantly obvious. The expression of caspase-8
showed no change when treated with nutlin-3 and Aspirin alone or in
combination (Fig. 3).
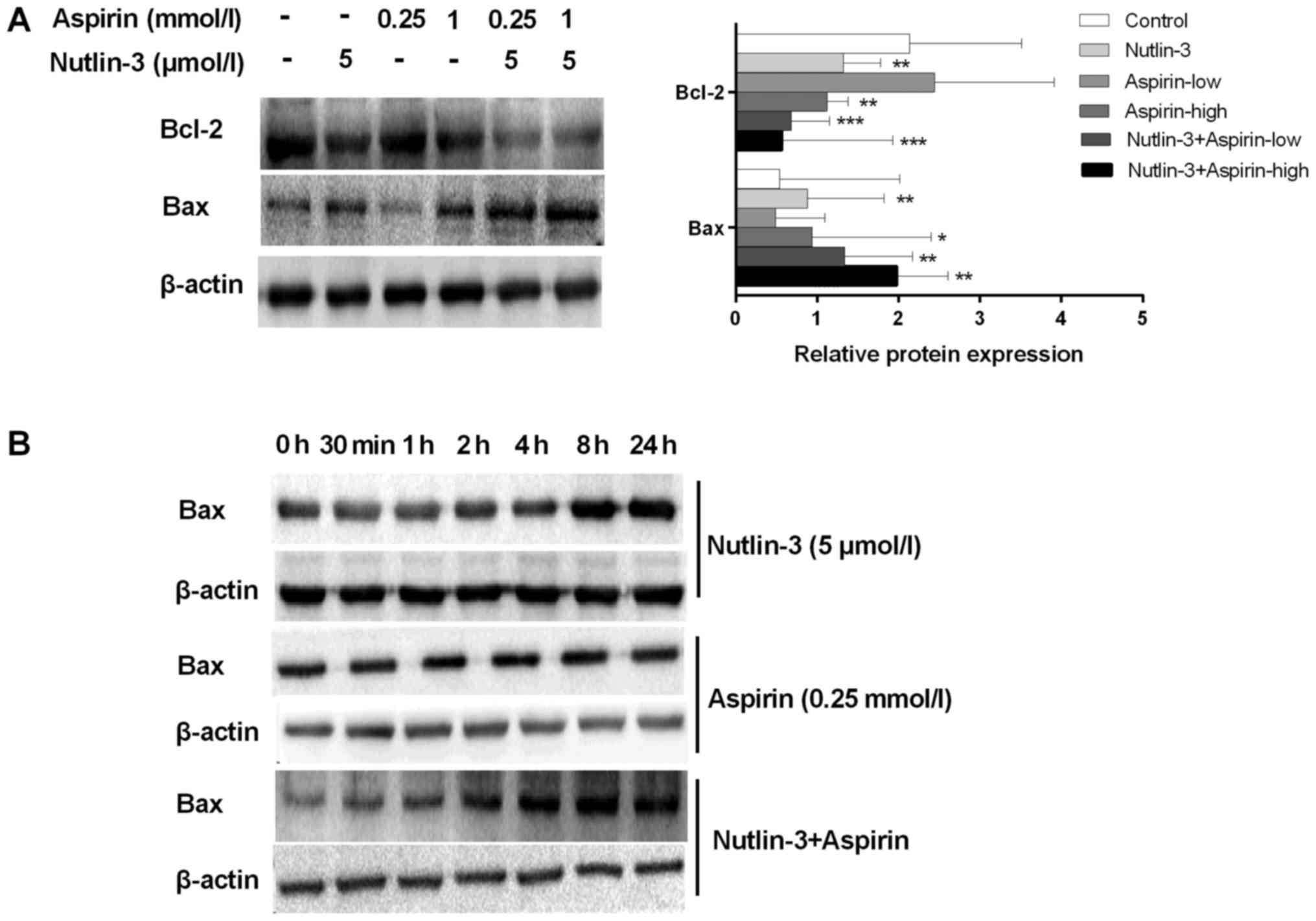
Nutlin-3 activates the Bax/Bcl-2
signaling pathway in HepG2 cells treated with A low dose of
aspirin
As we all know, Bcl-2/Bax is a specific signaling
pathways downstream of P53, and it is a downstream of caspase-3. We
wonder to know if Bcl-2/Bax pathway were involved in the
synergistic effect of nutlin-3 combined with Aspirin. The western
blot results found that nutlin-3 can increase the level of Bax and
decrease the level of Bcl-2 in HepG2 cells. The expression of Bax
was no change by aspirin (0.25 mmol/l) and enhanced by the high
dose of aspirin. In addition, the expression of Bcl-2 have no
change for aspirin. Furthermore, when treated with aspirin (0.25
mmol/l) and nutlin-3 (5 µmol/l), tumor cells showed high expression
of Bax levels than when treated with nutlin-3 or aspirin alone
(Fig. 4A). So we suggested that
nutlin-3 enhanced the apoptotic effect of aspirin through Bcl-2/Bax
signal pathway.
Next, we choose a low dose of aspirin (0.25 mmol/l)
to verify the mechanism. Nutlin-3 (5 µmol/l) and aspirin (0.25
mmol/l) were used for detecting the change of Bax at various time
points in HepG2 cells. After treated with nutlin-3 for 8 h in HepG2
cells, the total Bax expression was increased significantly in a
time-dependent manner (Fig. 4B),
but the group treated with aspirin at a low concentration have no
change. Furthermore, when nutlin-3 (5 µmol/l) and aspirin (0.25
mmol/l) were used together, the expression of Bax was significantly
increased beginning after treated with two drugs for 2 h.
Combination of nutlin-3 and aspirin
reduces proliferation and tumor angiogenesis of HCC cells in
vivo
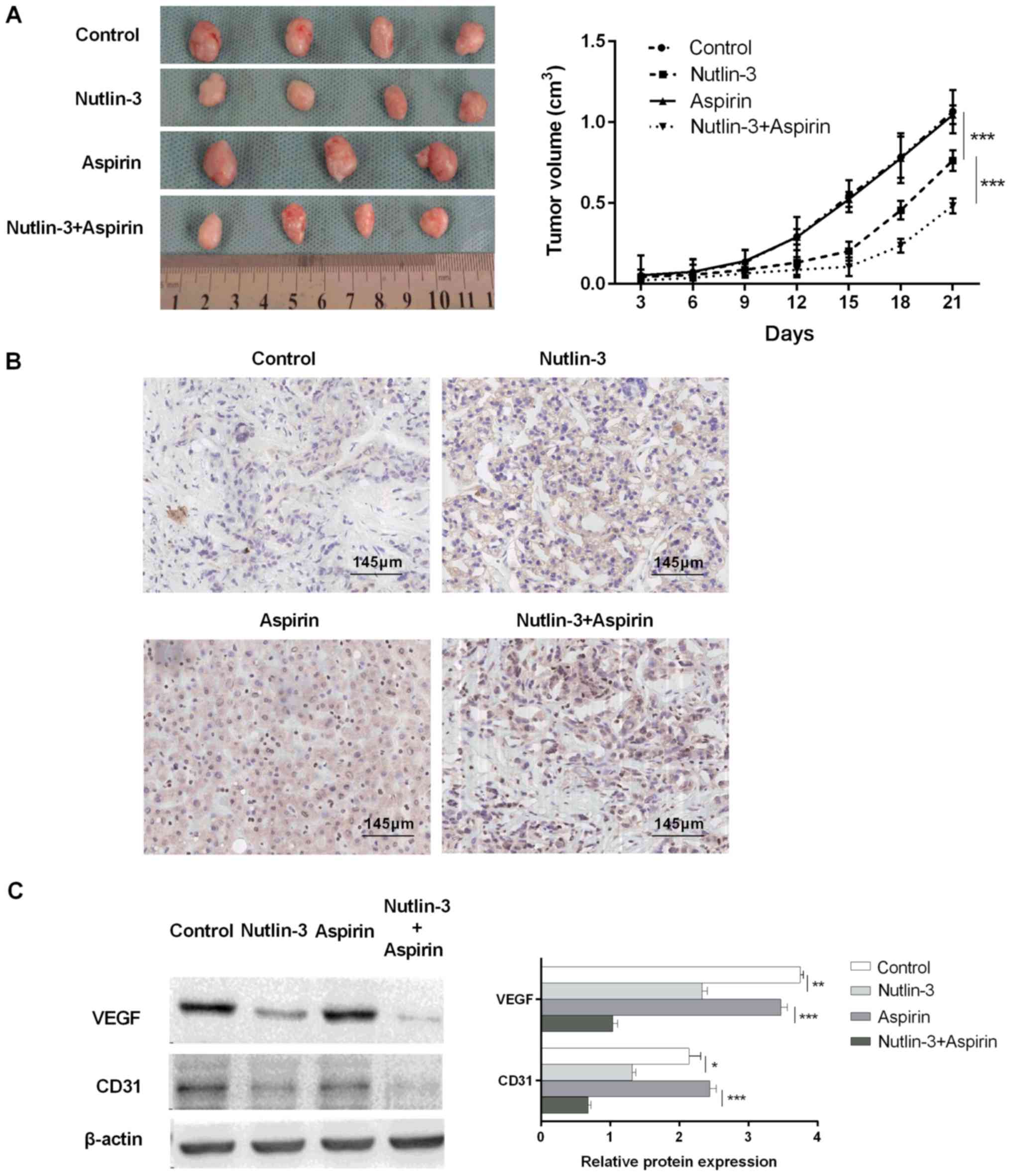
In order to verify whether the combination of
aspirin and nutlin-3 could inhibit tumor growth, the in vivo
activity of combination on HCC cells were also evaluated.
Subcutaneous inoculation of HepG2 cells into nude mice resulted in
a tumor formation at the site of injection in all mice. There are
four group in which mice administrated with PBS (0.2 ml/kg/day), a
low dose of aspirin (13 mg/kg/day, p.o.), nutlin-3 (200 mg/kg,
twice a day, p.o.) and a low dose of aspirin (13 mg/kg/day, p.o.)
combined with nutlin-3 (200 mg/kg, twice a day, p.o.),
respectively. One mice in the group treated with aspirin alone died
during the experiment because of gastric mucosa injury. But all the
mice from other groups survived until the experiment stopped. The
tumor growth ability of HepG2 cells in the combination group is
weakest among four groups. And no significant difference was
observed in tumor volume of control group and aspirin management
group (Fig. 5A). The Bax
immuno-histochemical reactions were performed in the tumor tissue
(Fig. 5B). Bax was markedly
expressed in group treated with aspirin and nutlin-3. Next, the
angiogenesis-associated proteins VEGF, and CD31 were detected in
the four group. VEGF, and CD31 protein expressions were decreased
in the group in which mice treated with nutlin-3. The expression of
VEGF, and CD31 is lowest in the four groups (Fig. 5C). Therefore, nutlin-3 cooperated
with aspirin might suppress the proliferation ability of tumor
cells and inhibit tumor angiogenesis via up-regulating the
expression of Bax in vivo. Moreover, nutlin-3 can attenuate
the toxicity in aspirin treatment.
Discussion
In 1979, the HepG2 cell line was firstly established
by Barbara Knowles and colleagues, and reported as a HCC (29). Over the next few decades, HepG2 has
been widely used to investigate metabolism, development,
oncogenesis (chemocarcinogenesis and mutagenesis), and
hepatotoxicity in HCC. However, in 2009, López-Terrada et al
reported HepG2 originated as a hepatoblastoma (HB) and not a HCC
according to array comparative genomic hybridization (CGH) analysis
and a series of experiments (30).
HB is a primary hepatic malignant tumor and originated from
primitive hepatic stem cells. It commonly occurred in children.
Now, HepG2 cell lines were used to explore molecular mechanism in
HB. Even so, more than 2012 researches used HepG2 to investigate
HCC from 2009 to 2017 according to PubMed. In the study, the
synergistic anti-cancer effects of aspirin and nutlin-3 have been
conformed in HepG2. There were even some hints the synergistic
anticancer effects of aspirin and nutlin-3 are exist not only in
liver cancer but also in hepatoblastoma. However, the general
applicability in other liver cancer type is still unknown.
Aspirin is a common clinic drugs which was approved
to control postoperative pain, inflammation and prevent
cardiovascular disease. Recently, a large number of researches and
epidemiological studies has demonstrated that the use of
nonsteroidal anti-inflammatory drugs (NSAIDs) protects against the
incidence and development of certain cancers. In our study, aspirin
can inhibit the proliferation and invasion of HepG2 cell line with
a dose-manner administration. The animal experiment has also
clarified the phenomena in vivo such as the smaller tumor
volume in treatment group with aspirin. In actually, Aspirin is the
most popular of being investigated in different types of cancers
(31–33). It has been used in chemoprevention
of many malignant cancers. And effective taking aspirin need to be
dose and time dependent. However, patients on high-dose aspirin
(≥500 mg daily) might induce bleeding complications which limit the
clinical application value of aspirin.
Drug-drug interactions (DDIs) is a new method to
deal with the problem of adverse drug events. The drug positively
or negatively impact the clinical effect or plasma concentration of
another drug even be a significant cause of morbidity and mortality
worldwide. Aspirin inhibit the prevention, progression, and
metastatic growth of cancer by the inhibition of COX-2 and platelet
aggregation (34–36). The expression of COX-2 is
associated with increased cell proliferation and tumor promotion
significantly. In addition, aspirin can promote platelet
aggregation which could contribute to immune evasion. And platelets
could induce abnormal COX-2 levels which can decrease the
oncosuppressor genes and decrease the oncogenes in colon carcinoma
(37). Moreover, some COX-2
independent pathways have been investigated to elucidate aspirin
mechanism including NF-κB, ERK, Bcl-2/Bax signaling pathway,
TGF-β/Smads signaling pathway, JAK1/STAT1 pathway, PI3K/Akt
signaling pathway and vascular endothelial growth factor (VEGF)
signaling (11,38–41).
Due to the mechanism is related with cells angiogenesis, cell
proliferation and migration, especially apoptosis, we wondered to
uncover a new drug to cooperate with aspirin for inducing tumor
cells' apoptosis. P53 is the classical anti-apoptosis gene and a
key protein in regulating tumor cells' apoptosis. Previous study
showed that inhibition of P53 suppression by MDM2 in cancer cells
promotes growth arrest, apoptosis and senescence in vitro
and in vivo. Nutlin-3 has been demonstrated that induces
cell cycle arrest and apoptosis by downregulates p53
phosphorylation on serine392 in HCC cells (27). In consist with previous researches,
in this study, a similar growth inhibition and apoptosis effect of
nutlin-3 were found in HepG2 cell lines. Moreover, combination
between nutlin-3 and aspirin not only reduced the proliferation of
HepG2 cells but also enhanced its apoptosis significantly. The
western blot result showed an outstanding upregulation of caspase-3
and caspase-9 in co-treatment group of nutlin-3 and aspirin
compared with other groups treated with nutlin-3 and aspirin alone.
These two proteinase paly essential role for being blocked the
recruitment of procadpase-9 to the Apaf-1/dATP/cytochrome c
apoptosome complex (42). As we
all know, Bax promotes the release of cytochrome c, and activates
caspase-9 and caspase-3. Bcl-2/Bax is directly regulated by the
tumor suppressor P53 and has been shown to be involved in
P53-mediated apoptosis (43,44).
We suggested that the synergistic effect between nutlin-3 and
aspirin exert apoptosis function by Bcl-2/Bax signaling pathway. In
consist with our assumption, the expression of Bax was raised
according different time continually. Therefore, these data
demonstrate that in HepG2 cell Bax is associated with the
apoptosis.
The applicable concentration of drug is a key issue
in clinic. Although aspirin has been a common therapy in clinic for
anti-inflammation and improving cardiovascular disease, the value
of preventing and controlling tumor is limited by its adverse drug
events. To make sure the effective clinical function, the blood
medicinal concentration of aspirin is 1–3 mmol/l. And a report
proved the safe concentration of aspirin is 30–1,500 mg/day
(45). However, approximately 90%
of the liver cancer patients have liver cirrhosis which show liver
dysfunction in different levels and portal hypertension. Using of
high-dose aspirin would induce gastrointestinal bleeding. But if
the dose lass than 100 mg daily, it would occur the risk of
bleeding. The application value of low dose aspirin has been
evidenced in some cancer (46,47).
There is a study indicated the low dose aspirin (300–75 mg daily)
also can reduce colorectal cancer incidence and mortality,
especially proximal colon cancer by a 20 years follow-up of five
randomized trials (47). In our
animal model, nude mouse were chosen to build model of
transplantable subcutaneously tumors in mouse. Because the nude
mice's capability of adapting themselves to the drug administration
was weaker than the normal mice. We determined the concentration of
aspirin as 15 mg/kg/day, p.o. according previous reports (40,48).
However, mice all died after treating with aspirin in succession.
Therefore, we lowered drug concentration (13 mg/kg/day, p.o.) to
fit experiment demands. The tumor volume for combination therapy of
aspirin and nutlin-3 is decrease compared with other groups and no
side effects were observed. It indicated that the combinations of
low dose aspirin and nutlin-3 regularly has no risk of bleeding and
assure the effective drug concentration in HCC. We can assume that
a study is feasible to determine an optimal concentration to
improve the efficacy of aspirin in clinic.
In conclusion, our present results indicated that
the treatment of aspirin and nutlin-3 alone or in combination both
inhibit growth and promote apoptosis in vitro and in
vivo. Nutlin-3 enhances the apoptosis effect of a low dose of
aspirin by upregulating Bax expression in HepG2 cell line and in
vivo. The synergistic effect of nutlin-3 in aspirin antitumor
therapy contribute to diminish the dose of aspirin and decrease the
rise of adverse drug events which would expand aspirin therapeutic
and/or adjuvant therapeutic application in the treatment of HCC in
clinical.
References
|
1
|
Gish RG: Hepatocellular carcinoma:
Overcoming challenges in disease management. Clin Gastroenterol
Hepatol. 4:252–261. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tsoulfas G, Agorastou P, Tooulias A and
Marakis GN: Current and future challenges in the surgical treatment
of hepatocellular carcinoma: A review. Int Surg. 99:779–786. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chattopadhyay M, Kodela R, Nath N,
Barsegian A, Boring D and Kashfi K: Hydrogen sulfide-releasing
aspirin suppresses NF-κB signaling in estrogen receptor negative
breast cancer cells in vitro and in vivo. Biochem Pharmacol.
83:723–732. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yiannakopoulou ECh: Aspirin and NSAIDs for
breast cancer chemoprevention. Eur J Cancer Prev. 24:416–421. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang X, Diao Y, Liu Y, Gao N, Gao D, Wan
Y, Zhong J and Jin G: Synergistic apoptosis-inducing effect of
aspirin and isosorbide mononitrate on human colon cancer cells. Mol
Med Rep. 12:4750–4758. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xiang S, Sun Z, He Q, Yan F, Wang Y and
Zhang J: Aspirin inhibits ErbB2 to induce apoptosis in cervical
cancer cells. Med Oncol. 27:379–387. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shen X, Han L, Ma Z, Chen C, Duan W, Yu S,
Li P, Zhang L, Li W, Xu Q and Ma Q: Aspirin: A potential
therapeutic approach in pancreatic cancer. Curr Med Chem.
20:4153–4162. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu JF, Jamieson GG, Wu TC, Zhu GJ and
Drew PA: A preliminary study on the postoperative survival of
patients given aspirin after resection for squamous cell carcinoma
of the esophagus or adenocarcinoma of the cardia. Ann Surg Oncol.
16:1397–1402. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mc Menamin ÚC, Cardwell CR, Hughes CM and
Murray LJ: Low-dose aspirin use and survival in breast cancer
patients: A nationwide cohort study. Cancer Epidemiol. 47:20–27.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Frouws MA, Bastiaannet E, Langley RE, Chia
WK, van Herk-Sukel MP, Lemmens VE, Putter H, Hartgrink HH, Bonsing
BA, Van de Velde CJ, et al: Effect of low-dose aspirin use on
survival of patients with gastrointestinal malignancies; an
observational study. Br J Cancer. 116:405–413. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kastrati I, Litosh VA, Zhao S, Alvarez M,
Thatcher GR and Frasor J: A novel aspirin prodrug inhibits NFκB
activity and breast cancer stem cell properties. BMC Cancer.
15:8452015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rao CV, Reddy BS, Steele VE, Wang CX, Liu
X, Ouyang N, Patlolla JM, Simi B, Kopelovich L and Rigas B: Nitric
oxide-releasing aspirin and indomethacin are potent inhibitors
against colon cancer in azoxymethane-treated rats: Effects on
molecular targets. Mol Cancer Ther. 5:1530–1538. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bhat IA, Rasool R, Qasim I, Masoodi KZ,
Paul SA, Bhat BA, Ganaie FA, Aziz SA and Shah ZA: COX-2
overexpression and −8473 T/C polymorphism in 3′UTR in non-small
cell lung cancer. Tumour Biol. 35:11209–11218. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Buskens CJ, Ristimäki A, Offerhaus GJ,
Richel DJ and van Lanschot JJ: Role of cyclooxygenase-2 in the
development and treatment of oesophageal adenocarcinoma. Scand J
Gastroenterol Suppl. 1–93. 2003.PubMed/NCBI
|
|
15
|
Singh Ranger G: The role of aspirin in
colorectal cancer chemoprevention. Crit Rev Oncol Hematol.
104:87–90. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sciulli MG, Filabozzi P, Tacconelli S,
Padovano R, Ricciotti E, Capone ML, Grana M, Carnevale V and
Patrignani P: Platelet activation in patients with colorectal
cancer. Prostaglandins Leukot Essent Fatty Acids. 72:79–83. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Polanski R, Noon AP, Blaydes J, Phillips
A, Rubbi CP, Parsons K, Vlatković N and Boyd MT: Senescence
induction in renal carcinoma cells by Nutlin-3: A potential
therapeutic strategy based on MDM2 antagonism. Cancer Lett.
353:211–219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shangary S and Wang S: Small-molecule
inhibitors of the MDM2-p53 protein-protein interaction to
reactivate p53 function: A novel approach for cancer therapy. Annu
Rev Pharmacol Toxicol. 49:223–241. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang P, Chen W, Li X, Eilers G, He Q, Liu
L, Wu Y, Wu Y, Yu W, Fletcher JA and Ou WB: Downregulation of
cyclin D1 sensitizes cancer cells to MDM2 antagonist Nutlin-3.
Oncotarget. 7:32652–32663. 2016.PubMed/NCBI
|
|
20
|
Pechackova S, Burdova K, Benada J,
Kleiblova P, Jenikova G and Macurek L: Inhibition of WIP1
phosphatase sensitizes breast cancer cells to genotoxic stress and
to MDM2 antagonist nutlin-3. Oncotarget. 7:14458–14475. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ghassemifar S and Mendrysa SM: MDM2
antagonism by nutlin-3 induces death in human medulloblastoma
cells. Neurosci Lett. 513:106–110. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Secchiero P, Bosco R, Celeghini C and
Zauli G: Recent advances in the therapeutic perspectives of
Nutlin-3. Curr Pharm Des. 17:569–577. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Endo S, Yamato K, Hirai S, Moriwaki T,
Fukuda K, Suzuki H, Abei M, Nakagawa I and Hyodo I: Potent in vitro
and in vivo antitumor effects of MDM2 inhibitor nutlin-3 in gastric
cancer cells. Cancer Sci. 102:605–613. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zanjirband M, Edmondson RJ and Lunec J:
Pre-clinical efficacy and synergistic potential of the MDM2-p53
antagonists, Nutlin-3 and RG7388, as single agents and in combined
treatment with cisplatin in ovarian cancer. Oncotarget.
7:40115–40134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zheng T, Yin D, Lu Z, Wang J, Li Y, Chen
X, Liang Y, Song X, Qi S, Sun B, et al: Nutlin-3 overcomes arsenic
trioxide resistance and tumor metastasis mediated by mutant p53 in
hepatocellular carcinoma. Mol Cancer. 13:1332014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shao QS, Ye ZY, Ling ZQ and Ke JJ: Cell
cycle arrest and apoptotic cell death in cultured human gastric
carcinoma cells mediated by arsenic trioxide. World J
Gastroenterol. 11:3451–3456. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shi X, Liu J, Ren L, Mao N, Tan F, Ding N,
Yang J and Li M: Nutlin-3 downregulates p53 phosphorylation on
serine392 and induces apoptosis in hepatocellular carcinoma cells.
BMB Rep. 47:221–226. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang J, Zheng T, Chen X, Song X, Meng X,
Bhatta N, Pan S, Jiang H and Liu L: MDM2 antagonist can inhibit
tumor growth in hepatocellular carcinoma with different types of
p53 in vitro. J Gastroenterol Hepatol. 26:371–377. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Aden DP, Fogel A, Plotkin S, Damjanov I
and Knowles BB: Controlled synthesis of HBsAg in a differentiated
human liver carcinoma-derived cell line. Nature. 282:615–616. 1979.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep G2 is a hepatoblastoma-derived cell line. Human
Pathol. 40:1512–1515. 2009. View Article : Google Scholar
|
|
31
|
Tougeron D, Sha D, Manthravadi S and
Sinicrope FA: Aspirin and colorectal cancer: Back to the future.
Clin Cancer Res. 20:1087–1094. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Maity G, De A, Das A, Banerjee S, Sarkar S
and Banerjee SK: Aspirin blocks growth of breast tumor cells and
tumor-initiating cells and induces reprogramming factors of
mesenchymal to epithelial transition. Lab Invest. 95:702–717. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Y, Shen C, Ge J and Duan H: Regular
aspirin use and stomach cancer risk in China. Eur J Surg Oncol.
41:801–804. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lin HY, Delmas D, Vang O, Hsieh TC, Lin S,
Cheng GY, Chiang HL, Chen CE, Tang HY, Crawford DR, et al:
Mechanisms of ceramide-induced COX-2-dependent apoptosis in human
ovarian cancer OVCAR-3 cells partially overlapped with resveratrol.
J Cell Biochem. 114:1940–1954. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu L, Stevens J, Hilton MB, Seaman S,
Conrads TP, Veenstra TD, Logsdon D, Morris H, Swing DA, Patel NL,
et al: COX-2 inhibition potentiates antiangiogenic cancer therapy
and prevents metastasis in preclinical models. Sci Transl Med.
6:242ra842014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cronin-Fenton DP, Heide-Jørgensen U, Ahern
TP, Lash TL, Christiansen P, Ejlertsen B and Sørensen HT: Low-dose
aspirin, nonsteroidal anti-inflammatory drugs, selective COX-2
inhibitors and breast cancer recurrence. Epidemiology. 27:586–593.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Santilli F, Boccatonda A and Davi G:
Aspirin, platelets, and cancer: The point of view of the internist.
Eur J Intern Med. 34:11–20. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Qin HX, Yang J, Cui HK, Li SP, Zhang W,
Ding XL and Xia YH: Synergistic antitumor activity of reversine
combined with aspirin in cervical carcinoma in vitro and in vivo.
Cytotechnology. 65:643–653. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shu Y, Liu Y, Li X, Cao L, Yuan X, Li W
and Cao Q: Aspirin-triggered resolvin D1 inhibits TGF-β1-induced
EndMT through increasing the expression of smad7 and is closely
related to oxidative stress. Biomol Ther (Seoul). 24:132–139. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li T, Dong ZR, Guo ZY, Wang CH, Tang ZY,
Qu SF, Chen ZT, Li XW and Zhi XT: Aspirin enhances IFN-α-induced
growth inhibition and apoptosis of hepatocellular carcinoma via
JAK1/STAT1 pathway. Cancer Gene Ther. 20:366–374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ding JH, Yuan LY, Huang RB and Chen GA:
Aspirin inhibits proliferation and induces apoptosis of multiple
myeloma cells through regulation of Bcl-2 and Bax and suppression
of VEGF. Eur J Haematol. 93:329–339. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Beere HM, Wolf BB, Cain K, Mosser DD,
Mahboubi A, Kuwana T, Tailor P, Morimoto RI, Cohen GM and Green DR:
Heat-shock protein 70 inhibits apoptosis by preventing recruitment
of procaspase-9 to the Apaf-1 apoptosome. Nat Cell Biol. 2:469–475.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lee SY, Choi HC, Choe YJ, Shin SJ, Lee SH
and Kim HS: Nutlin-3 induces BCL2A1 expression by activating ELK1
through the mitochondrial p53-ROS-ERK1/2 pathway. Int J Oncol.
45:675–682. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lee SY, Choe YJ, Park JY, Lee SS, Kim YH,
Shin SJ, Chung YJ and Kim HS: Wilms' tumor gene 1 enhances
nutlin-3-induced apoptosis. Oncol Rep. 31:131–136. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Patrono C, Coller B, Dalen JE, Fuster V,
Gent M, Harker LA, Hirsh J and Roth G: Platelet-active drugs: The
relationships among dose, effectiveness, and side effects. Chest.
114 5 Suppl:470S–488S. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jonsson F, Yin L, Lundholm C, Smedby KE,
Czene K and Pawitan Y: Low-dose aspirin use and cancer
characteristics: A population-based cohort study. Br J Cancer.
109:1921–1925. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Flossmann E and Rothwell PM: British
Doctors Aspirin Trial and the UK-TIA Aspirin Trial: Effect of
aspirin on long-term risk of colorectal cancer: Consistent evidence
from randomised and observational studies. Lancet. 369:1603–1613.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lu L, Sun HC, Zhang W, Chai ZT, Zhu XD,
Kong LQ, Wang WQ, Zhang KZ, Zhang YY, Zhang QB, et al: Aspirin
minimized the pro-metastasis effect of sorafenib and improved
survival by up-regulating HTATIP2 in hepatocellular carcinoma. PLoS
One. 8:e650232013. View Article : Google Scholar : PubMed/NCBI
|