Introduction
Cell surface glycans are molecules that regulate
interactions among neighboring cells or contact between cells and
the extracellular matrix during cell adhesion, recognition and
communication (1). An aberrant
glycosylation pattern promotes the development and progression of
certain pathologies, including cancer (1). Glycan alterations may result in tumor
development and progression, as well as tumor-cell dissociation and
invasion, which subsequently promote metastasis and
tumor-associated neoangiogenesis (2). Importantly, aberrant glycosylation of
the tumor cell surface results in the impairment of its recognition
by cells of the immune system, including natural killer (NK) cells
(2,3).
Alterations in glycosylation primarily arise from
changes in the expression of N-acetyl-D-glucosamine (GlcNAc)
transferases in the Golgi system (4). The family of
β-1,4-mannosyl-glycoprotein 4-β-N-acetyl
glucosaminyltransferases consists of several members, including
Mgat3 and Mgat5, which are involved in linking
terminal residues to glycans on the cell surface. The competition
between Mgat3 and Mgat5 results in the branching or
bisecting of surface glycans and this final pattern influences
intercellular recognition (5).
Mgat5 is responsible for adding β1-6 GlcNAc residues and
forming branched structures, which are particularly abundant in
cancer tissues with high metastatic potential (4). Mgat3 facilitates the addition
of β1-4 GlcNAc residues and constructs a bisecting structure that
inhibits further addition of GlcNAc by other
glucosaminyltransferases, including Mgat5 (6). Mgat3 and Mgat5 enzymes
tend to be overexpressed in tumor cells (2,7).
Song et al (8) reported
that bisecting GlcNAc on N-glycans inhibits the signaling of growth
factors and attenuates the progression of mammary tumors.
Therefore, the resulting effect of Mgat3/Mgat5
activity is likely to be tumor specific. The expression of
Mgat5 is regulated via the NM23 regulator, which is encoded
by Nme1 gene (9) and the
expression of glucosaminyltransferases, including Mgat3 and
Mgat5, is associated with their surface glycosylation
activity (10).
In vitro, cell glycosylation may be studied
using synthetic glycoconjugates (GCs) that modulate cell surface
glycosylation. GCs are able to interfere with cancer cell processes
and the cancer microenvironment; therefore they may be able to
modulate the malignant phenotype of cancer cells (11).
The present study focused on GC-triggered
alterations in the mRNA expression of surface glycosylation
regulators, as well as key components of signaling pathways
responsible for cell-extracellular matrix adhesion,
neovascularization and invasion, each of which contribute to the
metastatic potential of cancer cells. The GCs assessed included
GlcNAc moieties linked with a calix[4]arene or with a
polyamidoamine dendrimer (PAMAM) core. Calix[4]arenes possessing
distinctive geometry act as carriers of anticancer drugs (12) or as direct anticancer agents via
enzyme inhibition (13),
angiogenesis inhibition (14) or
innate immunity modulation (15).
PAMAM dendrimers have previously been used to deliver genes and
drugs (16).
Cells internalize exogenous glucosamine via the
glucose transporter and process it, as well as cellular
glucosamine, via lysosomal degradation into uridine diphosphate
glucose (UDP)-GlcNAc (17,18). This UDP-GlcNAc is further used in
the post-transcriptional modification of glycosylate substrates,
such as proteins. Glycosylation of nuclear proteins, including
histone H2B, contributes to transcriptional regulation (19). The genes encoding the sodium
glucose cotransporter 1 (Sglt1) and epidermal growth factor
receptor (Egfr1) are components of a glucose co-transporter.
Sglt1 is expressed primarily in the human intestine and
kidney (20). An example of a
Egfr1 ligand is N-acetylglucosamine (21). Glycosylation of the epidermal
growth factor receptor improves its ability to bind to epidermal
growth factor (22). The coupling
of Egfr1 to Sglt1 further stabilizes the whole
complex and enables cancer cells to take up high levels of glucose
(23).
The authors of the present study previously reported
that a GC consisting of four GlcNAc residues on a
N-acetyl-D-glucosamine-coated calix[4]arene core (GN4C)
alters the glycosylation of human NK cells and promotes the
cell-mediated cytotoxicity of human NK cells against K562 and HT-29
target cells via the phosphoinositide 3-kinase pathway (2). In addition, it has been demonstrated
that a GC consisting of eight GlcNAc residues on a PAMAM core
N-acetyl-D-glucosamine-coated polyamidoamine dendrimer
(GN8P) mediates alterations in cytokine profile specific to mouse
NK T-cells and macrophages (24).
Therefore, the present study aimed to determine whether these GCs
directly affect cancer cells by modulating the regulation of gene
expression and causing changes in the phenotypes of malignant
cells. The effect of synthetic GCs on the mRNA expression of the
cell glucosaminyltransferases Mgat3 and Mgat5,
members of the Wnt signaling family (Wnt2B and
Wnt9B), regulators of glucose metabolism (Sglt1 and
Egfr1) and regulators of cell adhesion and invasion matrix
metalloproteinase 3 (Mmp3) or transforming growth factor-β1
(Tgfb1) were measured.
Materials and methods
Preparation and treatment of cells
with GCs
The HT-29 adenocarcinoma cell line (ATCC number:
HTB-38) was obtained from American Type Culture Collection
(Manassas, VA, USA) and was authenticated following the guidelines
of the International Cell Line Authentication Committee (https://www.lgcstandards-atcc.org). Cells were
maintained in 37°C and in an RPMI-1640 medium with 10% fetal calf
serum (Sigma-Aldrich, Merck KGaA, Darmstadt, Germany). Peripheral
blood mononuclear cells (PBMC) were obtained from peripheral blood
of healthy donors (4 males, mean age 46.3±9.4; samples collected
August 2012) using a Ficoll-Paque (GE Healthcare Life Sciences,
Little Chalfont, UK) at density 1,077 g/ml and centrifugation for
40 min at room temperature and 400 × g. Blood samples were taken
from material remaining following a routine donor check-up at the
transfusion unit of the Thomayer Hospital (Prague, Czech Republic).
All donors signed informed consent for the use of their blood for
experimental purposes. The current study was approved by the
Ethical Committee of Thomayer Hospital. The chemically defined GC
calix[4]arene containing of four terminal N-acetyl-D-glucosamine
moieties (GN4C) was synthesized and kindly provided by Vladimir
Křen and Karel Křenek (Czech Academy of Sciences, Prague, Czech
Republic). A GC consisting of PAMAM with eight terminal
N-acetyl-D-glucosamine moieties (GN8P) was synthesized and kindly
provided by Thisbe Lindhorst (Christiana Albertina University,
Kiel, Germany). The synthesis, purity, nuclear magnetic resonance
data and dose-dependent effect of the GCs on human PBMC cells have
been previously described (2,24–26).
The optimal concentration of 10 nM GN4C or GN8P were used for the
experiments. Fludara® (1 mM, FLU; Genzyme, Cambridge,
MA, USA), which is a conventional anticancer drug, was used to
compare the anticancer effect of the tested GCs with a known
anticancer agent.
Glyco-gene profiling array
The HT-29 cell line was incubated with GN8P for 24 h
and total RNA was isolated using an RNAeasy Mini kit involving
DNAse I treatment following the manufacturer's protocol (DNAse I
was a component of RNAeasy Mini kit; Qiagen GmbH, Hilden, Germany).
A Glyco-gene Chip array (GLYCOv3 Gene Chip; Affymetrix; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) containing probe sets of
2,000 human transcripts was provided by the Consortium for
Functional Glycomics (http://www.functionalglycomics.org/static/consortium/resources/resourcecoree.shtml).
Microarray experiments were conducted by The Microarray Gene Core
of Consortium for Functional Glycomics, National Institutes of
Health (NIH)/National Institute of General Medical Sciences
(NIGMS), both Bethesda, MD, USA (http://www.functionalglycomics.org) and were performed
in triplicate. BRB ArrayTools version 4.3.0 beta 3 [Biometric
Research Branch, NIH/National Cancer Institute (NCI)] were used to
filter and analyze experimental data sets. Class comparison used a
two-sample t-test with a random variance model. A P-value of
log-ratio <0.05 was considered significant.
Gene Ontology analysis was performed using the
Database for Annotation, Visualization and Integrated Discovery
(DAVID) software version 6.7 available from NCI (http://david.abcc.ncifcrf.gov). The Gene Card database
was used for basic gene identification and characterization
(http://www.genecards.org; Weizmann Institute of
Science, Rehovot, Israel).
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
The HT-29 cell line was incubated with GCs and FLU
for 24 h and total RNA was isolated using an RNAeasy Mini kit
involving DNAse I treatment, as described by the manufacturer
(Qiagen). A total of 5 µg RNA was transcribed into cDNA using a
cDNA Archive kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.). qPCR was performed using PowerSybr Green Master mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.) on an iCycler5 (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). PCR product specificity was
analyzed by melt curve analysis. The cycling conditions recommended
by the Master mix manufacturer were followed: initiation 95°C 10
min and 40 cycles of 95°C 15 sec and 60°C 1 min. The primers used
for PCR were designed using Primer3 Input software version 0.4.0
(National Center for Biotechnology Information, Bethesda, MD, USA).
The sequences of primers were as follows: Mgat5, forward,
5′-CTTCTTTCTTCCAGCACCTCAAC-3′ and reverse,
5′-AAACACACAGTGCTTATTCTTAGGG-3′; Nme1, forward,
5′-ACCTTCATTGCGATCAAACC-3′ and reverse, 5′-GGCCCTGAGTGCATGTATTT-3′;
Siglec5, forward, 5′-CAAGTGCAGAAGTCGGTGAC-3′ and reverse,
5′-GGGTCTCTGGCTTCACTCTT-3′; Siglec8, forward,
5′-TGCAACCCTCAGCTTCCATA-3′ and reverse, 5′-ACTTCTTTGCTGGAGGGGTT-3′;
Wnt9B, forward, 5′-TGGGCAGACTGTCATCACAT-3′ and reverse,
5′-AACAAGGTTGGGGATGCTTG-3′. Sequences of the primers used to
amplify B2M, Egfr1 and Sglt1, Mki67, Mmp3, Tgfb1 and
Wnt2B were described previously (27–31):
B2M, forward, 5′-GAGTATGCCTGCCGTGTG-3′ and reverse,
5′-AATCCAAATGCGGCATCT-3′; Egfr1, forward,
5′-TTTCGATACCCAGGACCAAGCCACAGCAGG-3′ and reverse,
5′-AATATTCTTGCTGGATGCGTTTCTGTA-3′; Sglt1, forward,
5′-TGGCAGGCCGAAGTATGGTGT-3′ and reverse,
5′-ATGAATATGGCCCCCGAGAAGA-3′; Mki67, forward,
5′-GGAGGCAATATTACATAATTTCA-3′ and reverse,
5′-CAGGGTCAGAAGAGAAGCTA-3′; Mmp3, forward,
5′-ATGCCCACTTTGATGATGATGAAC-3′ and reverse,
5′-CCACGCCTGAAGGAAGAGATG-3′; Tgfb1, forward,
5′-TGACAGCAGGGATAACACACT-3′ and reverse,
5′-GTAGGGGCAGGGCCCGAGGCA-3′; Wnt2B, forward,
5′-CACCTGCTGGCGTGCACTCTCAGA-3′ and reverse,
5′-GGGCTTTGCAAGTATGGACGTCCACAGTA-3′. Primers for Mgat3 were
obtained from SA Biosciences; Qiagen (product ID: PPH01058A). The
expression of the genes of interest were normalized to that of the
control gene B2M and quantified using the 2-ΔΔCq
method (32). Differences in the
expression of genes between the untreated and GC-treated cells were
evaluated using Bio-Rad iQ5 version 2.0 (Bio-Rad Laboratories,
Inc.).
Cell proliferation and colony
formation assays
To determine the effect of GCs on cell growth,
5×103 cells/well were seeded in triplicate on a 96-well
plate and incubated with GCs for 126 h. FLU (1 mM) was used as a
positive control of tumor cell growth inhibition. Negative controls
represent cells with no additional treatment. Cell growth was
monitored continuously using a Real-Time Cell Analyzer (RTCA;
xCELLigence System; Acea Biosciences, San Diego, CA, USA;
https://www.aceabio.com/products/rtca-dp/) and
evaluated using RTCA software version 1.2 (Acea Biosciences).
To study colony formation, a standard colony forming
assay and an impedance-based assay using the RTCA instrument (Acea
Biosciences) were performed. For colony formation assays,
1×103 were seeded in a Petri dish 100 mm in diameter.
After 2 weeks, colonies were fixed for 30 min with 70% ethanol
(Sigma-Aldrich; Merck KGaA) at room temperature and subsequently
stained for 10 min at room temperature with 0.5% crystal violet
(Sigma-Aldrich; Merck KGaA). RTCA was also used to evaluate colony
growth in order to obtain quantitative data. Here, 100 cells/well
were seeded in triplicate on a 96-well plate and colony growth was
monitored using RTCA for 2 weeks.
Apoptosis assay and cell-mediated
cytotoxicity test
GC-mediated toxicity in the HT-29 cancer cell line
was assessed using propidium iodide (PI) staining and subsequent
flow cytometry with a BD LSRII (BD Biosciences, Franklin Lakes, NJ,
USA). Apoptosis was measured using an FITC Annexin V Apoptosis
Detection kit I (Annexin V-fluorescein isothiocyanate, BD
Biosciences) following the manufacturer's protocol. The results
were analyzed using FlowJo version 7.2.2 software (FlowJo LLC,
Ashland, OR, USA).
Cell-mediated cytotoxicity was performed using a
standard chromium release assay, as previously described (2). Briefly, HT-29 cells (2×104
cells/well) pretreated with or without GCs for 30 min were
incubated with 51Cr for 2 h and served as target cells
for PBMCs (total volume 0.1 ml/well). All samples were tested in
triplicate. The effector:target ratio of 16:1 was optimal for the
experimental conditions. Following 4 h co-incubation of target and
PBMC cells, 51Cr release was measured in cell-free
supernatants (obtained as supernatants following centrifugation of
cells at 2,000 × g for 5 min at room temperature) using a Wallac
Microbeta Trilux scintillation counter (PerkinElmer, Inc., Waltham,
MA, USA). The percentage specific lysis was calculated using the
following formula: [Experimental counts per minute (cpm) -
spontaneous cpm]/(maximum cpm - spontaneous cpm) × 100, where
maximum cpm was determined by addition of 10% Triton X-100.
Statistical analysis
Statistically significant differences in the
parameters tested in HT-29 cells cultured in the presence or
absence of GCs were assessed using a one-way analysis of variance
followed by Dunnett's post hoc test and a confidence interval of
95%. Statistical analysis was conducted using GraphPad Prism 5
software (GraphPad Software, Inc., La Jolla, CA, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
GC stimulation modulates the mRNA
expression of glucosaminyltransferases, components of glucose
metabolism and adhesion molecules
Previous research has noted alterations in the
phenotype of cancer cells following treatment with GCs. Therefore,
the present study investigated a wide range of candidate molecules
and the results revealed that the interaction of cancer cells with
GCs induced complex alterations in gene expression.
Candidate molecules were selected based on the novel
results of a glyco-gene array investigating GN8P, as well as
previously published results of glyco-gene profiling of GN4C in
NK-92 cancer cells (2). Regarding
the glyco-gene array performed on cell samples incubated for 24 h
in the presence or absence of GN8P, the present study excluded all
genes in which the percentage of absent data (all data where two
out of three parallels were missing) >50% and where P>0.05.
Subsequently, class comparison identified 22 genes that exhibited
significantly different expression between control and GN8P-treated
HT-29 cells (P<0.05). The responsive genes were functionally
categorized according to Gene Ontology classification and the Gene
Card database. A total of 64% of the genes listed were linked to
cancer and 14% of the genes were linked to inflammation.
Differentially expressed genes were primarily involved in signal
transduction (28%), carbohydrate binding (23%), proliferation (14%)
and immune processes (14%). A complete list of the differentially
expressed genes, including their function and disease association,
is presented in Table I. Along
with other genes, GN8P mediated the downregulation of Wnt
signaling molecules that serve an important role in cancer
progression; thus inhibition of their expression is of particular
interest. Upregulated genes included protein tyrosine phosphatase
Ptprt and tyrosine kinase Flt3 (Table I). Candidate genes were validated
by RT-qPCR (data not shown).
 | Table I.List of GN8P-responsive genes in the
HT-29 cell line. |
Table I.
List of GN8P-responsive genes in the
HT-29 cell line.
| Gene | Entrez gene ID | Biological
function | Disease
association | (Refs.) |
|---|
| Upregulated
genes |
|
|
|
|
|
Mpl | 4352 | GFR, ST, CP | CAMT, TR, PV,
MD | (33–35) |
|
Flt3 | 2322 | Ig, TK, ST, CP | C-L (AML, ALL) | (36,37) |
|
Bmp7 | 655 | GF, TGF | RD, OS, PA | (38–40) |
|
Ptprt | 11122 | ST, PP | CoCa, glioma | (41,42) |
|
Ppbp | 5473 | GF, GTTA, CP,
I | IN, ET, TP | (43–45) |
|
Cxcl2 | 2920 | I, CP | C, N, S | (46,47) |
|
Gpc3 | 2719 | CP | HepCa, WT | (48,49) |
| Downregulated
genes |
|
|
|
|
|
Siglec12 | 89858 | CB, adheze | – | (50) |
|
Nrg1 | 3084 | GF, TF | C, SC | (51) |
|
Ccl7 | 6354 | I, ST | IN, AS | (52) |
|
Lgals13 | 29124 | CB, GAL, LPL | Pre-eclampsia | (53) |
|
Csf2ra | 1438 | ST, CP | C, N | (54) |
|
Wnt9B | 7484 | ST | C | (55) |
|
Clec4C | 170482 | CB, I | – | (56) |
|
Wnt2B | 7482 | ST | C | (57) |
|
Mrc1 | 4360 | CB, L, RI | TBC, ALL | (58,59) |
|
Cxccl9 | 4283 | ST, I | N, IN | (60) |
|
Sgsh | 6448 | hydrolase, GB | MPS | (61) |
|
Clec3B | 7123 | CB, L, PAP | C | (62) |
|
Gdf11 | 10220 | TGF, GF | ALL | (63) |
|
Emr2 | 30817 | ST | CoCa | (64) |
|
Hs3st5 | 222537 | GB | CoCa | (65) |
For a detailed examination of the effect of GCs on
the glycosylation mechanism in HT-29 cells, the expression of
glucosaminyltransferases involved in terminal glycan elongation and
genes involved in glucose uptake and cell adhesion, were measured.
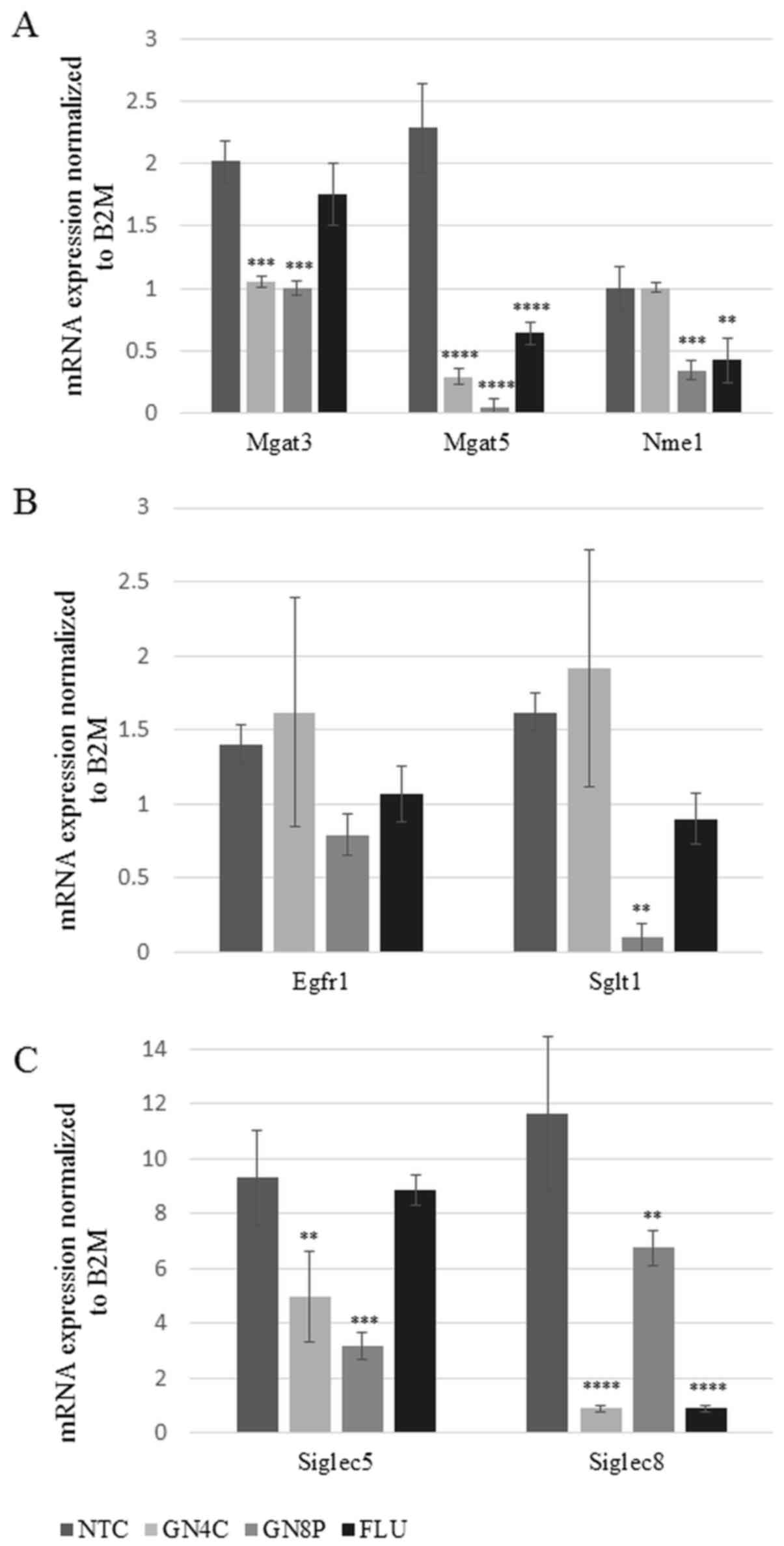
When compared with the NTC group, treatment with the GN4C and GN8P
significantly downregulated expression levels of Mgat3
(P=0.0002 and P=0.0001, respectively) and Mgat5 (each,
P=0.0001) glucosaminyltransferases. FLU also inhibited the
expression of Mgat5 (P=0.0001) compared with the NTC group.
The Nme1 gene encodes a transcription regulator controlling
expression of the Mgat5 gene (9). Compared with the NTC group, the mRNA
expression of this transcription factor was significantly
downregulated by GN8P and FLU (P=0.0009 and P=0.002, respectively;
Fig. 1A).
Sglt1 and Egfr1 are components of a
glucose cotransporter in human cells. The mRNA expression of these
genes was significantly reduced by GN8P (P=0.0054 for SGLT1).
However, the reduction of EGFR1 expression following exposure to
GN8P was of borderline significance (P=0.2) (Fig. 1B). Furthermore, the mRNA expression
of the glycan binding adhesion molecules Siglec5 and
Siglec8 were downregulated by GN4C (P=0.0078 and P=0.0001,
respectively) as well as by GN8P (P=0.0009 and P=0.0081,
respectively; all Fig. 1C).
However, FLU only inhibited the mRNA expression of Siglec8
(P=0.0001; Fig. 1C).
GCs inhibit HT-29 cell proliferation,
induce apoptosis and promote PBMC-mediated cytotoxicity
The present study questioned whether GCs alter the
proliferation of cancer cells. To test this hypothesis, real-time
monitoring of cell proliferation was performed for 5 days. A
proliferation protocol was optimized using an RTCA system allowing
the continuous monitoring of treated and untreated cells for a set
period of time. The results indicated that the proliferation of
HT-29 cells treated with GN4C, GN8P or FLU was significantly
inhibited from ~5 h following induction. The greatest inhibition of
cell growth was detected 60 h following treatment in the cells
treated with FLU, GN8P and GN4C (P=0.0001 for all treatments vs.
NTC group; Fig. 2A). A significant
reduction in the expression of the proliferation antigen
Mki67 48 h following treatment with GN4C (P=0.0017), GN8P
(P=0.0005) and FLU (P=0.0003; Fig.
2B), was observed, compared with the NTC group.
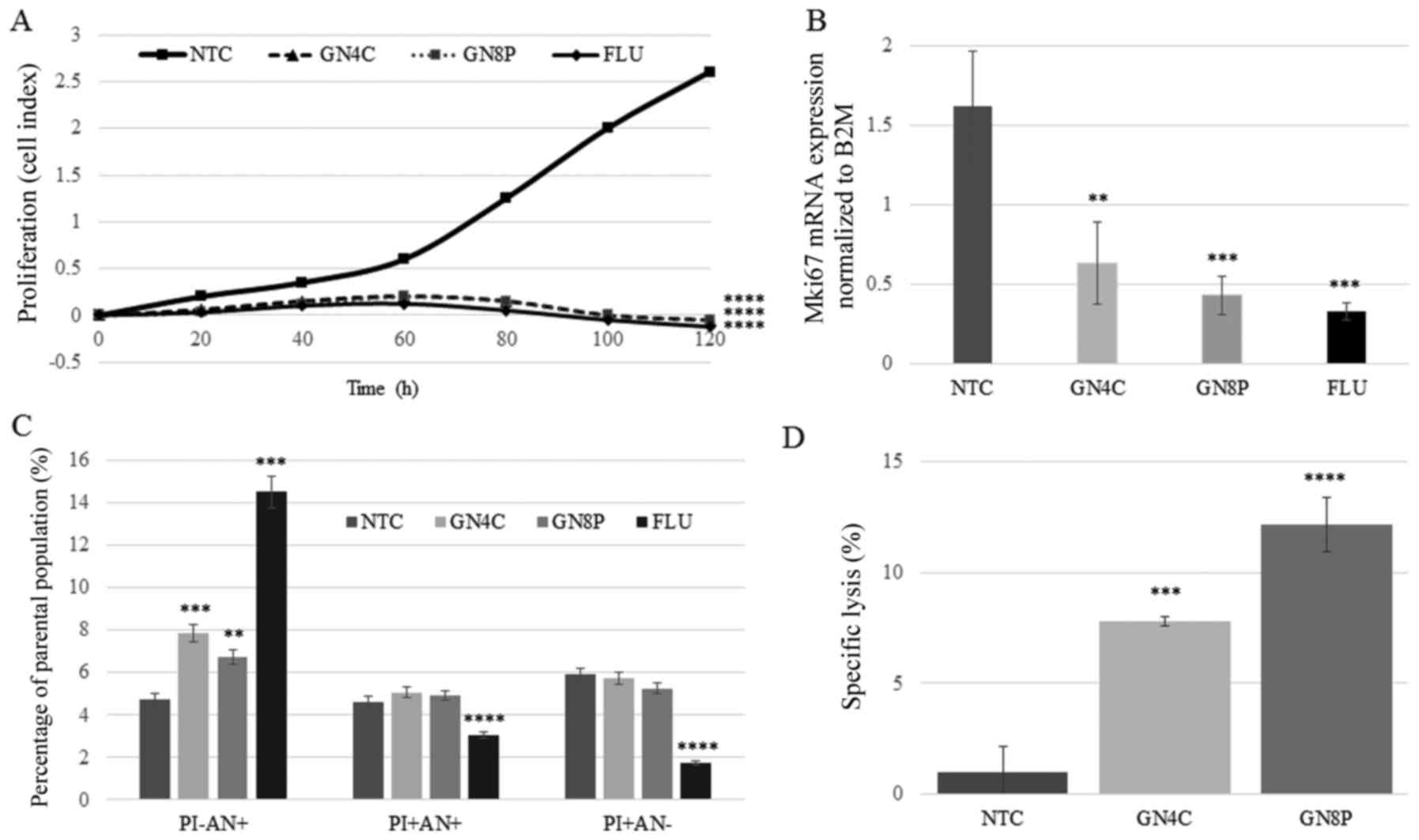 | Figure 2.Effect of synthetic glycoconjugates
on cell proliferation, apoptosis and sensitivity to cell-mediated
killing. (A) Proliferation curve of cancer cells measured using a
Real-Time Cell analyzer. The cell index reflects the number of
live, attached cells. Average values of triplicates are plotted.
(B) mRNA expression (fold change) of the proliferative antigen
Mki67. (C) Detection of apoptotic and necrotic cells by flow
cytometric analysis. The early apoptotic population was represented
by cells that were PI- and AN+; the late apoptotic population was
represented by cells that were PI+ and AN+; the necrotic population
was represented by cells that were PI+ and AN-. (D) The cell
mediated cytotoxicity assay employed PBMCs. Data are presented as
the mean ± standard deviation. Experiments were performed in
triplicate. **P<0.01, ***P<0.001 and ****P<0.0001 vs. NTC
control group. PBMCs, peripheral blood mononuclear cells; NTC,
glycoconjugate-non-treated control with basic toxicity of PBMC to
HT-29 cancer cells; GN4C and GN8P, cells treated with GN4C or GN8P,
respectively; FLU, cells treated with Fludara that represented the
positive control; GN4C, N-acetyl-D-glucosamine-coated
calix[4]arene; GN8P, N-acetyl-D-glucosamine-coated polyamidoamine
dendrimer; PI, propidium iodide; AN, Annexin V; +, positive; -,
negative. |
To determine the toxicity of GCs in the HT-29 cell
line, Annexin V positivity (specific marker of apoptosis) and the
incorporation of PI into cells were measured. The percentage of
early apoptotic, Annexin V-positive and PI-negative cells was
significantly increased following treatment with GN4C, GN8P and FLU
(P=0.0001, P=0.002 and P=0.0001, respectively), compared with the
NTC group. Treatment with FLU induced a significant decrease in the
population of cells in the late apoptotic and necrotic phases
compared with the NTC group (all P=0.0001; Fig. 2C) However, treatment with GCs had
no effect on the proportion of cells in the late apoptotic and
necrotic stages of apoptosis (Fig.
2C). The proportion of untreated HT-29 cells that underwent
spontaneous necrosis (PI positive, Annexin V-negative population)
was 5.8%; comparable with the percentage of necrotic cells found in
GN4C and GN8P treated cells (Fig.
2C). These results indicate that GCs preferably induce
apoptosis over necrosis.
Surface glycosylation is a tool facilitating the
recognition of cancer cells by immune cells. The current study
questioned whether alterations in cancer cell glycosylation
sensitizes them to PBMC-mediated toxicity. HT-29 cells, acting as
target cells, which were naturally resistant to human PBMC-mediated
cytotoxicity, were pretreated with GCs and subsequently exposed to
PBMCs from healthy donors, acting as effector cells. Significantly
higher cytolytic activity was observed in HT-29 cells pretreated
with GN4C and GN8P (P=0.0002 and P=0.0001, respectively; Fig. 2D). The rate of spontaneous toxicity
in untreated HT-29 cells against human PBMCs was ~1.2% (Fig. 2D).
Synthetic GCs reduce cancer cell
colony formation
As cell invasion is associated with extracellular
matrix remodeling and neoangiogenesis, levels of regulatory
components, including Mmp3, Tgfb1, Wnt2B and Wnt9B,
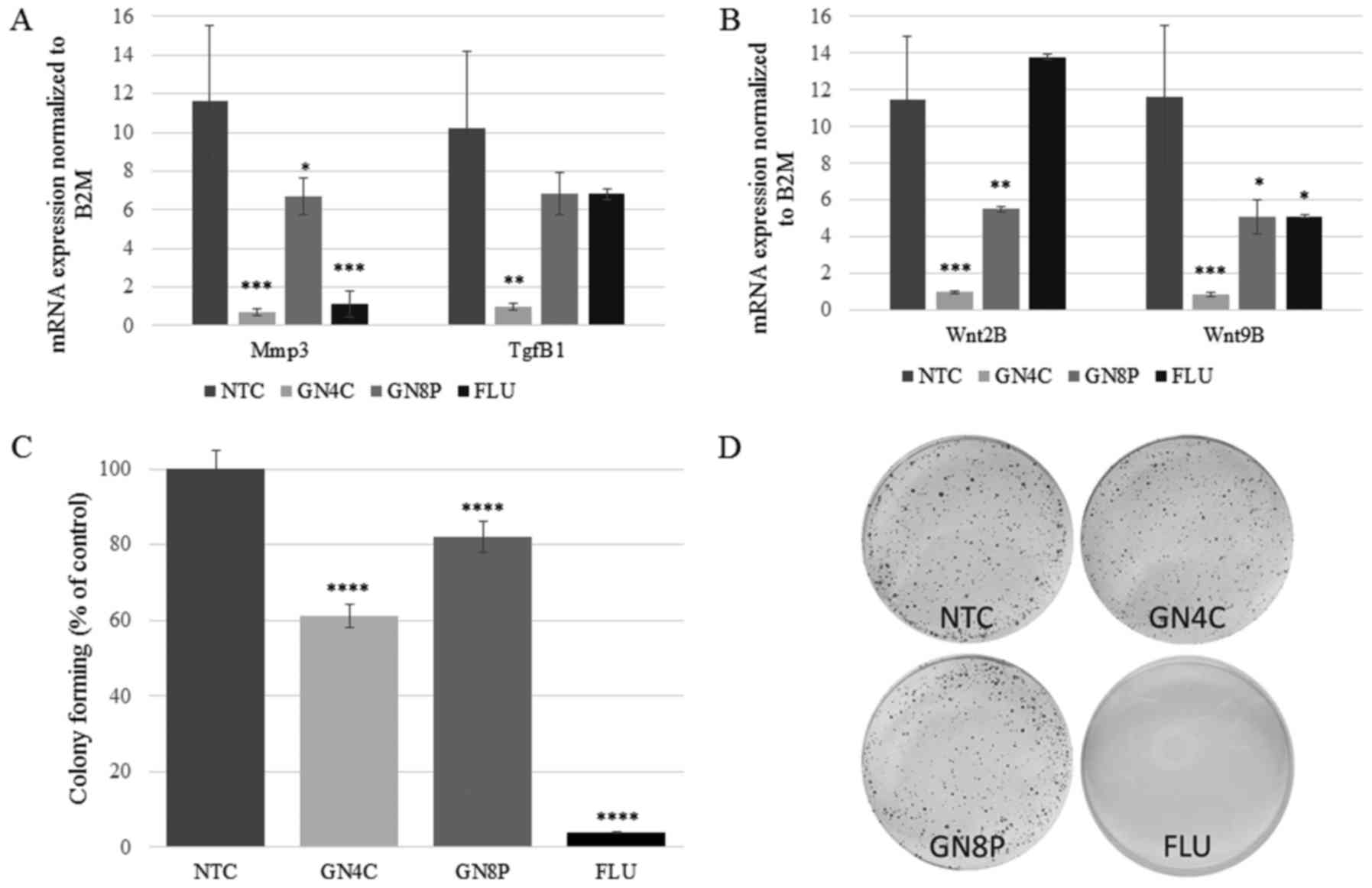
were measured. There was a significant decrease in the expression
of Mmp3 mRNA following treatment with GN4C and FLU (P=0.0005
and P=0.0006, respectively; Fig.
3A). GN4C inhibited the expression of Tgfb1 (P=0.0016;
Fig. 3A). GN4C and GN8P decreased
the expression of Wnt2B (P=0.0002 and P=0.0066,
respectively) and Wnt9B (P=0.0004 and P=0.0101,
respectively; Fig. 3B). FLU
significantly decreased the expression of Wnt9B (P=0.01;
Fig. 3B) but not Wnt2B.
Treatment of HT-29 cells with GN4C and GN8P led to decreases in
cell colony formation by 39 and 18%, respectively (P=0.0001 vs.
NTC). FLU treated (positive control) cells exhibited a decrease in
cell colony formation of 96% compared with the NTC group (P=0.0001;
Fig. 3C). These changes in colony
formation were confirmed following standard crystal-violet
staining. There was a reduction in colonies in samples incubated
with GCs and FLU (Fig. 3D).
Discussion
Previous studies have focused on the identification
of genes involved in the response of NK cells isolated from PBMC
and the permanent NK-92 cell line to the synthetic GC GN4C
(2), or on the role of GN8P in the
activation of immune cells (24).
Phenotypic alterations in cancer cells following
exposure to GCs has been observed. Calix[6]arenes exhibit an
anticancer effect by modulating AXL and Mer tyrosine kinase
receptor gene expression (66);
therefore, the present study questioned whether the phenotypic
changes in cancer cells were modulated by changes in gene
expression. The current study specifically focused on the
expression of cancer development-related genes and how gene
expression is affected by two GlcNAc-modified GCs that contain
different cores: First generation PAMAM or calix[4]arene.
Previous studies have reported that exogenous
glucosamine, the terminal moiety of the tested GCs, may be
internalized via a glucose transporter (17,18).
In HT-29 cells, clathrin-mediated endocytosis has been described as
a mechanism of uptake of third generation PAMAM (67). Cancer cells, such as HT-29, exhibit
a high glucose intake due to high-energy requirements (22). The sodium-glucose cotransporter
(SGLT1) transports glucose into cells independent of its
concentration (68). SGLT1 is
stabilized by interaction with EGFR1, facilitating cancer cell
survival (27). In colon cancer,
the high expression of Sglt1 and Egfr1 is associated
with poor patient prognosis (69).
The results of the present study demonstrated that the PAMAM-based
GC significantly decreased the expression of the two genes
Sglt1 and Egfr1 that code for the glucose
cotransporter complex. This, in turn, inhibited cell growth and
reduced levels of the proliferation antigen Mki67.
It has been reported that higher generation
dendrimers (G2, G4 and G6) promote cell growth at lower
concentrations but induce cell death at higher concentrations. The
critical concentration at which dendrimers induced cell death was
500 nM. Also, toxicity was enhanced by higher generation dendrimers
(70). The current study observed
that a first generation PAMAM dendrimer with eight GlcNAc moieties
induced alterations in gene expression and altered the properties
of cancer cells even at low concentrations (10 nM). Comparing the
responses of cells to GCs and FLU identified the different
underlying mechanisms of action of each compound. FLU is a potent
inducer of apoptosis and affected cells undergo rapid
disintegration, thus explaining the decrease in the number of cells
in the late apoptotic and necrotic phases following treatment with
FLU.
The growth of cancer cells was markedly affected by
the GC GN8P and changes in cell adhesion and invasion were
associated with the altered expression of specific mRNAs. The
Siglec family contains proteins that serve an important role in
cancer cell adhesion and invasion. Human cell-surface-receptors,
including sialic acid-binding Ig-like lectin (SIGLEC) 5 and
SIGLEC8, are members of the cluster of differentiation 33-related
Siglec subfamily, expressed predominantly by immune cells (71) and they are overexpressed in acute
myeloid leukemia, chronic eosinophilic and myelogenous leukemias
(72,73). In solid tumors, Siglecs are
overexpressed in tumor-associated immune cells, such as
macrophages; however there is little evidence regarding their
overexpression in actual cancer cells (74). The alteration in Siglec mRNA
expression observed in the present study following the incubation
of tumor cells with GC may be due to alterations in Mgat5
expression.
In colon cancer, glycosylation performed by MGAT5
may regulate colon cancer stem cells and tumor progression via Wnt
signaling (75). As GCs
downregulate the expression of Mgat5 and exhibit reduced
colony formation, the present study measured the expression of the
Wnt family members Wnt2B and Wnt9B, the
metalloproteinase Mmp3, which is responsible for
extracellular matrix remodeling (76) and Tgfb1, which is involved
in tumor neoangiogenesis (77).
These genes were selected based on the preliminary results of cDNA
profiling in GN8P-treated cancer cells (unpublished results). GCs
reduced the expression of Wnt2B and Wnt9B; however,
only GN4C inhibited the expression of Mmp3 and Tgfb1.
This may be due to the fact GN4C is more effective at inhibiting
cell invasion; indeed, it has been demonstrated that calix[4]arene
inhibits cancer angiogenesis (78).
The present study identified a specific pattern in
gene expression, common to the two GCs, which included
downregulation of i) the glucosaminyltransferases Mgat3 and
Mgat5, ii) the adhesion molecules Siglec5 and
Siglec8, iii) Wnt2B and Wnt9B and iv) the
proliferation marker Mki67. The two GCs increased the
proportion of cells in the early apoptotic phase and the
sensitivity of cancer cells to PBMCs. The current study focused the
effect of GCs in cancer cells and revealed associations that may
allow for the investigation of individual components in a different
perspective or focus on specific signaling pathways induced by the
GCs. It has been demonstrated that glycosylation in NK cells
involves the phosphoinositide 3-kinase signaling pathway (2). Based on information from a recently
published report (79),
alterations regarding the sensitivity to PBMCs may be due to the
GlcNAc section of the GCs. In the case of the downregulation of the
Sglt1/Egfr1, the PAMAM core may induce this change since
GN8P significantly lowered expression of the Sglt1 component but
GN4C demonstrated no such effect. In myotubes, Wnt signaling
affects glucose transport via a glucose transporter (80). In the present study, the same
members of the Wnt pathway (Wnt2B and Wnt9B) were
downregulated by GN4C. No differences in Sglt1/Egfr1
expression following treatment with GN4C were detected. This
suggests that these factors are not involved in colon cancer cell
signaling, however, further analysis may reveal if there is a
similar association between the SGLT1/EGFR1 cotransporter and other
members of Wnt pathway.
It is likely that other members of the Wnt signaling
pathway are involved and responsible for this distinctive response
to particular cores. The present study demonstrated that Wnt
signaling may be involved in the response of cancer cells to
synthetic GCs, possibly via modulation of glucosaminyltransferases.
Subsequent detailed studies focusing on the Wnt pathway may
identify response-specific members of the Wnt family.
The evolutionarily conserved Wnt signaling mechanism
is an important pathway and Wnt proteins undergo post-translational
glycosylation. Therefore, anticancer therapies that target
Wnt-signaling members, based on glycosylation modulation, may be
developed as a novel therapeutic strategy.
In conclusion, the present study demonstrated that
the interaction of colon cancer cells with specifically designed
GCs results in a complex commitment of different cellular pathways
and induces alterations in the phenotypes of cells. The results of
the current study revealed that alterations in the expression of
particular genes following treatment with GCs are associated with
specific outcomes in cancer cells, including their higher
sensitivity to immune cell-mediated killing. The GCs used in the
current study exhibited multiple effects following their
application to cancer cells. These results, together with those of
previous studies determining the immunostimulatory effects of GCs,
support the importance of glycosylation-targeted anticancer therapy
and provides a basis for further studies.
Acknowledgements
The authors wish to thank The Consortium for
Functional Glycomics/NIGMS/Gene Microarray Core (grant no. GM62116)
for their resources and collaboration. The authors thank Cory
Benson for editing the English language of the manuscript,
Professors Vladimır Křen and Thisbe Lindhorst for providing GCs and
Dr Lucie Vondráčková for providing the xCELLigence system. The
present study was supported by the Ministry of Health of the Czech
Republic (grant no. 15-33094A) and the Czech Science Foundation
(grant no. 14-10100S).
Glossary
Abbreviations
Abbreviations:
|
GC
|
glycoconjugate
|
|
GlcNAc
|
N-acetylglucosamine
|
|
GN4C
|
N-acetyl-D-glucosamine-coated
calix[4]arene
|
|
GN8P
|
N-acetyl-D-glucosamine-coated
polyamidoamine dendrimer
|
|
PAMAM
|
polyamidoamine
|
|
PBMC
|
peripheral blood mononuclear cells
|
|
RTCA
|
Real-Time Cell Analyzer
|
References
|
1
|
Kobata A and Amano J: Altered
glycosylation of proteins produced by malignant cells and
application for the diagnosis and immunotherapy of tumours. Immunol
Cell Biol. 83:429–439. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Benson V, Grobarova V, Richter J and
Fiserova A: Glycosylation regulates NK cell-mediated effector
function through PI3K pathway. Int Immunol. 22:167–177. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lau E, Feng Y, Claps G, Fukuda MN, Perlina
A, Donn D, Jilaveanu L, Kluger H, Freeze HH and Ronai ZA: The
transcription factor ATF2 promotes melanoma metastasis by
suppressing protein fucosylation. Sci Signal. 8:ra1242015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pinho SS and Reis CA: Glycosylation in
cancer: Mechanisms and clinical implications. Nat Rev Cancer.
15:540–555. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cipolla L, La Ferla B and Gregori M:
Combinatorial approaches to iminosugars as glycosidase and
glycosyltransferase inhibitors. Comb Chem High Throughput Screen.
9:571–582. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Abbott KL, Nairn AV, Hall EM, Horton MB,
McDonald JF, Moremen KW, Dinulescu DM and Pierce M: Focused
glycomic analysis of the N-linked glycan biosynthetic pathway in
ovarian cancer. Proteomics. 8:3210–3220. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kang X, Wang N, Pei C, Sun L, Sun R, Chen
J and Liu Y: Glycan-related gene expression signatures in human
metastatic hepatocellular carcinoma cells. Exp Ther Med. 3:415–422.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Song Y, Aglipay JA, Bernstein JD, Goswami
S and Stanley P: The bisecting GlcNAc on N-glycans inhibits growth
factor signaling and retards mammary tumor progression. Cancer Res.
70:3361–3371. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chakraborty AK, Sousa Jde F, Chakraborty
D, Funasaka Y, Bhattacharya M, Chatterjee A and Pawelek J: GnT-V
expression and metastatic phenotypes in macrophage-melanoma fusion
hybrids is down-regulated by 5-Aza-dC: Evidence for methylation
sensitive, extragenic regulation of GnT-V transcription. Gene.
374:166–173. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lau KS and Dennis JW: N-Glycans in cancer
progression. Glycobiology. 18:750–760. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yoshinaga A, Kajiya N, Oishi K, Kamada Y,
Ikeda A, Chigwechokha PK, Kibe T, Kishida M, Kishida S and Komatsu
M: NEU3 inhibitory effect of naringin suppresses cancer cell growth
by attenuation of EGFR signaling through GM3 ganglioside
accumulation. Eur J Pharmacol. 782:21–29. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Galindo-Murillo R, Sandoval-Salinas ME and
Barroso-Flores J: In Silico design of monomolecular drug carriers
for the tyrosine kinase inhibitor drug imatinib based on calix- and
thiacalix [n]arene host molecules: A DFT and molecular dynamics
study. J Chem Theory Comput. 10:825–834. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cherenok S, Vovk A, Muravyova I, Shivanyuk
A, Kukhar V, Lipkowski J and Kalchenko V: Calix[4]arene
alpha-aminophosphonic acids: Asymmetric synthesis and
enantioselective inhibition of an alkaline phosphatase. Org Lett.
8:549–552. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou H, Wang DA, Baldini L, Ennis E, Jain
R, Carie A, Sebti SM and Hamilton AD: Structure-activity studies on
a library of potent calix[4]arene-based PDGF antagonists that
inhibit PDGF-stimulated PDGFR tyrosine phosphorylation. Org Biomol
Chem. 4:2376–2386. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Geraci C, Consoli GM, Galante E, Bousquet
E, Pappalardo M and Spadaro A: Calix[4]arene decorated with four Tn
antigen glycomimetic units and P3CS immunoadjuvant: Synthesis,
characterization and anticancer immunological evaluation. Bioconjug
Chem. 19:751–758. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Esfand R and Tomalia DA: Poly(amidoamine)
(PAMAM) dendrimers: From biomimicry to drug delivery and biomedical
applications. Drug Discov Today. 6:427–436. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schleicher ED and Weigert C: Role of the
hexosamine biosynthetic pathway in diabetic nephropathy. Kidney Int
Suppl. 77:S13–S18. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hinderlich S, Berger M, Schwarzkopf M,
Effertz K and Reutter W: Molecular cloning and characterization of
murine and human N-acetylglucosamine kinase. Eur J Biochem.
267:3301–3308. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Phatnani HP and Greenleaf AL:
Phosphorylation and functions of the RNA polymerase II CTD. Genes
Dev. 20:2922–2936. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gorboulev V, Schürmann A, Vallon V, Kipp
H, Jaschke A, Klessen D, Friedrich A, Scherneck S, Rieg T, Cunard
R, et al: Na+-D-glucose cotransporter SGLT1 is pivotal
for intestinal glucose absorption and glucose-dependent incretin
secretion. Diabetes. 61:187–196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ferguson KM, Berger MB, Mendrola JM, Cho
HS, Leahy DJ and Lemmon MA: EGF activates its receptor by removing
interactions that autoinhibit ectodomain dimerization. Mol Cell.
11:507–517. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang XQ, Sun P, O'Gorman M, Tai T and
Paller AS: Epidermal growth factor receptor glycosylation is
required for ganglioside GM3 binding and GM3-mediated suppression
(correction of suppresion) of activation. Glycobiology. 11:515–522.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Grobárová V, Benson V, Rozbeský D, Novák P
and Cerný J: Re-evaluation of the involvement of NK cells and
C-type lectin-like NK receptors in modulation of immune responses
by multivalent GlcNAc-terminated oligosaccharides. Immunol Lett.
156:110–117. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Krenek K, Kuldová M, Hulíková K, Stibor I,
Lhoták P, Dudic M, Budka J, Pelantová H, Bezouska K, Fiserová A and
Kren V: N-acetyl-D-glucosamine substituted calix[4]arenes as
stimulators of NK cell-mediated antitumor immune response.
Carbohydr Res. 342:1781–1792. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vannucci L, Fiserová A, Sadalapure K,
Lindhorst TK, Kuldová M, Rossmann P, Horváth O, Kren V, Krist P,
Bezouska K, et al: Effects of N-acetyl-glucosamine-coated
glycodendrimers as biological modulators in the B16F10 melanoma
model in vivo. Int J Oncol. 23:285–296. 2003.PubMed/NCBI
|
|
27
|
Weihua Z, Tsan R, Huang WC, Wu Q, Chiu CH,
Fidler IJ and Hung MC: Survival of cancer cells is maintained by
EGFR independent of its kinase activity. Cancer Cell. 13:385–393.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Han Y, San-Marina S, Yang L, Khoury H and
Minden MD: The zinc finger domain of Wilms' tumor 1 suppressor gene
(WT1) behaves as a dominant negative, leading to abrogation of WT1
oncogenic potential in breast cancer cells. Breast Cancer Res.
9:R432007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ullmannová V, Stöckbauer P, Hradcová M,
Soucek J and Haskovec C: Relationship between cyclin D1 and
p21Waf1/Cip1 during differentiation of human myeloid
leukemia cell lines. Leuk Res. 27:1115–1123. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Klatt AR, Paul-Klausch B, Klinger G, Kühn
G, Renno JH, Banerjee M, Malchau G and Wielckens K: A critical role
for collagen II in cartilage matrix degradation: Collagen II
induces pro-inflammatory cytokines and MMPs in primary human
chondrocytes. J Orthop Res. 27:65–70. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Van Den Berg DJ, Sharma AK, Bruno E and
Hoffman R: Role of members of the Wnt gene family in human
hematopoiesis. Blood. 92:3189–3202. 1998.PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Besancenot R, Roos-Weil D, Tonetti C,
Abdelouahab H, Lacout C, Pasquier F, Willekens C, Rameau P, Lecluse
Y, Micol JB, et al: JAK2 and MPL protein levels determine
TPO-induced megakaryocyte proliferation vs differentiation. Blood.
124:2104–2115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Moliterno AR, Hankins WD and Spivak JL:
Impaired expression of the thrombopoietin receptor by platelets
from patients with polycythemia vera. N Engl J Med. 338:572–580.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Moliterno AR, Williams DM, Rogers O and
Spivak JL: Molecular mimicry in the chronic myeloproliferative
disorders: Reciprocity between quantitative JAK2 V617F and Mpl
expression. Blood. 108:3913–3915. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Marhäll A, Kazi JU and Rönnstrand L: The
Src family kinase LCK cooperates with oncogenic FLT3/ITD in
cellular transformation. Sci Rep. 7:137342017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rousset D, Agnès F, Lachaume P, André C
and Galibert F: Molecular evolution of the genes encoding receptor
tyrosine kinase with immunoglobulinlike domains. J Mol Evol.
41:421–429. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Higgins DF, Ewart LM, Masterson E, Tennant
S, Grebnev G, Prunotto M, Pomposiello S, Conde-Knape K, Martin FM
and Godson C: BMP7-induced-Pten inhibits Akt and prevents renal
fibrosis. Biochim Biophys Acta. 1863:3095–3104. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Anticevic D, Jelic M and Vukicevic S:
Treatment of a congenital pseudarthrosis of the tibia by osteogenic
protein-1 (bone morphogenetic protein-7): A case report. J Pediatr
Orthop B. 15:220–221. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Honda Y, Knutsen R, Strong DD, Sampath TK,
Baylink DJ and Mohan S: Osteogenic protein-1 stimulates mRNA levels
of BMP-6 and decreases mRNA levels of BMP-2 and −4 in human
osteosarcoma cells. Calcif Tissue Int. 60:297–301. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao Y, Scott A, Zhang P, Hao Y, Feng X,
Somasundaram S, Khalil AM, Willis J and Wang Z: Regulation of
paxillin-p130-PI3K-AKT signaling axis by Src and PTPRT impacts
colon tumorigenesis. Oncotarget. 8:48782–48793. 2017.PubMed/NCBI
|
|
42
|
Bourgonje AM, Verrijp K, Schepens JT,
Navis AC, Piepers JA, Palmen CB, van den Eijnden M, Hooft van
Huijsduijnen R, Wesseling P, Leenders WP and Hendriks WJ:
Comprehensive protein tyrosine phosphatase mRNA profiling
identifies new regulators in the progression of glioma. Acta
Neuropathol Commun. 4:962016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yeo L, Adlard N, Biehl M, Juarez M,
Smallie T, Snow M, Buckley CD, Raza K, Filer A and Scheel-Toellner
D: Expression of chemokines CXCL4 and CXCL7 by synovial macrophages
defines an early stage of rheumatoid arthritis. Ann Rheum Dis.
75:763–771. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sullam PM, Frank U, Yeaman MR, Täuber MG,
Bayer AS and Chambers HF: Effect of thrombocytopenia on the early
course of streptococcal endocarditis. J Infect Dis. 168:910–914.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Aziz H, Zaas A and Ginsburg GS: Peripheral
blood gene expression profiling for cardiovascular disease
assessment. Genomic Med. 1:105–112. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
De Plaen IG, Han XB, Liu X, Hsueh W, Ghosh
S and May MJ: Lipopolysaccharide induces CXCL2/macrophage
inflammatory protein-2 gene expression in enterocytes via NF-kappaB
activation: Independence from endogenous TNF-alpha and
platelet-activating factor. Immunology. 118:153–163. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sarvaiya PJ, Guo D, Ulasov I, Gabikian P
and Lesniak MS: Chemokines in tumor progression and metastasis.
Oncotarget. 4:2171–2185. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang HL, Anatelli F, Zhai QJ, Adley B,
Chuang ST and Yang XJ: Glypican-3 as a useful diagnostic marker
that distinguishes hepatocellular carcinoma from benign
hepatocellular mass lesions. Arch Pathol Lab Med. 132:1723–1728.
2008.PubMed/NCBI
|
|
49
|
Tretiakova M, Zynger DL, Luan C, Andeen
NK, Finn LS, Kocherginsky M, Teh BT and Yang XJ: Glypican 3
overexpression in primary and metastatic Wilms tumors. Virchows
Arch. 466:67–76. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mitra N, Banda K, Altheide TK, Schaffer L,
Johnson-Pais TL, Beuten J, Leach RJ, Angata T, Varki N and Varki A:
SIGLEC12, a human-specific segregating (pseudo)gene, encodes a
signaling molecule expressed in prostate carcinomas. J Biol Chem.
286:23003–23011. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Marballi K, Quinones MP, Jimenez F,
Escamilla MA, Raventós H, Soto-Bernardini MC, Ahuja SS and
Walss-Bass C: In vivo and in vitro genetic evidence of involvement
of neuregulin 1 in immune system dysregulation. J Mol Med.
88:1133–1141. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Girkin J, Hatchwell L, Foster P, Johnston
SL, Bartlett N, Collison A and Mattes J: CCL7 and IRF-7 mediate
hallmark inflammatory and IFN responses following rhinovirus 1b
infection. J Immunol. 194:4924–4930. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Than NG, Romero R, Xu Y, Erez O, Xu Z,
Bhatti G, Leavitt R, Chung TH, El-Azzamy H, LaJeunesse C, et al:
Evolutionary origins of the placental expression of chromosome 19
cluster galectins and their complex dysregulation in preeclampsia.
Placenta. 35:855–865. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Waghray M, Yalamanchili M, Dziubinski M,
Zeinali M, Erkkinen M, Yang H, Schradle KA, Urs S, Pasca Di
Magliano M, Welling TH, et al: GM-CSF mediates
mesenchymal-epithelial cross-talk in pancreatic cancer. Cancer
Discov. 6:886–899. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Akpa MM, Iglesias DM, Chu LL, Cybulsky M,
Bravi C and Goodyer PR: Wilms tumor suppressor, WT1, suppresses
epigenetic silencing of the β-catenin gene. J Biol Chem.
290:2279–2288. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Riboldi E, Daniele R, Parola C, Inforzato
A, Arnold PL, Bosisio D, Fremont DH, Bastone A, Colonna M and
Sozzani S: Human C-type lectin domain family 4, member C
(CLEC4C/BDCA-2/CD303) is a receptor for
asialo-galactosyl-oligosaccharides. J Biol Chem. 286:35329–35333.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Shi Q, Liu H, Han P, Li C, Wang Y, Wu W,
Zhu D, Amos CI, Fang S, Lee JE, et al: Genetic variants in WNT2B
and BTRC predict melanoma survival. J Invest Dermatol.
137:1749–1756. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lin SY, Li K, Stewart GS and Elledge SJ:
Human Claspin works with BRCA1 to both positively and negatively
regulate cell proliferation. Proc Natl Acad Sci USA. 101:pp.
6484–6489. 2004; View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Focarelli ML, Soza S, Mannini L, Paulis M,
Montecucco A and Musio A: Claspin inhibition leads to fragile site
expression. Genes Chromosomes Cancer. 48:1083–1090. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ohta K, Shigeishi H, Taki M, Nishi H,
Higashikawa K, Takechi M and Kamata N: Regulation of CXCL9/10/11 in
oral keratinocytes and fibroblasts. J Dent Res. 87:1160–1165. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sidhu NS, Schreiber K, Pröpper K, Becker
S, Usón I, Sheldrick GM, Gärtner J, Krätzner R and Steinfeld R:
Structure of sulfamidase provides insight into the molecular
pathology of mucopolysaccharidosis IIIA. Acta Crystallogr D Biol
Crystallogr. 70:1321–1335. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhang X, Wan JX, Ke ZP, Wang F, Chai HX
and Liu JQ: TMEM88, CCL14 and CLEC3B as prognostic biomarkers for
prognosis and palindromia of human hepatocellular carcinoma. Tumour
Biol. 39:10104283177089002017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Yu H, Gaochao Z, Huaquan W, Limin X,
Lijuan L, Huijuan J, Wei Z, Jinglian T and Zonghong S: GDF11 is
increased in patients with myelodysplastic syndrome. Int J Clin Exp
Pathol. 9:6031–6038. 2016.
|
|
64
|
Safaee M, Ivan ME, Oh MC, Oh T, Sayegh ET,
Kaur G, Sun MZ, Bloch O and Parsa AT: The role of epidermal growth
factor-like module containing mucin-like hormone receptor 2 in
human cancers. Oncol Rev. 8:2422014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Fernández-Vega I, García-Suárez O, García
B, Crespo A, Astudillo A and Quirós LM: Heparan sulfate
proteoglycans undergo differential expression alterations in right
sided colorectal cancer, depending on their metastatic character.
BMC Cancer. 15:7422015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Pelizzaro-Rocha KJ, de Jesus MB,
Ruela-de-Sousa RR, Nakamura CV, Reis FS, de Fátima A and
Ferreira-Halder CV: Calix[6]arene bypasses human pancreatic cancer
aggressiveness: Downregulation of receptor tyrosine kinases and
induction of cell death by reticulum stress and autophagy. Biochim
Biophys Acta. 1833:2856–2865. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Saovapakhiran A, D'Emanuele A, Attwood D
and Penny J: Surface modification of PAMAM dendrimers modulates the
mechanism of cellular internalization. Bioconjug Chem. 20:693–701.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Wright EM, Loo DD and Hirayama BA: Biology
of human sodium glucose transporters. Physiol Rev. 91:733–794.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Guo GF, Cai YC, Zhang B, Xu RH, Qiu HJ,
Xia LP, Jiang WQ, Hu PL, Chen XX, Zhou FF, et al: Overexpression of
SGLT1 and EGFR in colorectal cancer showing a correlation with the
prognosis. Med Oncol. 28 Suppl 1:S197–S203. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Parimi S, Barnes TJ, Callen DF and
Prestidge CA: Mechanistic insight into cell growth, internalization
and cytotoxicity of PAMAM dendrimers. Biomacromolecules.
11:382–389. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Crocker PR, McMillan SJ and Richards HE:
CD33-related siglecs as potential modulators of inflammatory
responses. Ann N Y Acad Sci. 1253:102–111. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Virgo P, Denning-Kendall PA,
Erickson-Miller CL, Singha S, Evely R, Hows JM and Freeman SD:
Identification of the CD33-related Siglec receptor, Siglec-5
(CD170), as a useful marker in both normal myelopoiesis and acute
myeloid leukaemias. Br J Haematol. 123:420–430. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Hudson SA, Herrmann H, Du J, Cox P, Haddad
el-B, Butler B, Crocker PR, Ackerman SJ, Valent P and Bochner BS:
Developmental, malignancy-related and cross-species analysis of
eosinophil, mast cell and basophil siglec-8 expression. J Clin
Immunol. 31:1045–1053. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Potapenko IO, Haakensen VD, Lüders T,
Helland A, Bukholm I, Sørlie T, Kristensen VN, Lingjaerde OC and
Børresen-Dale AL: Glycan gene expression signatures in normal and
malignant breast tissue; possible role in diagnosis and
progression. Mol Oncol. 4:98–118. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Guo H, Nagy T and Pierce M:
Post-translational glycoprotein modifications regulate colon cancer
stem cells and colon adenoma progression in Apcmin/+
mice through altered WNT receptor signaling. J Biol Chem.
289:31534–31549. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Radisky DC, Levy DD, Littlepage LE, Liu H,
Nelson CM, Fata JE, Leake D, Godden EL, Albertson DG, Nieto MA, et
al: Rac1b and reactive oxygen species mediate MMP-3-induced EMT and
genomic instability. Nature. 436:123–127. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ferrari G, Cook BD, Terushkin V, Pintucci
G and Mignatti P: Transforming growth factor-beta 1 (TGF-beta1)
induces angiogenesis through vascular endothelial growth factor
(VEGF)-mediated apoptosis. J Cell Physiol. 219:449–458. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Dings RP, Miller MC, Nesmelova I,
Astorgues-Xerri L, Kumar N, Serova M, Chen X, Raymond E, Hoye TR
and Mayo KH: Antitumor agent calixarene 0118 targets human
galectin-1 as an allosteric inhibitor of carbohydrate binding. J
Med Chem. 55:5121–5129. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Liu WC, Lin YL, Spearman M, Cheng PY,
Butler M and Wu SC: Influenza virus hemagglutinin glycoproteins
with different N-glycan patterns activate dendritic cells in vitro.
J Virol. 90:6085–6096. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Abiola M, Favier M,
Christodoulou-Vafeiadou E, Pichard AL, Martelly I and
Guillet-Deniau I: Activation of Wnt/beta-catenin signaling
increases insulin sensitivity through a reciprocal regulation of
Wnt10b and SREBP-1c in skeletal muscle cells. PLoS One.
4:e85092009. View Article : Google Scholar : PubMed/NCBI
|