Introduction
Lyme disease (LD) is a multisystem inflammatory,
tick-borne disease resulting from infection with Borrelia
burgdorferi (1,2). LD is present in >80 countries,
including China. A total of ~3,000,000 cases of LD are reported
each year worldwide, representing an incidence of ~0.111% (2,3). LD
seriously affects human health, in addition to economic
development. As a result, this widespread disease has been given
attention regarding its prevention and treatment by the World
Health Organization (3). Lyme
neuroborreliosis (LNB) is a principal manifestation of LD and is
caused by the inflammatory effects of the spirochete B.
burgdorferi on the nervous system. LNB causes extensive
neurological damage and encephalitic memory impairment, even
leading to dementia and personality disorders. Given the high
disability rate caused by the disease, researchers and clinicians
are giving more concentrated attention to LNB (4,5);
however, its pathogenesis remains incompletely understood. It is
generally accepted that LNB is caused by an autoimmune response
triggered by molecular mimicry (6,7).
Microglial cells, the resident macrophage cells
within the central nervous system (CNS), are important in
initiating an immune response to microbial products (8). As a previous study demonstrated, CNS
damage and inflammation may activate microglia (9). Reactive microglia produce various
cytokines and chemokines causing an acute inflammatory reaction,
which leads to neuronal damage and apoptosis. Microglia cells are
hypersensitive to CNS damage. These cells multiply rapidly, begin
to express major histocompatibility complex proteins, migrate and
subsequently differentiate into phagocytes that secrete cytokines
and other toxic substances (10,11).
Previous studies have suggested that B.
burgdorferi basic membrane protein A (BmpA), one of the primary
B. burgdorferi pathogenic substances, exhibits a potent
pro-inflammatory effect (11–13).
In the authors' previous study, an Escherichia coli
expression system was established and purified recombinant BmpA
(rBmpA) was successfully obtained. When stimulated with rBmpA,
murine microglia BV2 cells produced pro-inflammatory chemokines,
including C-X-C motif chemokine 2 (CXCL2), C-C motif chemokine
(CCL) 5 and CCL22, causing inflammation and damage in mice
(13). However, the signal
transduction mechanism involved is unclear (13).
The nuclear factor κ-B (NF-κB) is A protein complex
which was found to specifically bind to the κB sequence
(GGGACTTTCC) of the immunoglobulin K enhancer and regulated the
expression of target genes (14–16).
NF-κB has received widespread interest from researchers for its
crucial role in the immune system. It has been demonstrated in
unstimulated cells that inhibitor of NF-κB (IκB-β) combines with
NF-κB and maintains NF-κB an inactive state. The stimuli that cause
NF-κB activation target IκB-β for degradation via a
phosphorylation-dependent ubiquitination process (16,17).
With the degradation of IκB-β, NF-κB is activated, entering the
cell nucleus where it encounters a promoter with DNA binding sites
for NF-κB (17,18). The activation of target gene
transcription induces the expression of specific mRNAs, and the
production of cytokines and chemokines, thereby regulating the
activation, multiplication, infiltration, chemotaxis and secretion
of immune cells (18–20). As rBmpA stimulates the production
of CXCL and CCL chemokines in microglia, it was hypothesized that
NF-κB may be closely associated with rBmpA-induced chemokine
production in BV2 cells (13).
Based on the conclusions of the authors' previous
study demonstrating that rBmpA stimulates the production of
inflammatory chemokines in BV2 cells, the present study continued
to investigate the signal transduction mechanism of LNB (13). Additionally, the present study
investigated the role of NF-κB in the signaling pathway of
rBmpA-induced inflammatory chemokines and provides a scientific
basis for the prevention and treatment of LBN.
Materials and methods
rBmpA preparation
Recombinant BmpA proteins were produced in E.
coli using the bacterial expression vector pGEX-6P1 (GE
Healthcare, Chicago, IL, USA). Expression, purification and
enzymatic cleavage of the glutathione S-transferase fusion proteins
were performed as previously described (14,15).
Cell culture and groups
BV2 cells (Kunming Medical University Biological
Engineer Center, Kunming, China) were cultured in Dulbecco's
modified Eagle's medium (DMEM)/high glucose with 5% cattle serum
(CS; HyClone, Logan, UT, USA), 1% penicillin/streptomycin (Sangon
Biotech Co., Ltd., Shanghai, China) at 37°C with 5% CO2.
BV2 cells were seeded into a 6-well plate at a concentration of
3×105 cells/ml. Supernatants were discarded until the
cells fully adhered to the plate. Cells were divided into three
groups (negative control group, positive control group and
experimental group), and were stimulated with 2 ml/well PBS with 5%
CS-DMEM medium, 1 µg/ml lipopolysaccharide (LPS; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany), and 20 µg/ml rBmpA, respectively.
Radioimmunoprecipitation assay buffer (high; Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) was used to
lyse the cells and cell lysates were collected at 12, 24 and 48 h
following stimulation.
ELISA analysis
Culture supernatants were collected at 6, 12 and 24
h following stimulation with rBmpA and were analyzed by ELISA.
Mouse CXCL2 (MIP-2), mouse CCL22 (MDC) and mouse CCL5 (RANTES)
ELISA kits (RayBiotech, Inc., Norcross, GA, USA; cat. nos. P10889,
O88430 and P30882) was used following manufacturer's protocol. A
Bio-Rad microplate reader (Model 680; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) was used to measure the absorbance at 450
nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Harvested cells were lysed with RNAiso Plus (Takara
Biotechnology Co., Ltd., Dalian, China) and RNA was extracted,
following the manufacturer's protocol. Total RNA was reverse
transcribed to cDNA. The RT-qPCR was performed using the real-time
PCR System Cfx-Connect (Bio-Rad Laboratories, Inc.). The reaction
conditions of the RT-qPCR were as follows: Denaturation at 95°C for
30 min, annealing at 58°C for 1 h and extension at 65°C for 5 sec.
The total volume used in the PCR was 25 µl, including 2 µl cDNA,
12.5 µl SYBR (Takara Biotechnology, Co., Ltd.), 8.5 µl
dH2O, 1 µl forward primer and 1 µl reverse primer
(Tsingke Biotech Co., Ltd., Kunming, China). The sequences of the
specific primers used to amplify NF-κB p65 and GAPDH were as
follows: NF-κB p65 forward, 5′-GCTACACAGAGGCCATTGAA-3′ and reverse,
5′-TCCCGGAGTTCATCTATGTG-3′; IκB-β forward,
5′-GGGAACGTCAGTCTGTACCA-3′ and reverse, 5′-GCACCATCGCTCTCTGTTTT-3′;
GAPDH forward, 5′-TCCCAGAGCTGAACGGGAAG-3′ and reverse,
5′-TCAGTGGGCCCTCAGATGC-3′. The gene expression level was calculated
using the method of 2−ΔΔCq (21–23).
Western blot analysis
Total protein quantitation analysis was used as the
internal control for quantitative fluorescent western blot analysis
(24–27). Cells were lysed in
radioimmunoprecipitation lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) with 1 mM phenylmethylsulfonyl
fluoride (Beyotime Institute of Biotechnology), and quantified
using a bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology). Proteins were separated on 10% TGX FastCast
acrylamide gels (Bio-Rad Laboratories, Inc.) with a loading mass of
20 µg total protein, then subsequently transferred to a
polyvinylidene difluoride (PVDF) membranes (0.2 µm; Bio-Rad
Laboratories, Inc.) using Trans-Blot Turbo Transfer System (p65, 15
V in 4-min protocol; IκB-β, 15 V in 4-min protocol; Bio-Rad
Laboratories, Inc.) with Trans-Blot Turbo Midi Transfer Packs
(Bio-Rad Laboratories, Inc.). The PVDF membranes were blocked with
5% skim milk at 37°C for 2 h. Blots were incubated with specific
primary antibodies of NF-κB p65 (cat. no. ab16502; 1:2,000; Abcam,
Cambridge, UK) or IκB-β (cat. no. ab7574; 1:1,000; Abcam) overnight
at 4°C, and subsequently incubated with goat anti-rabbit secondary
antibodies (cat. no. BS13278; p65 1:10,000; Nanjing Bioworld
Biotech Co., Ltd., Nanjing, China; cat. no. ab150077; IκB-β;
1:3,000; Abcam) at room temperature for 2 h. The blots were washed
three times between the primary and secondary antibody incubations.
The immunoreactive bands were imaged using an infrared imaging
system (Bio-Rad Laboratories, Inc.). ChemiDoc™
XRS+Systems with Image Lab™ software (Bio-Rad
Laboratories, Inc.) was used to analyze the western blot results of
NF-κB p65 and IκB-β.
Statistical analysis
The results are expressed as the mean ± standard
error of the mean and were analyzed using the GraphPad Prism 6
software package (GraphPad Software, Inc., La Jolla, CA, USA)
(26). Two-way analysis of
variance was used to calculate the P-values, multiple comparison
between the groups was performed using the Student-Newman-Keuls
method. P<0.05 was considered to indicate a statistically
significant difference.
Results
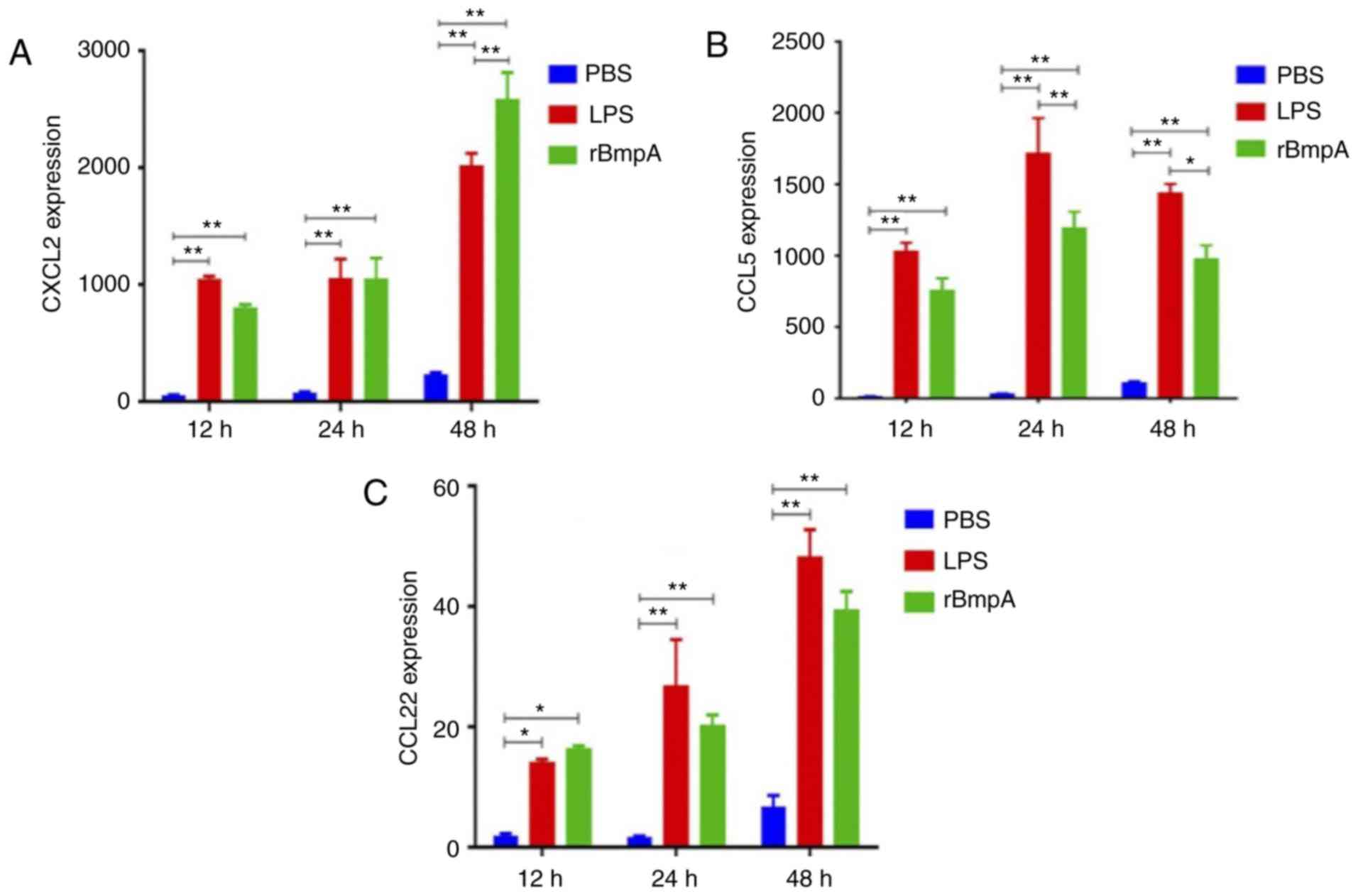
rBmpA significantly stimulates the
production of the inflammatory cytokines CXCL2, CCL5 and CCL22 in
BV2 cells
To determine whether rBmpA induces the production of
pro-inflammatory chemokines in BV2 cells, analysis of CXCL2, CCL5
and CCL22 concentrations in the cell supernatants was assayed by
ELISA. The results demonstrated that rBmpA significantly increased
CXCL2, CCL5 and CCL22 expression at all time points tested compared
with the PBS only group (P<0.05; Fig. 1). The expression of the three
chemokines demonstrated an increasing trend between 12 and 48 h,
except for a slight decline in CCL5 between 24 and 48 h (Fig. 1B).
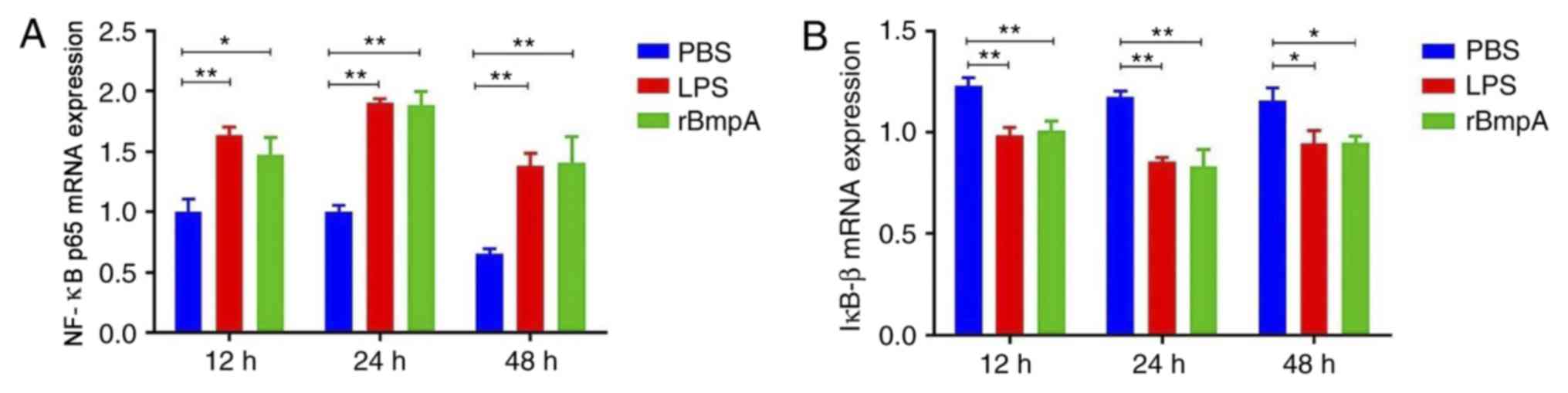
rBmpA stimulation induces an increase
in NF-κB and decrease in IκB-bmRNA in BV2 cells
rBmpA stimulation significantly increased CXCL2,
CCL5 and CCL22 expression compared with the PBS control
(P<0.05). Therefore, the rBmpA-induced nuclear translocation of
NF-κB p65 and degradation of cytosolic IκB-β was investigated.
RT-qPCR was used to analyze the expression of NF-κB p65 and IκB-β
mRNA. As exhibited in Fig. 2, the
expression of NF-κB mRNA in the group treated with rBmpA was
significantly increased (P<0.01) compared with the PBS-only
control at 12, 24 and 48 h and rBmpA treatment significantly
decreased the intracellular levels of IκB-β (P<0.05).
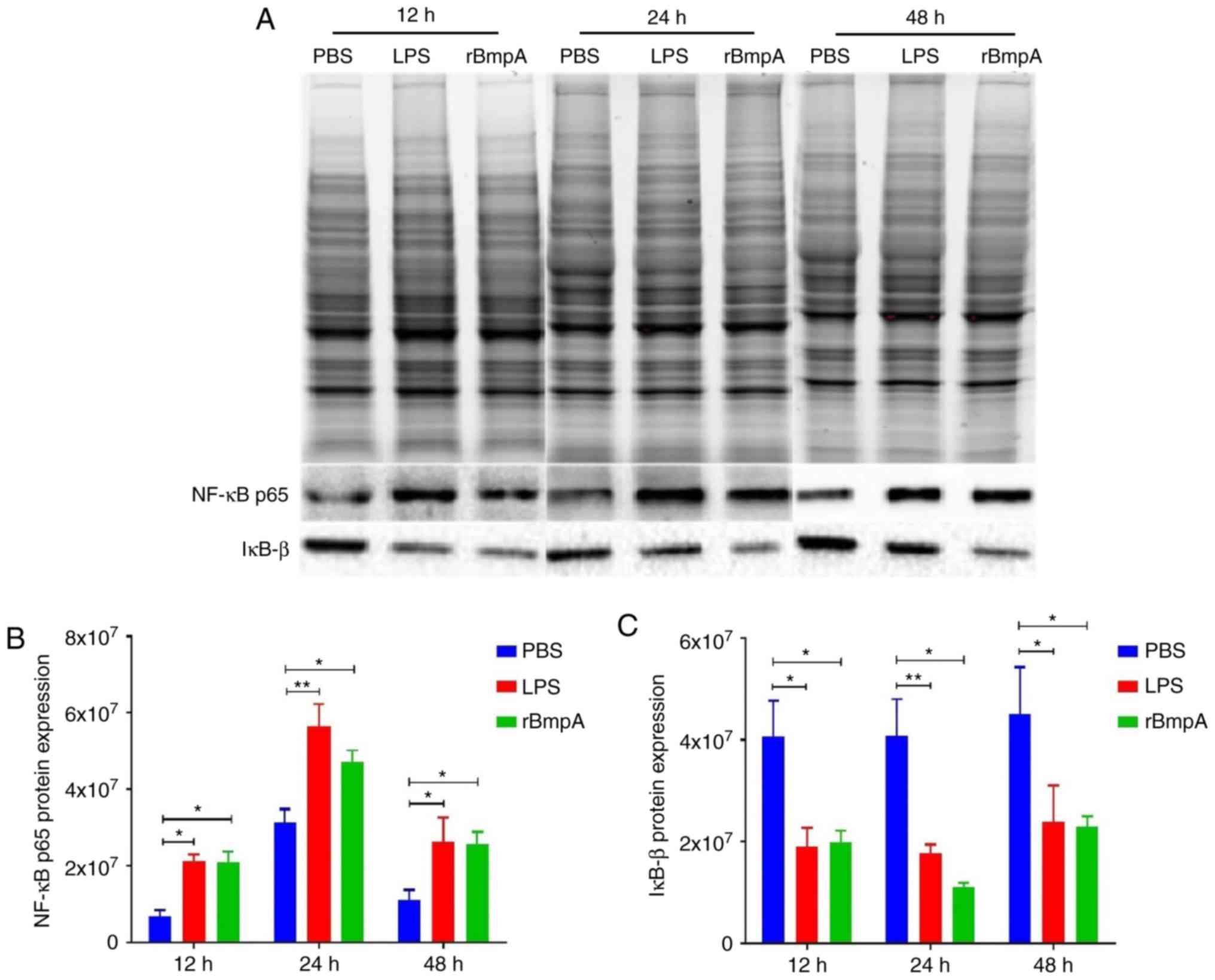
rBmpA stimulation induces an increase
in NF-κB p65 and the degradation of IκB-bproteins in BV2 cells
Western blot analysis was used to detect the protein
expression levels of NF-κB p65 and IκB-β in BV2 cells. As exhibited
in Fig. 3, following stimulation
with rBmpA for 12, 24 and 48 h, NF-κB p65 protein expression was
significantly increased (P<0.05) and the protein level of IκB-β
was significantly decreased (P<0.05) in the rBmpA treatment
group compared with the PBS-only control. Increased expression
levels of NF-κB p65 protein were observed in the rBmpA and LPS
treatment groups. rBmpA significantly decreased the intracellular
levels of IκB-β protein compared with the PBS control
(P<0.05).
Discussion
NF-κB is a nuclear transcription factor which
regulates the immune response, stress response and apoptosis
(28). Upregulation of NF-κB has
been demonstrated in various tumors, pulmonary disease and hepatic
disease. Previous in vitro studies have suggested that NF-κB
is a key signal transduction molecule in the downstream pathway of
Toll-like receptors (TLRs) (29,30).
Once the TLRs are stimulated by various pathogenic factors, the
degradation of IκB-β is initiated and NF-κB is free to enter the
cell nucleus, where NF-κB combines with the IκB motif controlling
the transcription of a number of cytokines, including tumor
necrosis factor-α (TNF-α), CXCL13 and interleukin (IL)-6. Studies
have demonstrated that NF-κB is a transducer of various common
inflammatory signaling pathway reactions (27–30).
Therefore, methods of inhibiting NF-κB signaling have potential
therapeutic applications in inflammatory diseases. At present, a
number of anti-inflammatory drugs and anti-rheumatic drugs,
including corticosteroids and aspirin, have been confirmed to be
inhibitors of NF-κB activation (31,32).
It has been reported that the B. burgdorferi
bmpA/B gene operon exhibits the most marked upregulation in
mouse and human joints (33). BmpA
possesses stimulatory activity through functional domains that
trigger the inflammatory response. For example, the activated NF-κB
kinase signalling pathway in articular synovial cells was
demonstrated to produce pro-inflammatory cytokines including TNF-α
and IL-1β, triggering prostaglandin E2 receptor EP4 subtype (PGE-2)
(15,34). Rasley et al (35) confirmed that B. burgdorferi
was an important stimulus that induced microglia to produce IL-6,
TNF-α and PGE-2, which was associated with increased expression of
NF-κB, TLR2 and cluster of differentiation 14 in BV2 cells. By
activating signaling molecules, B. burgdorferi stimulated
immune cells to produce multiple inflammatory substances (35). Further investigation by Sun et
al (36) suggested that
substance P was able to prompt B. burgdorferi-induced NF-κB
activation by upregulating NF-κB subunit p65, causing a significant
increase in the production of inflammatory cytokines (36). NF-κB serves an important role in
the inflammatory cytokine signaling pathway of immune cells.
However, rBmpA and NF-κB associated pathogenesis in LNB, to the
best of the authors' knowledge, has not been reported in the
literature. A recent report indicated that rBmpA induced activation
of BV2 cells with a concentration-dependent secretion of
inflammatory chemokines (13).
Therefore, rBmpA may be associated with the inflammatory chemokines
produced by BV2 cells and the occurrence of LNB. However, the
precise mechanism of the signal transduction pathway remains
uncertain.
In the present study, whether NF-κB was a key
modulator in the inflammatory chemokines (CXCL2, CCL5 and CCL22)
signaling pathway stimulated by rBmpA in BV2 cells was
investigated. The results of the present study demonstrated that
CXCL2, CCL5, CCL22, NF-κB mRNA and NF-κB protein increased and the
protein and mRNA levels of IκB-β decreased following stimulation
with rBmpA in BV2 cells. It was demonstrated that NF-κB served a
key role in the signaling pathways stimulated by rBmpA in BV2
cells, resulting in the production of various inflammatory
chemokines. This result increases the understanding of the
pathogenesis of LNB. Future studies will investigate the key
proteins in inflammatory chemokine signaling pathways stimulated by
rBmpA and, most importantly, investigate whether these signaling
pathways also modulate the pathogenesis of Lyme arthritis,
dermatitis and carditis. The long-term aim is to investigate
whether preventive and therapeutic medicines for LNB may be
developed that target these proteins (37).
In conclusion, following stimulation by rBmpA, BV2
cells overexpressed NF-κB and exhibited significantly reduced
expression of IκB-β in the inflammatory cytokine-signaling pathway.
This result suggested that NF-κB is important in the inflammatory
cytokines signaling pathways stimulated by rBmpA and may be
associated with the occurrence of LNB. The present study further
clarified the mechanism underlying the rBmpA-induced inflammatory
chemokines signaling pathway in microglial cells, and provides a
scientific basis for the prevention and treatment of LNB.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81560596 and
3156005) and the Natural Foundation of Yunnan Province (grant nos.
2012FB011, 2014FA011, 2014FB001 and 2017FE467-001).
References
|
1
|
Borchers AT, Keen CL, Huntley AC and
Gershwin ME: Lyme disease: A rigorous review of diagnostic criteria
and treatment. J Autoimmun. 57:82–115. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pal U, Wang P, Bao F, Yang X, Samanta S,
Schoen R, Wormser GP, Schwartz I and Fikrig E: Borrelia burgdorferi
basic membrane proteins A and B participate in the genesis of Lyme
arthritis. J Exp Med. 205:133–141. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ramamoorthi N, Narasimhan S, Pal U, Bao F,
Yang XF, Fish D, Anguita J, Norgard MV, Kantor FS, Anderson JF, et
al: The Lyme disease agent exploits a tick protein to infect the
mammalian host. Nature. 436:573–577. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lantos PM, Shapiro ED, Auwaerter PG, Baker
PJ, Halperin JJ, McSweegan E and Wormser GP: Unorthodox alternative
therapies marketed to treat Lyme disease. Clin Inf Dis.
60:1776–1782. 2015. View Article : Google Scholar
|
|
5
|
Marques AR: Lyme neuroborreliosis.
Continuum (Minneap Minn). 21(6 Neuroinfectious Disease): 1–1744.
2015.
|
|
6
|
Bremell D and Dotevall L: Oral doxycycline
for Lyme neuroborreliosis with symptoms of encephalitis, myelitis,
vasculitis or intracranial hypertension. Eur J Neurol.
21:1162–1167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cerar T, Ogrinc K, Lotričfurlan S,
Lotric-Furlan S, Kobal J, Levicnik-Stezinar S, Strle F and
Ruzić-Sabljic E: Diagnostic value of cytokines and chemokines in
Lyme neuroborreliosis. Clin Vaccine Immunol. 20:1578–1584. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim C, Cho ED, Kim HK, You S, Lee HJ,
Hwang D and Lee SJ: β1-integrin-dependent migration of microglia in
response to neuron-released α-synuclein. Exp Mol Med. 46:e912014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Togna AR, Latina V, Trefiletti G, Guiso M,
Moschini S and Togna GI: 1-Phenil-6,7-dihydroxy-isochroman inhibits
inflammatory activation of microglia. Brain Res Bull. 95:33–39.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brissette CA, Houdek HM, Floden AM and
Rosenberger TA: Acetate supplementation reduces microglia
activation and brain interleukin-1β levels in a rat model of Lyme
neuroborreliosis. J Neuroinflammation. 9:2492012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuhlow CJ, Garcia-Monco JC, Coleman JL and
Benach JL: Murine microglia are effective phagocytes for Borrelia
burgdorferi. J Neuroimmunol. 168:183–187. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Battisti JM, Bono JL, Rosa PA, Schrumpf
ME, Schwan TG and Policastro PF: Outer surface protein A protects
Lyme disease spirochetes from acquired host immunity in the tick
vector. Infect Immun. 76:5228–5237. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao H, Liu A, Cui Y, Liang Z, Li B and
Bao F: Borrelia burgdorferi basic membrane protein A could induce
chemokine production in murine microglia cell line BV2. Microb
Pathog. 111:174–181. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Simpson WJ, Schrumpf ME and Schwan TG:
Reactivity of human Lyme borreliosis sera with a 39-kilodalton
antigen specific to Borrelia burgdorferi. J Clin Microbiol.
28:1329–1337. 1990.PubMed/NCBI
|
|
15
|
Yang X, Izadi H, Coleman AS, Wang P, Ma Y,
Fikrig E, Anguita J and Pal U: Borrelia burgdorferi lipoprotein
BmpA activates pro-inflammatory responses in human synovial cells
through a protein moiety. Microbes Infect. 10:1300–1308. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baldwin AS Jr: The NF-kappa B and I kappa
B proteins: New discoveries and insights. Annu Rev Immunol.
14:649–683. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Whiteside ST, Epinat JC, Rice NR and
Israël A: I kappa B epsilon, a novel member of the I kappa B
family, controls RelA and cRel NF-kappa B activity. EMBO J.
16:1413–1426. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bell S, Degitz K, Quirling M, Jilg N, Page
S and Brand K: Involvement of NF-kappaB signaling in skin
physiology and disease. Cell Signal. 15:1–7. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vinayagamoorthi R, Koner BC, Kavitha S,
Nandakumar DN, Padma Priya P and Goswami K: Potentiation of humoral
immune response and activation of NF-kappaB pathway in lymphocytes
in experimentally induced hyperthyroid rats. Cell Immunol.
238:56–60. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Colleran A, Collins PE and Carmody RJ:
Assessing Sites of NF-κB DNA binding using chromatin
immunoprecipitation. Methods Mol Biol. 1280:47–59. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Leal VO, Saldanha JF, Stockler-Pinto MB,
Cardozo LF, Santos FR, Albuquerque AS, Leite M Jr and Mafra D: NRF2
and NF-κB mRNA expression in chronic kidney disease: A focus on
nondialysis patients. Int Urol Nephrol. 47:1985–1991. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Naranjo V, Ayllón N, Pérez de la Lastra
JM, Galindo RC, Kocan KM, Blouin EF, Mitra R, Alberdi P, Villar M
and de la Fuente J: Reciprocal regulation of NF-kB (Relish) and
subolesin in the tick vector, Ixodes scapularis. PLoS One.
8:e659152013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Irvine M, Okitsu C and Hsieh CL: Q-PCR in
combination with ChIP assays to detect changes in chromatin
acetylation. Methods Mol Biol. 791:213–223. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Eaton SL, Roche SL, Llavero Hurtado M,
Oldknow KJ, Farquharson C, Gillingwater TH and Wishart TM: Total
protein analysis as a reliable loading control for quantitative
fluorescent western blotting. PLoS One. 8:e724572013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
G5: ding Control for Quantitative
Fluorescent Western Blotting. Plos One. 8:e724572013.238: 56–605,
2097. and brain interleukin-1β lev View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vigelsø A, Dybboe R, Hansen CN, Dela F,
Helge JW and Guadalupe Grau A: GAPDH and β-actin protein decreases
with aging, making stain-free technology a superior loading control
in Western blotting of human skeletal muscle. J Appl Physiol
(1985). 118:386–394. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rivero-Gutiérrez B, Anzola A,
Martínez-Augustin O and de Medina FS: Stain-free detection as
loading control alternative to Ponceau and housekeeping protein
immunodetection in Western blotting. Anal Biochem. 467:1–3. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ruzehaji N, Avouac J, Elhai M, Frechet M,
Frantz C, Ruiz B, Distler JH and Allanore Y: Combined effect of
genetic background and gender in a mouse model of bleomycin-induced
skin fibrosis. Arthritis Res Ther. 17:1452015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peri S, Devarajan K, Yang DH, Knudson AG
and Balachandran S: Meta-analysis Identifies NF-κB as a therapeutic
target in renal cancer. PLoS One. 8:e767462013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Murqueitio MS, Ebner S, Hörtnagl P, Rakers
C, Bruckner R, Henneke P, Wolber G and Santos-Sierra S: Enhanced
immunostimulatory activity of in silico discovered agonists of
Toll-like receptor 2 (TLR2). Biochim Biophys Acta. 1861:2680–2689.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ottonello L, Bertolotto M, Montecucco F,
Bianchi G and Dallegri F: Delayed apoptosis of human monocytes
exposed to immune complexes is reversed by oxaprozin: Role of the
Akt/IkappaB kinase/nuclear factor kappaB pathway. Br J Pharmacol.
157:294–306. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Carvalho L, Jacinto A and Matova N: The
toll/NF-κB signaling pathway is required for epidermal wound repair
in Drosophila. Proc Natl Acad Sci USA. 111:pp. E5373–E5382. 2014;
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Baum E, Hue F and Barbour AG: Experimental
infections of the reservoir species Peromyscus leucopus with
diverse strains of Borrelia burgdorferi, a Lyme disease agent.
MBio. 3:e00434–12. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bernardino AL, Myers TA, Alvarez X,
Hasegawa A and Philipp MT: Toll-like receptors: Insights into their
possible role in the pathogenesis of lyme neuroborreliosis. Infect
Immun. 76:4385–4395. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rasley A, Marriott I, Halberstadt CR, Bost
KL and Anguita J: Substance P augments Borrelia burgdorferi-induced
prostaglandin E2 production by murine microglia. J Immunol.
172:5707–5713. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sun J, Ramnath RD, Zhi L, Tamizhselvi R
and Bhatia M: Substance P enhances NF-kappaB transactivation and
chemokine response in murine macrophages via ERK1/2 and p38 MAPK
signaling pathways. Am J Physiol Cell Physiol. 294:C1586–C1596.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sadik CD, Hunfeld KP, Bachmann M, Kraiczy
P, Eberhardt W, Brade V, Pfeilschifter J and Mühl H: Systematic
analysis highlights the key role of TLR2/NF-kappaB/MAP kinase
signaling for IL-8 induction by macrophage-like THP-1 cells under
influence of Borrelia burgdorferi lysates. Int J of Biochem Cell
Biol. 40:2508–2521. 2008. View Article : Google Scholar
|