Introduction
Ischemic heart disease (IHD) greatly threatens human
health leading to arrhythmia, heart failure and even mortality
(1). Coronary artery stenosis is
the main cause of ischemic heart disease, and cardiopulmonary
bypass-caused arrest also induces ischemic heart disease (2). The treatment of ischemic heart
disease relies on the recovery of blood supply, but the restored
blood supply may trigger myocardial injury, termed
ischemia-reperfusion injury (IRI), which contributes to a poor
prognosis in IHD patients.
The commonly used drugs for the prevention and
treatment of myocardial ischemia include calcium antagonists,
β-receptor blockers and angiotensin-converting enzyme inhibitors,
but they are not ideal due to their side effects and safety and
ethical issues (3). A previous
study demonstrated that the antioxidant N(2)-L-alanyl-L-glutamine
(NLAG) is a precursor of intracellular glutathione and has been
proved by the US Food and Drug Administration to treat
acetaminophen-induced acute liver failure (4). NLAG can directly scavenge,
significantly improve the anti-apoptotic ability, and reduce the
release of inflammatory factors, chemokines and adhesion molecules
(5,6). In addition, NLAG has a protective
effect on the kidney and liver against IRI (7,8).
However, any protective effects and the underlying mechanism of
NLAG on the myocardial IRI remain unclear.
Protein tyrosine kinase/signal transducer and
activator of transcription (JAK/STAT) is a signaling transduction
pathway that is closely associated with inflammation (9), oxidative stress (10), cell injury and apoptosis (11). Janus activated kinase signal
transducer 2 and activator of transcription 3 (JAK2/STAT3) is an
important signal transduction pathway in the JAK/STAT family.
Ischemic preconditioning, ischemic post-conditioning and
anti-myocardial IRI effects of some drugs are associated with
JAK2/STAT3 pathway activation (12,13).
The present study established IRI model to investigate the
protective effect of NLAG on myocardial IRI, and to observe the
effect of NLAG on JAK2/STAT3 signaling pathway-associated
molecules.
Materials and methods
Animals and ethical approval
Adult male Wistar rats (350–450 g; n=30) were
obtained from Military Medical Science Academy of the People's
Liberation Army (Beijing, China). Animals were housed at a constant
temperature (22±1°C), with 50% humidity, and a 12-h light-dark
cycle. The rats had ad libitum access to food and autoclaved
water. All animal procedures were approved by the Animal
Experiments Ethics Committee of the Military Medical Science
Academy of the People's Liberation Army (Beijing, China).
Establishment of the IRI model
Myocardial IRI model was established as has
previously been described (14).
Rats were anesthetized with intraperitoneal injection of 2%
pentobarbital sodium (cat. no. 57-33-0; 0.2 ml/100 g;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) the tracheotomy was
performed between the third and fourth cartilage rings, and rats
received mechanical ventilation. The left anterior descending
coronary artery (LAD) was ligated by a 7/0 thread inserting 32 mm
below the left auricle root, crossing the myocardium and suturing
below the pulmonary artery cone. Both ends of the thread passed
through the polyethylene tubule (epidural catheter), which reached
the ventricular wall to block coronary blood flow by tightening the
ends of the thread. Following that, the tube was clamped with
hemostatic forceps. Electrocardiography revealed ST segment
elevation and the myocardial tissue below ligature site became
darker; following this, hemostatic forceps were released and the
LAD blood flow was restored, elevated ST segment was reduced to
above 1/2 and the myocardial tissue became gradually red.
Groups and treatments
Rats were randomly divided into three groups: Sham
operation (Sham group; n=10), myocardial ischemia reperfusion (IRI
group; n=10) and NLAG treatment (NLAG group; n=10). In the Sham
group, rats received the tracheotomy alone. In the IRI group, the
IRI model was established. In the NLAG group, rats were injected
with 150 mg/kg NLAG (Chongqing Laimei Pharmaceutical Co., Ltd.,
Chongqing, China) intraperitoneally 30 min prior to IRI
establishment.
Cardiac hemodynamic changes
The hemodynamic parameters of the heart were
recorded using the Datex-Ohmeda S/5 Entropy Module (DRE, Inc.,
Louisville, KY, USA). Left ventricular diastolic pressure (LVDP),
heart rate (HR) and the maximum rate of left ventricular pressure
(+dP/dtmax) were recorded before ischemia, at 15, 30, 45 and 60 min
following reperfusion (HR was recorded every 15 min).
Preparation and treatment of rat
tissues
All rats were anesthetized with pentobarbital sodium
(cat. no. 57-33-0; Sigma-Aldrich; Merck KGaA) at 4 h following IRI.
Blood samples (3 ml) were taken from the internal jugular vein and
permitted to clot overnight at 4°C prior to centrifugation for 15
min at 1,000 × g at 4°C. Serum aliquots were then removed and
samples were incubated at −20°C or −80°C. The rat myocardial
tissues were collected and fixed in neutral formalin or stored in
liquid nitrogen.
Hematoxylin-eosin (HE) staining
Heart tissues were fixed in 10% formaldehyde for 24
h at room temperature (pH=7.2; cat. no. G2161; Beijing Solarbio
Science & Technology, Co., Ltd., Beijing, China), and then
decalcified, dehydrated, permeabilized using xylene (50% xylene for
1 h and 100% xylene for 2 h), embedded in wax and sliced into 5 µm
thick sections using a microtome. All of the following steps were
carried out at room temperature. Sections were then dewaxed using
xylene I for 15 min and then xylene II for 15 min, hydrated with
absolute ethanol for 5 min, 90% ethanol for 2 min and 70% ethanol
for 2 min, mounted with 10% hematoxylin (cat. no. G1120; Beijing
Solarbio Science & Technology, Co., Ltd.) for 10 min,
differentiated with 1% hydrochloric acid and ethanol for 3–5 sec,
stained with 0.5% eosin (cat. no. G1120; Beijing Solarbio Science
& Technology, Co., Ltd.) for 1 min, dehydrated in alcohol
gradients (70% for 2 sec, 90% for 2 min and absolute ethanol for 5
min) and xylene, cleared and then mounted. Using a light microscope
(magnification, ×200; Leica DM 4000B; Leica Microsystems, Inc.,
Buffalo Grove, IL, USA), pathological changes of myocardial tissues
were observed.
Masson staining
Heart tissues were fixed using 10% formaldehyde for
24 h at room temperature (pH=7.2; cat. no. G2161; Beijing Solarbio
Science & Technology, Co., Ltd.), decalcified, dehydrated,
permeabilized using xylene (50% xylene for 1 h and 100% xylene for
2 h), embedded in wax and then sliced into 5 µm thick sections
using a microtome. All the following steps were carried out at room
temperature. The tissues were routinely deparaffinized and
rehydrated in accordance. Weigert's hematoxylin (5%; cat. no.
G1340; Beijing Solarbio Science & Technology, Co., Ltd.) was
used to dye the cell nucleus for 5 min. Following rinsing with
distilled water three times, the sections were stained using 0.7%
Masson-Ponceau-acid fuchsin solution (cat. no. G1340; Beijing
Solarbio Science & Technology, Co., Ltd.) for 10 min. Samples
were then rinsed in 2% glacial acetic acid and differentiated in
phosphomolybdic acid for 4 min. The sections were directly stained
with 2% aniline blue dye solution (cat. no. G1340; Beijing Solarbio
Science & Technology, Co., Ltd.). Following dehydrating with
ethanol series, clearing with xylene and mounting with neutral
resins, digital images were captured using a light microscope
(magnification, ×200; Leica DM 4000B; Leica Microsystems, Inc.) to
analyze fiber formation in the early callus.
ELISA assay
Protein expression changes of markers of myocardial
injury [lactase dehydrogenase (LDH; cat. no. SEB864Ra; Uscn Life
Sciences, Inc., Wuhan, China), troponin I (cTnI; cat. no. SEA478Ra;
Uscn Life Sciences, Inc.), creatine kinase (CK; cat. no. SEA109Ra;
Uscn Life Sciences, Inc.) and heart type fatty acid binding protein
(hFABP; cat. no. SEB243Ra; Uscn Life Sciences, Inc.)], inflammatory
cytokines [interleukin (IL) 1, 6 (cat. nos. SEA563Ra and SEA079Ra,
respectfully; Uscn Life Sciences, Inc.) and tumor necrosis factor
(TNF)-α (cat. no. SEA133Ra; Uscn Life Sciences, Inc.)], and
oxidative stress factors [malondialdehyde (MDA; cat. no. CEA597Ge;
Uscn Life Sciences, Inc.) and sorbitol dehydrogenase (SDH; cat. no.
SEB495Ra; Uscn Life Sciences, Inc.)] were detected by ELISA kits.
The optical density (OD) at 450 nm was measured with microplate
reader.
Western blot analysis
A total of 100 mg myocardial tissue was homogenized
(IKA T10; IKA®-Werke GmbH & Co. KG, Breisgau,
Germany) and centrifuged at 12,000 × g for 15 min at 4°C.
Subsequently, the supernatant was collected and protein
quantification was performed by bicinchoninic acid assay, and equal
amounts of protein lysate (40 µg) were separated by 12% SDS-PAGE.
Transfer to nitrocellulose membranes was performed in transfer
buffer (12 mM Tris base, 96 mM glycine, pH 8.3, and 15% methanol).
Subsequently, the membranes were blocked for 2 h at room
temperature in TBS with 20% Tween 20 (TBST) buffer and probed with
rabbit anti rat/human/mouse/cow B-cell lyphoma (Bcl)-2 (1:1,000;
cat. no. ab59348; Abcam, Cambridge, UK), Bcl-associated X protein
(Bax; 1:1,000; cat. no. ab32503; Abcam), Caspase-3 (1:500; cat. no.
ab13847; Abcam), Janus kinase (JAK2; 1:5,000; cat. no. ab108596;
Abcam), phosphorylated (p)-JAK2 (1:1,000; cat. no. ab32101; Abcam),
signal transducer and activator of transcription (STAT) 3 (1:1,000;
cat. no. ab68153; Abcam), p-STAT3 antibodies (1:2,000; cat. no.
ab76315; Abcam), and a rabbit anti rat/human/mouse/monkey β-actin
antibody (monoclonal; 1:100; cat. no. 8457; Cell Signaling
Technology Inc., Danvers, USA) overnight at 4°C. The membranes were
washed with TBST buffer three times, followed by incubation with a
horseradish peroxidase-conjugated secondary antibody (monoclonal;
1:4,000; cat. no. HS101; Beijing Transgen Biotech Co., Ltd.,
Beijing, China) for 1 h at room temperature. Finally, the protein
expression levels were detected by using a Gel Doc™ EZ
System (cat. no. 1708270; Bio-Rad Laboratories, Inc., Hercules, CA,
USA), and densitometric analysis was then performed using Image
Lab™ software (version 6.0; Bio-Rad Laboratories,
Inc.).
Statistical analysis
All data are expressed as the mean ± standard
deviation. Analysis was performed using the GraphPad Prism
software, version 6.00 (GraphPad Software Inc., La Jolla, CA, USA).
Multiple comparisons were analyzed using one-way analysis of
variance followed by the Newman-Keuls post-hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of NLAG on hemodynamic
parameters
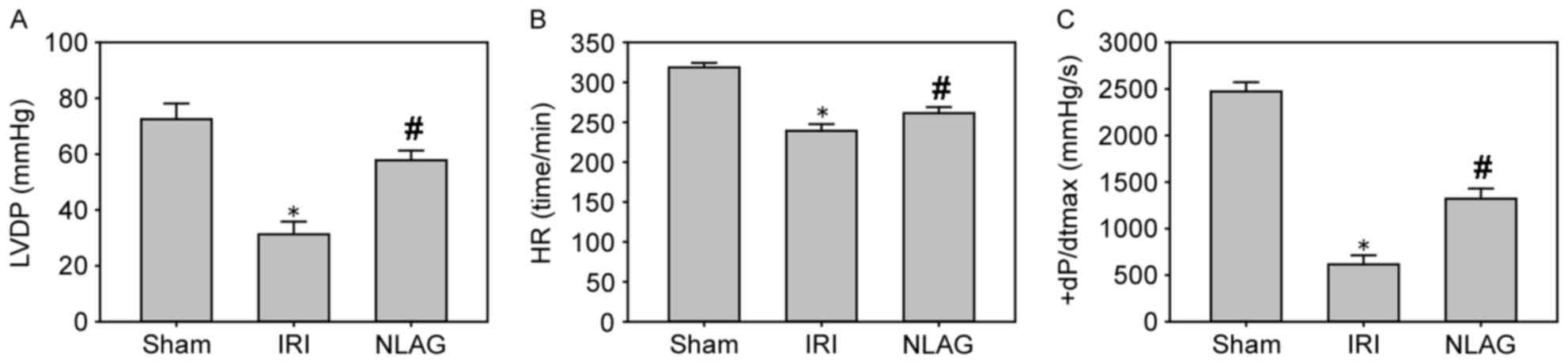
Compared with the sham group (LVDP, HR and +dP/dtmax
were 73.5±5.13 mmHg, 319.47±6.21 beats/min and 2,412.52±89.63
mmHg/sec, respectively), LVDP, HR and +dP/dtmax were significantly
decreased at 60 min following reperfusion in the IRI group (LVDP,
HR and +dP/dtmax were 31.3±4.53 mmHg, 239.17±8.45 beats/min and
615.17±98.23 mmHg/sec, respectively; P<0.05, Fig. 1). Furthermore, NLAG significantly
improved these levels (LVDP, HR and +dP/dtmax were 57.81±3.46 mmHg,
261.38±7.54 beats/min and 1320.15±110.12 mmHg/sec, respectively;
P<0.05, Fig. 1).
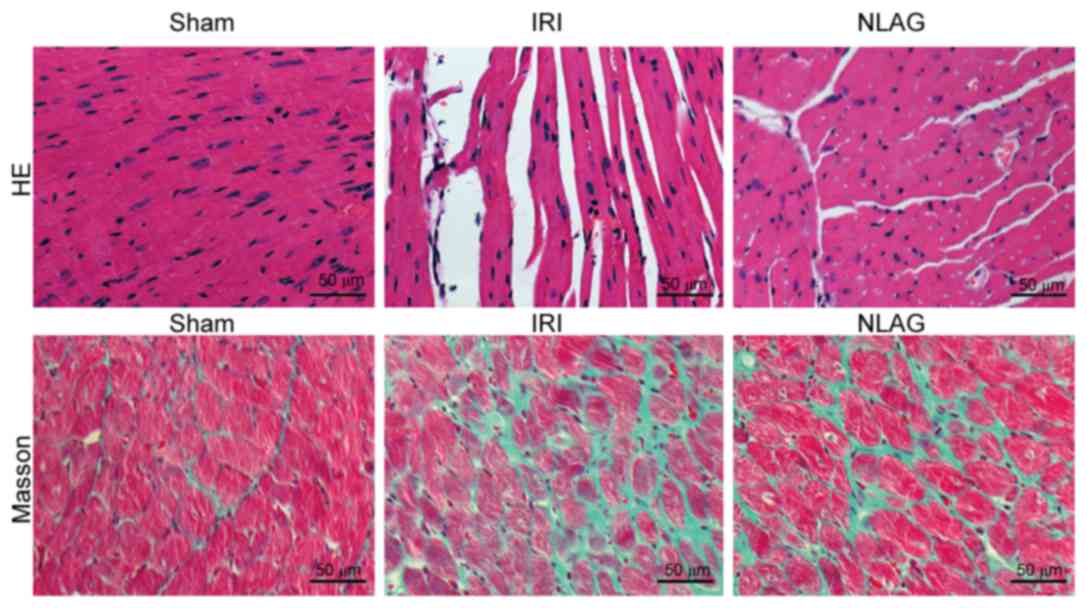
NLAG inhibits IRI-induced collagen
deposition in the heart
Myocardial cells were arranged orderly in the Sham
group, with clear boundaries and intact nuclei. On the contrary,
myocardial cells in the IRI group were arranged disorderly, with
unclear boundaries, myofiber rupture and nuclei disappearance. In
the NLAG group, the severity of myocardial injuries was improved to
a great extent (Fig. 2).
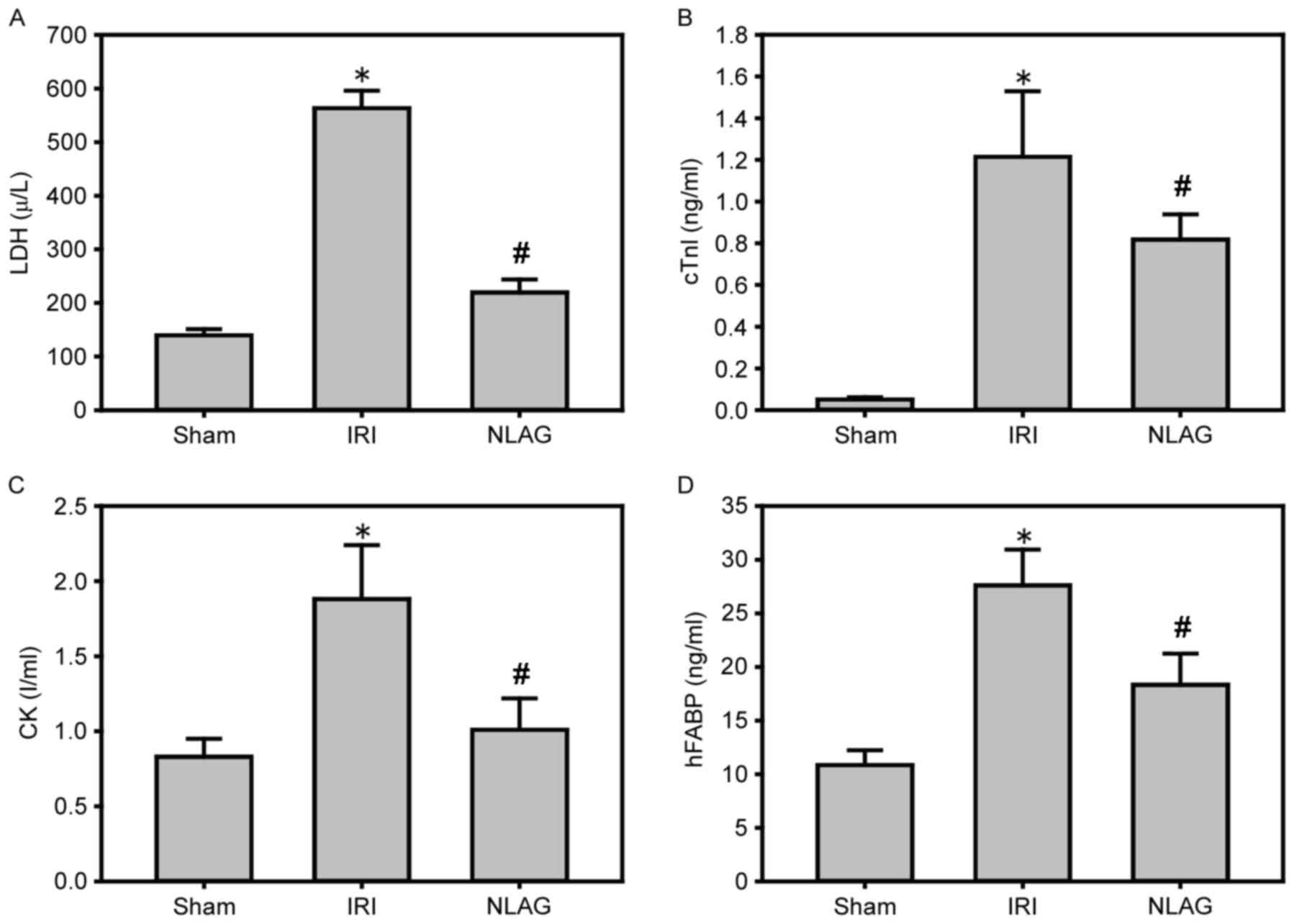
NLAG modulates IRI-induced protein
expression of cardiac enzymes
Compared with the Sham group, the protein levels of
LDH, cTnI, CK and hFABP in the plasma of the IRI group were
increased significantly (P<0.05). On the other hand, NLAG
effectively inhibited their expression levels, which was
statistically significant compared with the IRI group (P<0.05,
Fig. 3).
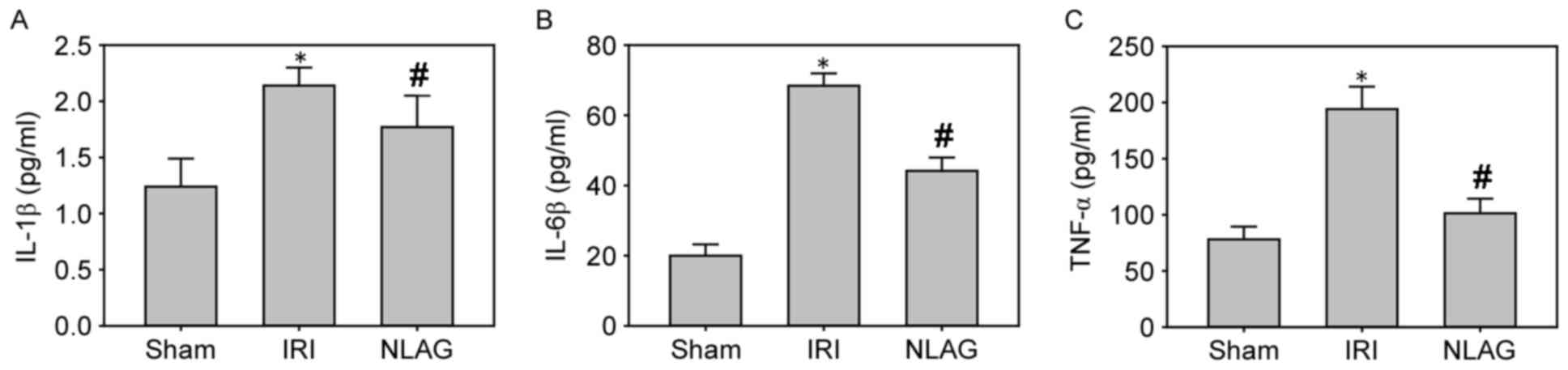
NLAG modulates IRI-induced protein
expression of inflammatory cytokines
Compared with the Sham group, the protein levels of
IL-1β, IL-6 and TNF-α in the plasma of the IRI group were increased
significantly (P<0.05). However, NLAG could effectively inhibit
the inflammatory response, which was statistically significant
compared with the IRI group (P<0.05, Fig. 4).
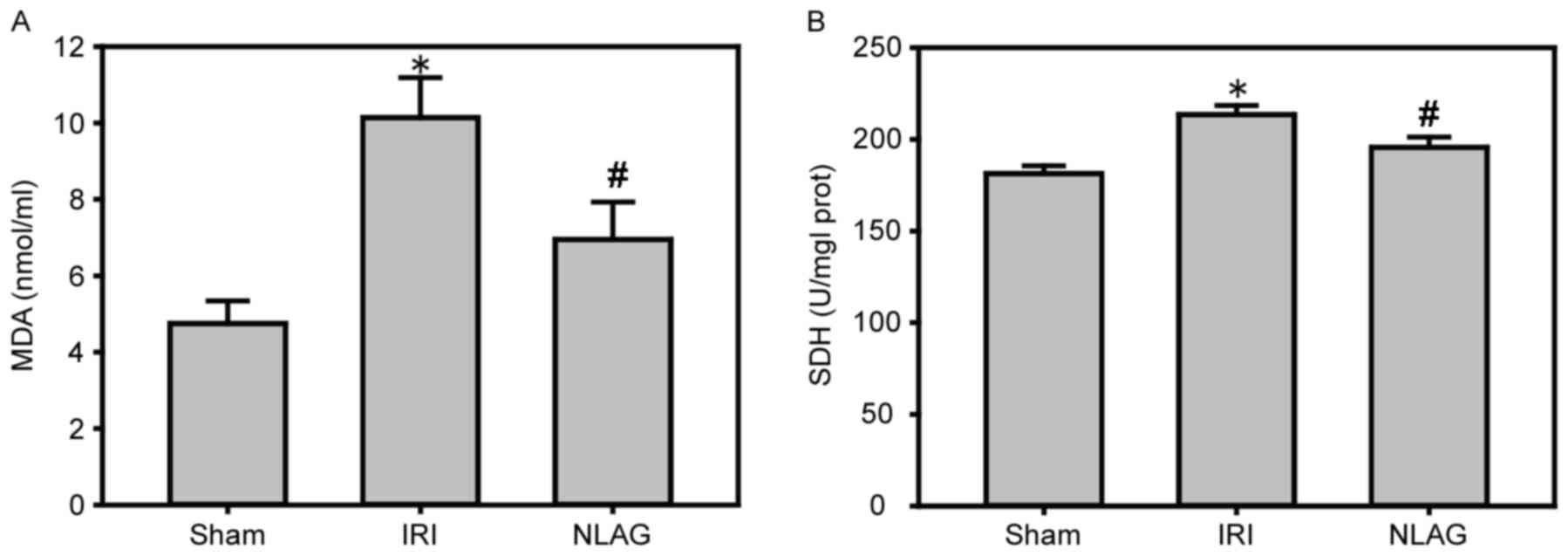
NLAG inhibits IRI-induced protein
expression of oxidative stress
Compared with the Sham group, the protein expression
levels of MDA and SDH were increased in the IRI group (P<0.05).
However, NLAG effectively inhibited the expression of the oxidative
stress indicators, which was statistically significant compared
with the IRI group (P<0.05, Fig.
5).
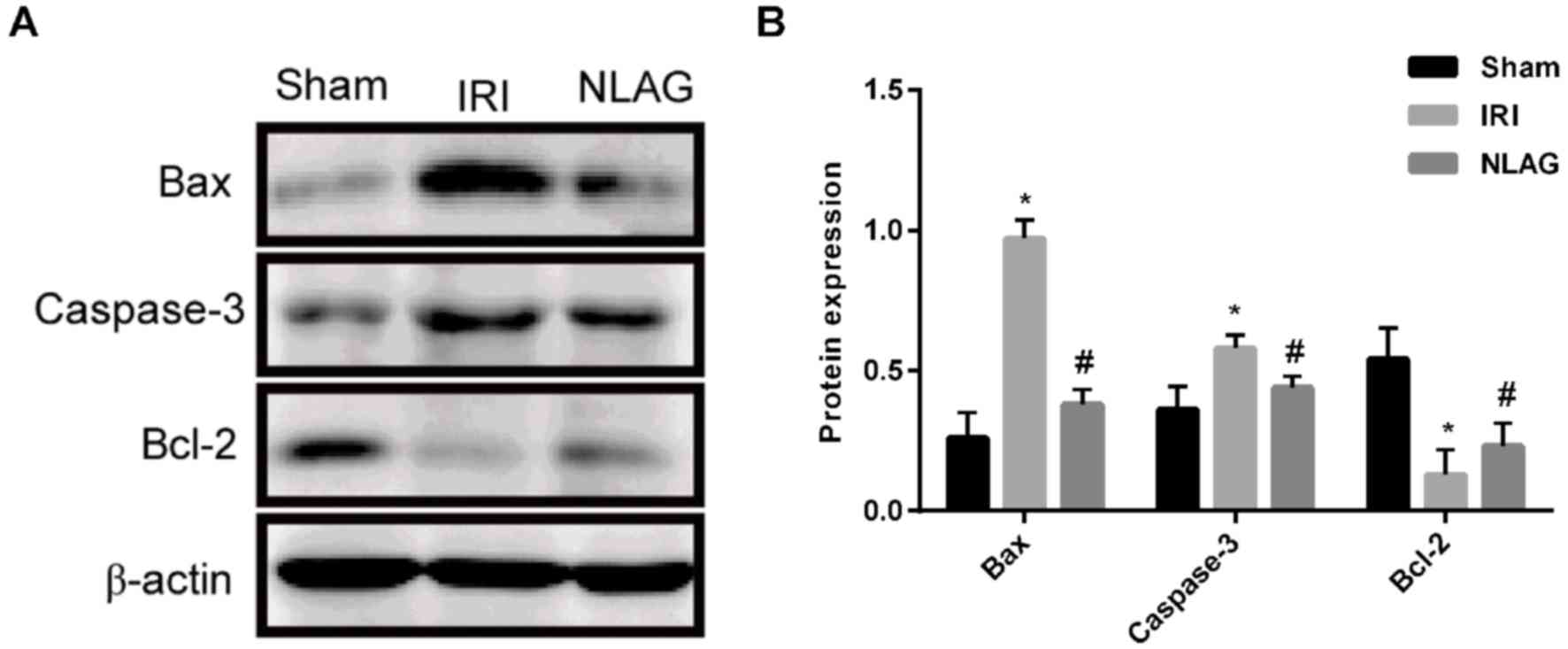
NLAG modulates IRI-induced protein
expression of Bcl-2, Bax and Caspase-3
Compared with the Sham group, the protein expression
levels of Bax and Caspase-3 in the IRI group were increased and the
expression of Bcl-2 was decreased, which was significant
(P<0.05). On the contrary, NLAG significantly increased Bcl-2
expression, and decreased Bax and Caspase-3 expression compared
with the IRI group (P<0.05, Fig
6).
NLAG modulates IRI induced-protein
expression of JAK2, p-JAK2, STAT3 and p-STAT3
Compared with the Sham group, the protein expression
levels of JAK2, p-JAK2, STAT3 and p-STAT3 were decreased
significantly in the IRI group (P<0.05). On the other hand, NLAG
significantly increased JAK2, p-JAK2, STAT3 and p-STAT3 expression
compared with the IRI group (P<0.05, Fig. 7).
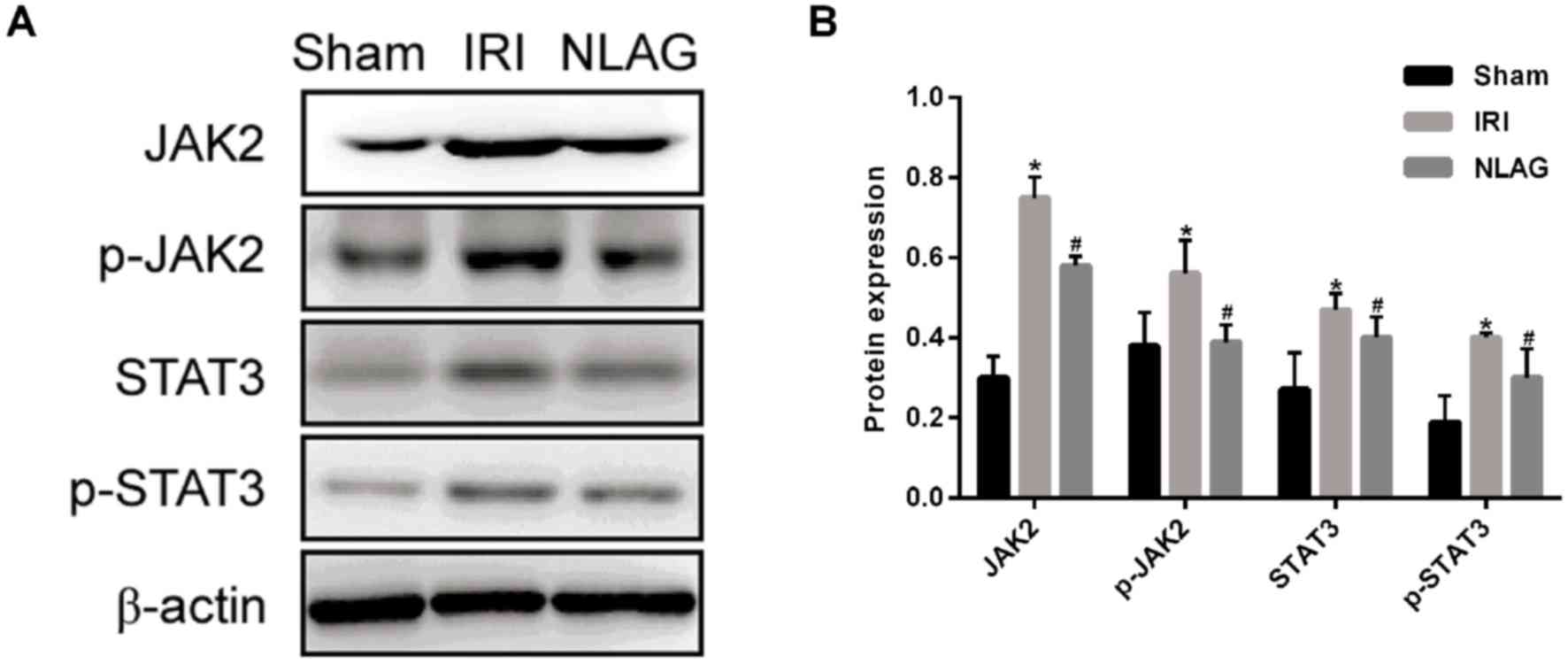 | Figure 7.Expression levels of JAK2, p-JAK2,
STAT3, p-STAT3 proteins were detected by western blot assay. (A)
Western blot analysis of JAK2, p-JAK2, STAT3 and p-STAT3 protein
expression levels. (B) Protein levels of JAK2, p-JAK2, STAT3 and
p-STAT3. Data are presented as the mean ± standard deviation.
*P<0.05 vs. sham group; #P<0.05 vs. IRI group.
NLAG, N(2)-L-alanyl-L-glutamine; IRI, ischemia-reperfusion injury;
JAK, Janus kinase; STAT, Signal transducer and activator of
transcription; p, phosphorylated. |
Discussion
In the present study, a rat IRI model was used to
explore the effects of NLAG on IRI and observe the
histopathological changes and protein expression changes of
inflammatory cytokines (such as IL-1β, IL-6 and TNF-α), of
oxidative stress markers (such as MDA and SDH), of
apoptosis-associated proteins (such as Bcl-2, Bax and Caspase-3)
and of the JAK2/STAT3 signaling pathway-associated proteins. It was
demonstrated that NLAG could reduce the IRI-induced injury on
cardiomyocytes, downregulate the expression of inflammatory
factors, and decrease the apoptosis to some extent. In addition,
NLAG prevented IRI-induced myocardial injury and inhibited JAK2,
STAT3, p-STAT3 and p-JAK2 expression. This study was aimed to
investigate the effects of NLAG on myocardial injury and to explore
the regulatory mechanism of JAK2/STAT3 signaling pathway on
myocardial injury, which provides a basis for cardiopulmonary
bypass.
Previous studies have demonstrated that IRI induces
death mechanisms in a variety of cells, including apoptosis,
necrosis and autophagy (15,16).
The STAT protein family serves an important role in the regulation
of cardiomyocyte apoptosis, whereas STAT3 serves a protective role.
Boengler et al (17)
identified that myocardial infarcted areas were greater in the
STAT3-knockout mice following IRI treatment than in the wild-type
mice, with more apoptotic cardiomyocytes and higher mortality. In
addition, cardiac-specific STAT3 knockout is more sensitive to
inflammatory injury, as the IRI-induced inflammation was more
obvious in the STAT3 knockout mice, especially following myocardial
necrosis. The JAK2/STAT3 signaling pathway is an important
component of the survivor activating factor enhancement, which
protects against the reperfusion injury through activation of TNF-α
(18). Butler et al
(19) have demonstrated that blood
supply from an ischemic preconditioning heart to another heart can
activate STAT3 in the recipient protecting the myocardial tissue.
In the present study, the rat IRI model exhibited myocardial
damage, disordered arrangement of myocardial cells and high levels
of LDH release, inflammatory factors (IL-1β, IL-6 and TNF-α) and
oxygen-free radicals (MDA and SDH), increased expression of
apoptosis-associated proteins, and decreased expression of
JAK2/STAT3-associated proteins. These results suggest that IRI can
induce myocardial injury through inflammation and apoptosis and the
JAK2/STAT3 signaling pathway serves an important role in
inflammation, oxidative stress, cell injury and apoptosis. Terrell
et al (20) have
demonstrated that IL-6 stimulates cardiac hypertrophy through
activation of the JAK/STAT signaling pathway. Park et al
(21) presented that the JAK/STAT
signaling pathway mediates oxidative stress-induced reduction of
pulmonary surfactant epithelium synthesis. In the present study,
inhibition of the JAK/STAT signaling pathway reduced the
IRI-induced myocardial injury and apoptosis. Li et al
(22) identified that fasudil
exerts its anti-IRI effect by inhibiting the endoplasmic reticulum
stress response and by activating the JAK2/STAT3 signaling pathway.
Luan et al (23)
demonstrated that hydrogen sulfide post-treatment can activate the
JAK2/STAT3 signaling pathway, thus protecting the isolated heart
from myocardial IRI. According to the study of Dong et al
(24), rapamycin can exert its
anti-IRI action in vivo and in vitro by activating
the JAK2/STAT3 signaling pathway. It has been widely used in the
treatment of respiratory diseases and digestion systemic diseases.
NLAG also improves ventricular remodeling and reduces myocardial
damage. In the present study, NLAG inhibited both inflammatory
factors and oxidative stress indicators, activated JAK2 and STAT3
proteins, and significantly increased the expression of p-JAK2 and
p-STAT3. These findings indicate that NLAG can attenuate myocardial
IRI, and that the JAK2/STAT3 signaling pathway serves an important
regulatory role in that process.
In conclusion, the results of the present study
suggest that myocardial ischemia-reperfusion may lead to myocardial
cell apoptosis and myocardial injury, and that NLAG attenuates
IRI-induced mitochondrial oxidative stress injury and apoptosis via
activation of the JAK2/STAT3 signaling pathway, thus exerting
protective effects against IRI.
Acknowledgements
The present study was supported by the Tianjin
Municipal Health and Family Planning Commission of Science and
Technology projects (grant no. 14KG125).
References
|
1
|
Igarashi M, Tada H, Yamasaki H, Kuroki K,
Ishizu T, Seo Y, Machino T, Murakoshi N, Sekiguchi Y, Noguchi Y, et
al: Fragmented QRS is a novel risk factor for ventricular
arrhythmic events after receiving cardiac resynchronization therapy
in non-ischemic cardiomyopathy. J Cardiovasc Electrophysiol.
28:327–335. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cordeiro B and Clements R: Murine isolated
heart model of myocardial stunning associated with cardioplegic
arrest. J Vis Exp. e524332015.PubMed/NCBI
|
|
3
|
Wong GW, Laugerotte A and Wright JM: Blood
pressure lowering efficacy of dual alpha and beta blockers for
primary hypertension. Cochrane Database Syst Rev: CD007449. 2015.
View Article : Google Scholar
|
|
4
|
Devi NR, Sumitra C and Singh NR:
Absorption spectral study of Pr(III) complexes with
L-Alanyl-L-Glutamine and N-Acetyl-L-Glutamine: Interaction
parameters, bonding and Judd-Ofelt intensity. Asian J Chem.
24:2863–2870. 2012.
|
|
5
|
Cunha Filho JF, Gonçalves II, Guimarães
SB, Jamacaru FV, Garcia JH and Vasconcelos PR: L-alanyl-glutamine
pretreatment attenuates acute inflammatory response in children
submitted to palatoplasty. Acta Cir Bras. 26 Suppl 1:S72–S76. 2011.
View Article : Google Scholar
|
|
6
|
Gabr A, Salem AAS, Samy HA, Tmam S and Ali
AM: N(2)-L-Alanyl-L-Glutamine dipeptide preventing
oxaliplatin-induced neurotoxicity in colorectal. cancer patients.
07:1–621. 2016.
|
|
7
|
Pires VL, Souza JR, Guimarães SB, Silva
Filho AR, Garcia JH and Vasconcelos PR: Preconditioning with
L-alanyl-L-glutamine in a Mongolian gerbil model of acute cerebral
ischemia/reperfusion injury. Acta Cir Bras. 26 Suppl 1:S14–S20.
2011. View Article : Google Scholar
|
|
8
|
Araújo Júnior RJ, Silva Júnior RG,
Vasconcelos MP, Guimarães SB, Vasconcelos PR and Garcia JH:
Preconditioning with L-alanyl-glutamine reduces hepatic
ischemia-reperfusion injury in rats. Acta Cir Bras. 26 Suppl
1:S8–S13. 2011. View Article : Google Scholar
|
|
9
|
Shen-Orr SS, Furman D, Kidd BA, Hadad F,
Lovelace P, Huang YW, Rosenberg-Hasson Y, Mackey S, Grisar FA,
Pickman Y, et al: Defective signaling in the JAK-STAT pathway
tracks with chronic inflammation and cardiovascular risk in aging
humans. Cell Syst. 3:374–384.e4. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
De S, Manna A, Kundu S, De Sarkar S,
Chatterjee U, Sen T, Chattopadhyay S and Chatterjee M:
Allylpyrocatechol attenuates collagen-induced arthritis via
attenuation of oxidative stress secondary to modulation of the
MAPK, JAK/STAT, and Nrf2/HO-1 pathways. J Pharmacol Exp Ther.
360:249–259. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim HC, Kim E, Bae JI, Lee KH, Jeon YT,
Hwang JW, Lim YJ, Min SW and Park HP: Sevoflurane postconditioning
reduces apoptosis by activating the JAK-STAT pathway after
transient global cerebral ischemia in rats. J Neurosurg
Anesthesiol. 29:37–45. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mudaliar H, Rayner B, Billah M, Lay W and
Bhindi R: Remote ischaemic preconditioning activates the JAK-STAT
pathway resulting in the attenuation of Egr-1 expression following
myocardial ischaemia reperfusion injury. Heart Lung Circulat. 24
Suppl 3:S1632015. View Article : Google Scholar
|
|
13
|
Wen SH, Li Y, Li C, Xia ZQ, Liu WF, Zhang
XY, Lei WL, Huang WQ and Liu KX: Ischemic postconditioning during
reperfusion attenuates intestinal injury and mucosal cell apoptosis
by inhibiting JAK/STAT signaling activation. Shock. 38:411–419.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baguisi A, Casale RA, Kates SA, Lader AS,
Stewart K and Beeuwkes R III: CMX-2043 efficacy in a rat model of
cardiac ischemia-reperfusion injury. J Cardiovasc Pharmacol Ther.
21:563–569. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shu S, Li CM, You YL, Qian XL, Zhou S and
Ling CQ: Electroacupuncture ameliorates cerebral
ischemia-reperfusion injury by regulation of autophagy and
apoptosis. Evid Based Complement Alternat Med. 2016:72974252016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Z, Yu J, Wu J, Qi F, Wang H, Wang Z
and Xu Z: Scutellarin protects cardiomyocyte ischemia-reperfusion
injury by reducing apoptosis and oxidative stress. Life Sci.
157:200–207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Boengler K, Buechert A, Heinen Y, Roeskes
C, Hilfiker-Kleiner D, Heusch G and Schulz R: Cardioprotection by
ischemic postconditioning is lost in aged and STAT3-deficient mice.
Circ Res. 102:131–135. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jacoby JJ, Kalinowski A, Liu MG, Zhang SS,
Gao Q, Chai GX, Ji L, Iwamoto Y, Li E, Schneider M, et al:
Cardiomyocyte-restricted knockout of STAT3 results in higher
sensitivity to inflammation, cardiac fibrosis, and heart failure
with advanced age. Proc Natl Acad Sci USA. 100:pp. 12929–12934.
2003; View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Butler KL, Huffman LC, Koch SE, Hahn HS
and Gwathmey JK: STAT-3 activation is necessary for ischemic
preconditioning in hypertrophied myocardium. Am J Physiol Heart
Circ Physiol. 291:H797–H803. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Terrell AM, Crisostomo PR, Wairiuko GM,
Wang M, Morrell ED and Meldrum DR: Jak/STAT/SOCS signaling circuits
and associated cytokine-mediated inflammation and hypertrophy in
the heart. Shock. 26:226–234. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park SK, Dahmer MK and Quasney MW: MAPK
and JAK-STAT signaling pathways are involved in the oxidative
stress-induced decrease in expression of surfactant protein genes.
Cell Physiol Biochem. 30:334–346. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Y, Zhu W, Tao J, Xin P, Liu M, Li J and
Wei M: Fasudil protects the heart against ischemia-reperfusion
injury by attenuating endoplasmic reticulum stress and modulating
SERCA activity: The differential role for PI3K/Akt and JAK2/STAT3
signaling pathways. PLoS One. 7:e481152012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luan HF, Zhen LI, Zhao QH, Wang L, Yong JI
and Zeng YM: Role of JAK2/STAT3 signaling pathway in hydrogen
sulfide postconditioning on isolated ischemia/reperfusion rat
hearts. Chin J Pharmacol Toxicol. 25:23–28. 2011.(In Chinese).
|
|
24
|
Dong J, Jiang Y, Liu A, Wang S, Yang Q and
Zhang Y: Expression of JAK/STAT signal transduction pathway and the
effects of rapamycin in lupus nephritis-prone MRL/lpr mice. Chin J
Microbiol Immunol. 15:73–77. 2007.
|