Introduction
Dental pulp stem cells (DPSCs) are highly
proliferative, multipotent, colorogenic type cells capable of
multilineage differentiation and self-renewal which may be used for
different regenerative medicine applications, including bone tissue
engineering (1–3). DPSCs have a natural function in the
production of odontoblasts and are capable of osteogenic
differentiation (4–6). In previous studies, it was
demonstrated that tumor necrosis factor-α (TNF-α) may successfully
promote the transition between DPSCs and bone cells through
step-wise, globally wide mRNA expressional alterations (7,8).
Nevertheless, it remains unknown whether other types of mechanisms,
including epigenetic regulation are involved in the osteogenic
differentiation of DPSCs.
Accumulating evidence have identified that
non-coding RNA transcripts, including microRNAs and long non-coding
RNAs (lncRNAs) are essential in stem cell proliferation and
differentiation (9–11). Different from mRNAs and microRNAs,
lncRNAs are transcribed by RNA polymerase II; however, lack stable
open reading frames (12–14). Functioning in the cis- or
trans-manners, lncRNAs may either serve as a platform to recruit
complex protein machinery to bind specific DNA loci or directly
bind RNA molecules to implement post-transcriptional regulation
(12–14). A number of lncRNAs have been
demonstrated to serve regulatory roles in osteogenic
differentiation from mesenchymal stem cells (15–17).
Nevertheless, whether lncRNAs are involved in the osteogenic
differentiation of DPSCs remains unclear.
The present study examined the involvement of
lncRNAs in the osteogenic differentiation of DPSCs. By
RNA-Sequencing (RNA-Seq) and reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) validation, alterations in
lncRNA expression at different phases of osteogenic differentiation
were identified. Further analysis identified that one lncRNA, X
inactive specific transcript (XISP), is required for this process.
These findings provide insight for the understanding of in
vitro induced differentiation of DPSC mechanisms, thus
identifying potential molecular targets which promote the
osteogenic differentiation from DPSCs, which may be useful for
translational studies using DPSCs for bone tissue engineering.
Materials and methods
Cell culture
All procedures in the present study involving human
participants were approved by the Ethics Committee of the
Affiliated Hospital of Nantong University (Nantong, China), and
performed according to the 1964 Helsinki declaration and its later
amendments or comparable ethical standards. To obtain the DPSCs,
normal human impacted third molars were first collected from 6
patients (age range: 22–41 years; 3 male, 3 female) with no carious
lesions and oral infection between May and Aug, 2016. Written
informed consent was obtained from all participants. The fresh
isolated teeth were subsequently washed and opened to reveal the
pulp chamber. A solution with 3 mg/ml collagenase type I was used
to digest the pulp cells at 37°C for 1 h. Single cell suspensions
were obtained and cultured with Dulbecco's modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 mg/ml streptomycin,
in a humidified atmosphere containing 5% CO2 and 95% air
at 37°C.
Osteogenic differentiation
Fresh DPSCs were cultured for three passages, prior
to being used in the differentiation assay. In total,
2×104 cells/cm2 were cultured in DMEM
supplemented with 0.1 mM dexamethasone, 10 mM β-glycerophosphate
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 50 mg/ml ascorbic
acid (Sigma-Aldrich; Merck KGaA) and 10 ng/ml TNF-α. DPSCs were
differentiated for 7 or 14 days prior to being subjected to RNA
extraction.
To transfect DPSCs at Day 1, control small
interfering (si)RNA (5′-ACGUGACACGUUCGGAGAA-3′; 200 nM) and XIST
siRNA (GCTTCTAACTAGCCTGAAT; 200 nM) were mixed with
Lipofectamine® RNAiMAX Reagent (1:1; cat. no. 13778030;
Thermo Fisher Scientific, Inc.) in opti-Minimum Essential Medium
(Gibco; Thermo Fisher Scientific, Inc.); the solution was
subsequently suspended in the culture DMEM. The culture medium was
changed 16 h after transfection.
RNA extraction, RNA-seq and
bioinformatics analysis
Total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and assessed
using the Agilent 2200 Bioanalyzer (Agilent Technologies, Inc.,
Santa Clara, USA) for RNA quality. Samples were processed using the
Illumina mRNA-Seq Sample Preparation kit (Illumina, Inc. San Diego,
CA, USA; cat. nos. 1004824 and 1004825). RNA-seq libraries were 100
bp, paired-end sequenced on an Illumina HiSeq 2000. Sequencing
reads following the removal of polymers, primer adaptors and
ribosomal RNAs were aligned to the human genome with
SOAPaligner/soap2 (version 2.21) (http://soap.genomics.org.cn/soapaligner.html). The
alignment data was utilized to calculate the distribution of reads
on reference genes and perform coverage analysis. The expression
levels of individual RNAs were measured by reads per kilo-base per
million following quality controls. Fisher's exact test was used
for pathway enrichment analysis and gene-act-network analysis. A
false discovery rate <0.05 was considered statistically
significant. Kyoto Encyclopedia of Genes and Genomes analysis
(KEGG; http://www.genome.jp/kegg/) was
performed to identify specific pathways involved. The lncRNA
expression pattern clustering was based on a previously published
algorithm (18). To perform the
coding non-coding co-expression network analysis, Pearson's
correlation was calculated and the significant correlation pairs
(>0.999) were selected, which were used to construct the network
(19).
RT-qPCR
Total RNA was extracted using TRIzol®
reagent (Invitrogen, USA). RT was performed with the high-capacity
cDNA RT kit (Applied Biosystems; Thermo Fisher Scientific, Inc.):
25°C for 10 min, 37°C for 120 min, 85°C for 5 min and then held at
4°C. Real-time PCR was performed in triplicate using SYBR green
qPCR master mix (Qiagen GmbH, Hilden, Germany) and the CFX96 qPCR
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA): 95°C for 5
min, 40 cycles of 95°C, 15 sec, 60°C 30 sec, 72°C 30 sec, and 72°C
10 min. The expression levels of GAPDH mRNA abundance were used for
normalization and 2−∆∆Cq method for quantification
(20). Primers used for validation
are listed in Table I.
 | Table I.Primers for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Sequence (5′-3′) |
|---|
| MKLN1-F |
CGCGGAGGACAACTTTTAGC |
| MKLN1-R |
TTAGCTCCTTGCCTCGTTCC |
| SH3BP5-F |
ATCAGGCTCAGGTTTGCTCC |
| SH3BP5-R |
AGTCTCCTGTTCTCTTGATCAGC |
| THAP9-F |
CGATGCGGAGATAATGGGGA |
| THAP9-R |
TCCTTCCCTGCATATTTTGAGTAA |
| XIST-F |
CCCTCATCCCCACTTTTCCC |
| XIST-R |
TGGAATGAGCAGTGTGCGAT |
| GAPDH-F |
AGAAGGCTGGGGCTCATTTG |
| GAPDH-R |
AGGGGCCATCCACAGTCTTC |
Alkaline phosphatase (ALP)
staining
Negative control or siRNA treated culture cells were
fixed (4% paraformaldehyde in 1X PBS, room temperature for 2 h) and
stained with the ALP assay kit (JianCheng, Nanjing, China)
according to the manufacturer's instructions as previously
described (8). Cultures were
imaged were imaged with a Zeiss Apotome microscope (magnification,
20×) equipped with a Zeiss Axiocam MRM REV2 camera (Zeiss AG,
Oberkochen, Germany). Cell counting was performed by eye.
Statistical analysis
For all figures, data are presented as the mean ±
standard error of the mean, and the number (n) of samples used is
indicated in the legends. Student's t-test and one-way analysis of
variance with Bonferroni's correction for multiple comparisons
(Prism 7.0, GraphPad Software, Inc., La Jolla, CA, USA) were
performed to determine the significant differences between
different groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Alterations of lncRNA expression
during osteogenic differentiation of DPSCs
As previously demonstrated, DPSCs were treated using
an osteogenic differentiation medium containing 10 ng/ml TNF-α
(7,8). In the present study, RNA was
collected after 7 and 14 days of treatment with TNF-α, when DPSCs
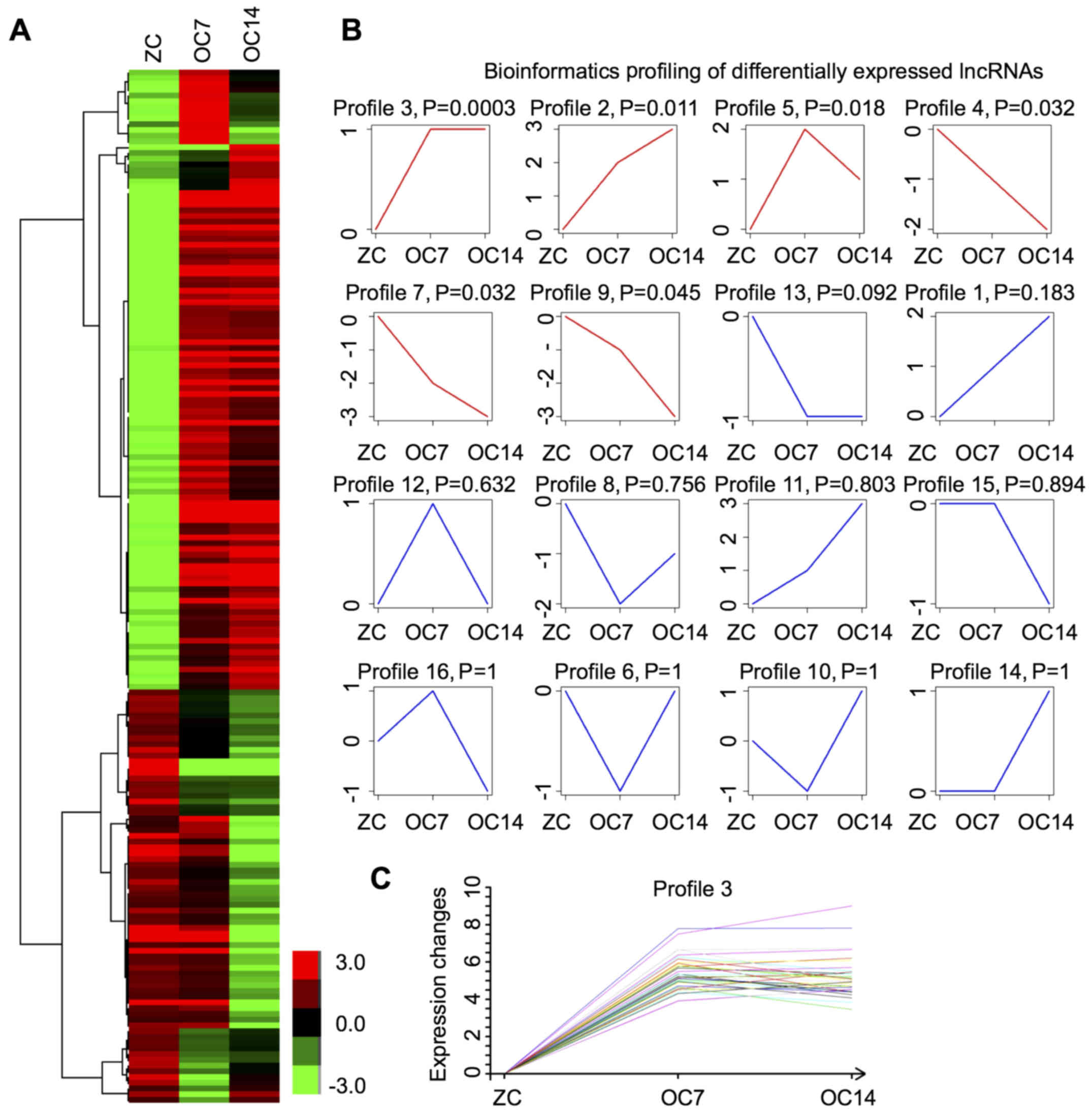
were undergoing and completing osteogenic differentiation (8). The next generation sequencing
analysis identified 77 and 133 lncRNAs, with 30 lncRNAs
overlapping, which were differentially expressed at days 7 and 14
post-treatment, respectively (Fig.
1A). In addition, 58 and 73 were upregulated, and 19 and 60
were downregulated at day 7 and 14, respectively. These results
demonstrated that lncRNAs underwent transitional alterations during
osteogenic differentiation of DPSCs.
Profiling of differentially expressed
lncRNAs
Subsequently, expression pattern analysis was
performed on the differentially expressed lncRNAs during osteogenic
differentiation of DPSCs. The bioinformatics analysis identified 16
different profiles (Fig. 1B).
Among them, six demonstrated statistical significance (Fig. 1B). Subsequently, Profile 3 was
analyzed for the following reasons: i) This profile demonstrated
the highest statistical significance and ii) notably, the
expression levels of all lncRNAs within this profile were increased
at day 7 and their expression was maintained at relatively high
expression levels until day 14 post-treatment with TNF-α (Fig. 1C).
To further validate the RNA-Seq results, the
expression alterations of four predicted lncRNAs in Profile 3 were
measured at 7 and 14 days post TNF-α induction by RT-qPCR. The
results demonstrated a concomitant increase of all four lncRNAs at
day 7, with different expression alterations at day 14 post-TNF-α
induction, which was consistent with RNA-Seq data (Fig. 2).
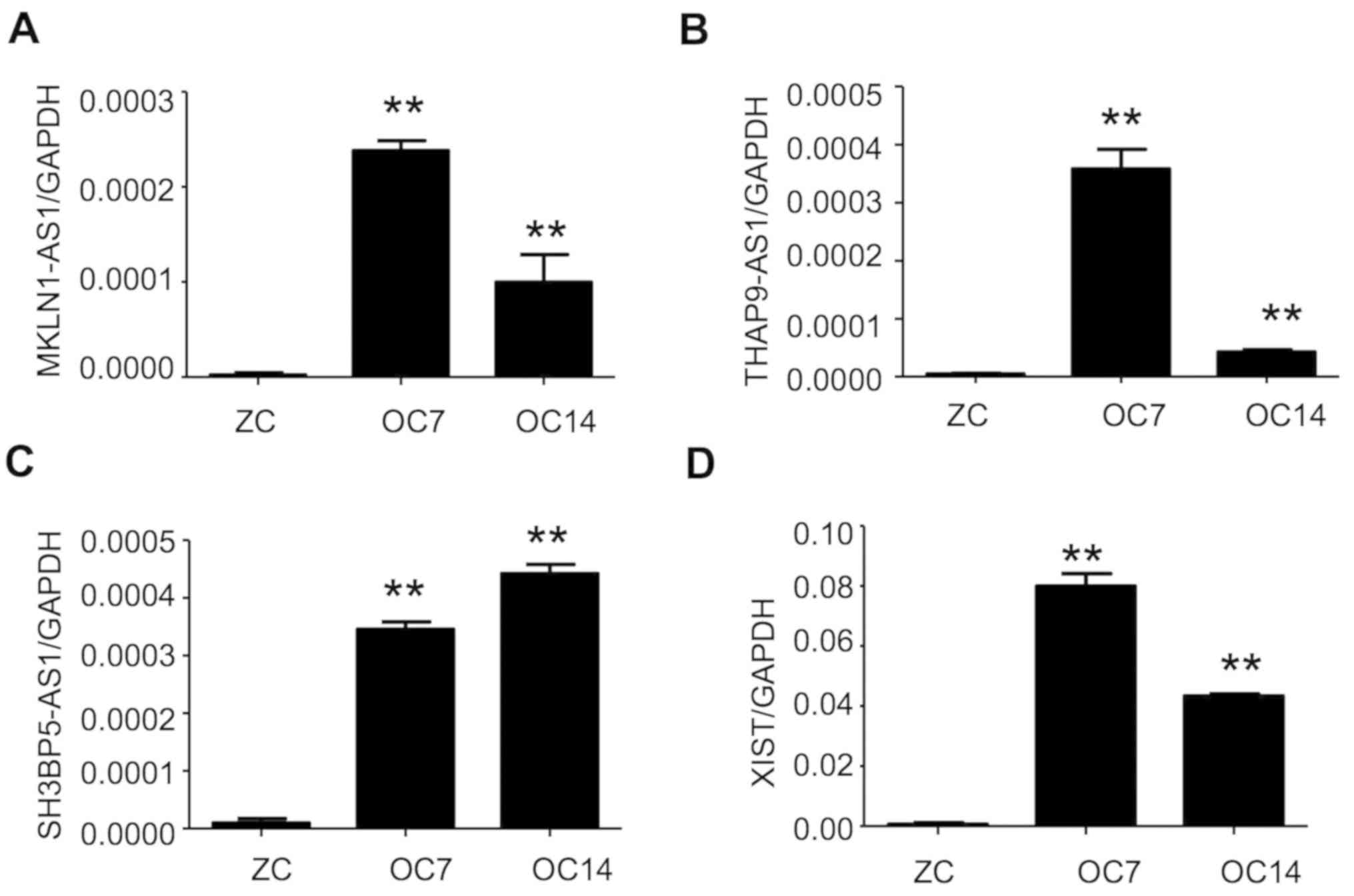 | Figure 2.Validation of key lncRNAs
demonstrating upregulation during osteogenic differentiation of
dental pulp stem cells. Expression alterations at day 7 and 14
following tumor necrosis factor-α induction of selected (A)
MKLN1-AS1, (B) THAP9-AS1, (C) SH3BP5-AS1 and (D) XIST in Profile 3
were validated by quantitative polymerase chain reaction. The GAPDH
mRNA expression level was used for normalization. n=3 in each
condition. **P<0.01 vs. ZC. lncRNAs, long non-coding RNAs;
MKLN1-AS1, MKLN1 antisense RNA 1; THAP9-AS1, THAP9 antisense RNA 1;
SH3BP5-AS1, SH3BP5 antisense RNA 1; XIST, X inactive specific
transcript; ZC, undifferentiated PDSCs; OC7, osteogenic
differentiation at 7 days; OC14, osteogenic differentiation at 14
days. |
Association between differentially
expressed lncRNAs and mRNAs during osteogenic differentiation of
DPSCs
In a previous study (8), mRNA alterations at day 7 and 14 post
TNF-α induction were observed. Therefore, the present study aimed
to investigate how lncRNAs alterations were associated with mRNA
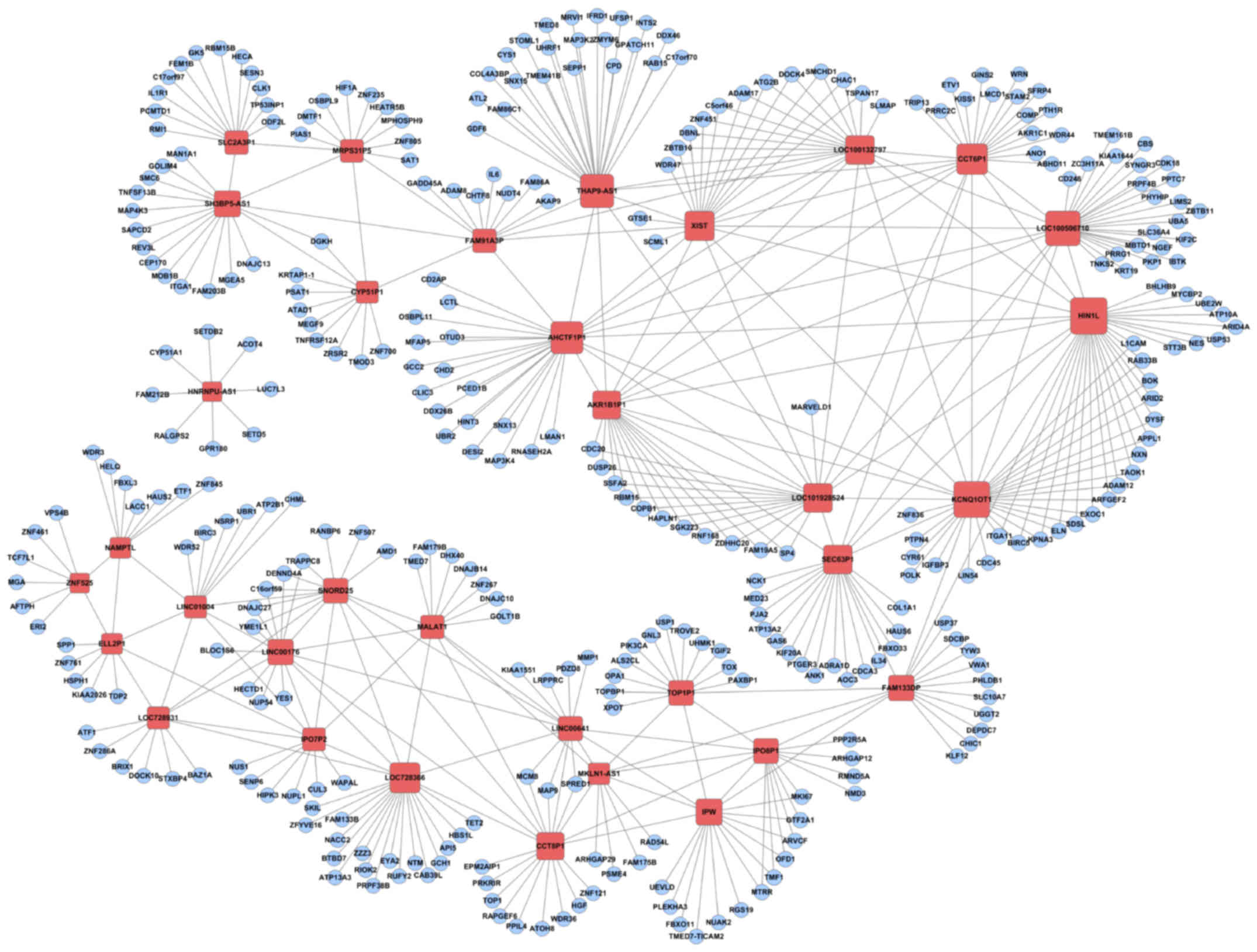
alterations during osteogenic differentiation of DPSCs. By
bioinformatics analysis, 34 lncRNAs were predicted to be associated
with 336 mRNA transcripts that underwent significant alterations
during osteogenic differentiation (Fig. 3). KEGG analysis identified the
‘PI3K-Akt signaling pathway’ and ‘MAPK signaling pathway’, which
have key roles in osteoblast differentiation (21–23)
(Table II).
| Table II.Characterization of functional
relevance to osteoblast differentiation in genes associated with
key lncRNAs. |
Table II.
Characterization of functional
relevance to osteoblast differentiation in genes associated with
key lncRNAs.
| KEGG ID | Pathway | Count | P-value |
|---|
| 04510 | Focal adhesion | 8 |
8.48×105 |
| 00260 | Glycine, serine and
threonine metabolism | 4 | 0.0003 |
| 04512 | ECM-receptor
interaction | 5 | 0.0005 |
| 03008 | Ribosome biogenesis
in eukaryotes | 5 | 0.0005 |
| 04151 | PI3K-Akt signaling
pathway | 9 | 0.0006 |
| 04141 | Protein processing in
endoplasmic reticulum | 6 | 0.001 |
| 04115 | p53 signaling
pathway | 4 | 0.002 |
| 04120 | Ubiquitin mediated
proteolysis | 5 | 0.004 |
| 00270 | Cysteine and
methionine metabolism | 3 | 0.007 |
| 04010 | MAPK signaling
pathway | 6 | 0.012 |
lncRNA XIST is required for efficient
osteogenic differentiation of DPSCs induced by TNF-α
At present, whether and how lncRNAs are involved in
osteoblast differentiation remains unclear. To investigate this, a
validated lncRNA, XIST (Fig. 2),
was selected and it was examined to determine how it may affect
osteoblast differentiation. RT-qPCR confirmed that a specific siRNA
was able to downregulate XIST expression in DPSCs (Fig. 4A). A total of 14 days after TNF-α
induction, inhibition of XIST by siRNA in primarily cultured DPSCs
significantly decreased the presence of alkaline phosphatase
positive osteoblast cells (P<0.01; Fig. 4B and C). Therefore, XIST, an lncRNA
is required for efficient osteogenic differentiation, possibly
through a regulatory role in a group of mRNAs associated with this
process.
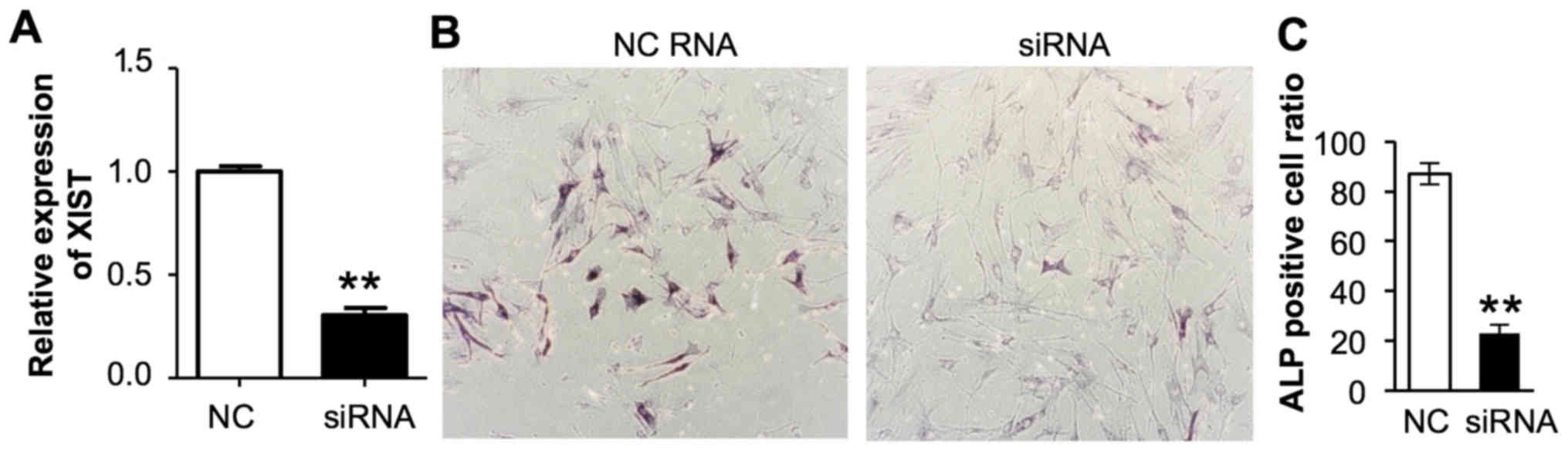 | Figure 4.Effects of XIST knockdown on
osteogenic differentiation of DSCPs. (A) Effects of XIST knockdown
by control or XIST siRNA in DSCPs. The GAPDH mRNA expression level
was used for normalization. **P<0.01, n=3,3 for control and XIST
siRNA treated groups. (B) Representative images (magnification,
×20) of control or XIST siRNA transfected DSCPs were stained for
ALP at 14 days post culture with (C) quantification. **P<0.01,
Student's t test. n=4,6 for control and XIST siRNA treated groups.
XIST, X inactive specific transcript; DSCPs, dental pulp stem
cells; siRNA, small interfering RNA; ALP, alkaline phosphatase; NC,
negative control. |
Discussion
DPSCs are considered as ideal candidates for
osteogenic differentiation, although their underlying mechanisms
remain largely unknown. In the present study, the expression
alterations of lncRNAs in TNF-α induced osteogenic differentiation
of DPSCs were investigated. The present results identified
transitional, global alterations of lncRNAs, which were associated
with mRNAs involved in key signaling pathways for osteoblast
differentiation. The present data further suggested that one lncRNA
in particular, XIST, is essential for efficient osteogenic
differentiation.
lncRNAs, as a type of non-coding RNAs, exhibit a
variety of different cytotopic localizations and functional
regulating modes. Such diversity and flexibility make lncRNAs good
candidates to regulate gene expression in a temporospatial manner
responding to complex situations, including during cellular
differentiation (10–14). For example, multiple lncRNAs were
demonstrated to serve important roles in neural, skin and muscle
stem cell differentiation (24).
XIST encodes a 17-kb long non-coding RNA. Silencing
factors are commonly recruited for global gene silencing on X
chromosome (25). However, its
role outside the X chromosome and during osteogenic differentiation
is largely unknown. The aim of future research is to identify
XIST-associated mRNA transcripts and to study their expressional
alterations during TNF-α induced osteogenic differentiation of
DPSCs. Due to the limitation of the current study, the
contributions of many lncRNAs were not examined in TNF-α induced
osteogenic differentiation. Nevertheless, the present findings
provide insight for the understanding of molecular mechanisms
underlying differentiation approaches of DPSCs and may be used to
promote the potential regenerative therapies that use DPSCs as
tissue resources.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
RT and YXL conceived and coordinated the study,
designed, performed and analyzed the experiments, and wrote the
paper. YKL, FL and ZYZ conducted the data collection and data
analysis, and revised the paper. All authors read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
All procedures performed in the present study
involving human participants were approved by the Ethics Committee
of the Affiliated Hospital of Nantong University (Nantong, China),
and according to the 1964 Helsinki declaration and its later
amendments or comparable ethical standards. Written informed
consent was obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DPSCs
|
dental pulp stem cells
|
|
lncRNA
|
long non-coding RNA
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
References
|
1
|
Gronthos S, Brahim J, Li W, Fisher LW,
Cherman N, Boyde A, DenBesten P, Robey PG and Shi S: Stem cell
properties of human dental pulp stem cells. J Dent Res. 81:531–535.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gronthos S, Mankani M, Brahim J, Robey PG
and Shi S: Postnatal human dental pulp stem cells (DPSCs) in vitro
and in. Proc Natl Acad Sci USA. 97:13625–13630. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tatullo M, Marrelli M, Shakesheff KM and
White LJ: Dental pulp stem cells: Function, isolation and
applications in regenerative medicine. J Tissue Eng Regen Med.
9:1205–1216. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
d'Aquino R, Graziano A, Sampaolesi M,
Laino G, Pirozzi G, De Rosa A and Papaccio G: Human postnatal
dental pulp cells co-differentiate into osteoblasts and
endotheliocytes: A pivotal synergy leading to adult bone tissue
formation. Cell Death Differ. 14:1162–1171. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Graziano A, d'Aquino R, Laino G and
Papaccio G: Dental pulp stem cells: A promising tool for bone
regeneration. Stem Cell Rev. 4:21–26. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lindroos B, Mäenpää K, Ylikomi T, Oja H,
Suuronen R and Miettinen S: Characterisation of human dental stem
cells and buccal mucosa fibroblasts. Biochem Biophys Res Commun.
368:329–335. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Feng X, Feng G, Xing J, Shen B, Li L, Tan
W, Xu Y, Liu S, Liu H, Jiang J, et al: TNF-α triggers osteogenic
differentiation of human dental pulp stem cells via the NF-kappaB
signalling pathway. Cell Biol Int. 37:1267–1275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu YK, Zhou ZY and Liu F: Transcriptome
changes during TNF-α promoted osteogenic differentiation of dental
pulp stem cells (DPSCs). Biochem Biophys Res Commun. 476:426–430.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gangaraju VK and Lin H: MicroRNAs: Key
regulators of stem cells. Nat Rev Mol Cell Biol. 10:116–125. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fatica A and Bozzoni I: Long non-coding
RNAs: New players in cell differentiation and development. Nat Rev
Genet. 15:7–21. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luo S, Lu JY, Liu L, Yin Y, Chen C, Han X,
Wu B, Xu R, Liu W, Yan P, et al: Divergent lncRNAs regulate gene
expression and lineage differentiation in pluripotent cells. Cell
Stem Cell. 18:637–652. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Geisler S and Coller J: RNA in unexpected
places: Long non-coding RNA functions in diverse cellular contexts.
Nat Rev Mol Cell Biol. 14:699–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guttman M and Rinn JL: Modular regulatory
principles of large non-coding RNAs. Nature. 482:339–346. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Engreitz JM, Ollikainen N and Guttman M:
Long non-coding RNAs: Spatial amplifiers that control nuclear
structure and gene expression. Nat Rev Mol Cell Biol. 17:756–770.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang Y, Zheng Y, Jia L and Li W: Long
noncoding RNA H19 promotes osteoblast differentiation via
TGF-β1/Smad3/HDAC signaling pathway by deriving miR-675. Stem
Cells. 33:3481–3492. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li H, Zhang Z, Chen Z and Zhang D:
Osteogenic growth peptide promotes osteogenic differentiation of
mesenchymal stem cells mediated by LncRNA AK141205-induced
upregulation of CXCL13. Biochem Biophys Res Commun. 466:82–88.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhuang W, Ge X, Yang S, Huang M, Zhuang W,
Chen P, Zhang X, Fu J, Qu J and Li B: Upregulation of lncRNA MEG3
promotes osteogenic differentiation of mesenchymal stem cells from
multiple myeloma patients by targeting BMP4 transcription. Stem
Cells. 33:1985–1997. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ramoni MF, Sebastiani P and Kohane IS:
Cluster analysis of gene expression dynamics. Proc Natl Acad Sci
USA. 99:9121–9126. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shi J, Chen X, Li H, Wu Y, Wang S, Shi W,
Chen J and Ni Y: Neuron-autonomous transcriptome changes upon
ischemia/reperfusion injury. Sci Rep. 7:58002017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
James AW: Review of signaling pathways
governing MSC osteogenic and adipogenic differentiation.
Scientifica (Cairo). 2013:6847362013.PubMed/NCBI
|
|
22
|
Ge C, Xiao G, Jiang D and Franceschi RT:
Critical role of the extracellular signal-regulated kinase-MAPK
pathway in osteoblast differentiation and skeletal development. J
Cell Biol. 176:709–718. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rahman MS, Akhtar N, Jamil HM, Banik RS
and Asaduzzaman SM: TGF-β/BMP signaling and other molecular events:
Regulation of osteoblastogenesis and bone formation. Bone Res.
3:150052015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Flynn RA and Chang HY: Long noncoding RNAs
in cell-fate programming and reprogramming. Cell Stem Cell.
14:752–761. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cerase A, Pintacuda G, Tattermusch A and
Avner P: Xist localization and function: New insights from multiple
levels. Genome Biol. 16:1662015. View Article : Google Scholar : PubMed/NCBI
|