Introduction
Osteogenic signaling pathways and transcription
factors tightly regulate normal skeletal development and renewal of
bone (1,2). The important role of Wnt signaling
during bone formation has been well established. Numerous human
genetic studies have revealed that mutations in Wnt proteins or
co-receptors result in osteoporosis, osteosclerosis and increased
fracture risk (3–6). In agreement with such results in
humans, genetic studies in mice have demonstrated similar roles of
Wnt signaling in bone formation (7–9). Wnt
proteins are a family of secreted glycoproteins, and associated
signals are transduced by the binding of a Wnt protein to the
frizzled receptor and co-receptor [such as low-density lipoprotein
receptor related protein (LRP)5 and LRP6], which activates numerous
distinct intracellular signaling cascades (10–13).
The most well characterized canonical Wnt pathway is the β-catenin
dependent pathway. The Wnt/β-catenin signaling pathway is an
evolutionarily conserved developmental signaling cascade that has
an important role in regulating organ development and tissue
homeostasis (14). β-catenin can
interact with the lymphoid enhancing binding factor (Lef)/T cell
factor 1 (Tcf-1) family of high mobility group-type transcription
factors to induce the expression of target genes (15–18).
Gene regulatory mechanisms are essential for bone
formation, including the epigenetic regulation of genes and
post-translational biochemical modifications of protein activity.
An increasing number of studies have revealed that micro (mi)RNAs,
which are small, non-coding RNAs, have essential roles in the
epigenetic control of bone development (19). miRNAs dynamically regulate
osteoblast differentiation through stages of maturation and mediate
the activities of osteogenic signaling pathways (20). Dicer, an endoribonuclease, is
responsible for the processing of the mature miRNA duplex (21). Dicer deletion in mesenchymal
progenitors decreases fetal survival and bone formation in mice,
whereas excision in differentiated osteoblasts increases bone mass
in adult mice (22). The
conditional deletion of Dicer in mesenchymes by Wnt1-Cre, or in the
epithelium by shh-Cre, revealed markedly different defects in tooth
development (23), which suggests
that deficiencies in mature miRNAs have the same effect on normal
bone homeostasis when compared with miRNAs.
Although numerous miRNAs have been identified to
target Wnt signaling components in osteoblasts (24–28),
it has not been established whether miRNAs represent antagonists or
activators of Wnt signaling during bone formation. In the present
study, Dicer was knocked down in mouse bone marrow stromal cells
(BMSCs) in order to establish an osteoprogenitor cell model with
miRNA production deficiency. The results revealed that significant
inhibition of osteogenic differentiation was associated with the
differential regulation of numerous Wnt signalling components via
the inhibition of Dicer in BMSCs. In contrast, the mechanism
regulating the differentiation of osteoblast cells (MC3T3-E1) was
identified to be different from BMSCs. The results of the present
study also suggested that Wnt4 and Wnt10a may have more important
roles during osteogenic differentiation compared with other Wnt
genes. Furthermore, five putative Wnt10a-targeting miRNAs were
demonstrated to be significantly upregulated during osteoblast
differentiation.
Materials and methods
Cell culture
MC3T3-E1 are mouse osteoblasts (American Type
Culture Collection, Manassas, VA, USA) and were cultured in
α-minimal essential medium (MEM) supplemented with 10% fetal bovine
serum (FBS; Atlanta Biologicals, Inc., Atlanta, GA, USA). NIH3T3
cells are mouse fibroblasts (American Type Culture Collection) and
were maintained in Dulbecco's modified Eagle's medium (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% FBS (Atlanta Biologicals, Inc.). Cells were cultured in a
humidified incubator at 37°C with 5% CO2 (Thermo Fisher
Scientific, Inc.). In order to investigate cell differentiation,
MC3T3-E1 cells were incubated at 100% confluency in the
aforementioned medium containing 10 mM β-glycerol phosphate and 50
µg/ml ascorbic acid (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
and fresh medium was added every other day, as previously reported
(29).
Isolation of BMSCs
BMSCs were isolated from the femurs and tibias of
five female and five male C57BL/6 mice (age, 6–8 weeks; weight,
20±2 g). All mice were purchased from and housed in a designated
and government-approved animal facility of The Fourth Hospital of
Hebei Medical University (Shijiazhuang, China). The mice were
maintained at 21±2°C, with 40–60% relative humidity, in a 12-h
light/dark cycle and with free access to food and water. The
present study was granted ethical approval by The Fourth Hospital
of Hebei Medical University. BMSCs were prepared as previously
reported (30). Soft tissues were
disassociated from tibias and femurs. Bones were scrubbed and
washed to remove impurities with complete α-MEM medium on ice. A
23-gauge needle was inserted into the bone cavity to extract the
bone marrow. Syringes with 20-gauge needles were used to dissociate
cell clumps in the crude suspension. Following this, erythrocytes
were eliminated using hypotonic lysis buffer and the collected
cells were then resuspended in MSC growth media [ascorbic acid-free
α-MEM medium supplemented with 1X penicillin/streptomycin and 18%
FBS (Hyclone; GE Healthcare)]. Single cell suspension was plated at
a density of 1.5×105 cells per 10 cm culture plate for
the expansion of the MSCs. BMSCs ranging from passages 6–10 were
used for all subsequent experiments.
Transduction of Dicer short hairpin
(sh)RNA and osteogenic differentiation in BMSCs
In total, 6 µg lentiviral plasmid encoding for
shRNAs against mouse Dicer1 (cat. no. MSH039774 GeneCopoeia, Inc.,
Rockville, MD, USA) or containing a scramble shRNA (cat. no.
CSHCTR001-LVRH1GP; GeneCopoeia, Inc.) were co-transfected with 4 µg
psPAX2 (Addgene, Inc., Cambridge, MA, USA) and 2 µg pMD2.G
(Addgene, Inc.) packaging plasmids into 293FT cells (American Type
Culture Collection) using Lipofectamine® 3000
transfection reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
(30). Following a total of 48 h
post-transfection, the viral particle-containing culture medium was
collected. The medium was subsequently concentrated and 0.5 ml
lentiviral suspension (1×108 IU/ml; multiplicity of
infection, 100) was used to infect BMSCs in the presence of 5 µg/ml
polybrene (Sigma-Aldrich; Merck KGaA), and the infected cells were
then selected using puromycin (Sigma-Aldrich; Merck KGaA). The
infection efficiency was investigated by determining Dicer mRNA
expression levels by reverse transcription-quantitative polymerase
chain reaction (RT-qPCR). Intact BMSCs and infected BMSCs were then
cultured at 37°C in α-MEM with 50 µg/ml ascorbic acid and 10 mM
β-glycerol phosphate (Sigma-Aldrich; Merck KGaA) in order to induce
osteoblast differentiation for 21 days. In order to perform miRNA
analysis and RT-qPCR, cells were harvested at the indicated days
(day 5, 7, 14 and or 21).
RNA isolation and RT-qPCR
RNA was exracted from cells (BMSCs, MC3T3 and
NIH3T3) using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Following purification using Direct-zol RNA
miniprep kit (Zymo Research Corp., Irvine, CA, USA), 1 µg RNA was
reverse transcribed using the qScript cDNA SuperMix for total cDNA
(thermocycling conditions: 25°C for 5 min, 42°C for 30 min and 85°C
for 5 min) or qScript microRNA cDNA Synthesis Kit specifically for
miRNA cDNA (thermocycling conditions: 60 min at 37°C, 5 min at
70°C, 20 min at 42°C and 5 min at 85°C) purchased from Quantabio
(Beverly, MA, USA) according to the manufacturer's protocol. TaqMan
probes (Thermo Fisher Scientific, Inc.) and SYBRGreen (Thermo
Fisher Scientific, Inc.) primers were used to determine relative
gene expression (Tables I and
II). The corresponding TaqMan and
SYBR green mastermix (Thermo Fisher Scientific, Inc.) were used for
the RT-qPCR reaction. All the related reagents and materials were
purchased from Thermo Fisher Scientific, Inc. miRNA expression
levels were detected using SYBR Green miRNA-specific primers
(Table I). RNA isolation and
RT-qPCR analyses were performed according to previously published
protocol (31). RT-qPCR was
performed using a 7300 sequence detection system (Invitrogen;
Thermo Fisher Scientific, Inc.). The qPCR reaction was performed as
follows: An initial incubation at 50°C for 2 min and at 95°C for 10
min, followed by 40 cycles at 95°C for 15 sec, 60°C for 30 sec,
72°C for 30 sec and a final incubation at 72°C for 10 min.
Expression levels were normalized against levels of GAPDH (for
mRNAs) or U6 small nuclear RNA (for miRNAs). The relative
expression levels and fold changes of each gene were calculated via
the 2−ΔΔCq method (32)
using Microsoft Excel 2007 (Microsoft Corporation, Redmond, WA,
USA).
 | Table I.Primer sequences used to investigate
gene expression and miRNA expression levels in the present
study. |
Table I.
Primer sequences used to investigate
gene expression and miRNA expression levels in the present
study.
| A, Genes |
|---|
|
|---|
| Name | Primer sequences
(5′-3′) |
|---|
| Runx2 | For:
CGCCCCTCCCTGAACTCT |
|
| Rev:
TGCCTGCCTGGGATCTGTA |
| ALP | For:
TTGTGCGAGAGAAAGGAGA |
|
| Rev:
GTTTCAGGGCATTTTTCAAGGT |
| OC | For:
CTGACAAAGCCTTCATGTCCAA |
|
| Rev:
GCGCCGGAGTCTGTTCACTA |
| Col1a1 | For: 5′
CCCAAGGAAAAGAAGCACGTC |
|
| Rev:
AGGTCAGCTGGATAGCGACATC |
| Dicer1 | For:
GGTGGTTCGTTTTGATTTGCC |
|
| Rev:
GGCAGTGTTGATTGTGACTC |
| CEBPa | For:
CAAAGCCAAGAAGTCGGTGGACAA |
|
| Rev:
TCATTGTGACTGGTCAACTCCAGC |
| PPARγ | For:
GAGTGTGACGACAAGATTTG |
|
| Rev:
GGTGGGCCAGAATGGCATCT |
| FABP4 | For:
ATGTGCGACCAGTTTGTG |
|
| Rev:
TTTGCCATCCCACTTCTG |
| GAPDH | For:
TCACCACCATGGAGAAGGC |
|
| Rev:
GCTAAGCAGTTGGTGGTGCA |
|
| B,
microRNAs |
|
| Name | Primer sequences
(5′-3′) |
|
| mmu-miR-28 |
AAGGAGCTCACAGTCTATTGAG |
| mmu-miR-149 |
TCTGGCTCCGTGTCTTCACTCCC |
| mmu-miR-212 |
ACCTTGGCTCTAGACTGCTTACT |
| mmu-let-7b |
TGAGGTAGTAGGTTGTGTGGTT |
| mmu-let-7c |
TGAGGTAGTAGGTTGTATGGTT |
| Table II.TaqMan probe IDs used to investigate
gene expression and miRNA expression levels in the present
study. |
Table II.
TaqMan probe IDs used to investigate
gene expression and miRNA expression levels in the present
study.
| Gene | TaqMan assay
ID |
|---|
| Wnt1 | Mm01300555_g1 |
| Wnt3 | Mm00437336_m1 |
| Wnt4 | Mm01194003_m1 |
| Wnt7a | Mm00437356_m1 |
| Wnt10a | Mm00437325_m1 |
| Wnt10b | Mm00442104_m1 |
| Tcf1 | Mm00493445_m1 |
| Lef1 | Mm00550265_m1 |
| SP1 | Mm00489039_m1 |
Alkaline phosphatase (ALP)
staining
ALP substrate solution was prepared using Sigma fast
red tablet sets (cat. no. F4523; Sigma-Aldrich; Merck KGaA)
according to the manufacturer's instructions. BMSCs were fixed at
room temperature for 10 min using 10% neutral buffered formalin
(Sigma-Aldrich; Merck KGaA) cultured in the ALP substrate solution
in a 37°C incubator for 30 min and subsequently monitored using a
light microscope (magnification, ×100) for the development of a red
reaction product, which indicated ALP activity.
miRNA prediction
TargetScan 7.0 (http://www.targetscan.org) and miRWalk 3.0 (http://mirwalk.umm.uni-heidelberg.de/)
were used to identify the potential miRNAs targeting Wnt10a mRNA.
In total, five putative Wnt10a-targeting miRNAs were selected based
on the previously reported function of these miRNAs in bone
development (33–37).
Statistical analysis
Data are presented as the mean ± standard error of
the mean. Student's t-test was calculated using Prism 7 (GraphPad
Software, Inc., La Jolla, CA, USA) were used to analyze the
statistical differences among different groups. P<0.05 was
considered to indicate a statistically significant difference. All
experiments were repeated three times independently.
Results
Profiling of Dicer and Wnt genes
during osteogenic differentiation in BMSCs
Firstly, mouse BMSCs were isolated and
differentiated into multiple lineages, as previously reported
(17). Adipogenic and osteoblastic
markers were detected via qPCR in order to further characterize
BMSCs (Fig. 1A and B). In order to
establish an osteogenic differentiation model, isolated BMSCs were
incubated for five days, which represented the proliferative
expansion period, until they reached 100% confluency. The results
revealed that there were increased levels of osteogenic markers,
which suggested that osteogenic differentiation had been
successfully induced (Fig. 1A).
Important transcription factors (Tcf-1 and Lef1) represent
downstream targets of the Wnt signaling pathway (15–18).
The results of the present study demonstrated Tcf-1 expression was
markedly upregulated during osteogenic differentiation, whereas
levels of Lef1 increased slightly between days 5–7, and then
markedly decreased between days 7–21 (Fig. 1C). Similar to the majority of Wnt
genes, Dicer expression was upregulated by 4 fold during the 21
days of differentiation, which suggested that increased levels of
Dicer is important for osteogenic differentiation in BMSCs
(Fig. 1D). To investigate the
association between Wnt signaling and osteogenic differentiation,
the expression levels of numerous Wnt genes were determined by
RT-qPCR. Four of the six Wnt genes analysed (Wnt1, Wnt3, Wnt7a and
Wnt10b) were revealed to be consistently upregulated throughout the
differentiation process (Fig. 1D).
Furthermore, Wnt4 was demonstrated to be downregulated between days
5–7, and upregulated between days 7–21 (Fig. 1D). However, Wnt10a levels were
revealed to be increased during the proliferative period between
days 5–7, and markedly decreased during the differentiation period
between days 7–21. Dynamic changes of numerous Wnt genes suggested
that members of the Wnt protein family exhibit different roles
during the early and late differentiation stages.
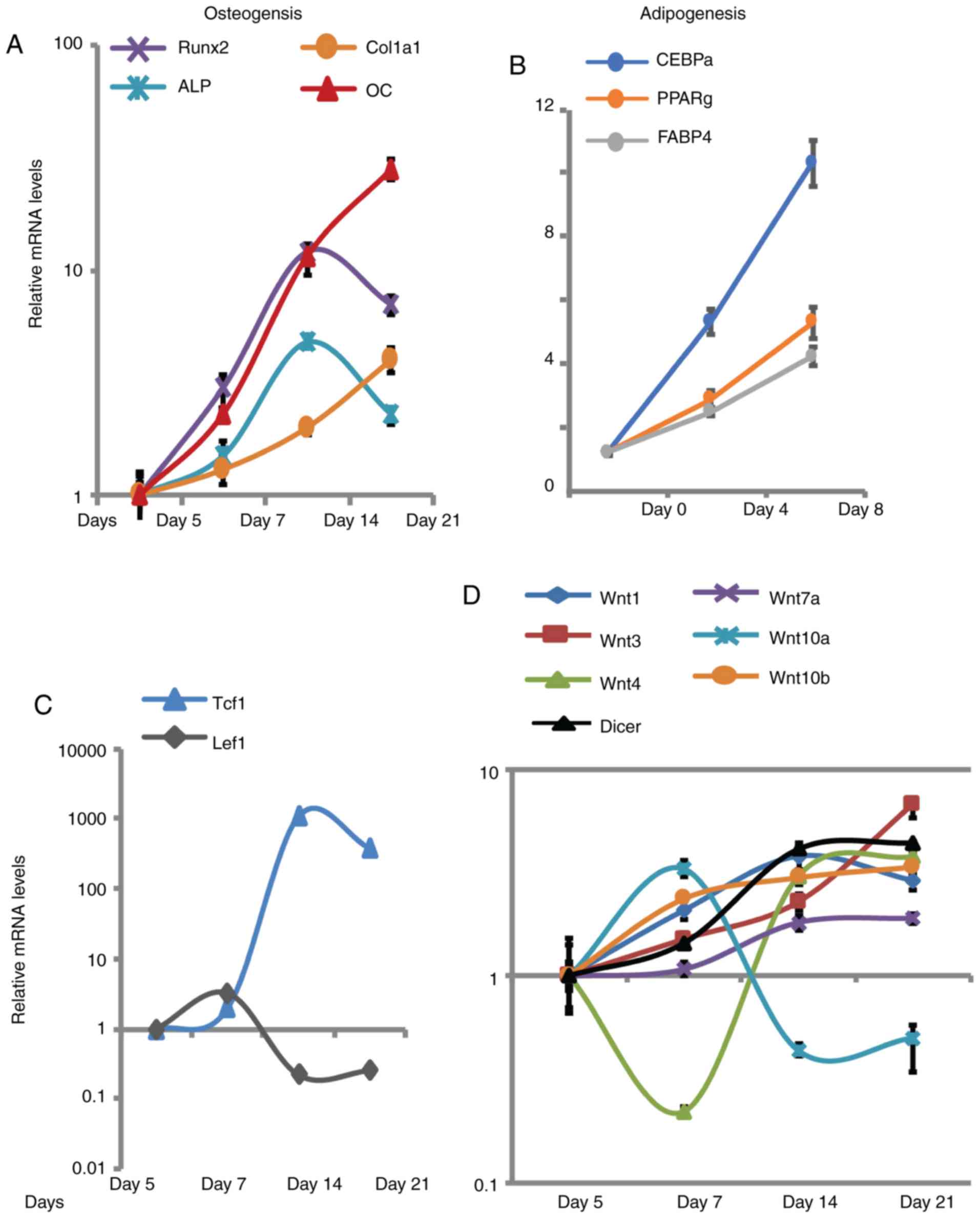 | Figure 1.Wnt genes exhibited dynamic changes
in expression levels during the osteogenic differentiation of
BMSCs. (A) Activation of osteogenic markers were detected during
the osteogenic differentiation of BMSCs. (B) Activation of
adipogenic markers was detected during the adipogenic
differentiation of BMSCs. (C) mRNA expression levels of Tcf-1 and
Lef1 transcription factors downstream of Wnt signalling were
detected in differentiating BMSCs. (D) Levels of Wnt genes and
Dicer were evaluated during differentiation in BMSCs. Gene
expression levels were investigated by reverse
transcription-quantitative polymerase chain reaction. The values
reported were compared with the determined values at day 5 of
osteogenic differentiation or day 0 of adipogenic differentiation
in BMSCs (set as 1). Values are presented as the mean ± standard
error of the mean (n=3). BMSCs, bone marrow stromal cells; ALP,
akaline phosphatase; Tcf1, T-cell factor 1; Lef1, lymphoid enhancer
binding factor 1; Col1a1, collagen 1a1; OC, osteocalcin; Runx2,
runt-related transcription factor 2; CEBPα, CCAAT enhancer binding
protein-α; PPARγ, peroxisome proliferator activated receptor-γ;
FABP4, fatty acid binding protein 4. |
Deletion of Dicer block osteogenic
differentiation, and the regulation of Wnt genes is
stage-specific
To investigate the role of Dicer during osteogenic
differentiation, stable transduction of BMSCs with shDicer was
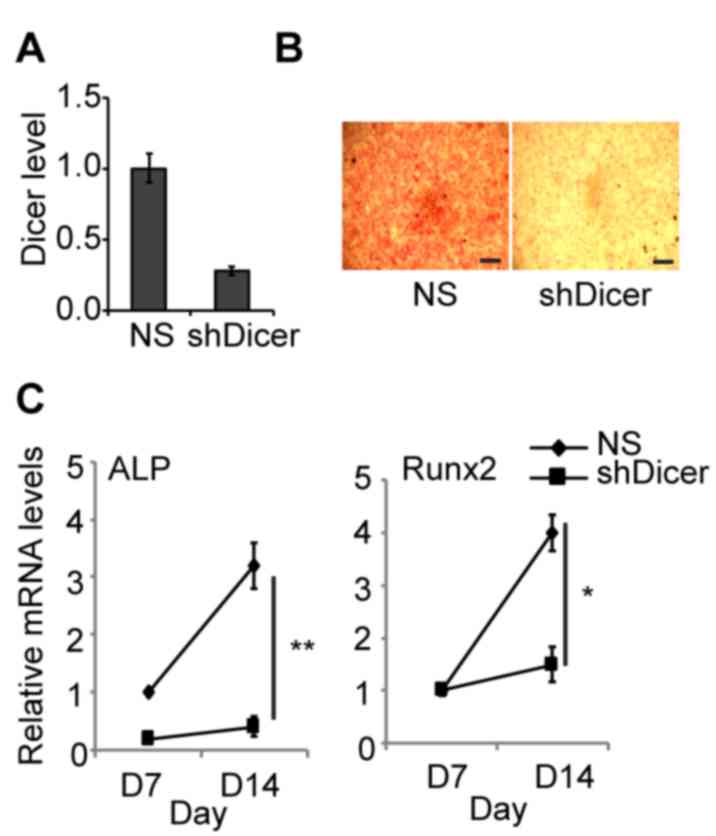
performed. The results revealed that ~80% of Dicer was knocked down
by shDicer (Fig. 2A). The results
of histological analyses further demonstrated that inhibition of
Dicer blocked osteogenic differentiation in BMSCs as assessed by
negative ALP staining, in yellow (Fig.
2B). In BMSCs transfected with scramble shRNA [not specific
(NS)], the expression levels of osteogenic differentiation markers
including Runx2 and ALP, were significantly upregulated by 4-fold
on day 14 of differentiation compared with day 7 (Fig. 2C). Therefore, the results suggested
that knockdown of Dicer almost completely blocked the activation of
these markers during osteogenic differentiation.
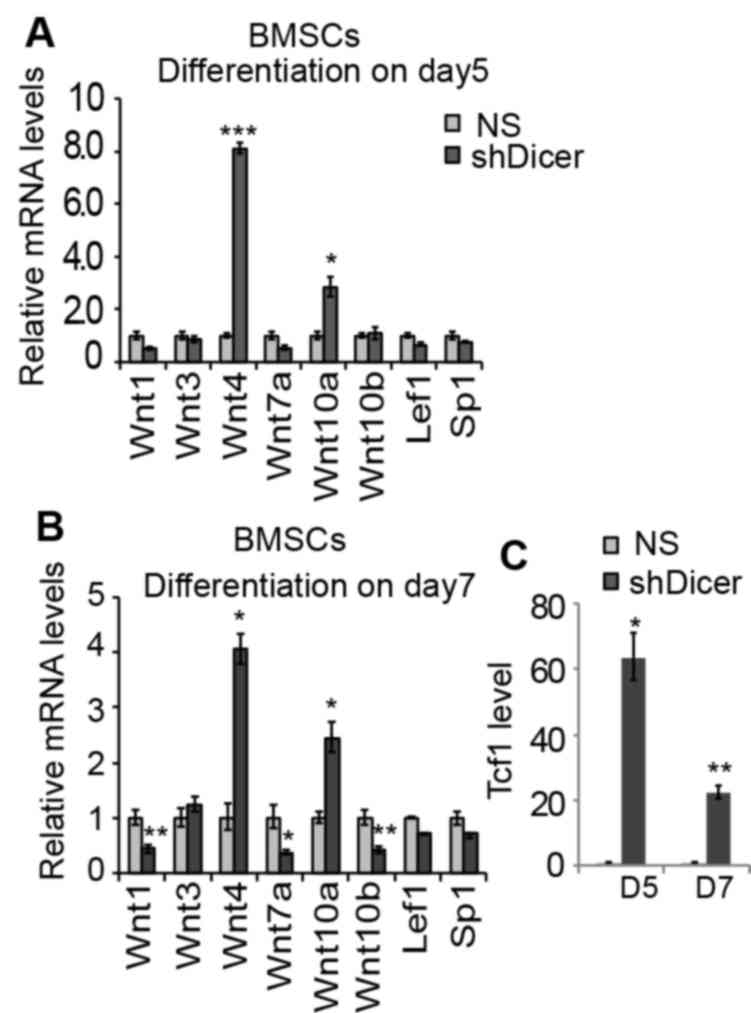
The effects of shDicer on the gene expression levels
of Wnt genes in differentiating BMSCs were also investigated. At
the day 5 time interval during the proliferative expansion period,
knockdown of Dicer suppressed the expression levels of Wnt1 and
Wnt7a by ~50%; whereas the expression levels of Wnt4, Wn10a and
Tcf-1 increased by 8.1-, 2.54- and 63.5-fold, respectively
(Fig. 3). Furthermore, at day 7
during differentiation, the expression levels of Wnt10b decreased
by 60% in shDicer-BMSCs compared with the NS group (Fig. 3B). The expression levels of other
Wnt components tested by qPCR was not altered. Transfection with
shDicer increased the expression of Wnt10a by 2.5-fold in BMSCs at
the 5 and 7 day time intervals compared with the NS group (Fig. 3A and B). The inhibition of Wnt1 and
Wnt7a by shDicer was also observed during early differentiation on
day 7; however, transfection with shDicer significantly increased
the expression levels of Wnt4 and Tcf-1 by 4- and 22-fold compared
with the NS group, respectively (Fig.
3B and C).
Considering that the knockdown of Dicer was revealed
to be associated with the regulation of Wnt genes and Tcf-1 at the
proliferation stage (day 5) and the early differentiation stage
(day 7), alterations in the expression levels of these genes in
shDicer-BMSCs at the 14 and 21 day time intervals during late
osteogenic differentiation were investigated. The results revealed
that knockdown of Dicer suppressed the expression levels of all
included Wnt genes by 50–90%, apart from Wnt10a (Fig. 4A and B). Notably, in contrast to
the activation of Tcf-1 by shDicer at the 5 and 7 day time
intervals, shDicer inhibited the expression levels of Tcf-1 by
>90% in BMSCs during the late stage of osteogenic
differentiation (Fig. 4A and B),
which may be due to the suppressed expression of numerous Wnt
genes, resulting in the inhibition of Tcf-1. Furthermore, knockdown
of Dicer was revealed to upregulate Wnt10a expression throughout
the osteogenic differentiation process, and this effect was most
significant at the 14 day time interval (>5.54 fold), which
suggested that Dicer may directly regulate the Wnt10a-targeting
miRNAs in order to mediate Wnt10a expression levels during the
osteogenic differentiation process. To investigate whether
shDicer-induced upregulation of Wnt10a is specific to osteogenic
differentiation, shDicer was additionally applied to mouse
fibroblasts (NIH3T3 cells). The results revealed that a total of
90% of Dicer was inhibited in transfected cells compared with the
NS group (Fig. 4C). Furthermore,
the expression levels of all of the examined Wnt genes, as well as
Tcf-1 and Lef1, were downregulated in NIH3T3 cells following
transfection with shDicer, thus suggesting that the regulation of
Wnt10a expression by Dicer is specific to the in vitro
osteogenic differentiation of BMSCs and osteoblasts (Fig. 4C). Furthermore, the expression
levels of SP1 were not affected by shDicer in either BMSCs or
fibroblasts, thereby suggesting that the upregulation of Wnt genes,
Tcf and Lef1 by Dicer is specific to osteoblasts MC3T3-1 (Figs. 3A, B and 4).
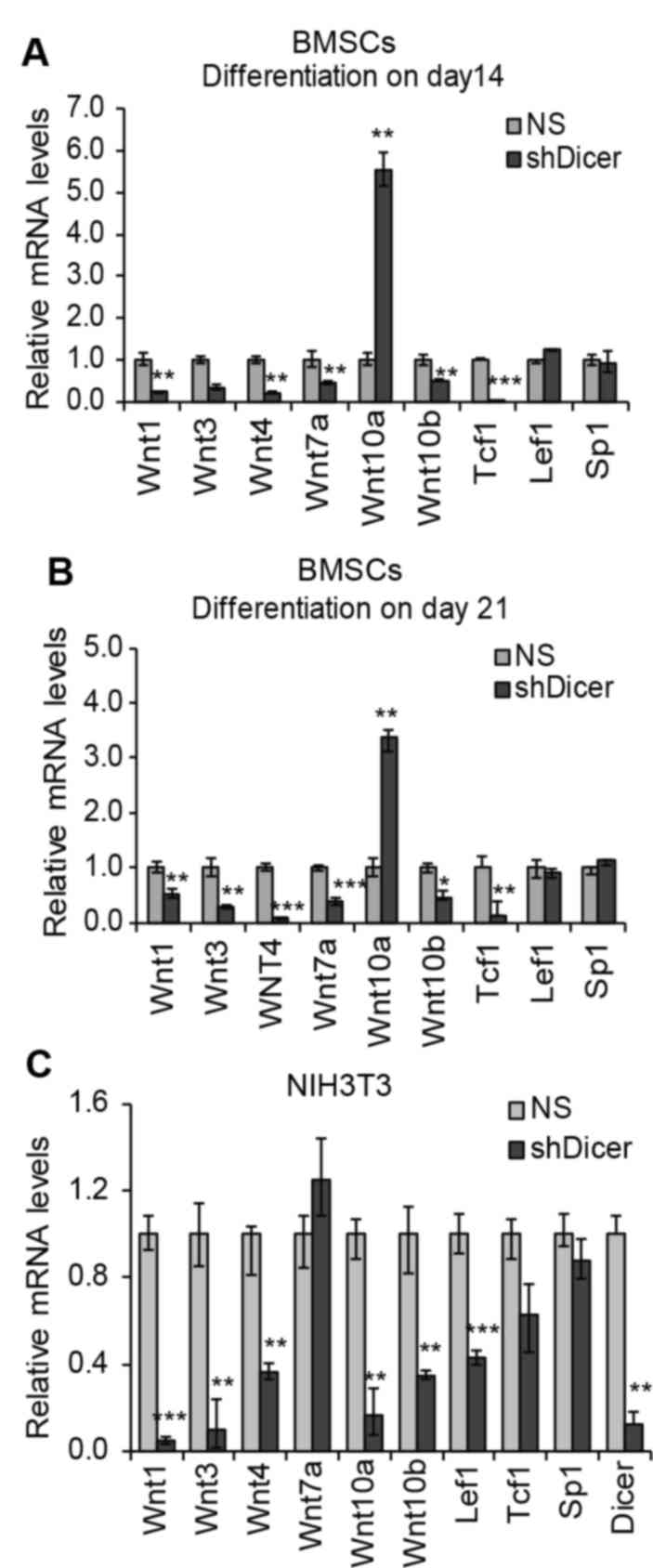 | Figure 4.Regulation of Wnt genes via knockdown
of Dicer during the late stage of osteogenic differentiation in
BMSCs. (A and B) In BMSCs transfected with NS shRNA and shDicer,
the expression levels of Wnt genes, Tcf-1, Lef1 and Sp1 were
detected on days (A) 14 and (B) 21 during late osteogenic
differentiation. (C) Expression levels of genes were investigated
in mouse fibroblast NIH3T3 cells transfected with shDicer. Gene
expression levels were investigated by reverse
transcription-quantitative polymerase chain reaction. Values are
presented as the mean ± standard error of the mean (n=3).
*P<0.05, **P<0.01 and ***P<0.001 vs. NS group. NS,
non-specific; BMSCs, bone marrow stromal cells; Tcf-1, T-cell
factor 1; Lef1, lymphoid enhancer binding factor 1; sh, short
hairpin; SP1, Sp1 transcription factor. |
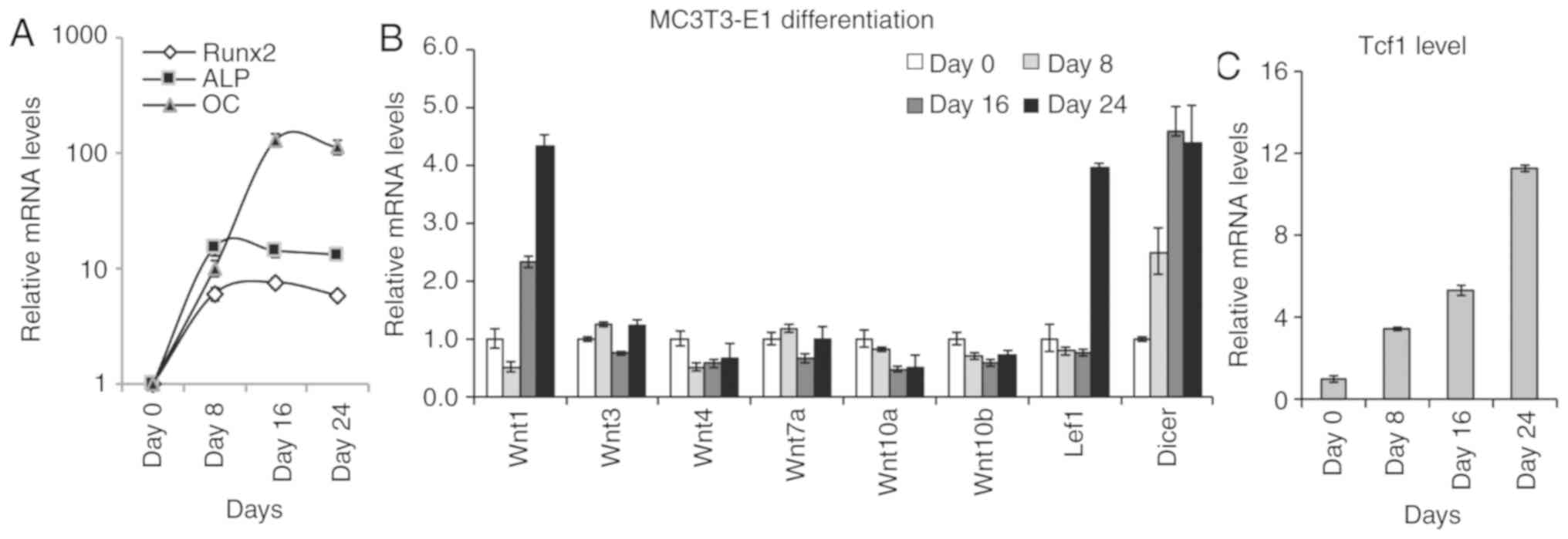
Profiling of Wnt genes in osteoblast
MC3T3-E1 cells
Considering that BMSCs have multi-lineage
differentiation potential (17),
MC3T3-E1 osteoblast cells were additionally investigated, as they
have the limited potential to differentiate selectively to mature
osteoblast cells. Differentiation was induced in MC3T3-E1
osteoblast cells, as assessed by a marked upregulation of Runx1,
ALP and Osteocalcin expression levels (Fig. 5A). The expression levels of Wnt1,
Tcf-1 and Lef1 were markedly upregulated; whereas the expression
levels of Wnt4 and Wnt10a were downregulated by 50% during the 24
days of differentiation (Fig. 5B and
C). Similar to BMSC osteogenic differentiation, the expression
level of Dicer was increased by 4.5-fold during the differentiation
period in MC3TC-E1 cells (Fig.
5B).
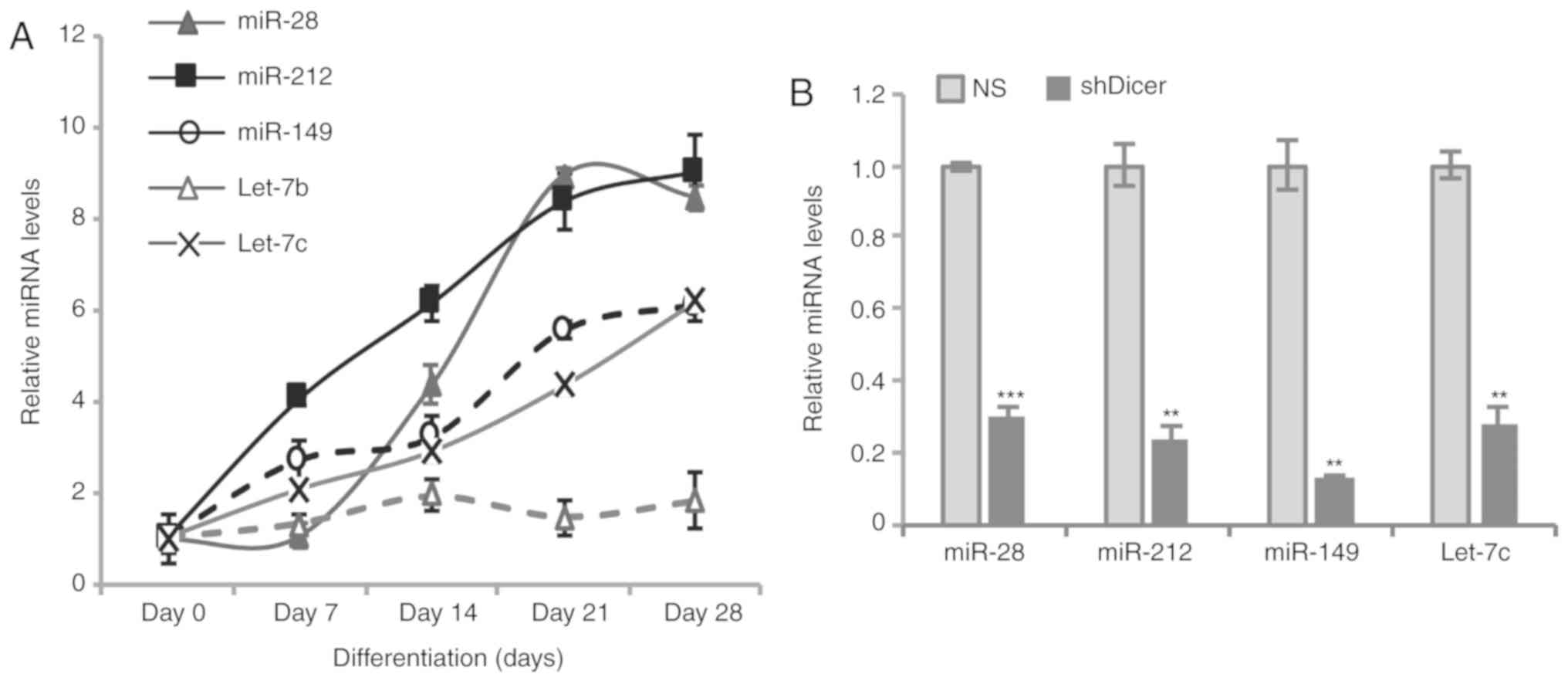
Expression levels of putative
Wnt10a-targeting miRNAs in BMSCs during osteogenic
differentiation
Dicer is an important enzyme for the production of
mature miRNAs. Therefore, deletion of Dicer may induce miRNA
deficiency during osteoblast differentiation, which may decrease
the suppression of Wnt10a expression by targeting miRNAs. Five
putative Wnt10a-targeting miRNAs were investigated during
osteogenic differentiation in BMSCs. These miRNAs were selected
based on the analysis using Targetscan and miRWalk prediction
software (38,39). In total, four of five miRNAs
(miR-28, miR-149, miR-212 and let-7b) were revealed to be markedly
upregulated during osteogenic differentiation (Fig. 6A). In addition, how the deletion of
Dicer affected the expression levels of the aforementioned miRNAs
was investigated. The results revealed that knockdown of Dicer
significantly suppressed the expression levels of the investigated
miRNAs in BMSCs (Fig. 6B).
Discussion
An increasing body of evidence has demonstrated that
miRNAs have important roles in bone development and homeostasis
(19). Supporting these findings,
it has also been previously reported that deletion of the key
enzyme Dicer in mice led to marked levels of abnormal bone
development (22). However, the
molecular mechanisms underlying such effects require further
investigation. In the present study, the dynamic changes associated
with numerous Wnt genes during osteogenic differentiation in mouse
BMSCs were investigated, as well as the regulation of these Wnt
genes and important transcriptional factors (Tcf-1 and Lef1) by
Dicer.
Extensive evidence has demonstrated that organ
development, tissue homeostasis and the development of numerous
diseases are associated with a number of signalling pathways, such
as inflammation, Wnt/β-catenin and metabolic dysregulation
(40–42). All of the Wnt genes included in the
present study have been reported to promote osteogenic
differentiation (43–47). To the best of the authors'
knowledge, the present study is the first to demonstrate that
various Wnt genes exhibit stage-specific patterns of expression
during osteogenic differentiation The activation of Wnt1 and Tcf-1
were observed in differentiating BMSCs and MC3T3-E1 osteoblasts,
thus suggesting that both Wnt1 and Tcf-1 have important roles in
osteoblast differentiation. The expression levels of Wnt3, Wnt7a
and Wnt10b were only upregulated in differentiating BMSCs, which
indicated that the upregulation of these genes may be important
regarding osteogenic differentiation. Wnt4, a prototypical ligand
for the non-canonical Wnt pathway, has been previously demonstrated
to promote osteoblast differentiation of mesenchymal stem cells
(48). Furthermore, in an in
vivo transgenic model, Wnt4 has been revealed to promote bone
formation and block bone resorption (49). The results of the present study
revealed that the expression levels of Wnt4 were decreased during
the proliferative expansion period and then significantly increased
during the differentiation stage. Notably, Wnt4 expression was
decreased during MC3T-E1 differentiation in the present study.
These results indicated that the upregulation of Wnt4 may be
involved in osteoblast proliferation; however, increased levels of
Wnt4 may not be essential for osteoblast differentiation. In
differentiating BMSCs, Wnt10a exhibited opposite expression
patterns to Wnt4. The result suggested that Wnt10a may be
associated with the proliferative expansion of BMSCs, but not with
differentiation. This hypothesis was further supported by the fact
that Wnt10a expression levels were downregulated during the
differentiation of osteoblasts in the present study.
In osteogenic and osteoblast differentiation models,
Dicer was upregulated, which suggested that it has an important
role during this biological process. Consistent with previous
studies, marked blockage of osteogenic differentiation in
shDicer-BMSCs was demonstrated in the present study (22). The differential regulation of the
included Wnt genes via knockdown of Dicer is coordinated well with
their dynamic expression pattern during differentiation. For
example, during the early stage of differentiation (days 5–7), Wnt4
and Tcf-1 levels were relatively low in undifferentiated BMSCs.
Knockdown of Dicer significantly increased Wnt4 and Tcf-1 levels on
days 5 and 7, which may be involved in cell proliferation. During
the late stage of differentiation, deficiency of Dicer resulted in
the downregulation of Wnt4 and Tcf-1 expression, delaying or
blocking osteogenic differentiation. The present results suggested
that Wnt4 and Tcf-1 may serve stage-specific roles.
Upregulation of Wnt10a via knockdown of Dicer may
represent the only protein included in the present study that
directly regulates miRNA deficiency. Activation of Wnt10a could
switch adipogenesis to osteogenesis (50); however, its function is not clear
during osteogenic differentiation. In the present study, numerous
putative Wnt10a-targeting miRNAs, which were upregulated during
osteoblast differentiation and inhibited via knockdown of Dicer,
were determined. In the future, the mechanisms associated with the
miRNAs targeting Wnt10a during osteogenesis require further
investigation. Previous studies identified that, besides directly
targeting Wnt genes, a number of Dicer-dependent miRNAs may target
inhibitors of Wnt genes, including Dickkopf-related protein 1 and
secreted frizzled-related protein 1 (51). Deletion of Dicer leads to a
decreased maturation of miRNAs that may target various Wnt
inhibitors, resulting in the downregulation of Wnt genes.
In conclusion, the dynamic expression pattern of
numerous Wnt genes during osteogenic differentiation was
investigated in the present study, as well as the direct and
indirect regulation of these genes by Dicer. Considering the
important roles of Wnt signaling in bone development, the results
suggested that the inhibition of Wnt signalling may have a major
effect on defective bone formation in Dicer-knockout or -knockdown
models. miRNAs generated by Dicer may represent important
regulators of Wnt signaling during osteogenic differentiation.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analysed during this study are
included in this published article.
Authors' contributions
HYW conceived the present study, provided resources,
was involved in the investigations, analysis and visualization of
data, supervised the study and wrote the manuscript. RB, TS and FX
performed the experiments and analysed the data. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was granted ethical approval by
the Fourth Hospital of Hebei Medical University (Shijiazhuang,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lian JB, Stein GS, Javed A, van Wijnen AJ,
Stein JL, Montecino M, Hassan MQ, Gaur T, Lengner CJ and Young DW:
Networks and hubs for the transcriptional control of
osteoblastogenesis. Rev Endocr Metab Disord. 7:1–16. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Soltanoff CS, Yang S, Chen W and Li YP:
Signaling networks that control the lineage commitment and
differentiation of bone cells. Crit Rev Eukaryot Gene Expr.
19:1–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maupin KA, Droscha CJ and Williams BO: A
comprehensive overview of skeletal phenotypes associated with
alterations in wnt/β-catenin signaling in humans and mice. Bone
Res. 1:27–71. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fahiminiya S, Majewski J, Mort J, Moffatt
P, Glorieux FH and Rauch F: Mutations in WNT1 are a cause of
osteogenesis imperfecta. J Med Genet. 50:345–348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Laine CM, Joeng KS, Campeau PM, Kiviranta
R, Tarkkonen K, Grover M, Lu JT, Pekkinen M, Wessman M, Heino TJ,
et al: WNT1 mutations in early-onset osteoporosis and osteogenesis
imperfecta. N Engl J Med. 368:1809–1816. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zheng HF, Tobias JH, Duncan E, Evans DM,
Eriksson J, Paternoster L, Yerges-Armstrong LM, Lehtimäki T,
Bergström U, Kähönen M, et al: WNT16 influences bone mineral
density, cortical bone thickness, bone strength, and osteoporotic
fracture risk. PLoS Genet. 8:e10027452012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kato M, Patel MS, Levasseur R, Lobov I,
Chang BH, Glass DA II, Hartmann C, Li L, Hwang TH, Brayton CF, et
al: Cbfa1-independent decrease in osteoblast proliferation,
osteopenia, and persistent embryonic eye vascularization in mice
deficient in Lrp5, a Wnt coreceptor. J Cell Biol. 157:303–314.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Riddle RC, Diegel CR, Leslie JM, Van
Koevering KK, Faugere MC, Clemens TL and Williams BO: Lrp5 and Lrp6
exert overlapping functions in osteoblasts during postnatal bone
acquisition. PloS One. 8:e633232013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cui Y, Niziolek PJ, MacDonald BT, Zylstra
CR, Alenina N, Robinson DR, Zhong Z, Matthes S, Jacobsen CM, Conlon
RA, et al: Lrp5 functions in bone to regulate bone mass. Nat Med.
17:684–691. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lu W, Yamamoto V, Ortega B and Baltimore
D: Mammalian Ryk is a Wnt coreceptor required for stimulation of
neurite outgrowth. Cell. 119:97–108. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He X, Semenov M, Tamai K and Zeng X: LDL
receptor-related proteins 5 and 6 in Wnt/beta-catenin signaling:
arrows point the way. Development. 131:1663–1677. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen DQ, Cao G, Chen H, Liu D, Su W, Yu
XY, Vaziri ND, Liu XH, Bai X, Zhang L and Zhao YY: Gene and protein
expressions and metabolomics exhibit activated redox signaling and
wnt/β-catenin pathway are associated with metabolite dysfunction in
patients with chronic kidney disease. Redox Biol. 12:505–521. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen L, Chen DQ, Wang M, Liu D, Chen H,
Dou F, Vaziri ND and Zhao YY: Role of RAS/Wnt/β-catenin axis
activation in the pathogenesis of podocyte injury and
tubulo-interstitial nephropathy. Chem Biol Interact. 273:56–72.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Komiya Y and Habas R: Wnt signal
transduction pathways. Organogenesis. 4:68–75. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hovanes K, Li TW, Munguia JE, Truong T,
Milovanovic T, Lawrence Marsh J, Holcombe RF and Waterman ML:
Beta-catenin-sensitive isoforms of lymphoid enhancer factor-1 are
selectively expressed in colon cancer. Nat Genet. 28:53–57. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Behrens J, von Kries JP, Kuhl M, Bruhn L,
Wedlich D, Grosschedl R and Birchmeier W: Functional interaction of
beta-catenin with the transcription factor Lef-1. Nature.
382:638–642. 1996. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang M, Chen DQ, Chen L, Cao G, Zhao H,
Liu D, Vaziri ND, Guo Y and Zhao YY: Novel inhibitors of the
cellular RAS components, poricoic acids, target Smad3
phosphorylation and Wnt/β-catenin pathway against renal fibrosis.
Br J Pharmacol. 175:2689–2708. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen H, Yang T, Wang MC, Chen DQ, Yang Y
and Zhao YY: Novel RAS inhibitor 25-O-methylalisol F attenuates
epithelial-to-mesenchymal transition and tubulo-interstitial
fibrosis by selectively inhibiting TGF-β-mediated Smad3
phosphorylation. Phytomedicine. 42:207–218. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Husain A and Jeffries MA: Epigenetics and
bone remodeling. Curr Osteoporos Rep. 15:450–458. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hassan MQ, Tye CE, Stein GS and Lian JB:
Non-coding RNAs: Epigenetic regulators of bone development and
homeostasis. Bone. 81:746–756. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ha M and Kim VN: Regulation of microRNA
biogenesis. Nat Rev Mol Cell Biol. 15:509–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gaur T, Hussain S, Mudhasani R, Parulkar
I, Colby JL, Frederick D, Kream BE, van Wijnen AJ, Stein JL, Stein
GS, et al: Dicer inactivation in osteoprogenitor cells compromises
fetal survival and bone formation, while excision in differentiated
osteoblasts increases bone mass in the adult mouse. Dev Biol.
340:10–21. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oommen S, Otsuka-Tanaka Y, Imam N,
Kawasaki M, Kawasaki K, Jalani-Ghazani F, Anderegg A, Awatramani R,
Hindges R, Sharpe PT and Ohazama A: Distinct roles of microRNAs in
epithelium and mesenchyme during tooth development. Dev Dyn.
241:1465–1472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li J, Zhang Y, Zhao Q, Wang J and He X:
MicroRNA-10a influences osteoblast differentiation and angiogenesis
by regulating β-catenin expression. Cell Physiol Biochem.
37:2194–2208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang X, Lin J, Wang G and Lu J:
MicroRNA-433-3p promotes osteoblast differentiation through
targeting DKK1 expression. PloS One. 12:e01798602017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Long H, Sun B, Cheng L, Zhao S, Zhu Y,
Zhao R and Zhu J: miR-139-5p represses BMSC osteogenesis via
targeting wnt/β-catenin signaling pathway. DNA Cell Biol.
36:715–724. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kureel J, John AA, Prakash R and Singh D:
MiR 376c inhibits osteoblastogenesis by targeting Wnt3 and
ARF-GEF-1-facilitated augmentation of beta-catenin transactivation.
J Cell Biochem. 119:3293–3303. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cao F, Zhan J, Chen X, Zhang K, Lai R and
Feng Z: miR-214 promotes periodontal ligament stem cell
osteoblastic differentiation by modulating Wnt/β-catenin signaling.
Mol Med Rep. 16:9301–9308. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bae JS, Gutierrez S, Narla R, Pratap J,
Devados R, van Wijnen AJ, Stein JL, Stein GS, Lian JB and Javed A:
Reconstitution of Runx2/Cbfa1-null cells identifies a requirement
for BMP2 signaling through a Runx2 functional domain during
osteoblast differentiation. J Cell Biochem. 100:434–449. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xu S, De Becker A, Van Camp B,
Vanderkerken K and Van Riet I: An improved harvest and in vitro
expansion protocol for murine bone marrow-derived mesenchymal stem
cells. J Biomed Biotechnol. 2010:1059402010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang M, Chen DQ, Wang MC, Chen H, Chen L,
Liu D, Zhao H and Zhao YY: Poricoic acid ZA, a novel RAS inhibitor,
attenuates tubulo-interstitial fibrosis and podocyte injury by
inhibiting TGF-β/Smad signaling pathway. Phytomedicine. 36:243–253.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dernowsek JA, Pereira MC, Fornari TA,
Macedo C, Assis AF, Donate PB, Bombonato-Prado KF, Passos-Bueno MR
and Passos GA: Posttranscriptional interaction between miR-450a-5p
and miR-28-5p and STAT1 mRNA triggers osteoblastic differentiation
of human mesenchymal stem cells. J Cell Biochem. 118:4045–4062.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dole NS and Delany AM: MicroRNA variants
as genetic determinants of bone mass. Bone. 84:57–68. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
de la Rica L, García-Gómez A, Comet NR,
Rodríguez-Ubreva J, Ciudad L, Vento-Tormo R, Company C,
Álvarez-Errico D, García M, Gómez-Vaquero C and Ballestar E:
NF-κB-direct activation of microRNAs with repressive effects on
monocyte-specific genes is critical for osteoclast differentiation.
Genome Biol. 16:22015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yuan H, Zhao H, Wang J, Zhang H, Hong L,
Li H, Che H and Zhang Z: MicroRNA let-7c-5p promotes osteogenic
differentiation of dental pulp stem cells by inhibiting
lipopolysaccharide-induced inflammation via HMGA2/PI3K/Akt signal
blockade. Clin Exp Pharmacol Physiol. 21:1440–1681. 2018.
|
|
37
|
Wei J, Li H, Wang S, Li T, Fan J, Liang X,
Li J, Han Q, Zhu L, Fan L and Zhao RC: let-7 enhances osteogenesis
and bone formation while repressing adipogenesis of human
stromal/mesenchymal stem cells by regulating HMGA2. Stem Cells Dev.
23:1452–1463. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dweep H, Sticht C, Pandey P and Gretz N:
miRWalk-database: prediction of possible miRNA binding sites by
‘walking’ the genes of three genomes. J Biomed Inform. 44:839–847.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
ELife. 4:e050052015. View Article : Google Scholar :
|
|
40
|
Nusse R and Clevers H: Wnt/β-catenin
signaling, disease, and emerging therapeutic modalities. Cell.
169:985–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao YY, Cheng XL, Vaziri ND, Liu S and
Lin RC: UPLC-based metabonomic applications for discovering
biomarkers of diseases in clinical chemistry. Clin Biochem.
47:16–26. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen L, Su W, Chen H, Chen DQ, Wang M, Guo
Y and Zhao YY: Proteomics for biomarker identification and clinical
application in kidney disease. Adv Clin Chem. 85:91–113. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cawthorn WP, Bree AJ, Yao Y, Du B, Hemati
N, Martinez-Santibañez G and MacDougald OA: Wnt6, Wnt10a and Wnt10b
inhibit adipogenesis and stimulate osteoblastogenesis through a
β-catenin-dependent mechanism. Bone. 50:477–489. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shao JS, Cheng SL, Pingsterhaus JM,
Charlton-Kachigian N, Loewy AP and Towler DA: Msx2 promotes
cardiovascular calcification by activating paracrine Wnt signals. J
Clin Invest. 115:1210–1220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Adamska M, MacDonald BT, Sarmast ZH,
Oliver ER and Meisler MH: En1 and Wnt7a interact with Dkk1 during
limb development in the mouse. Dev Biol. 272:134–144. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Minear S, Leucht P, Jiang J, Liu B, Zeng
A, Fuerer C, Nusse R and Helms JA: Wnt proteins promote bone
regeneration. Sci Transl Med. 2:29ra302010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rauch F: The brains of the bones: How
osteocytes use WNT1 to control bone formation. J Clin Invest.
127:2539–2540. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chang J, Sonoyama W, Wang Z, Jin Q, Zhang
C, Krebsbach PH, Giannobile W, Shi S and Wang CY: Noncanonical
Wnt-4 signaling enhances bone regeneration of mesenchymal stem
cells in craniofacial defects through activation of p38 MAPK. J
Biol Chem. 282:30938–30948. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yu B, Chang J, Liu Y, Li J, Kevork K,
Al-Hezaimi K, Graves DT, Park NH and Wang CY: Wnt4 signaling
prevents skeletal aging and inflammation by inhibiting nuclear
factor-κB. Nat Med. 20:1009–1017. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chen YS, Wu R, Yang X, Kou S, MacDougald
OA, Yu L, Shi H and Xue B: Inhibiting DNA methylation switches
adipogenesis to osteoblastogenesis by activating Wnt10a. Sci Rep.
6:252832016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kawano Y and Kypta R: Secreted antagonists
of the Wnt signalling pathway. J Cell Sci. 116:2627–2634. 2003.
View Article : Google Scholar : PubMed/NCBI
|