Introduction
Ulcerative colitis (UC) is a major type of
inflammatory bowel disease (IBD), which are a group of diseases
characterized by a chronic and relapsing inflammatory disorder of
the large intestine with subsequent injury and disruption of the
mucosal barrier (1). Bloody
mucopurulent stool, diarrhea, abdominal pain, and fistulization are
the most common symptoms of UC (2,3).
Although numerous studies suggest that genetic, immunological and
environmental factors are involved in the development of UC
(3–5), its cause and potential underlying
mechanisms are still not clearly understood. In addition, the types
of drugs available for the treatment of UC at present are not
abundant and their effects are unsatisfactory; thus, there remains
a need to develop novel, effective drugs (6,7).
Previous studies have revealed that the dysregulation of
inflammatory pathways and autophagy may be involved in the
pathogenesis of UC, and thus may be novel therapeutic targets for
UC treatment (8,9).
Curcumin is the main component of turmeric
(Curcuma longa) (10).
Curcumin has attracted considerable attention due to its
anti-inflammatory, anti-tumor and anti-oxidant properties (11–13).
The anti-inflammatory properties of curcumin have resulted in its
use in the treatment of colitis, and previous studies have
demonstrated that it is able to attenuate inflammation associated
with experimental colitis (14,15)
and is effective in patients with UC (16,17).
Resveratrol, a natural polyphenolic compound present in grapes and
red wine, has been reported to exhibit a variety of anti-oxidant,
anti-platelet aggregation, anti-tumor and anti-inflammatory
effects, in addition to cardioprotective effects (18,19).
Due to the anti-inflammatory characteristics of resveratrol, its
use may be therapeutically beneficial in the treatment of a number
of different diseases, including colitis (20), arthritis (21) and pancreatitis (22). However, the exact molecular
mechanisms involved remain unclear.
Autophagy is an intracellular pathway responsible
for turnover of long-lived proteins and damaged organelles to
maintain cellular homeostasis (23). Autophagy dysregulation is
associated with an alteration of both innate and adaptive immune
responses, which has been shown to be involved in many diseases
including IBD (24,25). The present study evaluated the
effect of curcumin and resveratrol on dextran sulfate sodium
(DSS)-induced colitis in BALB/c mice and preliminarily investigated
the autophagy-associated mechanisms.
Materials and methods
Animals
Male BALB/c mice (7–8 weeks old), weighing 20–22 g
were purchased from Shanghai SLAC Laboratory Animal Co., Ltd.
(Shanghai, China) and fed in a controlled environment at a
temperature of 25±2°C and a 12 h light/dark cycle at 50–70%
humidity. Animals were acclimatized for 1 week prior to the
initiation of the study and were maintained with ad libitum
access to standard laboratory chow and water. All animal procedures
were ethically approved by the Institutional Animal Care and Use
Committee of Qingdao Municipal Hospital (Qingdao, China).
Experimental design
A total of 80 mice were randomly divided into four
groups: Control group, DSS group, curcumin-treated (DSS + Cur)
group, and resveratrol-treated (DSS + Res) group, with 20 mice per
group. In the control group, mice were fed with a standard diet
throughout the course of the experiment (14 days). In the DSS
group, the mice received a standard diet for 14 days in addition to
DSS (3.5% w/v) during the first 7 days of the experiment (from day
1 to 7). In the DSS + Cur group, the mice received the standard
diet supplemented with 50 mg/kg curcumin for 14 days in addition to
3.5% DSS during the first 7 days of the study. In the DSS + Res
group, the mice received the standard diet supplemented with 80
mg/kg resveratrol for 14 days in addition to 3.5% DSS during the
first 7 days.
Assessment of colitis
The mice were assessed daily for colitis development
by monitoring body weight, gross rectal bleeding, stool consistency
and survival. Mice were sacrificed by cervical dislocation at day
15 or judged as moribund (inability or unwillingness to walk,
inability to reach water or food, palpable hypothermia, or lack of
overt response to manipulation) before day 15 and immediately
sacrificed, and the colons were removed, length and weight were
measured. Scoring systems are used to assess the severity of
overall disease (26), and the
disease activity index (DAI) was calculated daily for each mouse.
In brief, the scoring was as follows: 0, no weight loss, no occult
blood in the stools and normal stool consistency; 1, weight loss of
1–5% of total body mass, no occult blood and normal stool
consistency; 2, 5–10% weight loss of total body mass, positive for
fecal occult blood and loose stools; 3, 10–20% weight loss of total
body mass, positive for fecal occult blood and loose stools; and 4,
>20% weight loss of total body mass, gross rectal bleeding and
diarrhea.
Histopathological examinations
Mice were sacrificed on day 8 or 15 by cervical
dislocation, and the length of the colon was measured. Samples for
histology were excised from the distal 6–8 cm of the colon, fixed
in 10% formalin overnight at room temperature and embedded in
paraffin blocks. Paraffin blocks were sliced into sections, 4 µm in
thickness, and stained with hematoxylin for 6 min and eosin (cat.
no. C0105; Beyotime Biotechnology, Shanghai, China) for 1 min at
room temperature.
Measurement of cytokines
The concentrations of tumor necrosis factor-α
(TNF-α) and interleukin-6 (IL-6) in the culture supernatants of the
colon tissues were measured using a Bio-Rad Multiplex bead array
instrument and cytokine kits (cat. no. #7050; Bio-Rad Laboratories,
Inc., Hercules, CA, USA) according to the manufacturer's
protocol.
Immunofluorescence staining
Double immunofluorescence staining for
autophagy-related 12 (Atg12), Beclin-1, microtubule-associated
protein light chain 3 (LC3)II, phospho-mechanistic target of
rapamycin (mTOR) and sirtuin 1 (SIRT1) were performed on the
sections. Paraffin sections were deparaffinized with xylene and
rehydrated. After endogenous peroxidase activity was blocked with
3% H2O2 for 10 min at room temperature, the
sections were treated with 0.01 mol/l citrate (pH 6.0) in a 500-W
microwave oven for 15 min for antigen retrieval. Subsequently,
sections were blocked with normal goat serum (cat. no. 16210-064;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) for 1 h,
and incubated with the following primary antibodies overnight at
4°C: Atg12 (cat. no. ab155589), Beclin-1 (cat. no. ab62557), LC3B
(cat. no. ab48394), mTOR (phospho S2448; cat. no. ab84400) and
SIRT1 (cat. no. ab32441; all 1:1,000 dilution; all from Abcam,
Cambridge, MA, USA). Then, Alexa Fluor 594 (red)-conjugated goat
anti-rabbit immunoglobulin G secondary antibody (1:1,000; cat. no.
A-11032; Molecular Probes, Eugene, OR, USA) was incubated with the
sections for 1 h at room temperature. Cell nuclei were stained with
DAPI (1 µg/ml; cat. no. 12131; Roche Diagnostics, Basel,
Switzerland) for 5 min at room temperature. Images were obtained
using a fluorescence microscope (Leica SP-8; Leica Microsystems
GmbH, Wetzlar, Germany).
Western blot analysis
Colon samples were homogenized on ice in
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) containing phenylmethylsulfonyl
fluoride and protease inhibitor. The protein concentrations of the
samples were determined using the Bradford method (using a Bio-Rad
Protein assay; Bio-Rad Laboratories, Inc.). Equivalent amounts of
protein (50 µg) from each sample were separated on 12% sodium
dodecyl sulfate-polyacrylamide gels, and fractioned proteins were
transferred onto 0.22 µM nitrocellulose membranes (EMD Millipore,
Billerica, MA, USA) at 75 V for 45 min. Subsequently, the membranes
were blocked with Tris-buffered saline solution containing 5%
nonfat dried milk at room temperature for 1 h, and then incubated
with specific antibodies against Atg12 (cat. no. 2011), Beclin-1
(cat. no. 3495), LC3II (cat. no. 4108), mTOR (cat. no. 2972) and
SIRT1 (cat. no. 8469; all 1:1,000 dilution; all from Cell Signaling
Technology, Inc., Danvers, MA, USA) at 4°C overnight and then
incubated with horseradish peroxidase-conjugated secondary antibody
(cat. no. 7074; Cell Signaling Technology, Inc.) for 1 h at room
temperature. Bands were visualized using a chemiluminescence
detection system (cat. no. #34095; Pierce; Thermo Fisher
Scientific, Inc.) and quantified with Quantity One Software 4.6.7
(Bio-Rad Laboratories, Inc.).
Statistical analysis
Data analysis was performed using GraphPad Prism
software (version 4.03; GraphPad Software, Inc., La Jolla, CA,
USA). Parametric data are presented as the mean ± standard error of
at least three independent experiments. The statistical
significance of any difference in each parameter among the groups
was evaluated using one-way analysis of variance followed by
Tukey's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of curcumin or resveratrol on
survival, body weight and the clinical symptoms of mice
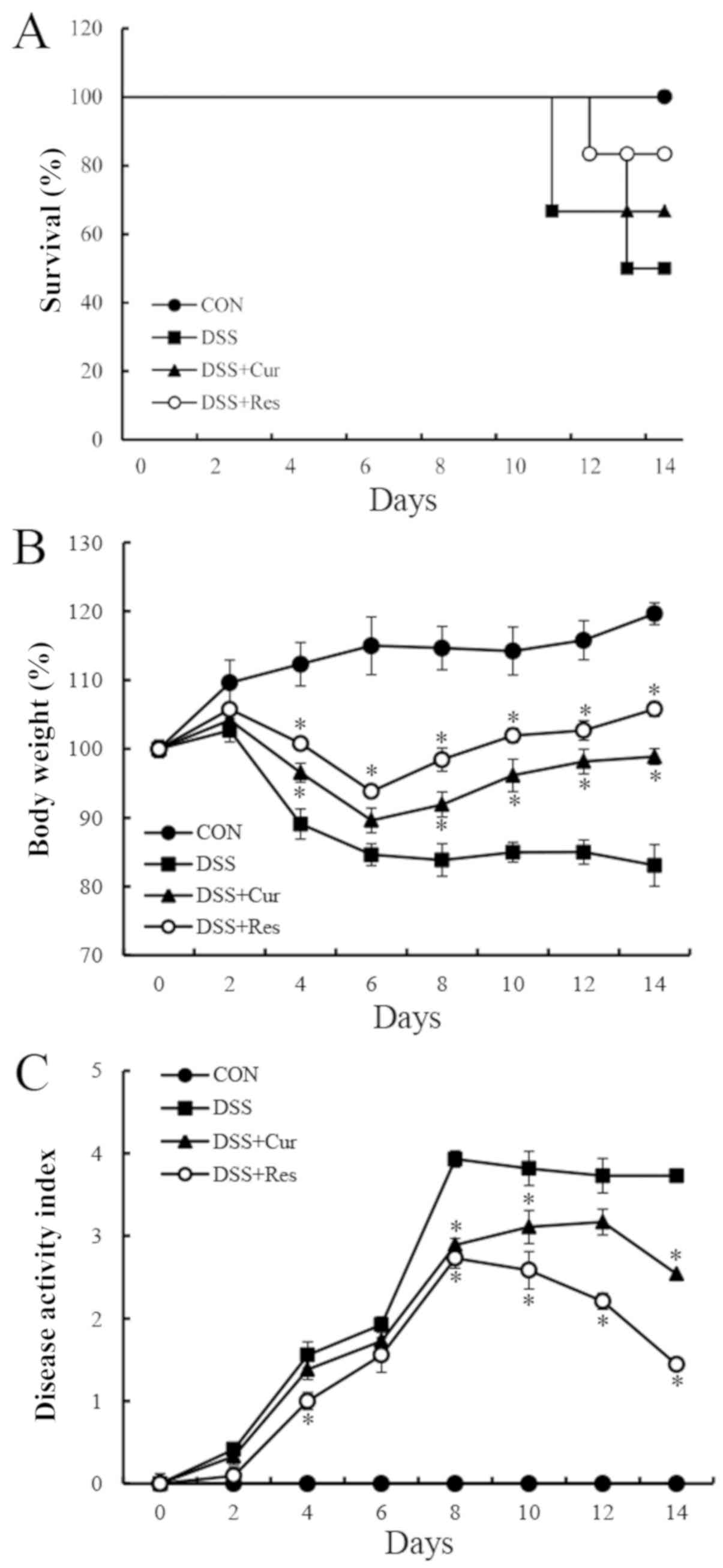
Symptomatic parameters, including the survival rate,
body weight loss and DAI, caused by colitis in mice were recorded.
On day 15, six mice were randomly selected from each group for body
weight and DAI analysis. As presented in Fig. 1A, DSS exposure resulted in a 50%
mortality rate at day 14, which was similar to the data reported in
a previous study (27). However,
the survival rate of the DSS-treated mice was improved in the
curcumin (66.7%) and resveratrol (83.3%)-treated groups. In the
DSS-treated group, a loss of body weight was recorded from day 4
onwards following the administration of DSS (4%) and this loss
remained significantly higher than the normal mice until day 8
(83.85±2.36%) (P<0.05). The administration of curcumin or
resveratrol significantly reduced the body weight loss in mice with
colitis compared with the DSS-treated mice (P<0.05; Fig. 1B). Another common feature of the
DSS-induced model of colitis is an increase in the DAI (28). Curcumin or resveratrol treatment
significantly ameliorated diarrhea and rectal bleeding compared
with the DSS alone-treated group (P<0.05; Fig. 1C).
Effect of curcumin or resveratrol on
the colon length and histopathology of mice with colitis
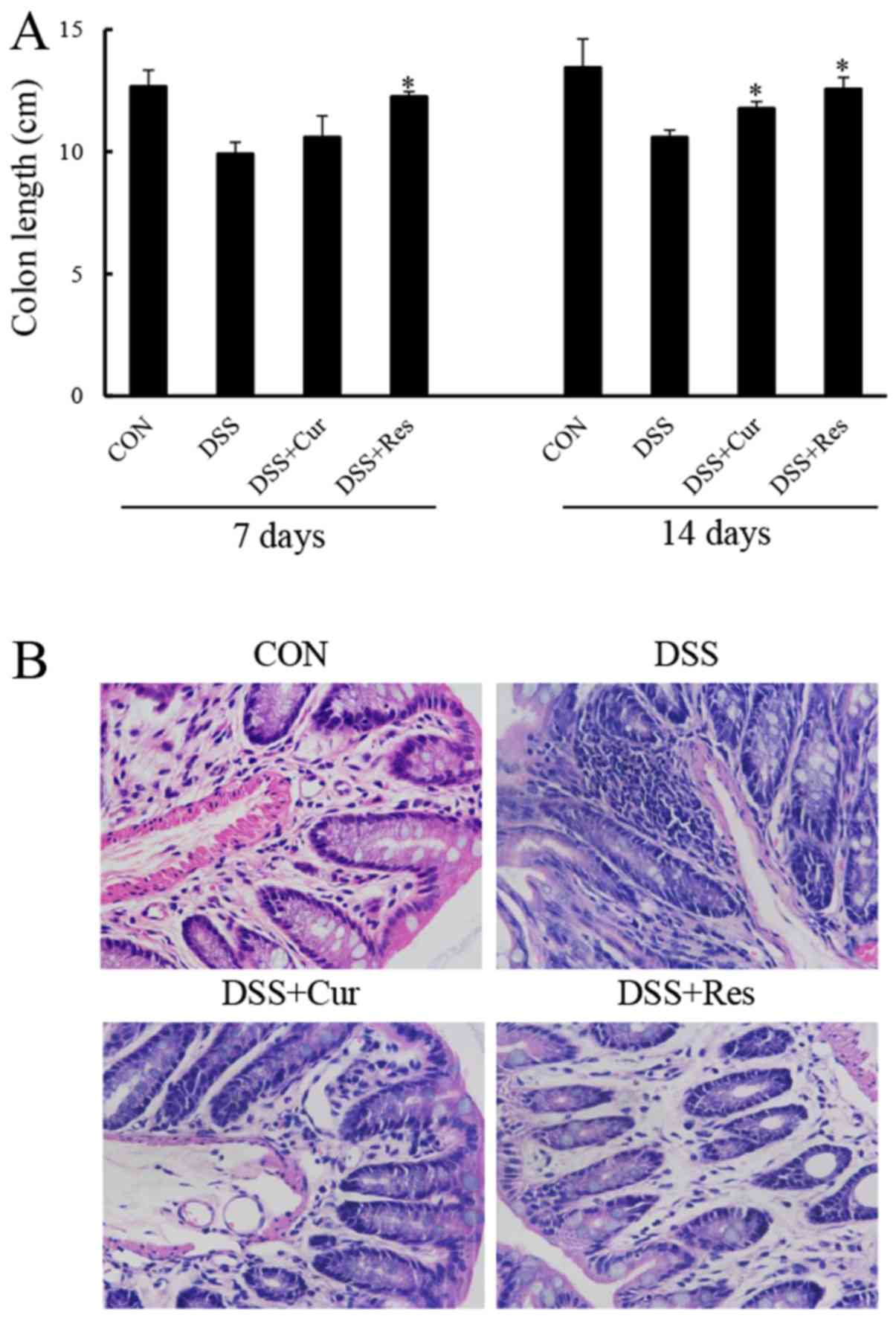
Colon tissues were collected and measured at days 7
and 14, in order to study the effect of curcumin or resveratrol on
the inflammation-induced decrease in colon length, a morphological
parameter used to assess the degree of inflammation in the
DSS-induced colitis mouse model. The colon length of the mice
exposed to DSS was significantly shorter compared with that of the
control mice (P<0.05; Fig. 2).
The colon of mice treated with DSS and curcumin, or DSS and
resveratrol exhibit a statistically significant increase in colon
length when compared with the DSS group and almost completely
reverted to a normal length (P<0.05).
Histological analysis of the distal colon tissue
revealed that the treatment of mice with DSS resulted in the
destruction of colonic architecture with ulcerations, crypt
dilation, goblet cell depletion, a disrupted epithelial layer and
intense infiltration of the inflammatory cells compared with the
normal morphology of colon tissue (Fig. 2B). By contrast, colon tissues from
the curcumin or resveratrol-treated mice exhibited predominantly
intact colon histology, with a preserved epithelial layer and crypt
structure, and reduced numbers of infiltrating cells (Fig. 2B).
Effect of curcumin and resveratrol on
the production of inflammatory cytokines in the colon tissue of
mice with colitis
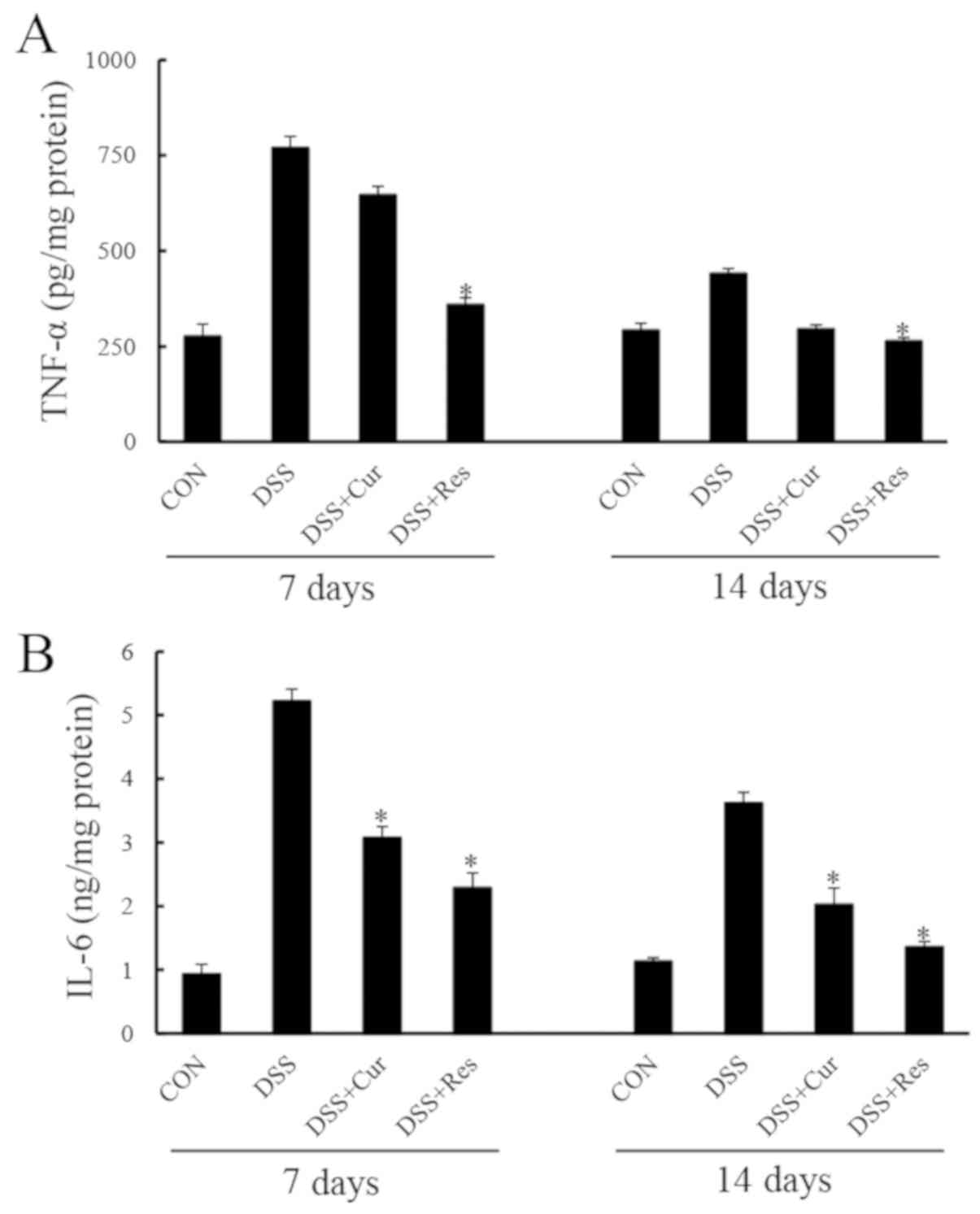
Colonic TNF-α and IL-6 levels were significantly
elevated in the DSS group compared with the controls (P<0.05;
Fig. 3). Curcumin or resveratrol
administration prevented significant increases in TNF-α and IL-6
expression levels at day 7 (P<0.05). Similar effects of curcumin
or resveratrol treatment were observed at day 14.
Effect of curcumin and resveratrol on
autophagy in DSS-induced UC in mice
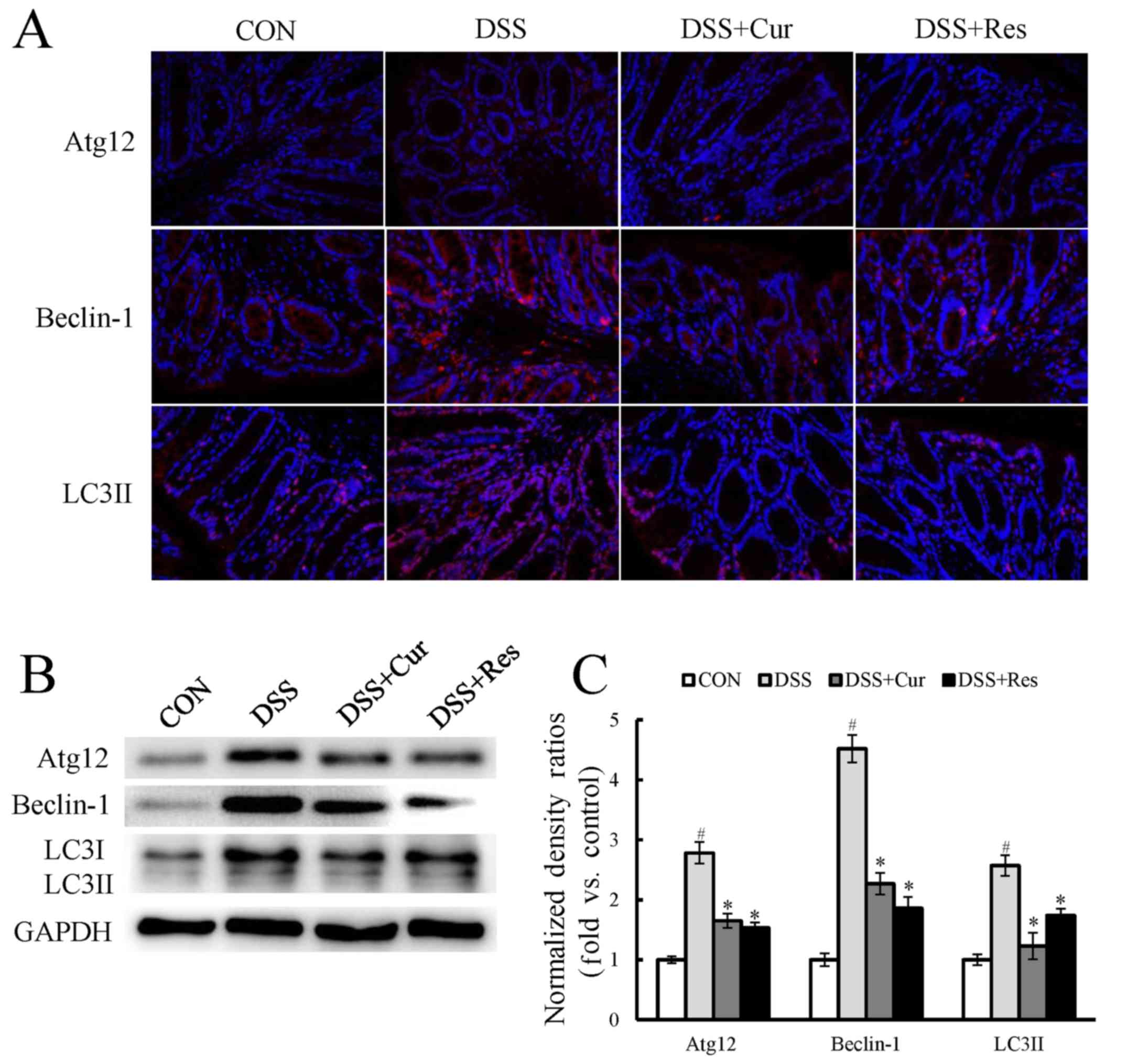
Immunofluorescence staining for components of
cellular autophagy signaling pathways, including Atg12, Beclin-1
and LC3II, was performed on colon tissues. As presented in Fig. 4A, a marked increase in the
expression of the autophagy-associated proteins Atg12, Beclin-1 and
LC3II was observed in the DSS-treated mice when compared with the
control mice (P<0.05). LC3I expression also appeared to
increase. Treatment with curcumin or resveratrol significantly
reversed the changes in the levels of autophagy-associated genes
when compared with the DSS-treated mice (P<0.05). To further
confirm this observation, the LC3II, Atg12 and Beclin-1 expression,
and the LC3II/I ratio were assessed using western blot analysis
(Fig. 4B). The expression levels
of Atg12, Beclin-1 and LC3II were significantly increased by
exposure to DSS (P<0.05), and curcumin or resveratrol treatment
significantly reduced their expression.
Effect of curcumin and resveratrol on
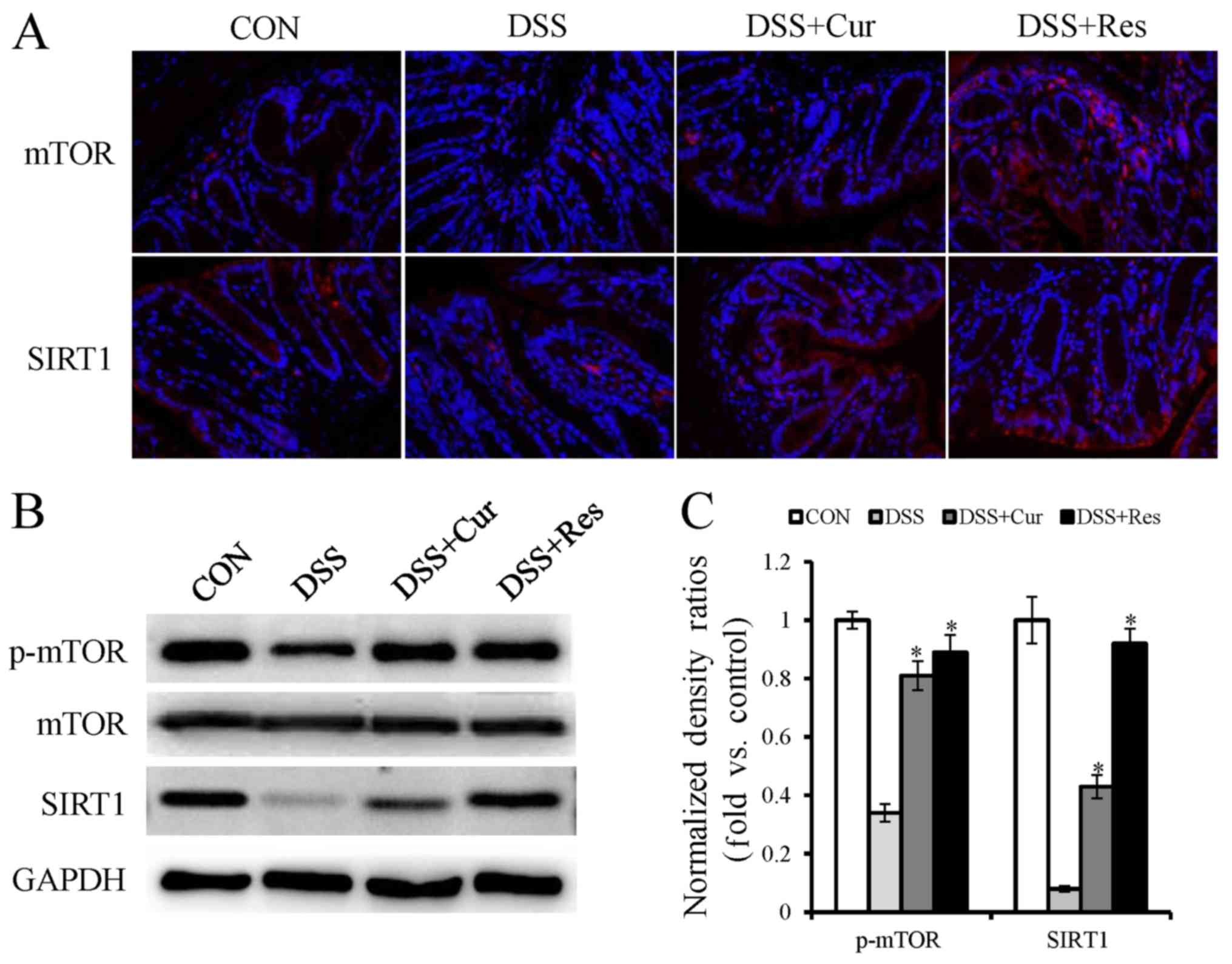
mTOR and SIRT1 expression in DSS-induced UC in mice
Furthermore, the present study intended to
investigate whether the inhibition of autophagy was mediated via
mTOR and SIRT1 activation. As determined by immunofluorescence
staining and western blot analysis, a substantial decrease in the
protein expression levels of phospho-mTOR and SIRT1 were observed
in the DSS group compared with the normal mice (Fig. 5A and B). Additionally, phospho-mTOR
and SIRT1 demonstrated a higher expression in the curcumin or
resveratrol-treated groups compared with the DSS alone treated
group.
Discussion
Administration of DSS to animals is a frequently
used as a model of UC, as the resulting pathological features
correspond well to those of UC in humans (29). In the present study, a UC model was
successfully established by treating mice with 3.5% DSS for 7 days,
and then the protective effects of curcumin and resveratrol were
investigated. Following the administration of DSS, the mice
exhibited considerable clinical signs of colitis, including weight
loss, crypt architectural distortion, colonic epithelial injury,
reduced colon length and infiltration of the inflammatory cells.
These macroscopic and microscopic alterations were consistent with
the findings of previous studies (30,31),
and were counteracted by the administration of curcumin and
resveratrol. In the survival experiment, 50 mg/kg curcumin or 80
mg/kg resveratrol treatment prolonged animal survival compared with
the DSS treatment alone. These observations clearly suggested that
curcumin and resveratrol suppress the overt clinical features of
DSS-induced colitis.
Curcumin and resveratrol treatment in the present
study resulted in the preservation of histological integrity in the
colon tissue. The DSS-induced mice developed immunological
deregulation (32), including the
prominent shortening of the large intestine, thickened muscular
layer, crypt damage and cellular infiltration in the inflamed
colon; however, these effects were reduced by curcumin and
resveratrol treatment.
In addition, curcumin and resveratrol treatment
significantly decreased the accumulation of pro-inflammatory
cytokines as compared with those in the colon tissue of DSS-treated
mice. Inflammatory responses have a pivotal role in the
pathogenesis of UC (33,34). A number of previous studies have
revealed that the interaction between inflammatory pathways and the
intestinal mucosal immune system may result in the disruption of
tight junction proteins in the intestinal epithelium and affect
intestinal homeostasis (35,36).
A number of studies support the idea that TNF-α and IL-6 are
important pathological mediators of IBD (37–39).
Increased serum and tissue levels of TNF-α and IL-6 are
characteristic features of colitis and numerous other chronic
inflammatory diseases (40). Among
these cytokines, the overexpression of TNF-α is vital in intestinal
mucosal impairment (41).
Adalimumab, a TNF-α blocker, has been successfully used for the
treatment of patients with IBD in a clinical setting (42). Another anti-TNF-α antibody,
infliximab, was also reported to exert an effective therapeutic
effect on UC in one clinical case (43). In addition, IL-6 is a key mediator
of the progression of UC. Inhibition the action of IL-6 may
attenuate the severity of diarrhea and reduce the infiltration of
inflammatory cells into the intestinal tissue (44,45).
In the present study, TNF-α and IL-6 production in DSS-exposed
colons were substantially higher at 14 days post-DSS compared with
curcumin or resveratrol-treated mice, which is consistent with the
delayed recovery from inflammation in these mice. These results are
supported by previous in vivo and in vitro
experiments where curcumin or resveratrol have been demonstrated to
suppress the chronic inflammation through the inhibition of these
inflammatory mediators (46–48).
Altogether, the present results suggest that the protective effect
of curcumin and resveratrol against colonic injury may be due to
the regulation of TNF-α and IL-6.
Autophagy is a degradative pathway involved in
recycling long-lived proteins and damaged organelles in order to
maintain cellular homeostasis. It is regulated by
autophagy-specific genes that include the Beclin-1 complex, Atg12
and LC3 systems (49).
Additionally, the dysregulation of autophagy has been shown to be
involved in multiple diseases, such as liver disease,
cardiomyopathy, diabetes, neurodegenerative disorders, autoimmune
diseases, malignancies and IBD (24,50,51).
Autophagy is regulated by autophagy specific genes, including
Beclin-1, which initiates autophagosome formation via the
phosphatidylinositol 3-kinase signaling pathway (25). Autophagosome formation is further
associated with the conversion of cytosolic-associated protein
light chain 3 (LC3-I) to the membrane bound LC3-II form (52). LC3-II is most commonly used for
autophagy assays since it is the only protein marker that is
reliably associated with the number of autophagosomes (52). However, LC3-II itself is degraded
by autophagy (53). The autophagy
factor Atg12-Atg5 conjugate has a novel E3-like activity for
facilitating the lipidation of members of the LC3 family (54). The results of the present study
demonstrated that LC3I, Beclin-1, and the Atg12 protein were highly
expressed in the colon of DSS-exposed mice, compared with the
control animals. Decreased LC3I, Beclin-1 and Atg12 protein
expression was detected in curcumin and resveratrol-treated mice
compared with the DSS-treated animals. Upon autophagy induction,
LC3-I is cleaved to LC3-II. However, the change of LC3-II in the
study is far lower than LC3I, which may be due to the degradation
by autophagy (53).
Autophagy is regulated by important nutrient-sensing
pathways, including the mTOR and SIRT1 pathways (55,56).
mTOR is an evolutionarily conserved serine/threonine kinase which
is a member of the phosphoinositide 3-kinase-associated kinase
group, and has emerged as a master regulator of cellular metabolism
and promotes cell growth in response to environmental cues
(57). Deregulation of the mTOR
pathway has been implicated in a number of human diseases including
diabetes, neurodegenerative diseases, cancer and IBD (57). Tuberous sclerosis complex (TSC) and
lymphangioleio-myomatosis are associated with the deregulation of
the raptor-mTOR pathway, which are likely caused by mutations in
TSC1 or TSC2 tumor suppressors (58). As a consequence, drugs that target
mTOR are used therapeutically (59). SIRT1 is a nuclear class III
deacetylase and has been demonstrated to extend lifespan in a
number of different species (60).
Furthermore, SIRT1 has been reported to improve the symptoms of
inflammation-associated diseases, including chronic obstructive
pulmonary disease, cardiovascular diseases, diabetes, renal
diseases and IBD (60,61). In the present study, the expression
of phosphorylated mTOR and SIRT1 was greatly increased by curcumin
and resveratrol in mice with DSS-induced colitis compared with the
DSS alone treated mice, suggesting that the anti-UC effect of
curcumin and resveratrol may be due to the promotion of mTOR and
SIRT1 signaling activation.
In conclusion, the present study is, to the best of
our knowledge, the first to demonstrate that the development and
progression of colitis symptoms caused by DSS administration are
attenuated by curcumin and resveratrol. The underlying molecular
mechanisms involved in inhibiting inflammatory responses, reducing
autophagy-associated gene expression and promoting autophagy
signaling activation were elucidated. Collectively, as relatively
non-toxic natural products combined with substantial
anti-inflammatory activity, curcumin and resveratrol have the
potential to function as effective anti-IBD therapeutic
methods.
Acknowledgements
Not available.
Funding
The present study was supported by the Project of
Traditional Chinese Medicine Technology Development Program in the
Shandong Province (grant no. 2017-201).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
Conception and design, LZ, DZ; data acquisition, LZ,
HX, GZ; data analysis and interpretation, CQ, XS, CP; manuscript
writing, LZ; final approval of manuscript, all authors. All authors
are accountable for all aspects of the study.
Ethics approval and consent to
participate
This study has been approved by the Institutional
Animal Care and Use Committee of Qingdao Municipal Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Baumgart DC and Sandborn WJ: Inflammatory
bowel disease: Clinical aspects and established and evolving
therapies. Lancet. 369:1641–1657. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stenson WF, Tremaine WJ and Cohen RD:
Ulcerative Colitis: Clinical manifestations and management. In:
Yamada's Atlas of Gastroenterol. John Wiley & Sons, Ltd. (New
York, NY). 216–224. 2016.
|
|
3
|
Baumgart DC and Carding SR: Inflammatory
bowel disease: Cause and immunobiology. Lancet. 369:1627–1640.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hou JK, Abraham B and El-Serag H: Dietary
intake and risk of developing inflammatory bowel disease: A
systematic review of the literature. Am J Gastroenterol.
106:563–573. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eaden JA, Abrams KR and Mayberry JF: The
risk of colorectal cancer in ulcerative colitis: A meta-analysis.
Gut. 48:526–535. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Laharie D, Bourreille A, Branche J, Allez
M, Bouhnik Y, Filippi J, Zerbib F, Savoye G, Nachury M, Moreau J,
et al: Ciclosporin versus infliximab in patients with severe
ulcerative colitis refractory to intravenous steroids: A parallel,
open-label randomised controlled trial. Lancet. 380:1909–1915.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kreijne JE, Lie MR, Vogelaar L and van der
Woude CJ: Practical guideline for fatigue management in
inflammatory bowel disease. J Crohns Colitis. 10:105–111. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pedersen J, Coskun M, Soendergaard C,
Salem M and Nielsen OH: Inflammatory pathways of importance for
management of inflammatory bowel disease. World J Gastroenterol.
20:64–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hooper KM, Barlow PG, Stevens C and
Henderson P: Inflammatory bowel disease drugs: A focus on
autophagy. J Crohns Colitis. 11:118–127. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jurenka JS: Anti-inflammatory properties
of curcumin, a major constituent of Curcuma longa: A review of
preclinical and clinical research. Altern Med Rev. 14:141–153.
2009.PubMed/NCBI
|
|
11
|
Bar-Sela G, Epelbaum R and Schaffer M:
Curcumin as an anti-cancer agent: Review of the gap between basic
and clinical applications. Curr Med Chem. 17:190–197. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aggarwal BB, Yuan W, Li S and Gupta SC:
Curcumin-free turmeric exhibits anti-inflammatory and anticancer
activities: Identification of novel components of turmeric. Mol
Nutr Food Res. 57:1529–1542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Naik SR, Thakare VN and Patil SR:
Protective effect of curcumin on experimentally induced
inflammation, hepatotoxicity and cardiotoxicity in rats: Evidence
of its antioxidant property. Exp Toxicol Pathol. 63:419–431. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Toden S, Theiss AL, Wang X and Goel A:
Essential turmeric oils enhance anti-inflammatory efficacy of
curcumin in dextran sulfate sodium-induced colitis. Sci Rep.
7:8142017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu L, Liu YL, Liu GX, Chen X, Yang K,
Yang YX, Xie Q, Gan HK, Huang XL and Gan HT: Curcumin ameliorates
dextran sulfate sodium-induced experimental colitis by blocking
STAT3 signaling pathway. Int Immunopharmacol. 17:314–320. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lang A, Salomon N, Wu JC, Kopylov U, Lahat
A, Har-Noy O, Ching JY, Cheong PK, Avidan B, Gamus D, et al:
Curcumin in combination with mesalamine induces remission in
patients with mild-to-moderate ulcerative colitis in a randomized
controlled trial. Clin Gastroenterol Hepatol. 13:1444–1449.e1.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baliga MS, Joseph N, Venkataranganna MV,
Saxena A, Ponemone V and Fayad R: Curcumin, an active component of
turmeric in the prevention and treatment of ulcerative colitis:
Preclinical and clinical observations. Food Funct. 3:1109–1117.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Baur JA and Sinclair DA: Therapeutic
potential of resveratrol: The in vivo evidence. Nat Drug Discov.
5:493–506. 2006. View
Article : Google Scholar
|
|
19
|
Tomé-Carneiro J, Larrosa M,
González-Sarrías A, Tomás-Barberán FA, García-Conesa MT and Espín
JC: Resveratrol and clinical trials: The crossroad from in vitro
studies to human evidence. Curr Pharm Des. 19:6064–6093. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Singh UP, Singh NP, Singh B, Singh B,
Hofseth LJ, Price RL, Nagarkatti M and Nagarkatti PS: Resveratrol
(trans-3,5,4′-trihydroxystilbene) induces SIRT1 and down-regulates
nuclear transcription factor-kappaB activation to abrogate dextran
sulfate sodium-induced colitis. J Pharmacol Exp Ther. 11:829–839.
2009.
|
|
21
|
Tian J, Chen JW, Gao JS, Li L and Xie X:
Resveratrol inhibits TNF-α-induced IL-1β, MMP-3 production in human
rheumatoid arthritis fibroblast-like synoviocytes via modulation of
PI3kinase/Akt pathway. Rheumatol Int. 33:1829–1835. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Carrasco C, Holguín-Arévalo MS,
Martín-Partido G, Rodríguez AB and Pariente JA: Chemopreventive
effects of resveratrol in a rat model of cerulein-induced acute
pancreatitis. Mol Cell Biochem. 387:217–225. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Komatsu M, Kurokawa H, Waguri S, Taguchi
K, Kobayashi A, Ichimura Y, Sou YS, Ueno I, Sakamoto A, Tong KI, et
al: The selective autophagy substrate p62 activates the stress
responsive transcription factor Nrf2 through inactivation of Keap1.
Nat Cell Biol. 12:213–223. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Baxt LA and Xavier RJ: Role of autophagy
in the maintenance of intestinal homeostasis. Gastroenterology.
149:553–562. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Garcia-Maurino S, Alcaide A and Dominguez
C: Pharmacological control of autophagy: Therapeutic perspectives
in inflammatory bowel disease and colorectal cancer. Curr Pharm
Des. 18:3853–3873. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cooper HS, Murthy S, Shah R and Sedergran
D: Clinicopathologic study of dextran sulfate sodium experimental
murine colitis. Lab Invest. 69:238–249. 1993.PubMed/NCBI
|
|
27
|
Vochyánová Z, Bartošová L, Bujdáková V,
Fictum P, Husník R, Suchý P, Šmejkal K and Hošek J: Diplacone and
mimulone ameliorate dextran sulfate sodium-induced colitis in rats.
Fitoterapia. 101:201–207. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chassaing B, Aitken JD, Malleshappa M and
Vijay-Kuma M: Dextran sulfate sodium (DSS)-induced colitis in mice.
Curr Protoc Immunol. 104:15.25.11–15.25.14. 2014.
|
|
29
|
Melgar S, Karlsson L, Rehnström E,
Karlsson A, Utkovic H, Jansson L and Michaëlsson E: Validation of
murine dextran sulfate sodium-induced colitis using four
therapeutic agents for human inflammatory bowel disease. Int
Immunopharmacol. 8:836–844. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Brown SR and Coviello LC: Extraintestinal
manifestations associated with inflammatory bowel disease. Surg
Clin North Am. 95:1245–1259. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhu H and Li YR: Oxidative stress and
redox signaling mechanisms of inflammatory bowel disease: Updated
experimental and clinical evidence. Exp Biol Med (Maywood).
237:474–480. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Håkansson Å, Tormo-Badia N, Baridi A, Xu
J, Molin G, Hagslätt ML, Karlsson C, Jeppsson B, Cilio CM and Ahrné
S: Immunological alteration and changes of gut microbiota after
dextran sulfate sodium (DSS) administration in mice. Clin Exp Med.
15:107–120. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nenci A, Becker C, Wullaert A, Gareus R,
van Loo G, Danese S, Huth M, Nikolaev A, Neufert C, Madison B, et
al: Epithelial NEMO links innate immunity to chronic intestinal
inflammation. Nature. 446:557–561. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Schwanke RC, Marcon R, Meotti FC, Bento
AF, Dutra RC, Pizzollatti MG and Calixto JB: Oral administration of
the flavonoid myricitrin prevents dextran sulfate sodium-induced
experimental colitis in mice through modulation of PI3K/Akt
signaling pathway. Mol Nutr Food Res. 57:1938–1949. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Abraham C and Medzhitov R: Interactions
between the host innate immune system and microbes in inflammatory
bowel disease. Gastroenterology. 140:1729–1737. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Neurath MF: Cytokines in inflammatory
bowel disease. Nat Rev Immunol. 14:329–342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Stillie R and Stadnyk AW: Role of TNF
receptors, TNFR1 and TNFR2, in dextran sodium sulfate-induced
colitis. Inflamm Bowel Dis. 15:1515–1525. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Geremia A, Biancheri P, Allan P, Corazza
GR and Di Sabatino A: Innate and adaptive immunity in inflammatory
bowel disease. Autoimm Rev. 13:3–10. 2014. View Article : Google Scholar
|
|
39
|
Strober W and Fuss IJ: Proinflammatory
cytokines in the pathogenesis of inflammatory bowel diseases.
Gastroenterology. 140:1756–1767.e1751. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Műzes G, Molnár B, Tulassay Z and Sipos F:
Changes of the cytokine profile in inflammatory bowel diseases.
World J Gastroenterol. 18:5848–5861. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Biasi F, Leonarduzzi G, Oteiza PI and Poli
G: Inflammatory bowel disease: Mechanisms, redox considerations,
and therapeutic targets. Antioxid Redox Signal. 19:1711–1747. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fiorino G, Szabo H, Fries W, Malesci A,
Peyrin-Biroulet L and Danese S: Adalimumab in Crohn's disease: Tips
and tricks after 5 years of clinical experience. Curr Med Chem.
18:1230–1238. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Rutgeerts P, Sandborn WJ, Feagan BG,
Reinisch W, Olson A, Johanns J, Travers S, Rachmilewitz D, Hanauer
SB, Lichtenstein GR, et al: Infliximab for induction and
maintenance therapy for ulcerative colitis. New Eng J Med.
353:2462–2476. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dionne S, D'Agata D, Hiscott J, Vanounou T
and Seidman E: Colonic explant production of IL-1 and its receptor
antagonist is imbalanced in inflammatory bowel disease (IBD). Clin
Exp Immunol. 112:435–442. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kwon KH, Murakami A, Hayashi R and
Ohigashi H: Interleukin-1beta targets interleukin-6 in progressing
dextran sulfate sodium-induced experimental colitis. Biochem
Biophys Res Commun. 337:647–654. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhu HT, Bian C, Yuan JC, Chu WH, Xiang X,
Chen F, Wang CS, Feng H and Lin JK: Curcumin attenuates acute
inflammatory injury by inhibiting the TLR4/MyD88/NF-κB signaling
pathway in experimental traumatic brain injury. J
Neuroinflammation. 11:592014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Epstein J, Sanderson IR and Macdonald TT:
Curcumin as a therapeutic agent: The evidence from in vitro, animal
and human studies. Br J Nutr. 103:1545–1557. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sánchez-Fidalgo S, Cárdeno A, Villegas I,
Talero E and de la Lastra CA: Dietary supplementation of
resveratrol attenuates chronic colonic inflammation in mice. Eur J
Pharmacol. 633:78–84. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Klionsky DJ, Cregg JM, Dunn WA Jr, Emr SD,
Sakai Y, Sandoval IV, Sibirny A, Subramani S, Thumm M, Veenhuis M
and Ohsumi Y: A unified nomenclature for yeast autophagy-related
genes. Dev Cell. 5:539–545. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tschurtschenthaler M, Adolph TE, Ashcroft
JW, Niederreiter L, Bharti R, Saveljeva S, Bhattacharyya J, Flak
MB, Shih DQ, Fuhler GM, et al: Defective ATG16L1-mediated removal
of IRE1α drives Crohn's disease-like ileitis. J Exp Med.
214:401–422. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Shintani T and Klionsky DJ: Autophagy in
health and disease: A double-edged sword. Science. 306:990–995.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Alayev A, Sun Y, Snyder RB, Berger SM, Yu
JJ and Holz MK: Resveratrol prevents rapamycin-induced upregulation
of autophagy and selectively induces apoptosis in TSC2-deficient
cells. Cell Cycle. 13:371–382. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Otomo C, Metlagel Z, Takaesu G and Otomo
T: Structure of the human ATG12~ATG5 conjugate required for LC3
lipidation in autophagy. Nat Struct Mol Biol. 20:59–66. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ou X, Lee MR, Huang X, Messina-Graham S
and Broxmeyer HE: SIRT1 positively regulates autophagy and
mitochondria function in embryonic stem cells under oxidative
stress. Stem Cells. 32:1183–1194. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
He C and Klionsky DJ: Regulation
mechanisms and signaling pathways of autophagy. Annu Rev Genet.
43:67–93. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Laplante M and Sabatini DM: mTOR signaling
in growth control and disease. Cell. 149:274–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Alves MM, Fuhler GM, Queiroz KC, Scholma
J, Goorden S, Anink J, Spek CA, Hoogeveen-Westerveld M, Bruno MJ,
Nellist M, et al: PAK2 is an effector of TSC1/2 signaling
independent of mTOR and a potential therapeutic target for Tuberous
Sclerosis Complex. Sci Rep. 5:145342015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Seinen ML, van Nieuw Amerongen GP, de Boer
NK and van Bodegraven AA: Rac attack: Modulation of the small
GTPase Rac in inflammatory bowel disease and thiopurine therapy.
Mol Diagn Ther. 20:551–557. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Satoh A, Brace CS, Rensing N, Cliften P,
Wozniak DF, Herzog ED, Yamada KA and Imai S: Sirt1 extends life
span and delays aging in mice through the regulation of Nk2
homeobox 1 in the DMH and LH. Cell Metab. 18:416–430. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Singh UP, Singh NP, Singh B, Hofseth LJ,
Price RL, Nagarkatti M and Nagarkatti PS: Resveratrol
(trans-3,5,4′-trihydroxystilbene) induces silent mating type
information regulation-1 and down-regulates nuclear transcription
factor-kappaB activation to abrogate dextran sulfate sodium-induced
colitis. J Pharmacol Exp Ther. 332:829–839. 2010. View Article : Google Scholar : PubMed/NCBI
|