Introduction
Bronchial asthma is a heterogeneous disease
characterized by chronic airway inflammation and often associated
with airway hyper-responsiveness (1,2).
Bronchial asthma usually results in extensive and variable
reversible airflow limitation resulting in recurrent attacks
(3,4). Severe cases can endanger life
(5,6). Common causes of asthma include air
pollution, weather changes, upper respiratory tract infection,
bronchitis and colds, allergic reactions and certain drugs, such as
aspirin and penicillin (7,8). Common substances such as dust,
pollen, carpets, clothing fibres and urine can trigger asthma.
Dermatophagoides pteronyssinus peptidase 1 (Der p1)
contained in air dust is the most frequent human allergen (9,10).
Thus, even innocuous compounds can act as allergens and have a very
serious impact on quality of life (11).
Airway epithelial cells constitute the first line of
defence against inflammatory triggers, and injury to the airway
epithelium is one of the characteristics of asthma (12,13).
Increased epithelial cell apoptosis is detectable in patients with
asthma and is associated with disease severity (14). Moreover, previous studies have
suggested that apoptosis results in loss of epithelium integrity
and exposes the airway and lungs to excess pathogens and allergens,
which in turn exacerbates inflammation and compromises homeostasis
of the airway epithelium (15,16).
Thus, apoptosis of bronchial epithelial cells contributes to asthma
(17,18). However, whether Der p1 can cause
apoptosis of bronchial epithelial cells remains unknown and the
underlying molecular mechanisms of Der p1-induced asthma are poorly
characterized.
Long non-coding RNAs (lncRNAs) serve an important
role in many bioactivities, such as the regulation of cell cycle
progression, apoptosis, cell differentiation and autophagy
(19). A previous study has
demonstrated that the levels of lncRNAs in serum vary greatly
between healthy people and patients with asthma, suggesting that
lncRNAs may participate in the occurrence and development of asthma
(20). In a recent study, 17
lncRNAs were detected in the serum of patients with asthma, and the
expression levels of several lncRNAs were significantly lower,
compared with serum from individuals without asthma (21). Other studies have also indicated
that lncRNAs TCF7 and GAS5 promote the development of asthma
(22,23). Among these lncRNAs, lncRNA
Opa-interacting protein 5 antisense RNA1 (OIP5-AS1) was
demonstrated to promote inflammation in several cell types,
including macrophages and endothelial cells (24,25).
Since inflammation is one of the pathological causes of asthma, it
may be hypothesized that OIP5-AS1 could serve a functional role in
bronchial asthma.
MicroRNAs (miRNAs) are endogenous small RNAs 20–24
nucleotides in length and have been implicated in asthma (26,27).
For instance, it has been reported that miR-19a was upregulated in
T cells and promoted the production of T helper 2 (Th2) cytokines
in the asthmatic airway (28). A
previous study also demonstrated that the levels of microRNA-18a,
−27a, −128 and −155 were decreased in asthmatic bronchial
epithelial cells (29). Moreover,
miR-143-3p was downregulated in smooth muscle cells (SMCs)
following stimulation with transforming growth factor-β1 (TGF-β1)
(30). In addition, miR-143-3p was
significantly downregulated in SMCs in patients with asthma
compared with healthy controls (31). However, the role of miR-143-3p in
asthma inflammation is still elusive and poorly understood.
The aim of the present study was to evaluate the
role of OIP5-AS1 in asthma using an in vitro model. lncRNA
OIP5-AS1 expression was evaluated in Der p1-exposed BEAS-2B cells.
Moreover, the function of OIP5-AS1 in Der p1-induced inflammation
and apoptosis was also evaluated. The findings of the present study
could provide deeper insight to the role of OIP5-AS1 in asthma
development and identify novel molecular mechanisms and treatment
targets for asthma research.
Materials and methods
Cell culture and transfection
The human bronchial epithelial cell line BEAS-2B was
obtained from American Type Culture Collection. Cells were cultured
in Dulbecco's Modified Eagle Medium (Gibco; Thermo Fisher
Scientific, Inc.) with 100 µg/ml penicillin-streptomycin
(Sigma-Aldrich; Merck KGaA) at 37°C with 5% CO2. To
establish an in vitro asthma model, BEAS-2B cells were
incubated in the presence of 10 µg/ml Der p1 (Sigma-Aldrich; Merck
KGaA) for 24 h. Untreated BEAS-2B cells were used as a control.
Cells were transfected with miR-143-3p mimics (5
nM), inhibitor (5 nM) or negative control (NC, 5 nM), as well as
OIP5-AS1 small interfering (si)-RNA (5 nM), si-NC (5 nM),
pcDNA3.1-HMGB1 (p-HMGB1; 5 nM), or pcDNA3.1 NC (p-NC; 5 nM). All
nucleic acids used for transfection were from Shanghai GeneChem
Co., Ltd. Transfections were performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) in serum-free Opti-MEM medium (Gibco; Thermo
Fisher Scientific, Inc.). The sequences of the nucleic acids used
were as follows: i) miR-143-3p mimics, 5′-UGAGAUGAAGCACUGUAGCUC−3′;
ii) mimics NC: 5′-UCACAACCUCCUAGAAAGAGUAGA-3′; iii) miR-143-3p
inhibitor, 5′-GAGCUACAGUGCUUCAUCUCA−3′; iv) inhibitor NC,
5′-UUCUCCGAACGUGUCACGUTT−3′; v) si-OIP5-AS1 sense,
5′-GCAGCAUGCUGUGUGCAAA-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT−3′; and vi) si-NC sense,
5′-UUCUCCGAACGUGUCACGUTT−3′ and antisense,
5′-CUUGAGGCUGUUGUCAUACTT−3′. The transfection efficiency was
evaluated 48 h after transfection.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from BEAS-2B cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). RNA was reverse transcribed using the Prime-Script One Step
RT-PCR kit (Takara Biotechnology Co., Ltd.) at 65°C for 5 min,
followed by 37°C for 50 min, then 70°C for 15 min. RT-qPCRs were
performed in an Applied Biosystems 7500 Real Time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.) using SYBR
GREEN Mastermix (Beijing Solarbio Science & Technology Co.,
Ltd.) according to manufacturer's instructions. The miR-143-3p,
OIP5-AS1, high mobility group box 1 (HMGB1), tumor necrosis
factor-α (TNF-α), interleukin (IL)-6, IL-8 genes were amplified,
and GAPDH and U6 served as internal controls. The primer sequences
were as follows: i) miR-143-3p-forward (F),
5′-CTTCTCAGACGTGCGCAAGG-3′; ii) miR-143-3p-reverse (R),
5′-GTTGATGTTGAAACAGCTCTC−3′; iii) OIP5-AS1-F,
5′-TGCGAAGATGGCGGAGTAAG-3′; iv) OIP5-AS1-R,
5′-TAGTTCCTCTCCTCTGGCCG-3′; v) HMGB1-F,
5′-TGATGCGAACACGGCGTGCTCTA-3′; vi) HMGB1-R,
5′-GCACAAAGAATGCATATGAGGAC-3′; vii) TNF-α-F,
5′-AAGCCTGTAGCCCACGTCGTA−3′; viii) TNF-α-R,
5′-GGCACCACTAGTTGGTTGTCTTTG−3′; ix) IL-6-F,
5′-CCGGAGAGGAGACTTCACAG-3′; x) IL-6-R, 5′-TCCACGATTTCCCAGAGAAC-3′;
xi) IL-8-F, 5′-GCTCTGTGTGAAGGTGCAGTT−3′; xii) IL-8-R,
5′-ACCCAGTTTTCCTTGGGGTC-3′; xiii) U6-F,
5′-ATTGGAACGATACAGAGAAGATT-3′; xiv) U6-R,
5′-GGAACGCTTCACGAATTTG-3′; xv) GAPDH-F, 5′-CCACAGTCCATGCCATCAC-3′;
and xvi) GAPDH-R, 5′-GCTTCACCACCTTCTTGATG-3′. Relative RNA levels
were calculated using the 2−ΔΔCq method (32).
Western blotting assay
Total protein was extracted from BEAS-2B cells using
radio-immunoprecipitation assay (RIPA) buffer (Vazyme Biotech Co.,
Ltd.), then quantified using the Protein Concentration
Determination kit (Bio-Rad Laboratories, Inc.). Proteins (20 µg)
were then resolved by SDS-PAGE on 10% gels, transferred to PVDF
membranes, and blocked with 5% non-fat milk for 2 h at room
temperature. The samples were then incubated with primary
antibodies specific for HMGB1 (1:10,000; cat. no. ab79823, Abcam)
or GAPDH (1:10,000; cat. no. ab181602; Abcam) at 4°C overnight and
subsequently probed with the HRP-conjugated secondary antibody
(goat anti-rabbit IgG H&L; 1:2,000; cat. no. ab205718; Abcam)
at 37°C for 45 min. Protein bands were scanned using an ECL reagent
(Bio-Rad laboratories, Inc.) and imaged. GAPDH served as an
internal control. Relative protein expression was quantified using
Image-Pro Plus software (version 6.0; Media Cybernetics, Inc.).
ELISA
ELISA was conducted to determine the levels of the
inflammatory factors. Briefly, cells were centrifuged at 1,500 × g
for 15 min at room temperature and the supernatant was collected.
The levels of TNF-α, IL-6 and IL-8 were evaluated using commercial
ELISA kits (cat. nos. ab181421, ab178013 and ab46032, respectively;
all from Abcam) according to the manufacturer's instructions.
Annexin V-Fluorescein isothiocyanate
(FITC) and propidium iodide (PI) staining
For cell apoptosis measurements, cells were
incubated with trypsin, then stained with Annexin V-FITC/PI double
staining kit (cat. no. 556547, BD Biosciences) according to the
manufacturer's instructions. A FACScan flow cytometer (BD
Biosciences,) was used for analysis. The data were analyzed using
CellQuest software (version 3.3, BD Biosciences).
Dual luciferase reporter assay
Binding between OIP5-AS1 and miR-143-3p was
predicted using Starbase v2.0 (http://starbase.sysu.edu.cn/) and TargetScan 7.2
(http://www.targetscan.org), and the
wild-type (WT) and mutant (MUT) fragments of OIP5-AS1 or HMGB1 were
designed accordingly.
A dual luciferase reporter assay was performed to
confirm the binding between OIP5-AS1 and miR-143-3p, as well as the
binding between miR-143-3p and HMGB1. For the construction of the
recombinant plasmid, the pre-miR-143-3p sequence was obtained from
the National Center for Biotechnology Information database
(accession no. MIMAT0000435; http://www.ncbi.nlm.nih.gov) and the fragment was
extended to 80 bp both downstream and upstream for amplification.
The sequences primer sequences were as follows: i)
Pre-miR-143-3p-F, 5′-TGAGATGAAGCACTGTAGCT-3′; ii) pre-miR-143-3p-R,
5′-TGAGATGAAGCACTGTAGCTC−3′; iii) OIP5-AS1-3′UTR-F,
5′-CCGTCTGAACTATCCTGCCC-3′; iv) OIP5-AS1-3′UTR-R,
5′-TCAACGTCAAGGAGTCGCAG-3′; v) HMGB1-3′UTR-F,
5′-CCGGATGCTTCTGTCAACTT-3′; and vi) HMGB1-3′UTR-R,
5′-GGGCGGTACTCAGAACAGAA-3′. The WT and MUT fragments of OIP5-AS1 or
HMGB1 containing the putative miR-124-3p binding sequence, were
amplified and subcloned into a pGL4.10 luciferase reporter vector
(Promega Corporation). The PCR reaction condition was as described
as above. The thermocycling conditions consisted of: i) 94°C for 2
min, 94°C for 30 sec, 60°C for 30 sec and 72°C for 30 sec; ii) 30
cycles of denaturation at 94°C for 2 min, then annealing at 60°C
for 25 sec; and iii) final extension at 72°C for 30 sec. Cells were
then co-transfected with 5 nM miR-143-3p mimics or mimics NC, as
well as luciferase reporter plasmids using
Lipofectamine® 2000. (Invitrogen; Thermo Fisher
Scientific, Inc.). After 48-h incubation, firefly and
Renilla luciferase activities were measured using a
Bright-Glo™ Luciferase Assay system (Promega Corporation). Firefly
luciferase activity was normalized to Renilla luciferase
activity.
Statistical analysis
Data are presented as the mean ± SD of three
experiments. An unpaired Student's t-test was used for comparisons
between two groups. Multi-group comparisons were performed using
one-way ANOVA followed by Tukey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference. Data
analysis was conducted using SPSS 20.0 statistical software (IBM
Corp.).
Results
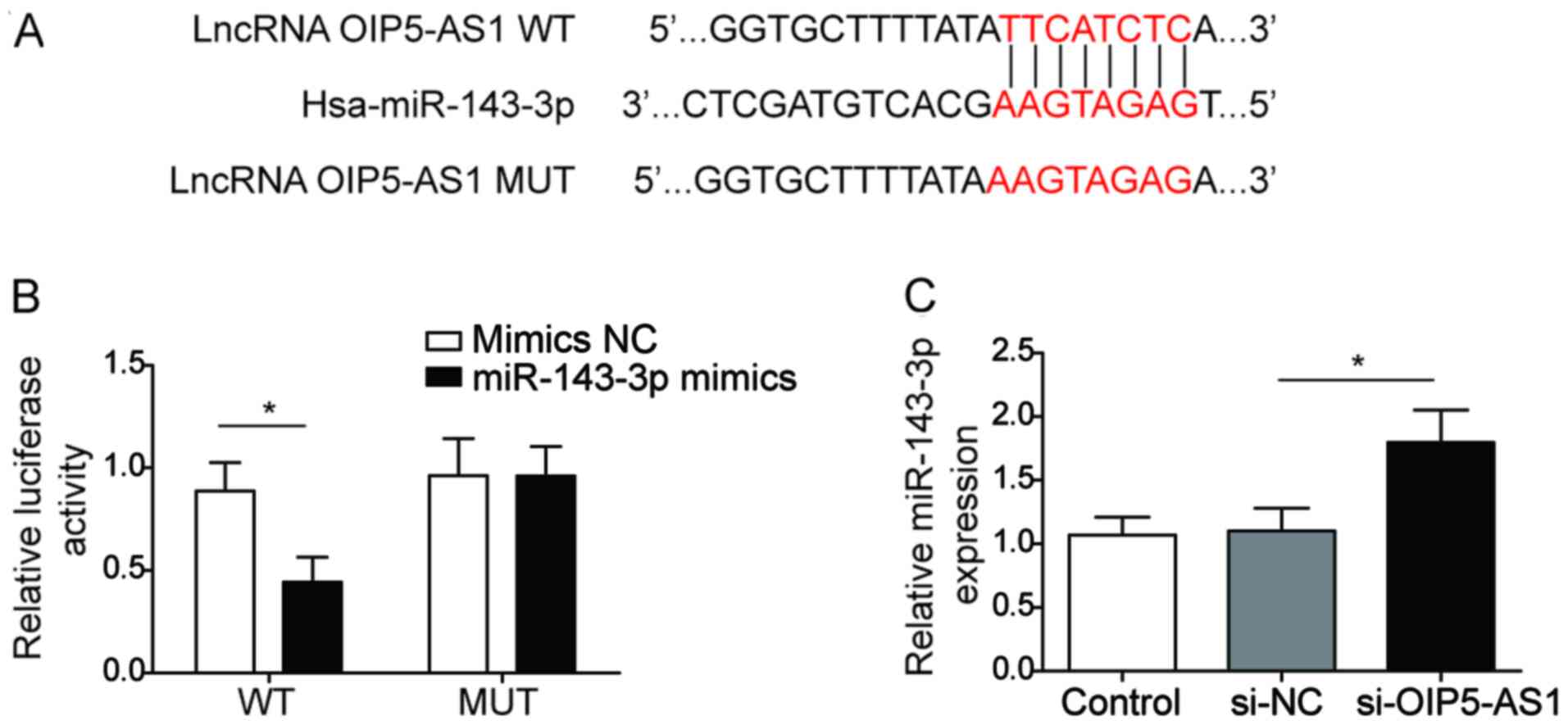
OIP5-AS1 knockdown reduces Der
p1-induced inflammatory responses and apoptosis in BEAS-2B
cells
OIP5-AS1 and miR-143-3p expression levels were
evaluated in BEAS-2B cells following Der p1 treatment. OIP5-AS1 was
significantly upregulated, while miR-143-3p was downregulated in
Der p1-treated BEAS-2B cells, compared with the control (Fig. 1A and B). To further examine the
potential function of OIP5-AS1, BEAS-2B cells were transfected with
si-OIP5-AS1. OIP5-AS1 expression was significantly reduced
si-PIP5-AS1-transfected cells, compared with si-NC, indicating
successful transfection (Fig.
1C).
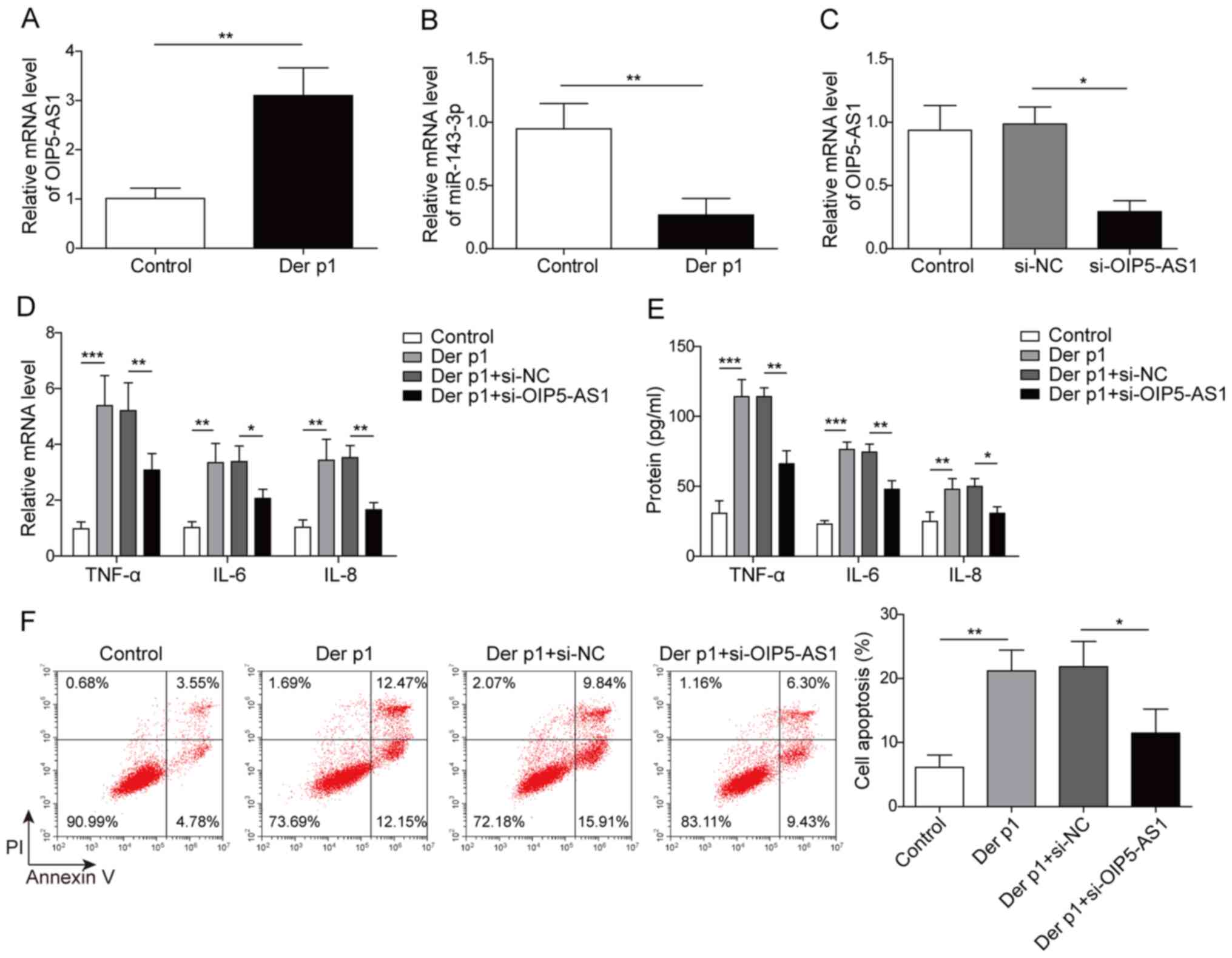 | Figure 1.OIP5-AS1 knockdown reduces Der
p1-induced inflammatory responses and apoptosis of BEAS-2B cells.
(A) OIP5-AS1 mRNA expression was measured in BEAS-2B cells treated
with 10 µg/ml Der p1 for 24 h. (B) Expression of miR-143-3p in
BEAS-2B cells treated with 10 µg/ml Der p1 for 24 h. (C) OIP5-AS1
expression in BEAS-2B cells following transfection with si-OIP5-AS1
or si-NC. IL-6, IL-8 and TNF-α (D) mRNA and (E) protein levels
following Der p1 treatment and OIP5-AS1 knockdown. (F) Apoptosis in
BEAS-2B cells Der p1-treated cells transfected with si-OIP5-AS1.
*P<0.05, **P<0.01, ***P<0.001. OIP5-AS1, Opa-interacting
protein 5 antisense RNA1; Der p1, Dermatophagoides
pteronyssinus peptidase 1; RT-qPCR, reverse
transcription-quantitative PCR; miR, microRNA; si, small
interfering; NC, negative control; IL, interleukin; TNF-α, tumor
necrosis factor-α; PI, propidium iodide. |
TNF-α, IL-6 and IL-8 levels were significantly
upregulated in cells treated with Der p1, compared with untreated
cells; however, OIP5-AS1 knockdown significantly reduced the
expression of these inflammatory factors, relative to si-NC
(Fig. 1D and E). Moreover,
treatment with Der p1 significantly enhanced the apoptotic rate of
BEAS-2B cells, compared with control. Transfection with si-OIP5-AS1
significantly reduced the frequency of apoptotic cells, compared
with si-NC (Fig. 1F). Altogether,
these results indicated that OIP5-AS1 silencing may attenuate Der
p1-induced inflammatory responses and apoptosis in
vitro.
LncRNA OIP5-AS1 directly targets and
negatively regulates miR-143-3p in BEAS-2B cells
To further confirm the association between OIP5-AS1
and miR-143-3p in BEAS-2B cells, potential binding partners for
OIP5-AS1 were predicted using the Starbase software, which
identified miR-143-3p as a target. Furthermore, luciferase activity
was significantly reduced in BEAS-2B cells co-transfected with
miR-143-3p and WT-OIP5-AS1 (Fig.
2A), but not with MUT-OIP5-AS1 (Fig. 2B), indicating that OIP5-AS1
directly targeted miR-143-3p. In addition, miR-143-3p was
significantly upregulated following transfection with si-OIP5-AS1,
compared with si-NC, which suggested that OIP5-AS1 may negatively
regulate miR-143-3p expression (Fig.
2C). Collectively, these findings suggested that lncRNA
OIP5-AS1 may target miR-143-3p and regulate its expression in
BEAS-2B cells.
Suppression of OIP5-AS1 exacerbates
Derp1-induced inflammation and apoptosis via miR-143-3p
The effect of miR-143-3p expression on Der
p1-treated BEAS-2B cells was examined using miR-143-3p inhibitor
transfection (Fig. 3A). Following
transfection with miR-143-3p inhibitor, TNF-α, IL-6 and IL-8 were
significantly upregulated at the mRNA and the protein levels,
compared with inhibitor NC (Fig. 3B
and C). However, in BEAS-2B cells co-transfected with the
inhibitor and OIP5-AS1, the levels of these cytokines significantly
decreased to levels comparable to inhibitor NC. Moreover,
miR-143-3p inhibitor transfection also promoted cell apoptosis.
This effect was also reversed following co-transfection with
si-OIP5-AS1, reducing the frequency of apoptotic cells to the same
levels as inhibitor NC (Fig. 3D).
These results indicated that miR-143-3p attenuated Der p1-induced
inflammatory responses and apoptosis in BEAS-2B cells and that
OIP5-AS1 exacerbated these effects via miR-143-3p.
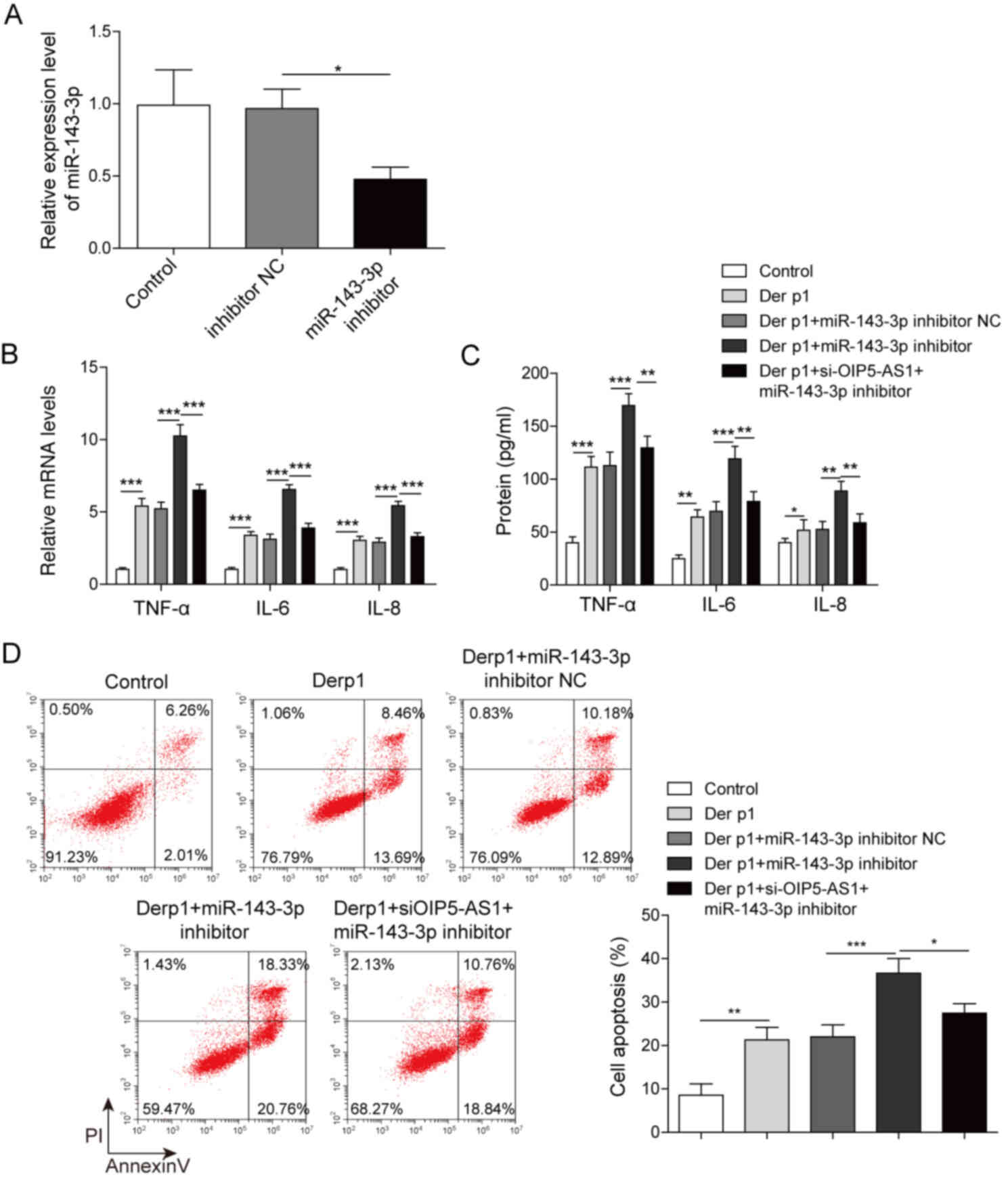 | Figure 3.OIP5-AS1 knockdown exacerbates
Derp1-induced inflammation and cell apoptosis through miR-143-3p.
(A) miR-143-p expression in BEAS-2B cells transfected with
miR-143-3p inhibitor or inhibitor NC. IL-6, IL-8 and TNF-α (B) mRNA
expression and (C) protein levels in Der p1-treated cells
co-transfected with miR-143-3p inhibitor and si-OIP5-AS1. (D)
Apoptosis in Der p1-treated cells co-transfected with miR-143-3p
inhibitor and si-OIP5-AS1. *P<0.05, **P<0.01, ***P<0.001.
OIP5-AS1, Opa-interacting protein 5 antisense RNA1; Der p1,
Dermatophagoides pteronyssinus peptidase 1; miR, microRNA;
si, small interfering; NC, negative control; IL, interleukin;
TNF-α, tumor necrosis factor-α; PI, propidium iodide. |
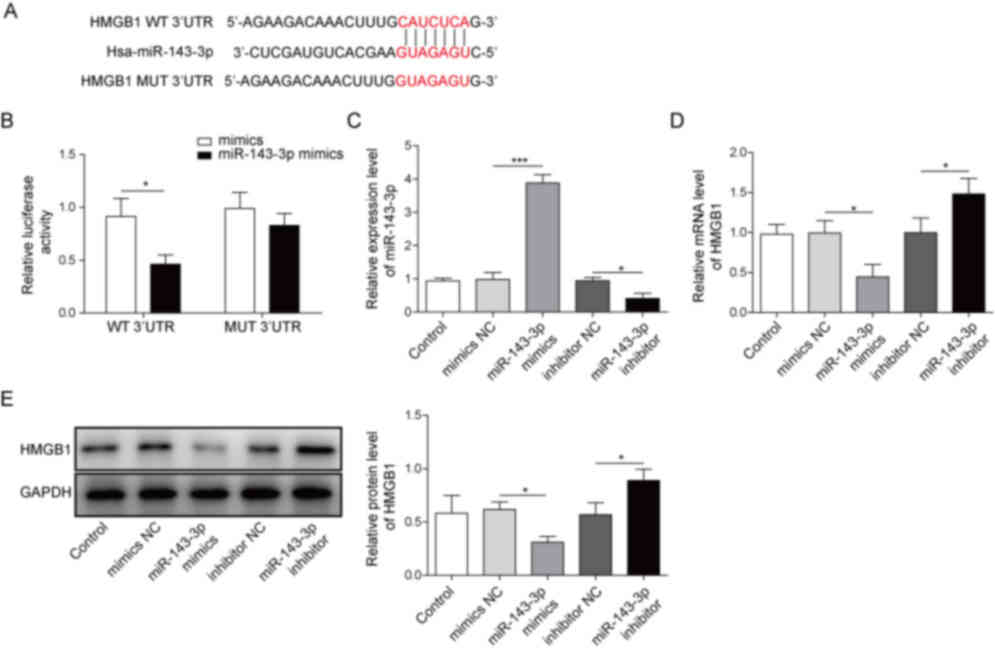
miR-143-3p directly targets and
negatively regulates HMGB1
To understand the mechanism by which miR-143-3p
regulated the Der p1-induced inflammatory responses and apoptosis
in BEAS-2B cells, potential binding targets for miR-143-3p were
screened using TargetScan 7.2, which identified HMGB1 as a
potential target gene (Fig. 4A).
Additionally, luciferase activity was significantly decreased in
BEAS-2B cells co-transfected with miR143-3p and WT-HMGB1, but not
MUT-HMGB1 (Fig. 4B). Transfections
with miR-143-3p mimics and miR-143-3p inhibitor were also carried
out in order to assess the effects of miR-143-3p expression on
BEAS-2B cells (Fig. 4C).
Transfection with miR-143-3p mimics significantly knocked down
HMGB1 expression, compared with mimics NC. Conversely, BEAS-2B
cells transfected with miR-143-3p inhibitor displayed significantly
increased levels of HMGB1, compared with the inhibitor NC (Fig. 4D and E). Altogether, these
observations suggested that miR-143-3p directly targeted HMGB1 and
negatively regulated its expression in vitro.
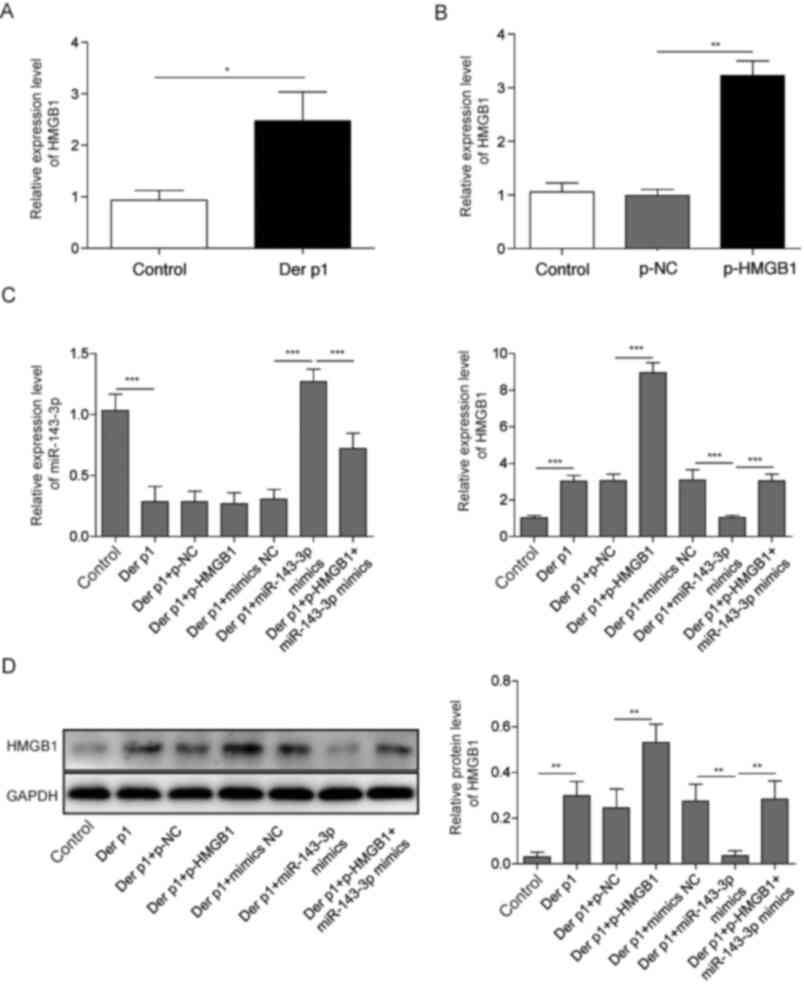
Overexpression of miR-143-3p reduces
Der p1-induced inflammation and apoptosis of BEAS-2B cells via
HMGB1
The role of HMGB1 in Der p1-induced inflammatory
response and apoptosis of BEAS-2B cells was further examined. HMGB1
was significantly upregulated in cells treated with Der p1
(Fig. 5A). BEAS-2B cells were then
transfected with miR-143-3p mimics, p-HMGB1 or both. Transfection
with the p-HMGB1 vector significantly increased the expression of
HMGB1 (Fig. 5B). miR-143-3p mimics
transfection increased the levels of miR-143-3p in BEAS-2B cells.
However, co-transfection with p-HMGB1 reduced miR-143-3p
expression, compared with cells transfected with mimics alone
(Fig. 5C). Moreover, BEAS-2B cells
transfected with miR-143-3p mimics significantly decreased HMGB1
expression, both at the mRNA and protein levels, compared with
controls. However, co-transfection with miR-143-3p mimics and
p-HMGB1 increased HMGB1 levels, compared with mimics alone
(Fig. 5D).
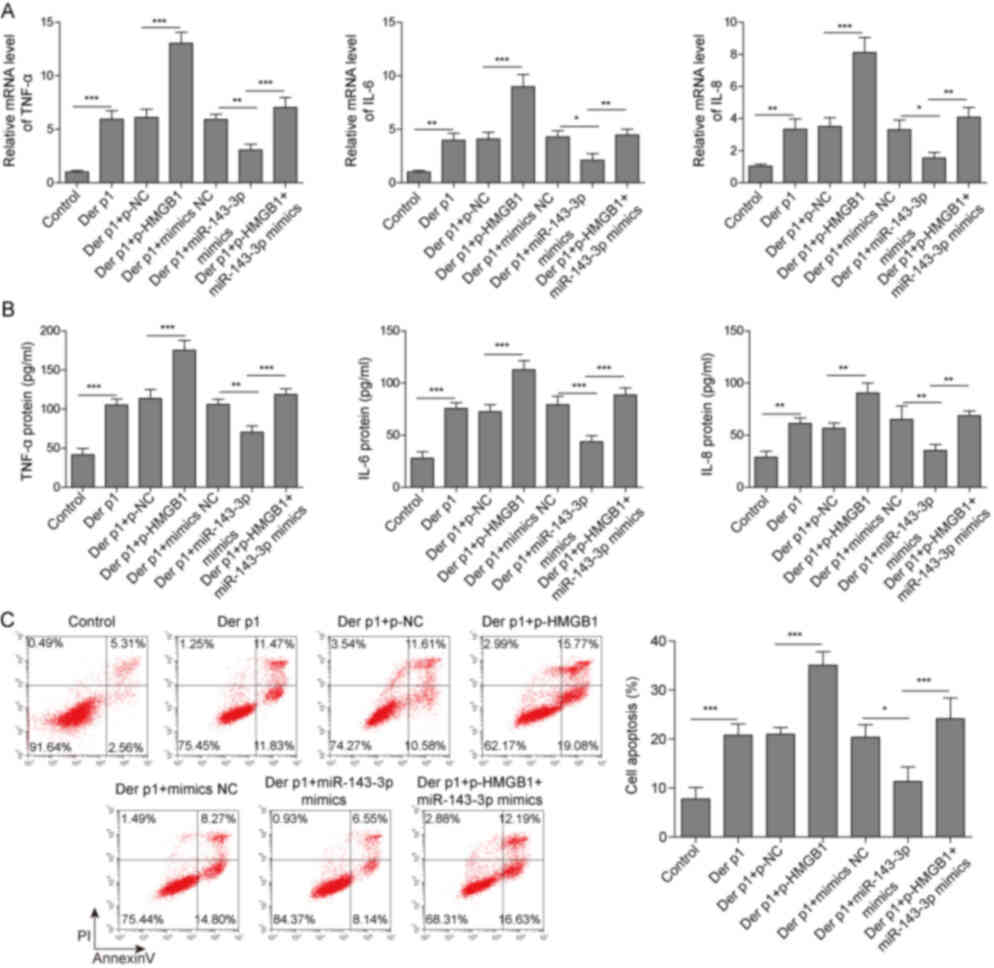
Furthermore, miR-143-3p mimics transfection
decreased TNF-α, IL-6 and IL-8 secretion, compared with mimics NC.
By contrast, HMGB1 overexpression significantly increased the
levels of these cytokines, compared with mimics alone (Fig. 6A and B). Apoptosis was also reduced
following miR-143-3p overexpression, but this effect was reversed
by transfection with p-HMGB1 (Fig.
6C). Thus, miR-143-3p reduced Der p1-induced inflammation and
apoptosis via HMGB1.
OIP5-AS1 regulates the expression of
HMGB1 via miR-143-3p
Further validation experiments were conducted using
si-OIP5-AS1, miR-143-3p inhibitor and p-HMGB1 co-transfection.
OIP5-AS1 silencing resulted in a significant increase in miR-143-3p
and a concomitant decrease in HMGB1 expression. However,
co-transfection of si-OIP5-AS1 together with miR-143-3p inhibitor
p-HMGB1 reversed these effects. Moreover, transfection miR-143-3p
inhibitor alone or overexpression of HMGB1 both resulted in an
increase in HMGB1. However, co-transfection with si-OIP5-AS1
downregulated HMGB1, compared with transfection with miR-143-3p
inhibitor or pHMGB1 alone. Altogether, these findings suggested
that OIP5-AS1 may regulate HMGB1 via miR-143-3p in Der p1-treated
BEAS-2B cells (Fig. 7).
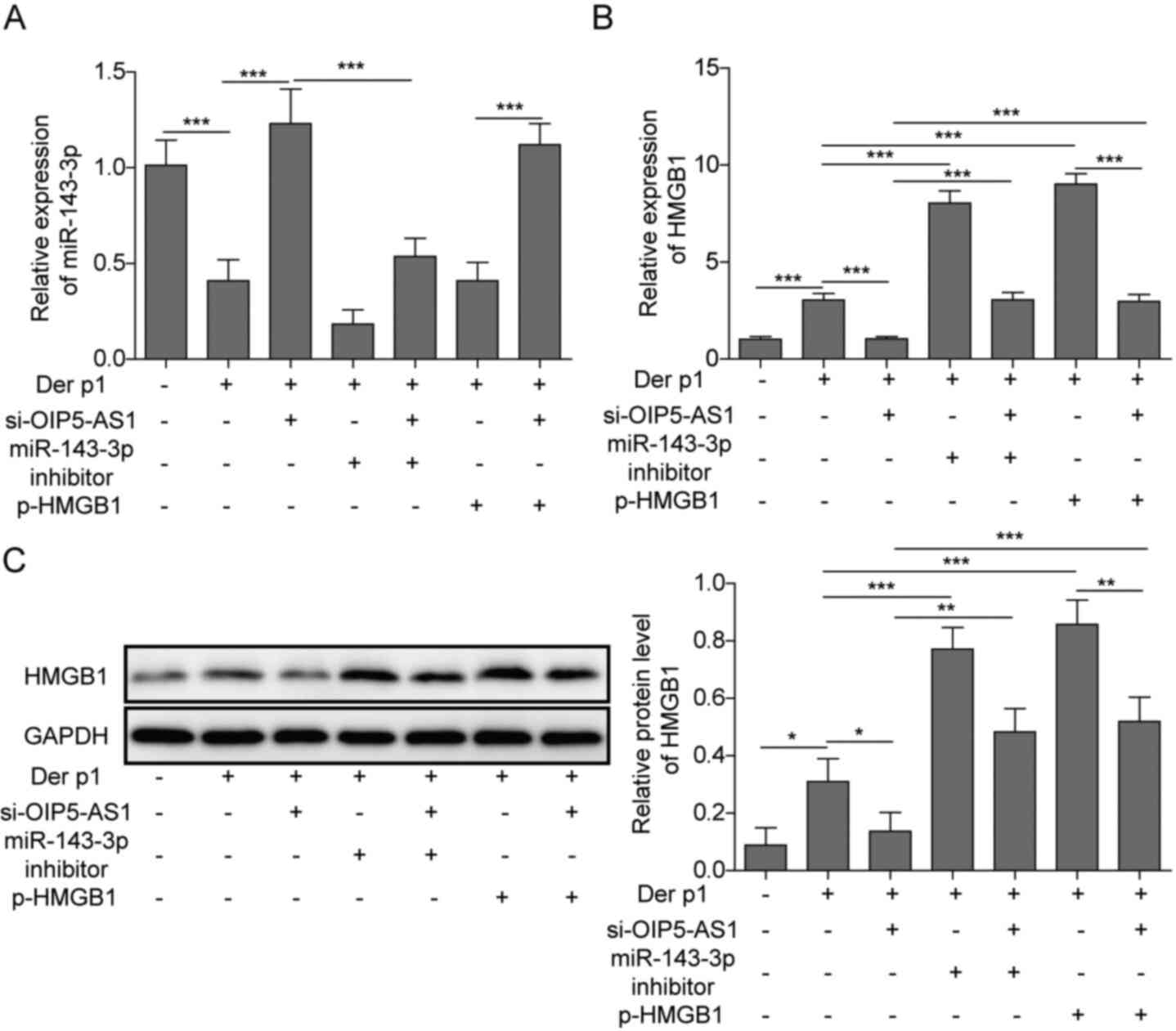 | Figure 7.OIP5-AS1 regulates the expression of
HMGB1 through miR-143-3p. (A) miR-143-3p and (B) HMGB1 expression
in cells co-transfected with si-OIP5-AS1, miR-143 inhibitor or
p-HMGB1. (C) Protein levels of HMGB1 in cells co-transfected with
si-OIP5-AS1, miR-143 inhibitor or p-HMGB1. *P<0.05, **P<0.01,
***P<0.001. OIP5-AS1, Opa-interacting protein 5 antisense RNA1;
Der p1, Dermatophagoides pteronyssinus peptidase 1; RT-qPCR,
reverse transcription-quantitative PCR; miR, microRNA; si, small
interfering; NC, negative control. |
Discussion
Bronchial asthma is a chronic airway inflammatory
disease characterized by allergic inflammation and airway
hyper-responsiveness. Bronchial asthma can cause irreversible
airway stenosis and remodelling, with serious adverse consequences
on quality of life and mortality (7). During an asthma attack, inflammatory
responses are initiated, releasing pro-inflammatory cytokines, such
as IL-1β, IL-6, IL-8 and TNF-α (33,34).
In addition, apoptosis is also involved in the development of
asthma, and the inhibition of cell apoptosis may improve asthma
(35,36). To the best of the authors'
knowledge, few studies have reported the role of OIP5-AS1 in
asthma. The present study demonstrated that OIP5-AS1 was
upregulated in Der p1-treated BEAS-2B cells. OIP5-AS1 promoted Der
p1-induced inflammation and apoptosis via miR-143-3p and HMGB1.
Furthermore, OIP5-AS silencing resulted in a reduction in the
expression levels of pro-inflammatory cytokines.
OIP5-AS1 can regulate several physiological
functions, and several studies have documented its role in cancer
development. For instance, Tao et al (37) demonstrated that inhibition of
OIP5-AS1 inhibited the development of multiple myeloma. OIP5-AS1
also promoted cell apoptosis of myeloma cells (38). However, whether OIP5-AS1 also
serves a role in asthma remains unknown. Since both activation of
inflammation and induction of cell apoptosis, such as apoptosis of
airway epithelial cells, are associated with incidence of asthma,
we noticed the promotion effects of OIP5-AS1 on cell apoptosis and
inflammation and speculate it may also participate in asthma.
Previous studies have demonstrated the effects of OIP5-AS1 on cell
apoptosis. For instance, OIP5-AS1 promoted the apoptosis of
oxidized low density lipoprotein (ox-LDL)-mediated vascular
endothelial cells by targeting glycogen synthase kinase-3β, and
silencing OIP5-AS1 reduced the apoptosis rate (25). Moreover, OIP5-AS1 silencing also
inhibited cell apoptosis of myeloma cells (38). In a recent study, OIP5-AS1 could
promote apoptosis of bladder cancer cells (39). Besides, OIP5-AS1 could also
facilitate ox-LDL-induced inflammation and oxidative stress in
macrophages, as well as induce ox-LDL mediated vascular endothelial
cells apoptosis (24,25). Consistent with these previous
findings, the present study demonstrated that OIP5-AS1 knockdown
inhibited Der p1-induced apoptosis.
LncRNAs regulate gene expression by serving as miRNA
sponges by competitively binding to miRNA and acting as a competing
endogenous RNA (ceRNA) of miRNA (40). Tao et al (37) suggested that OIP5-AS1 served as an
endogenous sponge of miR-367-3p in gastric cancer cells. Moreover,
OIP5-AS1 functioned as a sponge for miR-129-5p (38). In the present study, an association
between miR-143-3p and OIP5-AS1 was predicted using bioinformatics
analysis then validated in a dual-luciferase reporter assay. These
results indicated that OIP5-AS1 acted as a sponge for miR-143-3p
and negatively regulated its expression in vitro.
miR-143-3p is a short non-coding RNA that regulates
the survival pathways of many cancer cell types (41–43).
The role of miR-143-3p in asthma has also been reported in several
studies. Indeed, the expression of miR-143-3p was lower in
asthmatic patients, compared with healthy controls (31). Cheng et al (30) also demonstrated that overexpression
of miR-143-3p inhibited the abnormal proliferation of SMCs induced
by TGF-β1, suggesting a potential role for miR-143-3p in asthma.
Consistent with these previous findings, miR-143-3p was
downregulated in Der p1-induced BEAS-2B cells in the present
study.
Mu et al (44) suggested that miR-143-3p inhibited
hyperplastic scar formation by targeting connective tissue growth
factor. However, it was also reported that miR-143-3p promoted
inflammation in cardiac hypertrophy, suggesting that the roles of
miR-143-3p might vary under different pathophysiological conditions
(45). The present study
demonstrated that miR-143-3p inhibited the expression of
pro-inflammatory factors in Der p1-induced cells. Moreover,
miR-143-3p was a direct target of OIP5-AS1, and its effects on Der
p1-induced cells were mediated through HMGB1.
HMGB1 is associated with inflammation (46). HMGB1 activates NLR family pyrin
domain containing 3 and initiates inflammatory responses (47–49).
Several studies have demonstrated that HMGB1 is associated with the
development of asthma (50–52).
Zhang et al (50)
demonstrated that recombinant HMGB1 inhibited Th17 responses in
mice with neutrophilic asthma. Yanhua et al (51) also suggested that HMGB1 may induce
asthmatic airway inflammation through 75-kDa glucose-regulated
protein. HMGB1 also facilitates allergen-induced airway remodelling
in chronic asthma and promotes lung fibrosis (52). In agreement with these previous
findings, the presented study suggested that HMGB1 overexpression
promoted inflammation and apoptosis in Der p1-treated cells.
Besides, HMGB1 was reported to be a target of miR-143-3p in bladder
cancer cells (47). In the present
study, miR-143-3p directly targeted and negatively regulated the
expression of HMGB1 in BEAS-2B cells. miR-143-3p attenuated
apoptosis and inflammation in vitro by targeting HMGB1.
In conclusion, the present study demonstrated the
effect of OIP5-AS1 on Der p1-induced inflammation and apoptosis
in vitro and suggested a potential mechanism implicating
OIP5-AS1, miR-143-3p and HMGB1 in bronchial asthma. Clinical
evidence supporting the use of non-coding RNA as therapeutic
targets may provide insight into novel treatment options for
bronchial asthma.
Acknowledgements
Not applicable.
Funding
The present study was supported by The Key Research
and Development Program of Hainan Province (grant no. ZDYF2018150)
and The National Natural Science Foundation of China (grant no.
81860007).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XJC conceived the study and methodology, acquired
funding, wrote the original draft and validated the study. LHH
conducted data curation, and contributed to data collection and
analysis. YKZ performed data analysis and contributed to data
collection. YJH supervised the study, reviewed and revised the
manuscript, and contributed to data validation. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HDM
|
house dust mite
|
|
lncRNA
|
long non-coding RNA
|
|
Der p1
|
Dermatophagoides pteronyssinus
peptidase 1
|
|
miRNA/miR
|
microRNA
|
|
FITC
|
fluorescein isothiocyanate
|
|
HMGB1
|
high mobility group box 1
|
References
|
1
|
Meldrum K, Robertson SB, Römer I, Marczylo
T, Dean LSN, Rogers A, Gant TW, Smith R, Tetley TD and Leonard MO:
Cerium dioxide nanoparticles exacerbate house dust mite induced
type II airway inflammation. Part Fibre Toxicol. 15:242018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Izuhara K, Matsumoto H, Ohta S, Ono J,
Arima K and Ogawa M: Recent developments regarding periostin in
bronchial asthma. Allergol Int. 64 (Suppl):S3–S10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alam S, Li Z and Mahadeva R: S86 Formation
of Oxidised Alpha-1 antitrypsin induces inflammatory response in
human bronchial epithelial cells. Thorax. 67:A41–A42. 2012.
View Article : Google Scholar
|
|
4
|
Ren YF, Li H, Xing XH, Guan HS, Zhang BA,
Chen CL and Zhang JH: Preliminary study on pathogenesis of
bronchial asthma in children. Pediatr Res. 77:506–510. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hellermann GR, Nagy SB, Kong X, Lockey RF
and Mohapatra SS: Mechanism of cigarette smoke condensate-induced
acute inflammatory response in human bronchial epithelial cells.
Respir Res. 3:222002. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tauber E, Herouy Y, Urbanek R, Urbanek R,
Goetz M and Hagel E: Assessment of serum myeloperoxidase in
children with bronchial asthma. Allergy. 54:177–182. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fattouh R, Algarawi A, Fattouh M, Arias K,
Walker TD, Goncharova S, Coyle AJ, Humbles AA and Jordana M:
Eosinophils are dispensable for allergic remodeling and immunity in
a model of house dust mite–induced airway disease. Am J Respir Crit
Care Med. 183:179–188. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Larsen BB, Nielsen LP, Engelstätter R,
Steinijans V and Dahl R: Effect of ciclesonide on allergen
challenge in subjects with bronchial asthma. Allergy. 58:207–212.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cozens AL, Yezzi MJ, Kunzelmann K, Ohrui
T, Chin L, Eng K, Finkbeiner WE, Widdicombe JH and Gruenert DC:
CFTR expression and chloride secretion in polarized immortal human
bronchial epithelial cells. Am J Respir Cell Mol Biol. 10:38–47.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hamasaki Y, Kohno Y, Ebisawa M, Kondo N,
Nishima S, Nishimuta T, Morikawa A, Aihara Y, Akasawa A, Adachi Y,
et al: Japanese pediatric guideline for the treatment and
management of bronchial asthma 2012. Pediatr Int. 56:441–450. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Viau E, Levischaffer F and Peccia J:
Respiratory toxicity and inflammatory response in human bronchial
epithelial cells exposed to biosolids, animal manure, and
agricultural soil particulate matter. Environ Sci Technol.
44:3142–3148. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mertens TCJ, Karmouty-Quintana H, Taube C
and Hiemstra PS: Use of airway epithelial cell culture to unravel
the pathogenesis and study treatment in obstructive airway
diseases. Pulm Pharmacol Ther. 45:101–113. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang H, Sun Y, Rong W, Fan L, Cai Y, Qu
Q, Gao Y and Zhao H: miR-221 participates in the airway epithelial
cells injury in asthma via targeting SIRT1. Exp Lung Res.
44:272–279. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou C, Yin G, Liu J, Liu X and Zhao S:
Epithelial apoptosis and loss in airways of children with asthma. J
Asthma. 48:358–365. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang LI, Na CL, Luo S, Wu D, Hogan S,
Huang T and Weaver TE: The phosphatidylcholine transfer protein
stard7 is required for mitochondrial and epithelial cell
homeostasis. Sci Rep. 7:464162017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pu Y, Liu YQ, Zhou Y, Qi YF, Liao SP, Miao
SK, Zhou LM and Wan LH: Dual role of RACK1 in airway epithelial
mesenchymal transition and apoptosis. J Cell Mol Med. 24:3656–3668.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee HS, Park DE, Song WJ, Park HW and Sohn
SW: Effect of 1.8-cineole in Dermatophagoides
pteronyssinus-stimulated bronchial epithelial cells and mouse
model of asthma. Biol Pharm Bull. 39:946–952. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bucchieri F, Gammazza AM, Pitruzzella A,
Fucarino A, Farina F, Howarth P, Holgate ST, Zummo G and Davies DE:
Cigarette smoke causes caspase-independent apoptosis of bronchial
epithelial cells from asthmatic donors. PLoS One. 10:e01205102015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim J, Abdelmohsen K, Yang X, De S,
Grammatikakis I, Noh JH and Gorospe M: LncRNA OIP5-AS1/cyrano
sponges RNA-binding protein HuR. Nucleic Acids Res. 44:2378–2392.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Austin PJ, Tsitsiou E, Boardman C, Jones
SW, Lindsay MA, Adcock IM, Chung KF and Perry MM: Transcriptional
profiling identifies the long noncoding RNA plasmacytoma variant
translocation (PVT1) as a novel regulator of the asthmatic
phenotype in human airway smooth muscle. J Allergy Clin Immunol.
139:780–789. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Naemura M, Kuroki M, Tsunoda T, Arikawa N,
Sawata Y, Shirasawa S and Kotake Y: The long noncoding RNA OIP5-AS1
is involved in the regulation of cell proliferation. Anticancer
Res. 38:77–81. 2018.PubMed/NCBI
|
|
22
|
Fan M, Xu J, Xiao Q, Chen F and Han X:
Long non-coding RNA TCF7 contributes to the growth and migration of
airway smooth muscle cells in asthma through targeting TIMMDC1/Akt
axis. Biochem Biophys Res Commun. 508:749–755. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang XY, Tang XY, Li N, Zhao LM, Guo YL,
Li XS, Tian CJ, Cheng DJ, Chen ZC and Zhang LX: GAS5 promotes
airway smooth muscle cell proliferation in asthma via controlling
miR-10a/BDNF signaling pathway. Life Sci. 212:93–101. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li X, Cao Q and Wang Y and Wang Y: LncRNA
OIP5-AS1 contributes to ox-LDL-induced inflammation and oxidative
stress through regulating the miR-128-3p/CDKN2A axis in
macrophages. RSC Advances. 9:41709–41719. 2019. View Article : Google Scholar
|
|
25
|
Wang M, Liu Y, Li C, Zhang Y, Zhou X and
Lu C: Long noncoding RNA OIP5-AS1 accelerates the ox-LDL mediated
vascular endothelial cells apoptosis through targeting GSK-3β via
recruiting EZH2. Am J Transl Res. 11:1827–1834. 2019.PubMed/NCBI
|
|
26
|
Williams AE, Larnersvensson H, Perry MM,
Campbell GA, Herrick SE, Adcock IM, Erjefalt JS, Chung KF and
Lindsay MA: MicroRNA expression profiling in mild asthmatic human
airways and effect of corticosteroid therapy. PLoS One.
4:e58892009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Feng MJ, Shi F, Qiu C and Peng WK:
MicroRNA-181a, −146a and −146b in spleen CD4+ T lymphocytes play
proinflammatory roles in a murine model of asthma. Int
Immunopharmacol. 13:347–353. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Simpson LJ, Sana P, Bhakta NR, Choy DF,
Brightbill HD, Ren X, Wang Y, Pua HH, Baumjohann D, Montoya MM, et
al: A microRNA upregulated in asthma airway T cells promotes TH2
cytokine production. Nat Immunol. 15:1162–1170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Martinez-Nunez RT, Bondanese VP, Louafi F,
Francisco-Garcia AS, Rupani H, Bedke N, Holgate S, Howarth PH,
Davies DE and Sanchez-Elsner T: A microRNA network dysregulated in
asthma controls IL-6 production in bronchial epithelial cells. PLoS
One. 9:e1116592014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng W, Yan K, Xie LY, Chen F, Yu HC,
Huang YX and Dang CX: MiR-143-3p controls TGF-β1-induced cell
proliferation and extracellular matrix production in airway smooth
muscle via negative regulation of the nuclear factor of activated T
cells 1. Mol Immunol. 78:133–139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sorianoarroquia A, Mccormick R, Molloy AP,
Mcardle A and Goljanekwhysall K: Age-related changes in miR-143-3p:
Igfbp5 interactions affect muscle regeneration. Aging Cell.
15:361–369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Persson IM, Menzel M, Ramu S, Cerps S,
Akbarshahi H and Uller L: IL-1β mediates lung neutrophilia and
IL-33 expression in a mouse model of viral-induced asthma
exacerbation. Respir Res. 19:162018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jiang XG, Yang XD, Lv Z and Zhuang PH:
Elevated serum levels of TNF-α, IL-8, and ECP can be involved in
the development and progression of bronchial asthma. J Asthma.
55:111–118. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Reece SW, Kilburg-Basnyat B, Madenspacher
JH, Luo B, Capen A, Fessler MB and Gowdy KM: Scavenger receptor
class B type I (SR-BI) modulates glucocorticoid mediated lymphocyte
apoptosis in asthma. Am Assoc Immnol. 198((1 Suppl)): S532017.
|
|
36
|
Lv J, Su W, Yu Q, Zhang M, Di C, Lin X, Wu
M and Xia Z: Heme oxygenase-1 protects airway epithelium against
apoptosis by targeting the proinflammatory NLRP3-RXR axis in
asthma. J Biol Chem. 293:18454–18465. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tao Y, Wan X, Fan Q, Wang Y, Sun H, Ma L
and Wu Y: Long non-coding RNA OIP5-AS1 promotes the growth of
gastric cancer through the miR-367-3p/HMGA2 axis. Dig Liver Dis.
52:773–779. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang N, Chen J, Zhang H, Wang X, Yao H,
Peng Y and Zhang W: LncRNA OIP5-AS1 loss-induced microRNA-410
accumulation regulates cell proliferation and apoptosis by
targeting KLF10 via activating PTEN/PI3K/AKT pathway in multiple
myeloma. Cell Death Dis. 8:e29752017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang Y, Shi F, Xia Y and Zhao H: LncRNA
OIP5-AS1 predicts poor prognosis and regulates cell proliferation
and apoptosis in bladder cancer. J Cell Biochem. 120:7499–7505.
2019. View Article : Google Scholar
|
|
40
|
Huang YA, Chan KCC and You ZH:
Constructing prediction models from expression profiles for large
scale lncRNA-miRNA interaction profiling. Bioinformatics.
34:812–819. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sun X, Dai G, Yu L, Hu Q, Chen J and Guo
W: miR-143-3p inhibits the proliferation, migration and invasion in
osteosarcoma by targeting FOSL2. Sci Rep. 8:6062018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen L, Yao H, Wang K and Liu X: Long
non-coding RNA MALAT1 regulates ZEB1 expression by sponging
miR-143-3p and promotes hepatocellular carcinoma progression. J
Cell Biochem. 118:4836–4843. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Du J, Zhang Y, Shen L, Luo J, Lei H, Zhang
P, Pu Q, Liu Y, Shuai S, Li Q, et al: Effect of miR-143-3p on C2C12
myoblast differentiation. Biosci Biotechnol Biochem. 80:706–711.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mu S, Kang B, Zeng W, Sun Y and Yang F:
MicroRNA-143-3p inhibits hyperplastic scar formation by targeting
connective tissue growth factor CTGF/CCN2 via the Akt/mTOR pathway.
Mol Cell Biochem. 416:99–108. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yu B, Zhao Y, Zhang H, Xie D, Nie W and
Shi K: Inhibition of microRNA-143-3p attenuates myocardial
hypertrophy by inhibiting inflammatory response. Cell Biol Int.
42:1584–1593. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yang H and Tracey KJ: Targeting HMGB1 in
inflammation. Biochim Biophys Acta. 1799:149–156. 2020. View Article : Google Scholar
|
|
47
|
Luo J, Chen J, Li H, Yang Y, Yun H, Yang S
and Mao X: LncRNA UCA1 promotes the invasion and EMT of bladder
cancer cells by regulating the miR-143/HMGB1 pathway. Oncol Lett.
14:5556–5562. 2017.PubMed/NCBI
|
|
48
|
Thankam FG, Roesch ZK, Dilisio MF, Radwan
MM, Kovilam A, Gross RM and Agrawal DK: Association of inflammatory
responses and ECM disorganization with HMGB1 upregulation and NLRP3
inflammasome activation in the injured rotator cuff tendon. Sci
Rep. 8:89182018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Falcão AS, Carvalho LA, Lidónio G, Vaz
AR, Lucas SD, Moreira R and Brites D: Dipeptidyl vinyl sulfone as a
novel chemical tool to inhibit HMGB1/NLRP3-inflammasome and
inflamma-miRs in Aβ-mediated microglial inflammation. ACS Chem
Neurosci. 8:89–99. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang F, Huang G, Hu B, Qian GS and Song
Y: Recombinant HMGB1 A box protein inhibits Th17 responses in mice
with neutrophilic asthma by suppressing dendritic cell-mediated
Th17 polarization. Int Immunopharmacol. 24:110–118. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yanhua L, Yanli L, Dandan Z, Anbing Z,
Weihong G and Shunfang Z: HMGB1-induced asthmatic airway
inflammation through GRP75-mediated enhancement of ER-Mitochondrial
Ca2+ transfer and ROS increased. J Cell Biochem.
119:4205–4215. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hou C, Kong J, Liang Y, Huang H, Wen H,
Zheng X, Wu L and Chen Y: HMGB1 contributes to allergen-induced
airway remodeling in a murine model of chronic asthma by modulating
airway inflammation and activating lung fibroblasts. Cell Mol
Immunol. 12:409–423. 2015. View Article : Google Scholar : PubMed/NCBI
|