Introduction
The chronic progression of ankylosing spondylitis
(AS) is characterized by inflammatory bone erosion, intravertebral
bone loss and abnormal bone overgrowth (1,2).
Sacroiliac joints, the vertebral column and ligaments affected by
inflammation have been discovered to gradually cause extensive bone
generation, leading to stiffness and fusion of the spine and
syndesmophyte formation (3).
Although previous studies have examined genetic and environmental
factors, sex, age and ethnicity as causative factors, to the best
of our knowledge, the etiology of AS remains unclear (3–5).
Genetic factors underlying AS pathogenesis have been widely studied
(6–8), and several genes that have been
identified as susceptibility factors of AS may support the
diagnosis of AS, such as HLA-B27, HLA-B51 and ERAP1 (5,9,10).
In addition, genetic associations with AS may also provide insight
into the etiopathogenesis of this disease (11). Although the molecular functions of
several genes have been studied in the context of AS, their
potential pathogenic role and their possible therapeutic use have
not been evaluated (12).
Ankylosis progressive homolog (ANKH), an amino acid
of 492 base pairs in length, is a multichannel transmembrane
protein that transports intracellular pyrophosphate (PPi) to the
extracellular milieu (13).
Extracellular PPi was discovered to be a regulator of pathological
calcification and a potential inhibitor of calcium phosphate
mineralization (14–16). ANKH is the human homolog of the
murine ANK gene (15). It has been
demonstrated that the loss of ANK transporter function in mice led
to excessive mineralization due to reduced PPi transport (17), which was later confirmed by other
studies (18,19). Gurley et al (18) illustrated the role of ANK in PPi
transport and suggested that ANKH mutations may cause skeletal
diseases in humans. Ho et al (19) reported that ANK-deficient mice
displayed increased PPi levels and presented with mineralization
and skeletal ankylosis resembling AS. Thus, these previous studies
indicated that ANK may regulate ectopic mineralization in animal
models.
AS is characterized by pathological osteogenesis
(20). Previous studies suggested
that fibroblasts may be the target of heterotopic ossification
because these cells belong to the same lineage as osteoblasts and
were discovered to express osteogenesis-related genes, such as
BMP2, osteocalcin and Runx2 (21,22).
Fibroblasts are resident cells that preserve the extracellular
matrix, both in healthy conditions and during inflammation
(23). Thus, fibroblasts were
suggested as likely contenders to be involved in osteogenesis
(24). Fibroblasts and osteoblasts
both originate from mesenchymal stem cell with overlapping
phenotypes and through similar differentiation pathways, allowing
fibroblasts to differentiate into osteoblasts and osteoblasts to
differentiate into fibroblasts (25). In addition, the proliferative and
osteogenic potential of fibroblasts was discovered to increase
following the stimulation with fibronectin or BMP2 (26,27).
Therefore, fibroblasts may serve as important mediators of
osteogenesis.
The aim of the present study was to determine the
function of ANKH in fibroblasts isolated from patients with AS.
ANKH expression levels were analyzed in ligament tissue collected
from patients with AS, and explored the effect of ANKH on the
fibroblast cell viability, mineralization and ossification
differentiation. From the results, it was hypothesized that the
regulation of ANKH may affect fibroblast cell mineralization and
ossification in AS.
Materials and methods
Patients and tissue samples
Ligament tissue samples were collected from four
patients with AS (male; age, 18–42) and four patients with spinal
fracture (JK) (male; age, 22–44) who attended The People's Hospital
of Xinchang Hospital in March 2019 for treatment, according to the
modified New York criteria (28).
The patients used nonsteroidal anti-inflammatory drugs or
disease-modifying anti-rheumatic drugs, but not anti-tumor necrosis
factor. Written informed consent forms were signed by all patients.
The study was approved by The Ethics Committees of the People's
Hospital of Xinchang (Xinchang, China; approval no.
XC201902114).
Cell culture
The ligaments samples from patients with AS and JK
were separated in 60-mm sterile Petri dishes, and washed three
times with D-Hank's balanced salt solution (HBSS; cat. no.
14170161; Gibco; Thermo Fisher Scientific, Inc.). The tissues were
then sectioned into 1–3-mm3 blocks and washed with HBSS
twice. The blocks were maintained in a 25-cm2 culture
flask, with 4 mm space in between tissue blocks. Next, 2 ml
high-glucose DMEM (cat. no. 11965118; Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (cat. no. A3160901;
Gibco; Thermo Fisher Scientific, Inc.) was added to the flask. The
flask was placed vertically inside an incubator at 37°C with 5%
CO2 for 4 h. The bottle was gently laid flat at the end
of this incubation, and the medium was changed every three days
thereafter. After 4–8 days, when the cells reached 80% confluence,
they were washed with HBSS and added to 1 ml 0.25% trypsin solution
(cat. no. R001100; Gibco; Thermo Fisher Scientific, Inc.) at 37°C.
After 2 min, 3 ml DMEM supplemented with 100 U/ml penicillin, 100
mg/ml streptomycin and 10% FBS was immediately added to terminate
cell dissociation. The cells were collected and centrifuged in a
15-ml centrifuge tube at 1,000 × g for 5 min at 4°C. The cell
suspension was then divided (1:2) for secondary culture at 37°C
with 5% CO2. The medium was changed every two days, and
the cells were further subcultured in an incubator at 37°C with 5%
CO2 whenever confluence reached 80–90%. Cells in the
third generation were used for experimentation following
examination of cell morphology under an inverted phase-contrast
microscope (magnification, ×100; Chongqing UOP Photoelectric
Technology Co., Ltd.).
Immunocytochemistry
Immunocytochemistry was performed to analyze the
expression levels of vimentin in fibroblasts isolated from patients
with AS and JK. Slides covered with fibroblast cells were washed
with PBS three times for 5 min each time. After washing, 4%
paraformaldehyde was added to each slide for 15 min at room
temperature for fixed slides. Excess amounts of paraformaldehyde
was removed, and the slices were then washed again with PBS three
times for 5 min each time. Subsequently, 0.03% Triton X-100 was
added to the slides for 25 min at room temperature and further
incubated with 3% H2O2 for 10 min at room
temperature. After blocking with 2% BSA (cat. no. 11021037; Gibco;
Thermo Fisher Scientific, Inc.) for 30 min at room temperature.
After washing with PBS three times for 5 min each time, the slides
were then incubated with 50 µl anti-vimentin antibody (1:200; cat.
no. ab193555; Abcam) overnight at 4°C. Following the primary
antibody incubation, the slides were washed with PBS three times
for 5 min each time, then incubated with 50 µl goat anti-rabbit IgG
H&L (DyLight® 488) preabsorbed secondary antibody
(1:100; cat. no. ab96899; Abcam) for 30 min at 37°C and then washed
again with PBS three times. Staining was then visualized using a
DAB staining kit (cat. no. E670033; Sangon Biotech Co., Ltd.) for
3–5 min at room temperature, and the slides were examined under an
inverted phase-contrast microscope (magnification, ×400).
Cell transfection
The ANKH-pCMV6-XL5 overexpression plasmid (cat. no.
SC120218) and empty vector negative control (NC; cat. no. PCMV6XL5)
were purchased from OriGene Technologies, Inc. Fibroblasts from
patients with AS were transfected with ANKH overexpression vector
or NC. Fibroblasts from patients with JK were transfected with
small interfering RNA (si) specific for ANKH (siANKH) or siNC,
siANKH (5′-UCACUAUAAGCUAUCAGUGUG-3′) and siNC
(5′-UUCUCCGAACGUGUCACGU-3′) were obtained from Guangzhou RiboBio
Co., Ltd.
Fibroblasts were seeded into 24-well culture plates
at a density of 2×104 cells/well one day before
transfection and cultured to 90% confluence in 0.5 ml medium with
10% FBS without antibiotics. On the day of transfection, 20 pmol
siANKH/siNC, or 2 µg ANKH/NC were separately diluted in 50 µl DMEM
(cat. no. 12491015; Gibco; Thermo Fisher Scientific, Inc.). A
volume of 1 µl Lipofectamine® 2000 (cat. no. 11668019;
Invitrogen; Thermo Fisher Scientific, Inc.) was separately diluted
in 50 µl DMEM and incubated at room temperature for 5 min. The
diluted siANKH, siNC, ANKH or NC was then mixed with diluted
Lipofectamine® 2000 and placed at room temperature for
20 min, then added to the cells. Finally, the cells were incubated
at 37°C with 5% CO2 for 48 h, then harvested for
subsequent experimentation. Untransfected cells also served as an
additional control group.
Cell viability assay
Fibroblasts from patients with AS or JK were seeded
into 96-well culture plates at 200 µl per well, then incubated for
1–6 days. Following the incubation, an MTT assay was used to detect
cell viability. Briefly, 20 µl MTT (5 mg/ml; cat. no. M6494;
Invitrogen; Thermo Fisher Scientific, Inc.) was added to the cells
for 4 h at 37°C. After discarding the supernatant, 100 µl DMSO
(cat. no. D12345; Invitrogen; Thermo Fisher Scientific, Inc.) was
added to each well to dissolve the formazan crystals. The
absorbance was measured at 490 nm at the initial timepoint, then
every day over a time course of 6 days using a microplate reader.
In a separate experiment, absorbance was read at 24, 48 and 72 h
following transfection; the rest of the experimental details were
unchanged.
Flow cytometric analysis of
apoptosis
Apoptosis was analyzed using an Annexin
V-FITC/propidium iodide (PI) staining kit (cat. no. V13242;
Invitrogen; Thermo Fisher Scientific, Inc.). Fibroblasts
(2×105 cells/well) were cultured in 6-well plates for 24
h at 37°C, then washed with cold PBS and centrifuged at 1,000 × g
(5 min, 4°C). The cells were subsequently resuspended in 1X Annexin
binding buffer, 5 µl Annexin V-FITC and 5 µl PI was added to the
cells. The cells were incubated at room temperature for 15 min,
then added to 400 µl 1X Annexin binding buffer. Cell apoptosis was
assessed by flow cytometry using a FACScan instrument (BD
Biosciences) equipped with CellQuest software (version 5.1; BD
Biosciences). The apoptosis rate was calculated as the sum of
early-apoptotic and late-apoptotic ell frequencies. Unstained
cells, as well as Annexin V and PI single-stained cells were used
as controls, and the gates for positive staining were set according
to these controls.
Reverse transcription-quantitative PCR
(RT-qPCR)
Relative ANKH mRNA expression levels were analyzed
using RT-qPCR. Briefly, total RNA was extracted from fibroblasts
using TRIzol® reagent (cat. no. 15596018; Invitrogen;
Thermo Fisher Scientific, Inc.). RNA concentration was measured
using a NanoDrop™ 8000 spectrophotometer (Thermo Fisher Scientific,
Inc.). In addition, RNA integrity was confirmed by visualization of
the fragments in 1.5% agarose gel electrophoresis (Invitrogen;
Thermo Fisher Scientific, Inc.) followed by ethidium bromide and
visualized under ultraviolet light.
RT into cDNA was performed using a PrimeScript RT
Reagent kit with gDNA Eraser (cat. no. RR047A; Clontech
Laboratories, Inc.). Briefly, 7 µl RNA, 1 µl gDNA Eraser and 2 µl
5X gDNA Eraser buffer were added into a reaction tube and incubated
at 42°C for 2 min. The 10 µl sample was then mixed with 1 µl
PrimeScript RT enzyme mix I, 1 µl RT primer mix, 4 µl 5X
PrimeScript buffer II and 4 µl RNase-free distilled water. All
samples were incubated at 37°C for 15 min and at 85°C for 5 sec,
then allowed to cool down on ice. The resulting cDNA concentration
and OD values were measured on a NanoDrop 8000 spectrophotometer.
RT-qPCR was subsequently performed using a 7500 Real-Time PCR
system (cat. no. 4351105; Applied Biosystems; Thermo Fisher
Scientific, Inc.) with SYBR® Green PCR Master Mix
(4312704, Thermo Fisher Scientific, Inc.). The thermocycling
conditions consisted of an initial denaturation at 95°C for 10 min
followed by 40 cycles at 94°C for 5 sec and 60°C for 1 min. The
experiment was carried out three times. The relative expression
levels of the mRNA was analyzed using the 2−ΔΔCq method
(29), and ANKH expression was
normalized to GAPDH. All primers used in in the present study were
purchased from Sangon Biotech Co., Ltd. and the primer sequences
are presented in Table I.
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| Ankylosis
progressive | F:
GTGGGCCTGGTGTTTGTGAA |
| homolog | R:
CCTTCTCGTCTTGCTCCCC |
| GAPDH | F:
TGGATTTGGACGCATTGGTC |
|
| R:
TTTGCACTGGTACGTGTTGAT |
Alizarin Red staining
A 0.1% Alizarin Red dye liquor (pH 8.3) was prepared
by dissolving 0.1 g Alizarin Red powder (cat. no. 130-22-3;
Guidechem) in 100 ml Tris-HCl buffer (0.1 mol/l; cat. no.
AP-9005-125; Thermo Fisher Scientific, Inc.). Fibroblasts isolated
from the ligament tissue of patients with AS or JK
(2×104 cells/well) were seeded into six-well plates. A
cover glass was added to the 6-well plate before the cells were
inoculated. When the cells reached ~50% confluence, the samples
were collected, washed in PBS, then fixed in 4% methanol for 15 min
at room temperature. The cells were then washed again with PBS
three times for 3 min each time. Each cover glass was incubated
with 0.1% Alizarin Red dye liquor for 30 min at 37°C, then washed
using double steaming water three times for 3 min each time. The
slides were examined under an optical microscope (magnification,
×400), and Alizarin Red staining absorbance was read at 510 nm for
quantification.
Western blotting
Proteins were extracted from ligament tissue and
fibroblast by using RIPA (cat. no. 89900; Thermo Fisher Scientific,
Inc.). Protein concentration in the supernatant was measured using
a Pierce™ BCA Protein assay kit (cat. no. 23225; Thermo Fisher
Scientific, Inc.). A total of 20–40 µg protein lysate was separated
by SDS-PAGE on 12% gels (cat. no. P0012A; Beyotime Institute of
Biotechnology), then transferred onto PVDF membranes (cat. no.
FFP28; Beyotime Institute of Biotechnology). The membranes were
subsequently blocked using 5% non-fat milk in TBS containing 0.1%
Tween 20 (cat. no. TA-999-TT; Thermo Fisher Scientific, Inc.) at
room temperature for 1 h. The membranes were then incubated
overnight at 4°C with the following primary antibodies: Anti-GADPH
(cat. no. ab8245; 1:1,000; 36 kDa; Abcam), anti-ANKH (cat. no.
ab90104; 1:1,000, 54 kDa; Abcam), anti-alkaline phosphatase (ALP;
cat. no. ab83259; 1:1,000, 39 kDa; Abcam), anti-osteocalcin (OCN;
cat. no. ab93876; 1:500; 11 kDa; Abcam), anti-Runt-related
transcription factor 2 (Runx2; cat. no. ab23981; 1:1,000; 60 kDa;
Abcam), anti-phosphorylated (p)-β-catenin (cat. no. 9561; 1:1,000,
92 kDa; Cell Signaling Technology, Inc.), anti-β-catenin (cat. no.
9562; 1:1,000, 92 kDa; Cell Signaling Technology, Inc.) and
anti-c-Myc (cat. no. ab32072; 1:1,000; 57 kDa; Abcam). Following
the primary antibody incubation, the membranes were incubated with
a goat anti-rabbit IgG horseradish peroxidase-conjugated secondary
antibody (cat. no. ab205718; 1:5,000, 42 kDa; Abcam) at room
temperature for 1 h. Protein bands were visualized using an
enhanced chemiluminescence reagent kit (cat. no. 345818; EMD
Millipore). Protein expression levels were analyzed and normalized
to GAPDH using ImageJ software (version 1.5, National Institutes of
Health).
Statistical analysis
Statistical analysis was performed using SPSS 16.0
software (SPSS, Inc.) and data are presented as the mean ± SEM of
three experiments. An unpaired Student's t-test was used to analyze
statistical differences between two groups. Multi-group comparisons
were performed using one-way ANOVA followed by a Tukey's post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
ANKH expression levels in ligament
tissue from patients with AS
The expression levels of ANKH were investigated in
ligament tissues collected from patients with AS and JK controls
(n=4/group) using western blot analysis. The results revealed that
ANKH expression levels were significantly downregulated in the AS
group compared with the JK group (P<0.001; Fig. 1A and B).
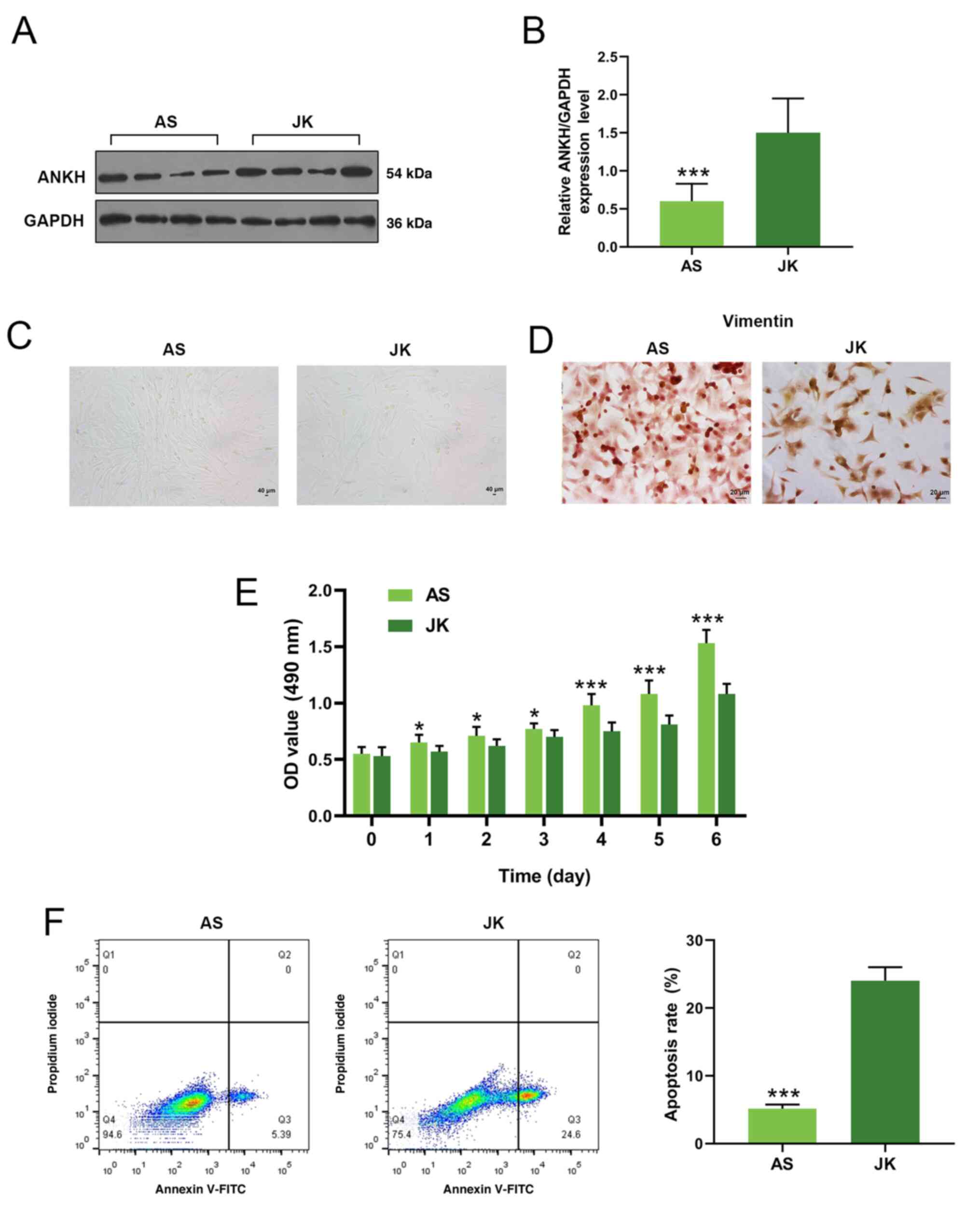 | Figure 1.ANKH protein expression levels, cell
viability and apoptosis in fibroblasts from patients with AS and JK
controls. (A) ANKH protein expression levels in ligament tissue
collected from patients with AS and JK controls were analyzed using
western blotting. (B) Densitometry data for ANKH expression levels
from part (A). Data are presented as the mean ± SEM. (C)
Fibroblasts morphology was examined under a light microscope.
Magnification, ×100. (D) Immunocytochemistry was performed to
analyze vimentin expression levels in AS and JK fibroblasts.
Magnification, ×400. (E) MTT assays were performed to measure cell
viability over a time course of six days. (F) Apoptotic rates of AS
or JK fibroblasts were analyzed using flow cytometry. Each dot plot
represents necrotic cells (Q1), late apoptotic cells (Q2), early
apoptotic cells (Q3) and viable cells (Q4). The apoptosis rate
(late and early apoptosis) is summarized as a graph on the
right-hand side. *P<0.05, ***P<0.001 vs. JK. ANKH, ankylosis
progressive homolog; AS, ankylosing spondylitis, JK, spinal
fracture; OD, optical density; Q, quadrant. |
Fibroblast isolation and culture
Fibroblasts from the AS and JK groups were isolated
from the ligament and cultured. The majority of the cells were
shuttle-shaped and flat, with 2–3 protuberance, displaying the
typical characteristics of fibroblasts (Fig. 1C). Notably, there were no
observable differences in the morphology between the two groups.
The intermediate filament protein vimentin is an important protein
in mammalian fibroblasts (30).
Fibroblasts from both groups were used for immunocytochemical
staining of vimentin; the expression levels of vimentin in the AS
and JK groups were positive (Fig.
1D).
Cell viability and apoptosis of
fibroblasts
Cell viability and apoptosis were investigated in
the fibroblasts from patients with AS and JK controls. The
fibroblast viability in the AS group was significantly increased
compared with the JK group (P<0.05 at days 1–3; P<0.001 at
days 4–6; Fig. 1E). Moreover, the
apoptotic rate, representing both early and late apoptosis, was
significantly reduced in fibroblasts from patients with AS compared
with the JK group (P<0.001; Fig.
1F).
ANKH overexpression inhibits AS
fibroblast viability, mineralization and ossification
The effect of ANKH on cell viability, mineralization
and ossification differentiation in AS was evaluated by
transfecting ANKH overexpression vectors into fibroblasts from
patients with AS and siANKH into fibroblasts from patients with JK.
In the AS group, the relative ANKH mRNA expression levels were
significantly upregulated following the transfection with the ANKH
overexpression vector compared with the control and NC groups
(P<0.001; Fig. 2A). By
contrast, the transfection with siANKH in JK-derived fibroblasts
significantly downregulated ANKH expression levels compared with
the control and siNC group (P<0.001; Fig. 2B).
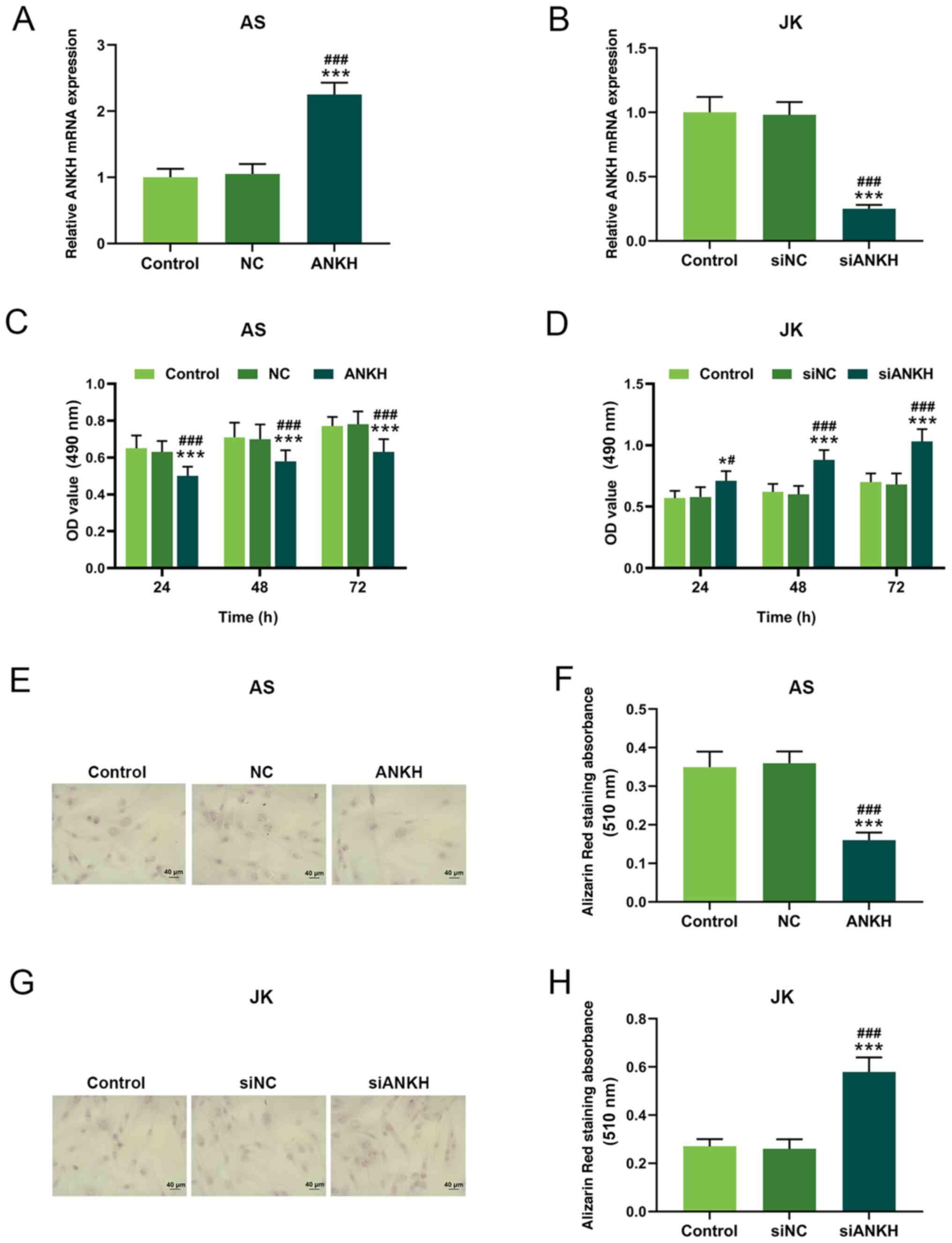 | Figure 2.ANKH expression levels affect cell
viability and mineralization. ANKH mRNA expression levels in
fibroblasts from (A) patients with AS transfected with an ANKH
overexpression vector or NC or (B) patients with JK transfected
with siANKH or siNC. Cell viability following (C) ANKH
overexpression in the AS group and (D) ANKH silencing in the JK
group. (E) Mineralization levels were evaluated using Alizarin Red
staining in ANKH-overexpressing AS fibroblasts. Magnification,
×400. (F) Mineralization levels were semi-quantified in fibroblasts
from patients with AS following ANKH overexpression. (G)
Mineralization levels were evaluated using Alizarin Red staining
following ANKH silencing in JK fibroblasts. Magnification, ×400.
(H) Mineralization levels were semi-quantified in fibroblasts from
patients with JK following siANKH transfection. *P<0.05,
***P<0.001 vs. control; #P<0.05,
###P<0.001 vs. NC or siNC. ANKH, ankylosis
progressive homolog; AS, ankylosing spondylitis, JK, spinal
fracture; OD, optical density; si, small interfering RNA; NC,
negative control. |
An MTT assay was performed to determine cell
viability in both groups at 24, 48 and 72 h following transfection.
Following transfection with the ANKH overexpression vector, the
viability of fibroblasts in the AS group was significantly reduced
compared with the control and siNC groups at each time point
(P<0.05; Fig. 2D). However, the
cell viability in the JK group was significantly increased,
particularly 48 and 72 h after transfection, compared with the
control group and the siNC group at each time point (P<0.05;
Fig. 2D).
Moreover, Alizarin Red absorbance in
ANKH-overexpressing fibroblasts from the AS group was significantly
reduced compared with the control and NC groups (P<0.001;
Fig. 2E and F). In comparison,
Alizarin Red absorbance in the ANKH-silenced JK group was
significantly increased compared with the siNC and control groups
(P<0.001; Fig. 2G and H).
The expression levels of ossification markers were
analyzed following transfection using western blotting analysis.
ALP, OCN and Runx2 expression levels were significantly
downregulated following ANKH overexpression in AS fibroblasts
compared with the control and NC groups (P<0.001; Fig. 3A and B). By contrast, the
expression levels of these ossification markers were significantly
upregulated following siANKH silencing in JK fibroblasts compared
with the control and siNC groups (P<0.001; Fig. 3C and D).
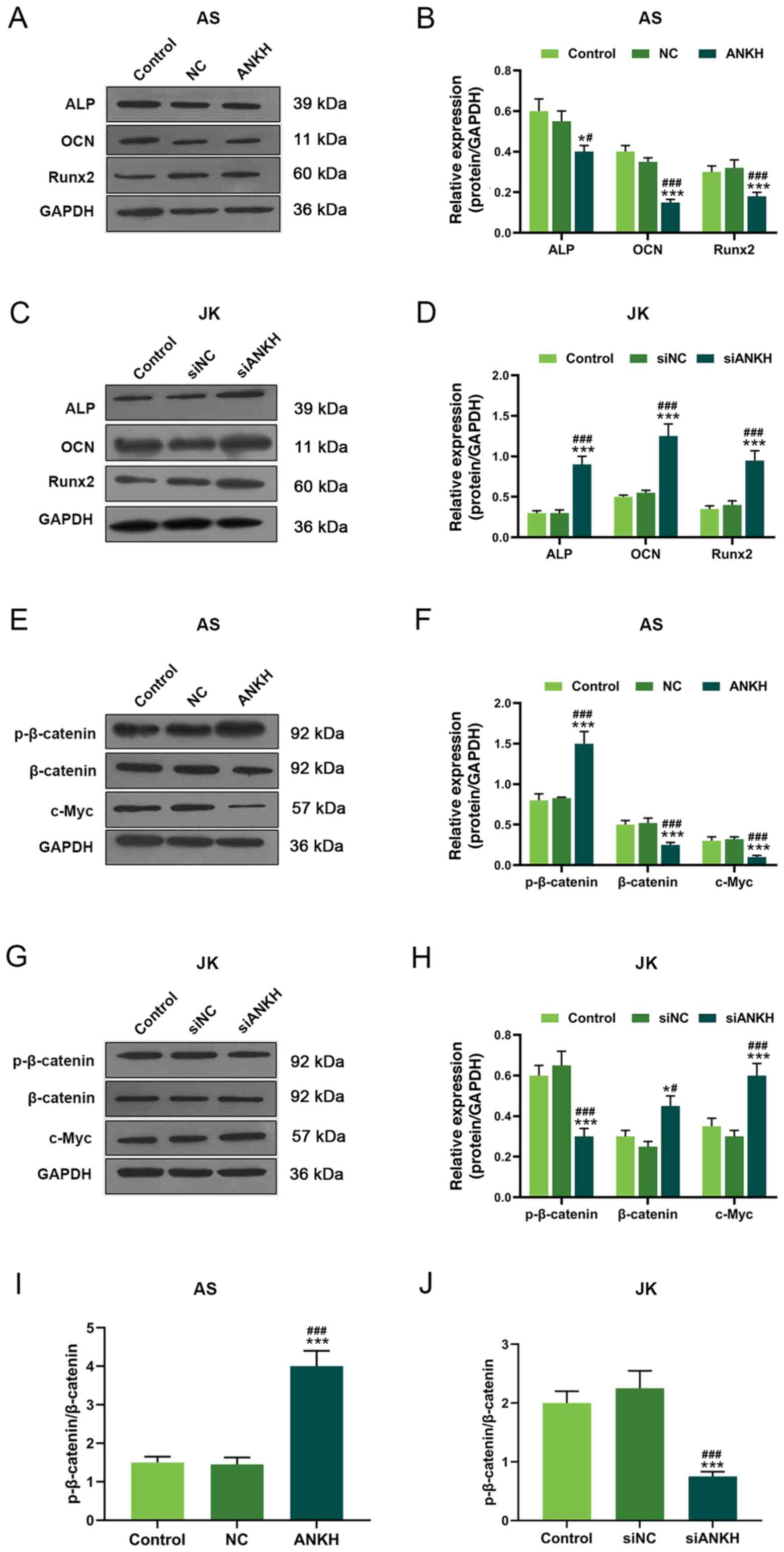 | Figure 3.ANKH regulates the expression levels
of ossification markers and Wnt signaling-related proteins. (A)
Expression levels of ALP, OCN and Runx2 in AS fibroblasts
overexpressing ANKH were analyzed using western blotting. (B)
Semi-quantification of the expression levels presented in part (A).
(C) Expression levels of ALP, OCN and Runx2 in JK fibroblasts
transfected with siANKH were analyzed using western blotting. (D)
Semi-quantification of the expression levels presented in part (C).
Expression levels of p-β-catenin, β-catenin and c-Myc in AS
fibroblasts overexpressing ANKH were analyzed using western
blotting. (F) Semi-quantification of the expression levels
presented in part (E). (G) Expression levels of p-β-catenin,
β-catenin and c-Myc in JK fibroblasts transfected with siANKH. (H)
Semi-quantification of the expression levels presented in part (G).
p-β-catenin: Total β-catenin expression ratios following (I) ANKH
overexpression in AS fibroblasts or (J) ANKH silencing in JK
fibroblasts. *P<0.05, ***P<0.001 vs. control;
#P<0.05, ###P<0.001 vs. NC or siNC.
ANKH, ankylosis progressive homolog; AS, ankylosing spondylitis,
JK, spinal fracture; si, small interfering RNA; NC, negative
control; p-, phosphorylated; ALP, alkaline phosphatase; OCN,
osteocalcin; Runx2, Runt-related transcription factor 2. |
ANKH overexpression affects the
Wnt/β-catenin signaling pathway in fibroblasts
The Wnt/β-catenin signaling pathway governs the
differentiation of ossification and plays an important role in
heterotopic ossification (25,31).
The overexpression of ANKH significantly promoted β-catenin
phosphorylation, whilst significantly downregulating the expression
levels of β-catenin and c-Myc in AS fibroblasts compared with the
control and NC groups (P<0.001; Fig. 3E and F). However, siANKH
significantly upregulated the expression levels of β-catenin
(P<0.05) and c-Myc (P<0.001), while inhibiting β-catenin
phosphorylation (P<0.001) compared with the control and siNC
groups in JK fibroblasts (Fig. 3G and
H). In addition, the p-β-catenin/β-catenin ratios were
significantly increased in the ANKH group compared with the
untransfected control and NC groups in the AS-derived fibroblasts;
however, the p-β-catenin/β-catenin ratio was significantly reduced
in the siANKH group compared with the untransfected control and
siNC groups in JK fibroblasts (P<0.001; Fig. 3I and J).
Discussion
The aim of the present study was to evaluate the
role of ANKH in the ossification and mineralization of fibroblasts
in patients with AS. The overexpression of the ANKH protein
inhibited the viability and mineralization of fibroblasts in
patients with AS. However, ANKH silencing led to the opposite
effect, indicating a potential anti-osteogenic role for ANKH in
AS.
The ANKH gene encodes a multi-channel transmembrane
protein, which has a previously characterized role in genetic
susceptibility to AS (32,33). A previous study suggested that ANKH
did not significantly affect the susceptibility to or clinical
manifestations of AS (32), which
contradicted the results of another study (33), in which ANKH was associated with
genetic susceptibility to AS in a sex-specific manner (33).
Previous studies have also evaluated the role of
ANKH in heterotopic ossification (34,35).
For instance, Gurley et al (34) demonstrated that the deletion of the
ANK gene caused progressive mineralization and joint disease,
leading to stiffness of the spine, a symptom similar to AS in
humans. Moreover, loss of ANKH function resulted in pathological
hydroxyapatite formation (35).
Consistent with these previous studies, the present findings
demonstrated that the expression levels of ANKH in ligaments from
patients with AS were downregulated compared with the control
subjects.
Moreover, the cellular morphology of fibroblasts
from patients with AS and control subjects was also examined.
Typical fibroblast cellular characteristics and positive vimentin
expression confirmed that cultured cells isolated from the ligament
retained a fibroblast phenotype. However, fibroblasts from patients
with AS displayed increased viability and reduced apoptotic rates
compared with the controls, indicating that AS pathogenesis may be
related to fibroblast growth. This observation is consistent with a
previous study, in which fibroblasts modulated osteoblast
metabolism and osteogenesis (24).
Therefore, it was hypothesized that inhibiting fibroblast viability
or promoting apoptosis may prevent pathological osteogenesis in
patients with AS.
In order to investigate the role of ANKH in
fibroblast viability and differentiation, the effects of ANKH
overexpression and silencing on fibroblasts were examined. The
present findings demonstrated an association between ANKH
expression levels and fibroblast viability and mineralization. ANKH
overexpression in fibroblasts from patients with AS reduced cell
viability and mineralization. Indeed, Alizarin Red staining
indicated that the number of mineralized nodules was reduced
following ANKH overexpression, suggesting a negative association
between ANKH expression levels and fibroblast cell mineralization.
Skubutyte et al (36)
demonstrated that ANK prevented pathological mineralization.
Similarly, Ho et al (19)
revealed that the ANK gene prevented mineralization, whereas ANK
deficiency accelerated mineralization in joints. Thus, it may be
possible to reduce fibroblast mineralization through ANKH
overexpression in order to prevent pathological bone formation.
The expression levels of osteogenic markers, such as
ALP, OCN and Runx2, were analyzed to determine the levels of
osteogenic differentiation and mineralization in fibroblasts from
patients with AS. ALP is a phenotypic marker for early-stage
osteoblast differentiation that serves a role in bone
mineralization (37). In the
present study, ANKH overexpression downregulated ALP expression
levels in fibroblasts from patients with AS. This result was
consistent with a previous study, which demonstrated that ANK was
negatively associated with ALP expression levels in bone marrow
stromal cells (38). OCN is a
marker unique to osteoblasts and a late marker of osteoblast
differentiation (38,39). In the present study, the expression
levels of OCN and Runx2 were downregulated in ANKH-overexpressing
cells. Runx2 can stimulate the transcription of osteoblast-related
genes, such as those encoding OPN and OCN (40). Hill et al (41) also demonstrated that ANKH
overexpression suppressed the mineralization and ossification of
fibroblasts.
Wnt/β-catenin signaling induces mesenchymal stem
cell differentiation, a precursor of osteoblastic activity
(42). Day et al (43) suggested that Runx2 expression
levels were possibly regulated by β-catenin upregulation.
Conversely, the inhibition of Wnt signaling was illustrated to
impair osteogenic differentiation in another previous study
(44). β-catenin serves an
important role in osteocyte viability, differentiation,
proliferation and new bone formation (45,46).
In fact, during the early and late stages of fracture repair,
β-catenin activation is essential for osteoblast differentiation
(45,46), and the loss of β-catenin resulted
in increased bone resorption (47). The present findings were consistent
with these aforementioned previous studies. Canonical Wnt/β-catenin
signaling was reported to stimulate the proliferation and
differentiation from fibroblasts to myofibroblast cell (48). In the present study, ANKH silencing
reduced p-β-catenin expression levels, thereby offering a possible
mechanism through which the cell viability and mineralization of
normal fibroblasts were stimulated. However, ANKH overexpression in
fibroblasts from patients with AS led to the opposite result. In
addition, c-Myc is an important downstream target protein of the
Wnt signaling pathway that can regulate proliferation,
differentiation and apoptosis (49). Loveridge et al (50) reported that downregulated c-Myc
protein levels prevented chondrocyte mineralization. In the present
study, a negative association was identified between ANKH and c-Myc
expression levels. Indeed, the overexpression of ANKH in
fibroblasts from patients with AS downregulated the expression
levels of c-Myc, resulting in reduced mineralization, whereas ANKH
silencing in normal fibroblasts led to c-Myc upregulation. Thus,
ANKH overexpression may reduce mineralization and ossification in
AS through c-Myc downregulation and increased β-catenin
phosphorylation and p-β-catenin/β-catenin, thereby inhibiting
pathological bone formation.
However, there were some limitations to the present
study. The immunofluorescence identification of JK fibroblasts by
vimentin is also required. In addition, the positive effects of
ANKH on AS fibroblasts was only determined in vitro; thus,
the present findings should be further validated in vivo.
Moreover, the potential use of the ANKH gene in clinical treatment
also requires validation. Finally, although this preliminary study
suggested that ANKH may regulate the Wnt/β-catenin pathway to
inhibit the viability, mineralization and ossification of
fibroblasts, the regulatory mechanism underlying this pathway
requires further investigation.
In conclusion, the findings of the present study
revealed the role of ANKH in fibroblasts isolated from the
ligaments of patients with AS. The results discovered that ANKH
expression levels were downregulated in ligaments of patients with
AS. Moreover, fibroblasts from patients with AS displayed increased
cell viability and reduced levels of apoptosis. ANKH overexpression
was discovered to inhibit cell mineralization and ossification,
which was likely mediated through its effect on ossification
markers and the Wnt/β-catenin signaling pathway. Therefore, these
results suggested that ANKH overexpression may prevent or delay new
bone formation in AS.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XH designed the study. XH and YD acquired, analyzed
and interpreted the data, drafted the manuscript and critically
revised it for important intellectual content. XH and YD authors
agree to be accountable for all aspects of the work in ensuring
that questions related to the accuracy or integrity of the work are
appropriately investigated and resolved. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The Ethics
Committees of The People's Hospital of Xinchang (approval no.
XC201902114). All procedures involving human participants were in
accordance with the 1964 Declaration of Helsinki and its later
amendments or comparable ethical standards. Written informed
consent was provided by all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ANKH
|
ankylosis progressive homolog
|
|
AS
|
ankylosing spondylitis
|
|
ALP
|
alkaline phosphatase
|
|
OCN
|
osteocalcin
|
|
Runx2
|
Runt-related transcription factor
2
|
|
RT-qPCR
|
reverse transcription- quantitative
PCR
|
References
|
1
|
Smith JA: Update on ankylosing
spondylitis: Current concepts in pathogenesis. Curr Allergy Asthma
Rep. 15:4892015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Leone A, Marino M, Dell'Atti C, Zecchi V,
Magarelli N and Colosimo C: Spinal fractures in patients with
ankylosing spondylitis. Rheumatol Int. 36:1335–1346. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Woodward LJ and Kam PC: Ankylosing
spondylitis: Recent developments and anaesthetic implications.
Anaesthesia. 64:540–548. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brown MA and Wordsworth BP: Genetics in
ankylosing spondylitis-current state of the art and translation
into clinical outcomes. Best Pract Res Clin Rheumatol. 31:763–776.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hanson A and Brown MA: Genetics and the
causes of ankylosing spondylitis. Rheum Dis Clin North Am.
43:401–414. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen S, Li Y, Deng C, Li J, Wen X, Wu Z,
Hu C, Zhang S, Li P, Zhang X, et al: The associations between PD-1,
CTLA-4 gene polymorphisms and susceptibility to ankylosing
spondylitis: A meta-analysis and systemic review. Rheumatol Int.
36:33–44. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Karami J, Mahmoudi M, Amirzargar A,
Gharshasbi M, Jamshidi A, Aslani S and Nicknam MH: Promoter
hypermethylation of BCL11B gene correlates with downregulation of
gene transcription in ankylosing spondylitis patients. Genes Immun.
18:170–175. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang P, Li Q, Qi J, Lv Q, Zheng X, Wu X
and Gu J: Association between vitamin D receptor gene polymorphism
and ankylosing spondylitis in Han Chinese. Int J Rheum Dis.
20:1510–1516. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Golder V and Schachna L: Ankylosing
spondylitis: An update. Aust Fam Physician. 42:780–784.
2013.PubMed/NCBI
|
|
10
|
Wellcome Trust Case Control Consortium;
Australo-Anglo- American Spondylitis Consortium (TASC), ; Burton
PR, Clayton DG, Cardon LR, Craddock N, Deloukas P, Duncanson A,
Kwiatkowski DP, McCarthy MI, et al: Association scan of 14,500
nonsynonymous SNPs in four diseases identifies autoimmunity
variants. Nat Genet. 39:1329–1337. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brown MA, Kenna T and Wordsworth BP:
Genetics of ankylosing spondylitis-insights into pathogenesis. Nat
Rev Rheumatol. 12:81–91. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ranganathan V, Gracey E, Brown MA, Inman
RD and Haroon N: Pathogenesis of ankylosing spondylitis-recent
advances and future directions. Nat Rev Rheumatol. 13:359–367.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kirsch T, Kim HJ and Winkles JA:
Progressive ankylosis gene (ank) regulates osteoblast
differentiation. Cells Tissues Organs. 189:158–162. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kanaujiya J, Bastow E, Luxmi R, Hao Z,
Zattas D, Hochstrasser M, Reichenberger EJ and Chen IP: Rapid
degradation of progressive ankylosis protein (ANKH) in
craniometaphyseal dysplasia. Sci Rep. 8:157102018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mitton-Fitzgerald E, Gohr CM, Bettendorf B
and Rosenthal AK: The role of ANK in calcium pyrophosphate
deposition disease. Curr Rheumatol Rep. 18:252016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang W, Xu J, Du B and Kirsch T: Role of
the progressive ankylosis gene (ank) in cartilage mineralization.
Mol Cell Biol. 25:312–323. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Williams CJ: The role of ANKH in
pathologic mineralization of cartilage. Curr Opin Rheumatol.
28:145–151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gurley KA, Reimer RJ and Kingsley DM:
Biochemical and genetic analysis of ANK in arthritis and bone
disease. Am J Hum Genet. 79:1017–1029. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ho AM, Johnson MD and Kingsley DM: Role of
the mouse ank gene in control of tissue calcification and
arthritis. Science. 289:265–270. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qin X, Jiang T, Liu S, Tan J, Wu H, Zheng
L and Zhao J: Effect of metformin on ossification and inflammation
of fibroblasts in ankylosing spondylitis: An in vitro study. J Cell
Biochem. 119:1074–1082. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu F, Cui Y, Zhou X, Zhang X and Han J:
Osteogenic differentiation of human ligament fibroblasts induced by
conditioned medium of osteoclast-like cells. Biosci Trends.
5:46–51. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rutherford RB, Moalli M, Franceschi RT,
Wang D, Gu K and Krebsbach PH: Bone morphogenetic
protein-transduced human fibroblasts convert to osteoblasts and
form bone in vivo. Tissue Eng. 8:441–452. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Turner JD, Naylor AJ, Buckley C, Filer A
and Tak PP: Fibroblasts and osteoblasts in inflammation and bone
damage. Adv Exp Med Biol. 1060:37–54. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
da Costa Fernandes CJ, do Nascimento AS,
da Silva RA and Zambuzzi WF: Fibroblast contributes for
osteoblastic phenotype in a MAPK-ERK and sonic hedgehog
signaling-independent manner. Mol Cell Biochem. 436:111–117. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zou YC, Yang XW, Yuan SG, Zhang P, Ye YL
and Li YK: Downregulation of dickkopf-1 enhances the proliferation
and osteogenic potential of fibroblasts isolated from ankylosing
spondylitis patients via the Wnt/β-catenin signaling pathway in
vitro. Connect Tissue Res. 57:200–211. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang WH, Li XL, Guo Y and Zhang Y:
Proliferation and osteogenic activity of fibroblasts induced with
fibronectin. Braz J Med Biol Res. 50:e62722017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen F, Bi D, Cao G, Cheng C, Ma S, Liu Y
and Cheng K: Bone morphogenetic protein 7-transduced human
dermal-derived fibroblast cells differentiate into osteoblasts and
form bone in vivo. Connect Tissue Res. 59:223–232. 2018.PubMed/NCBI
|
|
28
|
van der Linden S, Valkenburg HA and Cats
A: Evaluation of diagnostic criteria for ankylosing spondylitis. A
proposal for modification of the New York criteria. Arthritis
Rheum. 27:361–368. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Turowski P, Myles T, Hemmings BA,
Fernandez A and Lamb NJ: Vimentin dephosphorylation by protein
phosphatase 2A is modulated by the targeting subunit B55. Mol Biol
Cell. 10:1997–2015. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tamamura Y, Otani T, Kanatani N, Koyama E,
Kitagaki J, Komori T, Yamada Y, Costantini F, Wakisaka S, Pacifici
M, et al: Developmental regulation of Wnt/beta-catenin signals is
required for growth plate assembly, cartilage integrity, and
endochondral ossification. J Biol Chem. 280:19185–19195. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Timms AE, Zhang Y, Bradbury L, Wordsworth
BP and Brown MA: Investigation of the role of ANKH in ankylosing
spondylitis. Arthritis Rheum. 48:2898–2902. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tsui HW, Inman RD, Paterson AD, Reveille
JD and Tsui FW: ANKH variants associated with ankylosing
spondylitis: gender differences. Arthritis Res Ther. 7:R513–R525.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gurley KA, Chen H, Guenther C, Nguyen ET,
Rountree RB, Schoor M and Kingsley DM: Mineral formation in joints
caused by complete or joint-specific loss of ANK function. J Bone
Miner Res. 21:1238–1247. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pendleton A, Johnson MD, Hughes A, Gurley
KA, Ho AM, Doherty M, Dixey J, Gillet P, Loeuille D, McGrath R, et
al: Mutations in ANKH cause chondrocalcinosis. Am J Hum Genet.
71:933–940. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Skubutyte R, Markova D, Freeman TA,
Anderson DG, Dion AS, Williams CJ, Shapiro IM and Risbud MV:
Hypoxia-inducible factor regulation of ANK expression in nucleus
pulposus cells: Possible implications in controlling dystrophic
mineralization in the intervertebral disc. Arthritis Rheum.
62:2707–2715. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sharma U, Pal D and Prasad R: Alkaline
phosphatase: An overview. Indian J Clin Biochem. 29:269–278. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Minashima T, Quirno M, Lee YJ and Kirsch
T: The role of the progressive ankylosis protein (ANK) in
adipogenic/osteogenic fate decision of precursor cells. Bone.
98:38–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wei J and Karsenty G: An overview of the
metabolic functions of osteocalcin. Rev Endocr Metab Disord.
16:93–98. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
An S, Gao Y and Ling J: Characterization
of human periodontal ligament cells cultured on three-dimensional
biphasic calcium phosphate scaffolds in the presence and absence of
L-ascorbic acid, dexamethasone and β-glycerophosphate in
vitro. Exp Ther Med. 10:1387–1393. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hill TP, Später D, Taketo MM, Birchmeier W
and Hartmann C: Canonical Wnt/beta-catenin signaling prevents
osteoblasts from differentiating into chondrocytes. Dev Cell.
8:727–738. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xu C, Wang J, Zhu T, Shen Y, Tang X, Fang
L and Xu Y: Cross-talking between PPAR and WNT signaling and its
regulation in mesenchymal stem cell differentiation. Curr Stem Cell
Res Ther. 11:247–254. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Day TF, Guo X, Garrett-Beal L and Yang Y:
Wnt/beta-catenin signaling in mesenchymal progenitors controls
osteoblast and chondrocyte differentiation during vertebrate
skeletogenesis. Dev Cell. 8:739–750. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Leucht P, Jiang J, Cheng D, Liu B,
Dhamdhere G, Fang MY, Monica SD, Urena JJ, Cole W, Smith LR, et al:
Wnt3a reestablishes osteogenic capacity to bone grafts from aged
animals. J Bone Joint Surg Am. 95:1278–1288. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Duan P and Bonewald LF: The role of the
wnt/β-catenin signaling pathway in formation and maintenance of
bone and teeth. Int J Biochem Cell Biol. 77:23–29. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen Y, Whetstone HC, Lin AC, Nadesan P,
Wei Q, Poon R and Alman BA: Beta-catenin signaling plays a
disparate role in different phases of fracture repair: Implications
for therapy to improve bone healing. PLoS Med. 4:e2492007.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Albers J, Keller J, Baranowsky A, Beil FT,
Catala-Lehnen P, Schulze J, Amling M and Schinke T: Canonical Wnt
signaling inhibits osteoclastogenesis independent of
osteoprotegerin. J Cell Biol. 200:537–549. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xu L, Cui WH, Zhou WC, Li DL, Li LC, Zhao
P, Mo XT, Zhang Z and Gao J: Activation of Wnt/β-catenin signalling
is required for TGF-β/Smad2/3 signalling during myofibroblast
proliferation. J Cell Mol Med. 21:1545–1554. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shi L, Wu YX, Yu JH, Chen X, Luo XJ and
Yin YR: Research of the relationship between β-catenin and
c-myc-mediated Wnt pathway and laterally spreading tumors
occurrence. Eur Rev Med Pharmacol Sci. 21:252–257. 2017.PubMed/NCBI
|
|
50
|
Loveridge N, Farquharson C, Hesketh JE,
Jakowlew SB, Whitehead CC and Thorp BH: The control of chondrocyte
differentiation during endochondral bone growth in vivo: Changes in
TGF-beta and the proto-oncogene c-myc. J Cell Sci. 105:949–956.
1993.PubMed/NCBI
|














