Introduction
As the first functional organ during embryonic
development, the heart must maintain a high level of efficiency
during the whole life cycle of the organism, from the early
formation of the primitive cardiac tube to the formation of the
chamber, and even in the later stages of development (1). Cardiac development involves numerous
genes regulated by multiple signaling pathways. Any genetic
dysregulation may lead to abnormal cardiac development and
congenital heart disease (CHD) (2). In previous decades, novel molecular
technologies have focused on the molecular mechanism of cardiac
development. It has been found that Nkx2 homeobox 5 (3), T-box transcription factor 5 (4) and GATA binding protein 4 (5) are closely related to heart
development and function. However, the gene regulatory network for
heart development remains to be elucidated.
MicroRNAs (miRs/miRNAs) are endogenous small RNAs
harboring 20–24 nucleotides. miRNAs interact with the
3-untranslated region (UTRs) of specific RNA targets to promote
homologous mRNA degradation, inhibit protein translation and
regulate gene expression at the post-transcriptional level
(6). Previous data have reported
that miRNAs play a critical role in various biological processes,
including cell proliferation, cell cycle, apoptosis (7), differentiation (8), cell migration and intracellular
signal transduction (9,10). Previous research has confirmed that
miRNAs are involved in the regulation of normal heart development
(11). The miR-1/miR-133 cluster
is the most abundant small RNA expressed in the heart (12), and is the first and most widely
studied miRNA among all heart development-specific miRNAs (1). miR-1 overexpression resulted in
thinning of the ventricular wall and embryo death at embryonic day
13.5 (E13.5) (13), whereas
deficiency of miR-1-2 led to embryo death at E15.5 due to poor
ventricular myocyte proliferation (12). Additionally, miR-133a was revealed
to negatively regulate cardiac myocyte proliferation (14,15).
Furthermore, miR-1 promoted the differentiation of embryonic stem
cells into cardiomyocytes during in vitro studies. However,
to investigate the molecular mechanisms underpinning these
processes, more regulatory miRNAs need to be studied.
Previously, it has been reported that miR-29c
expression in fetal CHDs was significantly upregulated (16,17),
which suggested that miR-29c may be another cardiac
development-specific miRNA. Further research revealed that miR-29c
could promote the differentiation and apoptosis, and inhibit the
proliferation of cardiomyocyte-like cells, possibly by suppressing
Wnt4 signaling in P19 embryonal carcinoma cells (17,18).
The present study aimed to observe the mechanism of miRNA-29c in
cardiac development.
Zebrafish have only one atrium and one ventricle. A
previous study reported that the embryonic cardiac development of
zebrafish is similar to that of mammals (19). Zebrafish have a shorter growth
cycle compared with mammals. Zebrafish embryos are transparent, and
the heart is located on the ventral side; hence early-stage cardiac
physiology can be easily observed. Consequently, zebrafish are
commonly used to model cardiovascular diseases. In the present
study, using a zebrafish model injected with miR-29c mimics, the
effect of miR-29c overexpression on embryonic heart development and
the activation of Wnt4 signaling pathway were observed. The present
study demonstrated that miR-29c regulated the lateral development
and cardiac circulation of zebrafish embryo by targeting Wnt4.
Materials and methods
Zebrafish feeding
According to protocols approved by the Research
Ethics Committee of Jiangsu Taizhou People's Hospital (Taizhou
Clinical Medical School of Nanjing Medical University), Female
zebrafish were reared in the zebrafish installation of the Model
Animal Research Center, Nanjing Medical University. As previously
described (20), the embryos
obtained from the natural oviposition of wild-type (WT) adults were
cultured in embryo medium, as previously described (21), at 28.5°C. As previously described
(22–24), embryonic development was staged
based on morphological characteristics. The zebrafish embryo
appears in a transparent form, but after ~24 h of development, it
begins to generate melanin, which hinders in vivo
observation and signal detection with fluorescence. Therefore, it
is necessary to inhibit the generation of melanin to maintain the
transparency of zebrafish embryo, thereby reducing errors in the
results. Thus, the embryos were incubated with 0.003%
1-phenyl-2-thiourea (Sigma-Aldrich; Merck KGaA) at 8 h
post-fertilization (hpf) to prevent melanocyte development.
miR-29c mimic synthesis and
microinjection
The present study was divided into three groups:
wild-type (WT), experimental (miR-29c mimics) and negative control
(NC) groups. The miR-29c mimics (5′-UGACCGAUUUCUCCUGGUGUUC-3′) and
NC (5′-UUCUCCGAACGUGUCACGUTT-3′) were synthesized by Shanghai
GenePharma Co., Ltd., and dissolved in 20 µm diethylpyrocarbonate.
Using a capillary needle filled with fine borosilicate glass on the
back, miR-29c mimics (1, 1.6 and 2 µM) or equivalent concentrations
of NC was injected into single to four-cell-stage embryos,
respectively. The injected embryos were cultured in a constant
temperature incubator at 28.5°C for future use. The interval
between transfection and subsequent experimentation was at least 24
h. Degenerated zebrafish embryos were removed at any time and the
solutions were replaced daily.
In situ hybridization probe
preparation
Probes of the following genes were synthesized for
in situ hybridization: Ventricular myosin heavy chain
(vmhc), atrial myosin heavy chain (amhc), natriuretic peptide
precursor A (nppa), cardiac myosin light chain-2 (cmlc2), bone
morphogenetic protein 4 (bmp4), notch homolog 1b (notch1b), fli-1
proto-oncogene ETS transcription factor a (fli1a). The
hybridization probes were amplified from the cDNA generated from
the embryos at 120 h post-fertilization. cDNA was synthesized from
500 ng RNA using a reverse transcriptase kit (Vazyme Biotech Co.,
Ltd.) with the following temperature protocol: 25°C for 5 min,
followed by 50°C for 15 min and 85°C for 5 min (25). Then, cDNA was subcloned into a
pGEM-T plasmid vector (Takara Bio, Inc.). The plasmid vector was
linearized and transcribed using the Ambion®
MAXIscript® T7 In Vitro Transcription kit
(Ambion; Thermo Fisher Scientific, Inc.). The T3 RNA polymerase was
purchased from Ambion (Thermo Fisher Scientific, Inc.).
Single-strand RNA probes were prepared using a Digoxigenin RNA
Labeling kit, according to the manufacturers instructions (Roche
Diagnostics). The template was removed using 2 U/l
ribonuclease-free DNase I (Ambion; Thermo Fisher Scientific, Inc.)
for 15 min at 37°C. The RNase inhibitor used was purchased from
Ambion (Thermo Fisher Scientific, Inc.). Digoxigenin-labeled probes
(DIG-dUTP) were frozen at −80°C. The primer sequences and
restriction sites are shown in Table
I.
 | Table I.Description of probes used for in
situ hybridization. |
Table I.
Description of probes used for in
situ hybridization.
| Probe name | Amplification
primers | Restriction
sites | Transcription
direction |
|---|
| vmhc | Forward:
CTCCTGGTGCAAAGAATC | SalI | T7 |
|
| Reverse:
TTCAGCTCAGAGTGGCATTCGTCC |
|
|
| amhc | Forward:
AAGCATTCGCTCGTGGACT | NcoI | SP6 |
|
| Reverse:
CATCCAGTGTCTGCTGGT |
|
|
| bmp4 | Forward:
TGCCAAGTCCTACTGGGAG | SacII | SP6 |
|
| Reverse:
CGTGATTGGTGGAGTTGAG |
|
|
| cmlc2 | Forward:
CTCTTCCAATGTCTTCTCC | SalI | T73 |
|
| Reverse:
TATTTCCAGCCACGTCTA |
|
|
| notch1b | Forward:
GGCCAAACATGTGAGGTG | SacII | SP6 |
|
| Reverse:
GCTGTATCTTGTGCCGCT |
|
|
| nppa | Forward:
ATGGCCGGGGGACTAATTC | SacI | T7 |
|
| Reverse:
CCGCGTATTGCAGCTAACC |
|
|
| has2 | Forward:
ACGACACTGTTCGGCATTT | ApaI | SP6 |
|
| Reverse:
CAGCGGGTTTGTTGGTTG |
|
|
| Fli1a | Forward:
GTCTTATGATGCTGTACGGAGG | NcoI | SP6 |
|
| Reverse:
CCATCTTCGAGTGCAGTTCAAG |
|
|
Observation of malformation rate,
lethality, hatch rate and zebrafish cardiac phenotypes
Following injection, the malformation rate and
lethality at 24, 48, 72 and 96 hpf, and the hatch rate at 72 hpf
were measured with an Olympus SZ61 dissecting microscope as
previously described (25,26). The cardiac phenotypes of the
embryos at 48 and 72 hpf were observed. Cardiac-specific phenotypes
were imaged with a digital camera (Olympus DP71; Olympus
Corporation). Adobe Photoshop CS5 (Adobe Systems, Inc.) was used to
process the digital images.
Measurement of heart rate
Every 20 sec, the heart rate was recorded as the
number of consecutive contractions during one diastole phase under
a light dissecting microscope (MVX10; Olympus Corporation;
magnification, ×8) every 20 sec. The method is briefly described as
follows (26): Tricaine stock
solution (Sigma-Aldrich; Merck KGaA) was prepared for anesthesia
using 97.9 ml double-distilled water, 400 mg tricaine powder and
2.1 ml of 1 mol/l Tris (pH 9). Subsequently, 400 µl tricaine (4
mg/ml) was diluted in 8 ml double-distilled water. Following
anesthesia, the embryos were transferred to a recording chamber
filled with modified Tyrodes solution. The heart rates of the
embryos were then measured at different time points as previously
described (25).
Histological staining
Zebrafish were euthanized in a mixture of ice and
water for 20 min and then placed in a refrigerator at −20°C.
Cessation of the heartbeat was used to confirm death. To observe
histological changes, zebrafish embryos were fixed with 4%
paraformaldehyde at 4°C overnight and embedded in paraffin. The
embryos were sectioned consecutively at 5-µm thickness. Embryonic
tissues were stained with hematoxylin (1 min) and eosin (5 sec) at
room temperature. Photomicrographs were taken using an Olympus DP71
digital camera (Olympus Corporation) under a polarizing light
microscope (Olympus BX51; Olympus Corporation; magnification,
×200). At least three consecutive sections were selected to ensure
that the section contained the embryonic heart structures of
atrioventricular canal (AVC), and/or leaflets of the
atrioventricular valve.
Whole mount in situ hybridization
The whole in situ hybridization was performed
using the whole in situ hybridization kit for zebrafish
embryos (Nanjing YSY Biotech Company, Ltd.) according to the
manufacturers protocol. Riboprobes were used for confirming the
sequences of amhc, vmhc, nppa, cmlc2, bmp4, notch1b, appa,
hyaluronan synthase 2 (has2), fli-1 proto-oncogene ETS
transcription factor a (fli1a) and versican a (vcana) as previously
described (22–24). The detailed procedure of the whole
mount in situ hybridization was performed as previously
described (23,25,26).
Embryos were immobilized in 4% paraformaldehyde for 12–16 h at 4°C,
dehydrated continuously by graded methanol solution and stored at
−20°C for at least 30 min. The antisense probe (1 ng/µl) of
digoxigenin-labeled RNA was hybridized overnight in 50% formamide
buffer at 65°C. The non-specific binding probes were washed in the
SSC solution (cat. no. S6639; Sigma-Aldrich; Merck KGaA). Following
washing with SSC, the embryos were immersed in blocking solution
(10% sheep serum, 70% MAB and 0.1% Tween; Roche Diagnostics) at
room temperature for 1 h. Subsequently, the embryos were incubated
in blocking solution containing 1/5,000 anti-Digoxigenin-AP Fab
fragment (Roche Diagnostics) at 4°C overnight. Hybridization was
detected using nitroblue tetrazolium
chloride/5-bromo-4-chloro-3-indolyl phosphate and anti-DIG-AP Fab
fragment staining. The experimental and control embryos were
treated concurrently. The results of in situ hybridization
were observed under a light dissecting microscope (Olympus
Corporation; magnification, ×8) and recorded using the RetigaExit
Fast 1394 CCD camera (Olympus Corporation). The images were
processed with Adobe Photoshop CS5 Extended software (Adobe
Systems, Inc.) for contrast and light intensity processing.
Reverse transcription-quantitative PCR
(RT-qPCR)
The relative expression levels of Wnt4 and β-catenin
were examined by RT-qPCR and normalized to the reference gene
β-actin. Total RNA was extracted from at least 60 embryos for each
measurement using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). cDNA was synthesized from 500 ng RNA
using a reverse transcriptase kit (Vazyme Biotech Co., Ltd.) with
the following temperature protocol: 25°C for 5 min, followed by
50°C for 15 min and 85°C for 5 min. qPCR was performed using
SYBR-Green (Vazyme Biotech Co., Ltd.) on an ABI 7500 detection
system (Applied Biosystems; Thermo Fisher Scientific, Inc.)
according to the manufacturers protocol. The following primer pairs
were used for qPCR: Wnt4 forward, 5′-TGTGCAAACGGAACCTTGAG-3′ and
reverse, 5′-ATGCCCTTGTCACTGCAAAG-3′; β-catenin forward,
5′-AGACAGCTCGTTGTACTGCT-3′ and reverse, 5′-GTGTCGTGATGGCGTAGAA-3′;
and β-actin forward, 5′-CAACAGAGAGAAGATGACACAGATCA-3′ and reverse,
5′-GTCACACCATCACCAGAGTCCATCAC-3′. The following thermocycling
conditions were used for qPCR: Initial denaturation (95°C, 10 sec),
followed by 40 cycles of amplification and quantification (94°C, 5
sec and 72°C, 30 sec) with a final extension (7 min, 72°C).
Relative gene expression levels were quantified based on the cycle
quantification (Cq) values and normalized to the reference gene
β-actin. Gene expression levels were calculated using the
2−∆∆Cq method (27).
Stem-loop RT-qPCR
Stem-loop RT-PCR was performed to examine the
relative expression levels of miR-29c following microinjection.
miRNA levels were measured using the BulgeLoop™ miRNA qPCR Primer
Set (Vazyme Biotech Co., Ltd.) according to the manufacturers
instructions. The following primer pairs were used for PCR: U6
forward, 5′-TTGGTCTGATCTGGCACATATAC-3′ and reverse,
5′-AAAAATATGGAGCGCTTCACG-3′ and miR-29c forward,
5′-GCCTAGCACCATTTGAAATCG-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′.
Relative miR-29c expression levels were quantified using the
2−∆∆Cq method and normalized to the reference gene U6.
Stem-loop RT-qPCR was performed according to the aforementioned
RT-qPCR protocol.
Statistical analysis
Quantitative data from three independent experiments
were expressed as the mean ± SD. Statistical analyses were
performed using SPSS 24.0 software (IBM Corp.). Categorical data
were tested using χ2 and Fishers exact tests. For
comparison of multiple rates, the Scheffe post hoc test was
performed following the chi-squared test. Measurement data were
analyzed with one-way ANOVA followed by Student-Newman-Keuls post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effects of miR-29c overexpression on
mortality and hatchability
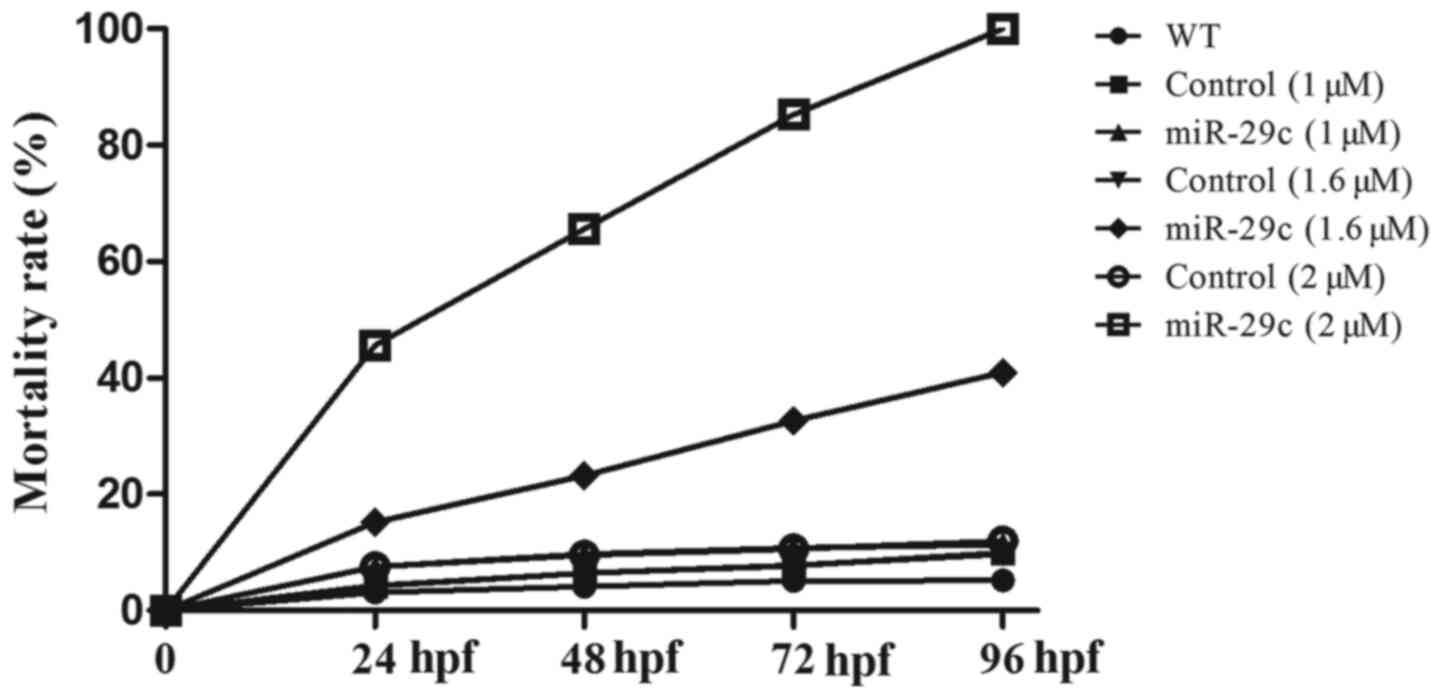
The effects of miR-29c mimics (1, 1.6 and 2 µM) on
the mortality and hatch rate at different stages of embryonic
development (24, 48, 72 and 96 hpf) were assessed. The lethal rate
of miR-29c mimic-injected embryos ranged between 18.52–100%
(Table II), which was
significantly higher (1.6 and 2 µM miR-29c mimic concentrations)
compared with WT and NC embryos. Compared with the WT and NC
groups, the hatchability of miR-29c mimic-injected embryos (1.6 and
2 µM miR-29c mimic concentrations) was significantly reduced
(Table III). The data indicated
that with the increase of miR-29 mimic concentration, the mortality
increased, and hatchability decreased, showing a dose-dependent
trend (Fig. 1). The results
suggested that miRNA-29c overexpression may hinder development and
increase the mortality of zebrafish embryos.
| Table II.Mortality rates (%) of zebrafish
embryos. |
Table II.
Mortality rates (%) of zebrafish
embryos.
|
| Time, hpf |
|---|
|
|
|
|---|
| Group | 24 | 48 | 72 | 96 |
|---|
| WT | 7.11 | 13.12 | 14.26 | 14.59 |
| Control (1 µM) | 7.18 | 13.42 | 14.67 | 14.93 |
| miR-29c (1 µM) | 18.52 | 26.74 | 28.81 | 31.36 |
| Control (1.6
µM) | 8.26 | 16.42 | 16.76 | 16.69 |
| miR-29c (1.6
µM) | 21.19a | 25.21a | 32.62a | 39.95a |
| Control (2 µM) | 9.51 | 15.52 | 17.66 | 17.99 |
| miR-29c (2 µM) | 40.61a | 58.62a | 89.26a | 100.00a |
| Table III.Hatch rate of zebrafish embryos at 72
h post-fertilization. |
Table III.
Hatch rate of zebrafish embryos at 72
h post-fertilization.
| Group | n | Hatch number | Hatch rate, % |
|---|
| WT | 87 | 85 | 97.70 |
| Control (1 µM) | 87 | 85 | 97.70 |
| miR-29c (1 µM) | 68 | 64 | 94.11 |
| Control (1.6
µM) | 85 | 83 | 97.65 |
| miR-29c (1.6
µM) | 68 | 58 | 85.29a |
| Control (2 µM) | 84 | 81 | 96.42 |
| miR-29c (2 µM) | 38 | 10 | 26.32a |
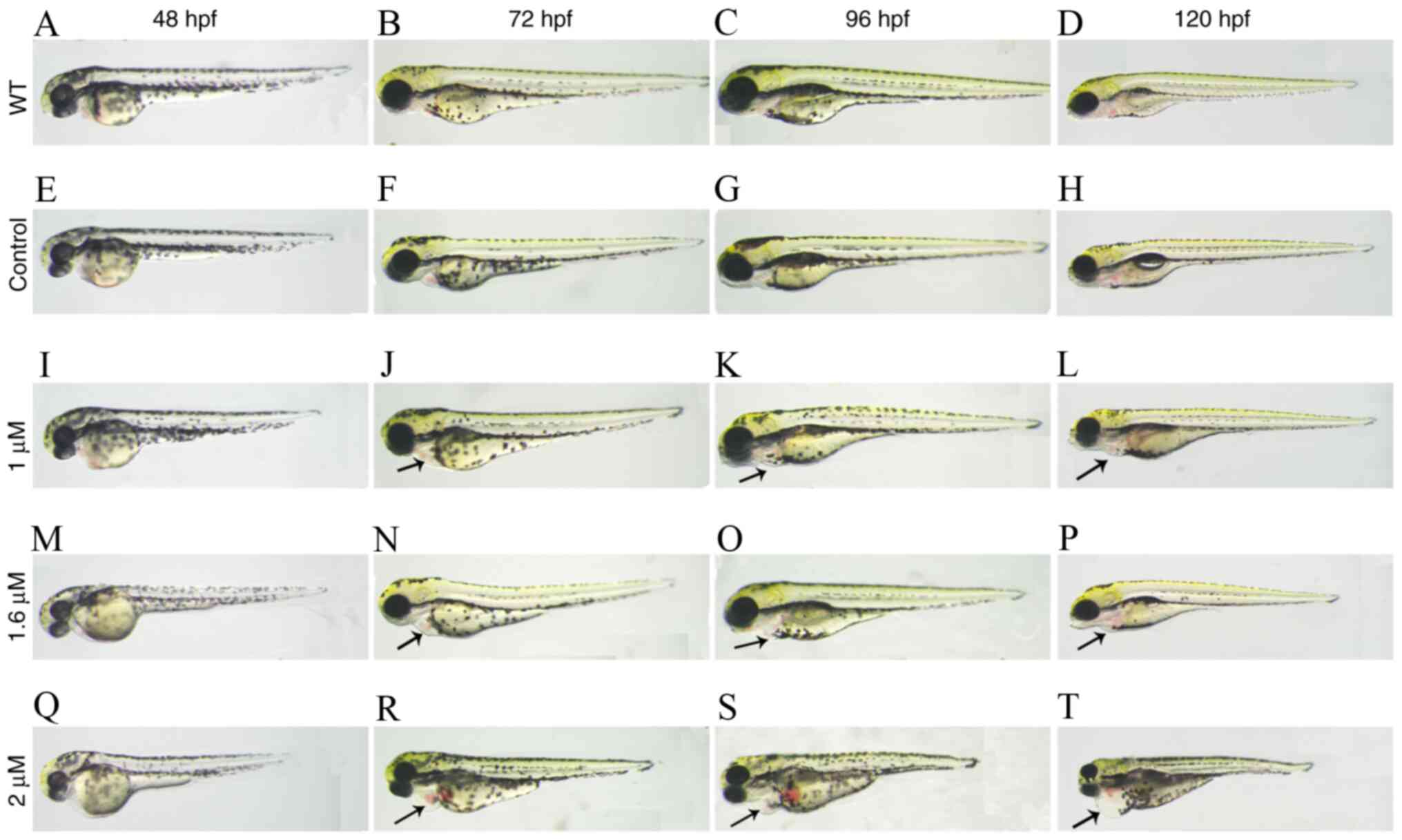
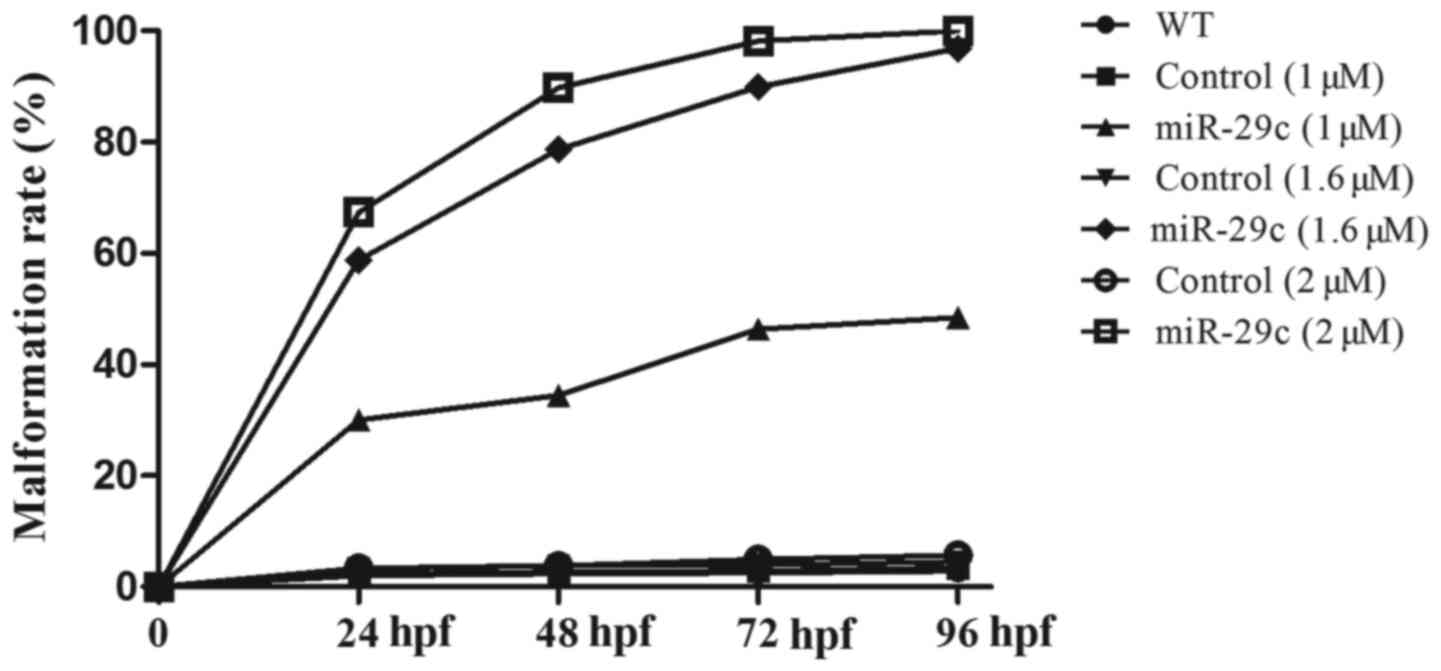
Effects of miR-29c overexpression on
the morphology of zebrafish embryos
The malformation rate is the proportion of zebrafish
embryos and larvae with obvious morphological abnormalities, such
as pericardial edema and shrinkage. The effect of increasing
concentrations of miR-29c mimics on the malformation rate was
evaluated at different stages (24, 48, 72 and 96 hpf) (Fig. 2). Quantitative analysis indicated
that the malformation rate of zebrafish embryos injected with
miR-29c mimics increased in a dose-dependent manner (Fig. 3). In particular, the malformation
rate of miR-29c mimic-injected embryos ranged between 30.08–100%,
which was significantly higher compared with NC and WT groups
(Table IV). The phenotype was
observed after injection of 1 µM and 1.6 µM miR-29c mimic.
Following injection of 2 µM miR-29c mimic, no embryos survived to
96 hpf. Therefore, 1.6 µM miR-29c mimic was chosen for further
experiments.
| Table IV.Malformation rate (%) of zebrafish
embryos at different time points. |
Table IV.
Malformation rate (%) of zebrafish
embryos at different time points.
|
| Time, hpf |
|---|
|
|
|
|---|
| Group | 24 | 48 | 72 | 96 |
|---|
| WT | 2.08 | 2.35 | 2.52 | 2.86 |
| Control (1 µM) | 2.36 | 2.72 | 2.86 | 3.46 |
| miR-29c (1 µM) | 30.08a | 34.44a | 46.40a | 48.45a |
| Control (1.6
µM) | 3.25 | 3.89 | 3.92 | 4.56 |
| miR-29c (1.6
µM) | 58.75a | 78.69a | 89.92a | 96.89a |
| Control (2 µM) | 3.35 | 3.78 | 4.92 | 5.69 |
| miR-29c (2 µM) | 67.38a | 89.75a | 98.20a | 100.00a |
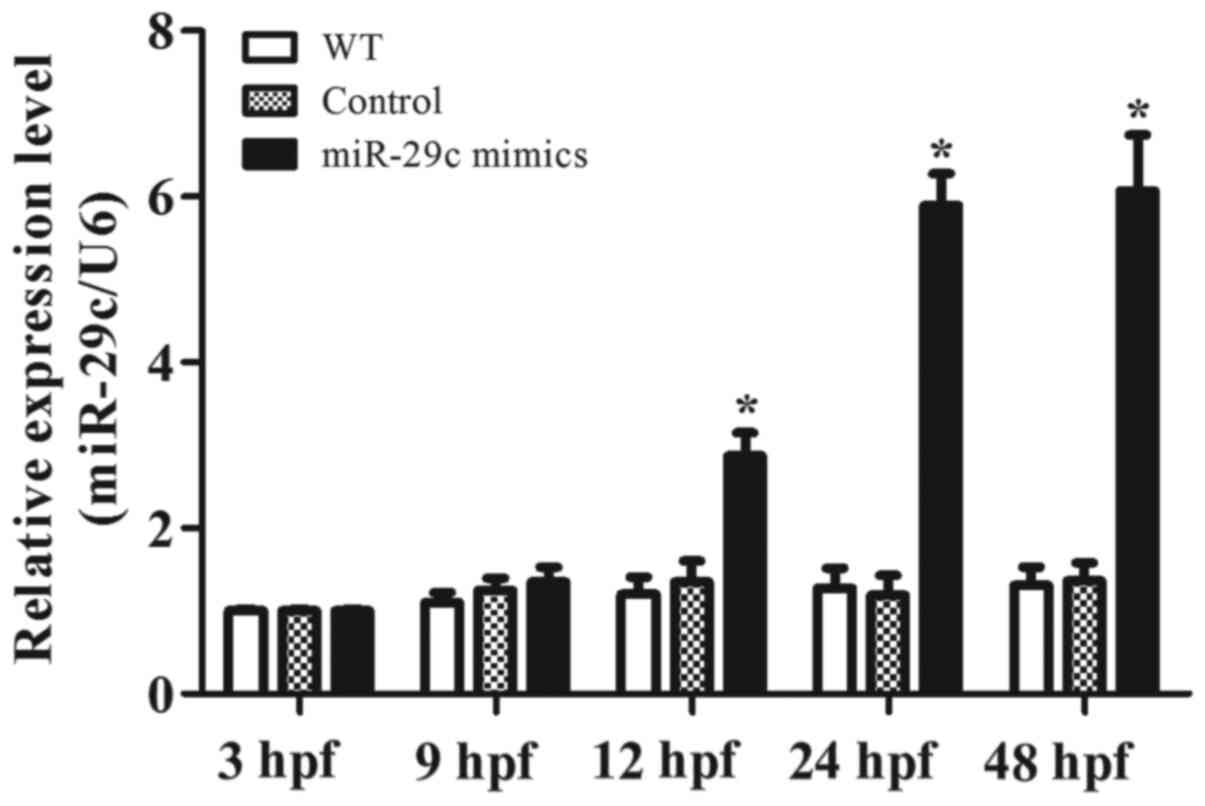
Effects of miR-29c overexpression on
heart rate and cardiac function
A dose of 1.6 µM miR-29c mimic or 1.6 µM NC was
chosen to explore the effects of miR-29c overexpression on the
average heart rate (AHR) and cardiac function. RT-qPCR was used to
confirm miR-29c overexpression (Fig.
4). At ~16 hpf, the cardiac progenitor cells of zebrafish begin
to differentiate; cardiac precursor cells reach the midline and
bind to each other and the cardiac cone extends. At ~24 hpf, the
cardiac cone gradually grows into a linear cardiac tube, then curls
to the left to form the anatomical atrium, ventricle and
atrioventricular valve. The heart development of zebrafish embryo
is completed within 48 hpf (28).
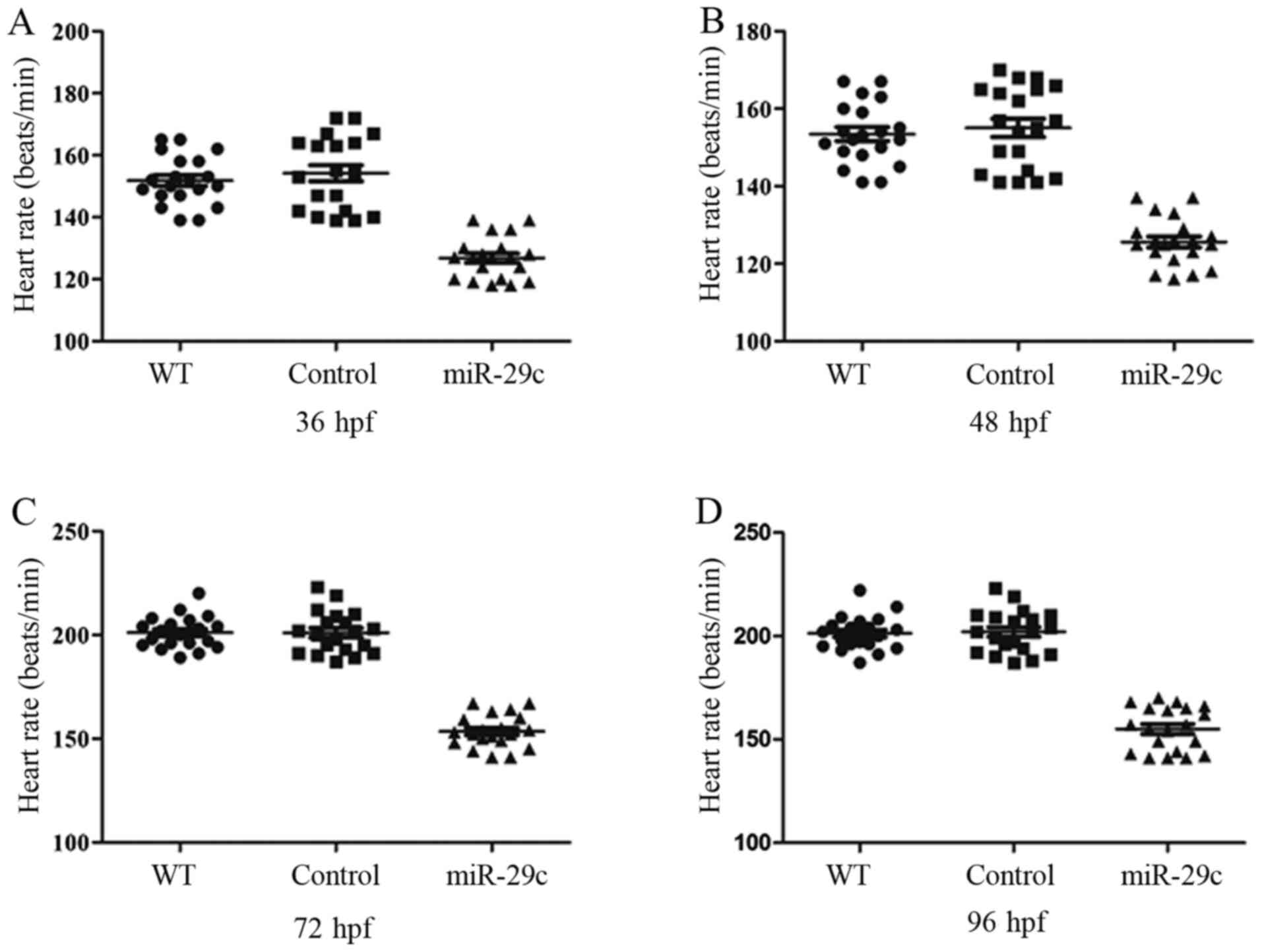
Therefore, the heart rate at 36, 48, 72 and 96 hpf were next
measured to assess the effect of miR-29c overexpression on heart
contraction. At 36 hpf, the AHR was 148.8±6.6 beats per minute
(bpm) in the WT group, 147.3±7.9 bpm in the NC group and 126.8±4.9
bpm in 1.6 µM miR-29c mimic injection group. At 48 hpf, these AHR
changed to 158.6±7.6 bpm, 154.7±5.8 bpm and 125.6±3.8 bpm. At 72
hpf, the AHRs were 202.5±8.4 bpm, 202.6±9.4 bpm and 164153.5±7.6
bpm. At 96 hpf, the AHRs increased to 206.2±9.5 bpm, 204.7±8.2 bpm
and 155.5±9.8 bpm, respectively (n=20; Fig. 5). The aforementioned data indicated
that the AHR significantly reduced following miR-29c mimic
injection. The atria and ventricle of control embryos exhibited
rhythmic and robust contraction, which ensured circulation in the
whole body. By contrast, weak and irregular contractions were more
prominent in the miR-29c mimic injection group. Taken together, the
results suggested that miR-29c overexpression could significantly
alter cardiac systolic function.
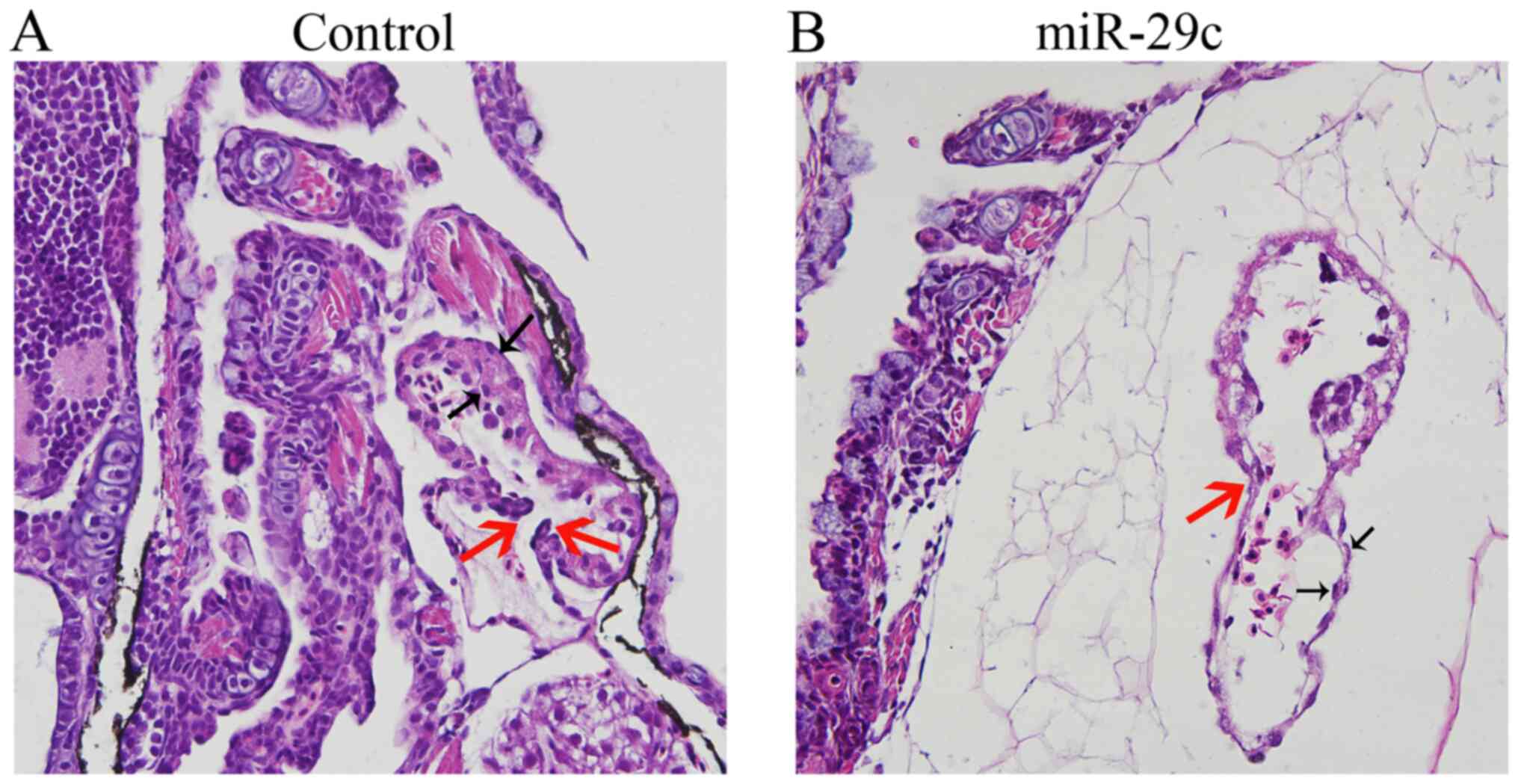
Effects of miR-29c overexpression on
the histological structure of zebrafish embryonic heart
Following injection of 1.6 µM NC or 1.6 µM miR-29c
mimics into zebrafish fertilized eggs, seven typical embryos were
sectioned at 120 hpf. The results showed that the myocardial layer,
endocardium and inter atrioventricular valvular lobule of the
embryonic heart in the control group developed normally (Fig. 6A). In the 1.6 µM miR-29c mimic
injection group, however, pericardial edema presented, as exhibited
by cavities outside the heart. The ventricular layer became thinner
and the number of endocardial cells decreased (Fig. 6B). The results showed that miR-29c
overexpression notably affected zebrafish embryonic
cardiogenesis.
Effects of miR-29c overexpression on
cardiac cyclization and expression of myocardial and endocardial
molecular markers
Under normal conditions, cmlc2 is expressed in the
whole heart, vmhc is expressed in the ventricle, amhc is expressed
in the atrium, and fli1a is expressed in the endocardium in the
embryo heart of zebrafish (22).
To study the effects of miR-29c on ventricular and atrioventricular
tube development at the molecular level, in situ
hybridization was performed to detect the expression of amhc,
cmlc2, fli1a and vmhc at 48 hpf. Compared with the control group
(embryos injected with 2 μM negative control mimics), embryos
injected with 1.6 and 2 µM miR-29c mimics displayed abnormal
expression patterns of the gene cmlc2, which marks cardiac
cyclization, and loss of left-right asymmetry (Fig. 7A), suggesting that miR-29c
overexpression altered embryonic cardiac cyclization. The genes
amhc (Fig. 7B) and vmhc (Fig. 7C), which mark atrio-genesis, were
also abnormally expressed in embryos overexpressing miR-29c, and
the atrial expression regions were significantly reduced,
suggesting that miR-29c overexpression affected atrial and
ventricular genesis in embryonic hearts. Similarly, the expression
of fli1a, a marker gene of cardiac endocardio-genesis, was notably
downregulated (Fig. 7D) compared
with control group, suggesting that miR-29c overexpression affected
embryonic cardiac endocardio-genesis.
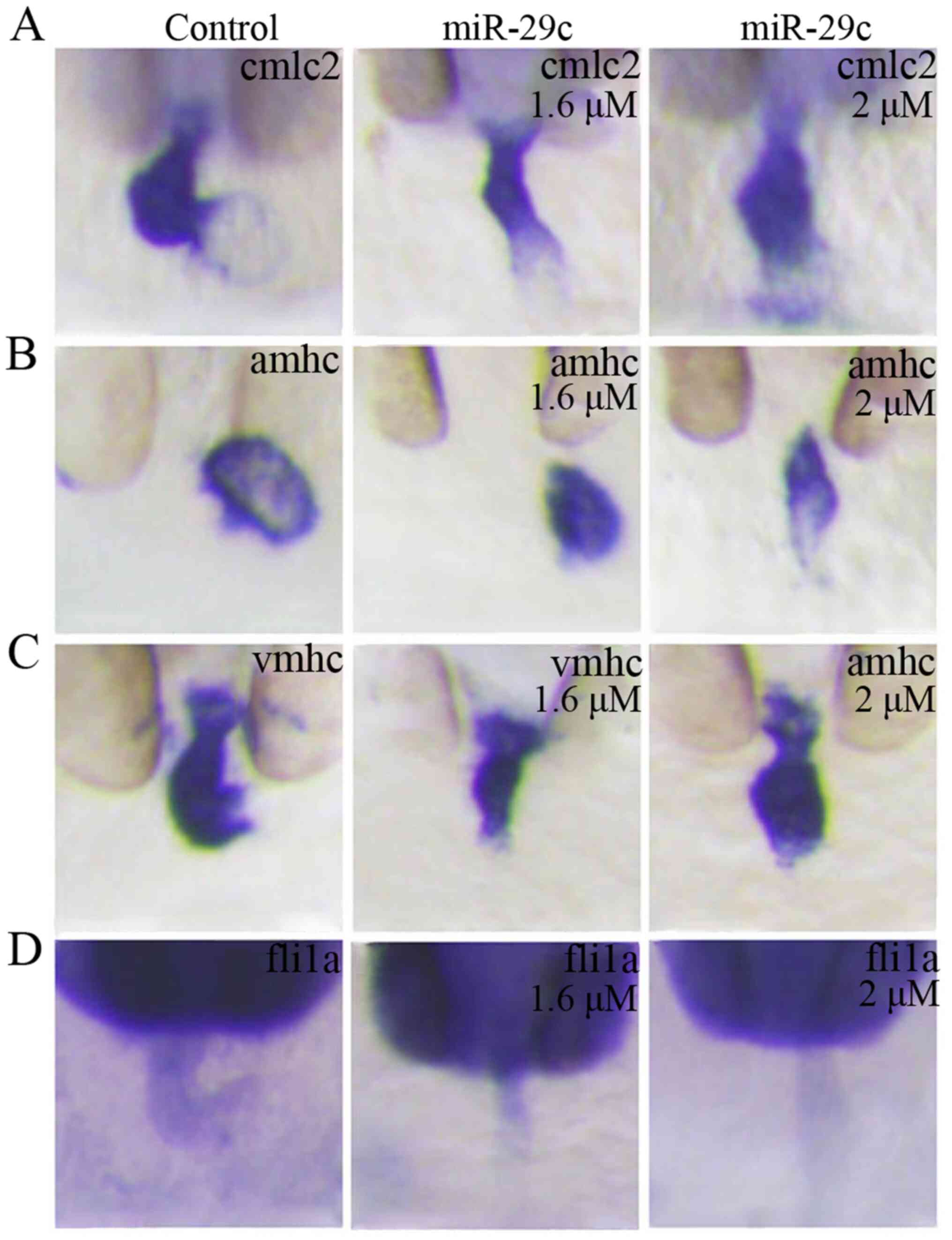 | Figure 7.Overexpression of miR-29c affects the
cardiac cyclization and the expression of myocardial and
endocardial molecular markers in the embryonic hearts of zebrafish.
At 48 hpf following injection, abnormal expression of (A) cmlc2,
(B) amhc, (C) vmhc and (D) fli1a was observed in the miR-29c mimic
injection group (1.6 and 2 µM miR-29c mimic concentrations)
compared with the control group (magnification, ×8). miR, microRNA;
hpf, hours post-fertilization; cmlc2, cardiac myosin light chain-2;
amhc, atrial myosin heavy chain; vmhc, ventricular myosin heavy
chain; fli1a, fli-1 proto-oncogene ETS transcription factor a; NC,
negative control. |
Effects of miR-29c overexpression on
the precursor cells of the atrioventricular valve in zebrafish
embryos
The embryos were fixed for in situ
hybridization at 48 hpf. The results showed that compared with the
control group (Fig. 8A-E), the
expression of bmp4 (Fig. 8F), has2
(Fig. 8G) and notch1b (Fig. 8H) in the atrioventricular pathway
diminished at 48 hpf in the miR-29c mimics injection group. In
miR-29c overexpression embryos, nppa (Fig. 8I) was heterotopically expressed in
the atrioventricular pathway, and highly expressed in the atrium,
whereas vcana (Fig. 8J) was not
expressed in the atrioventricular pathway, but highly expressed
near the ventricular outflow tract. These data suggested that
miR-29c overexpression affected the formation of precursor cells of
the atrioventricular valves in embryonic hearts.
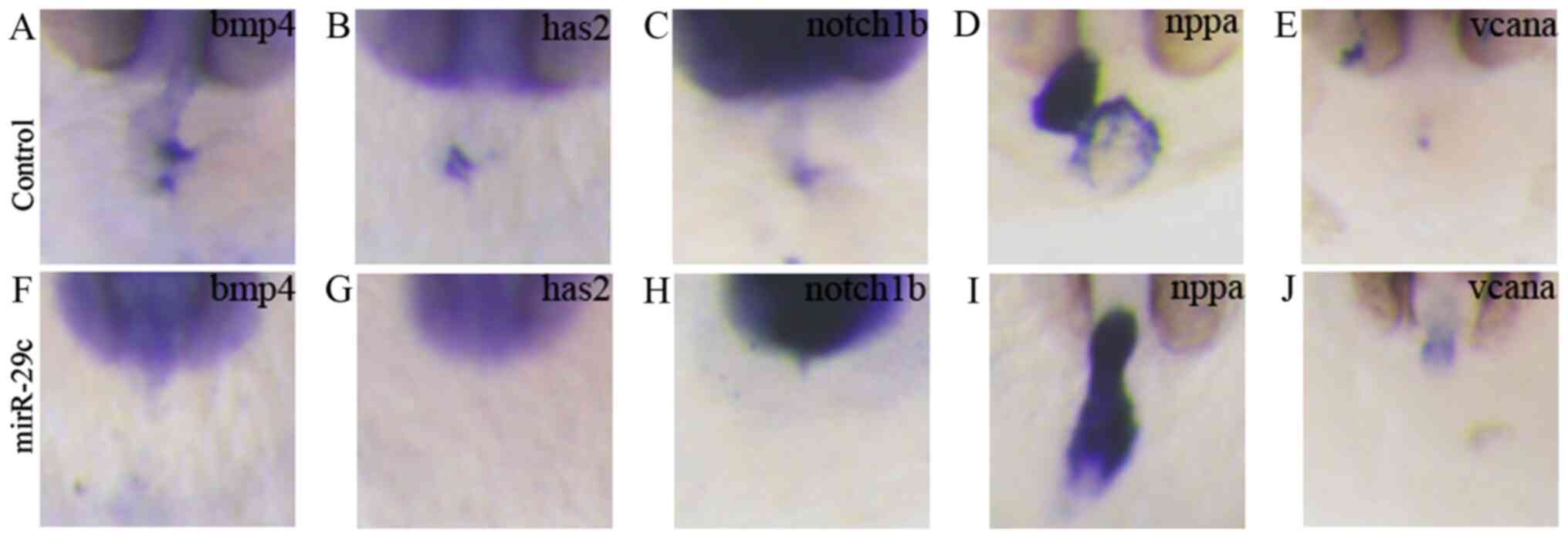 | Figure 8.miR-29c overexpression affects the
expression of marker genes in zebrafish embryonic heart valves. 1.6
µM miR-29c mimics or 1.6 µM NC were injected into fertilized eggs
for 48 hpf, and the zebrafish embryos were hybridized in
situ. Control embryos were injected with 1.6 µM NC and the
expression patterns of cardiac valve precursor marker genes (A)
bmp4, (B) has2, (C) notch1b, (D) nppa and (E) vcana were measured.
Embryos were injected with 1.6 µM miR-29c mimics and the expression
patterns of cardiac valve precursor marker genes (F) bmp4, (G)
has2, (H) notch1b, (I) nppa and (J) vcana were measured
(magnification, ×8). miR, microRNA; hpf, hours post-fertilization;
NC, negative control; bmp4, bone morphogenetic protein 4; has2,
hyaluronan synthase 2; notch1b, notch homolog 1b; nppa, natriuretic
peptide precursor A; vcana, versican a; AVC, atrioventricular
canal. |
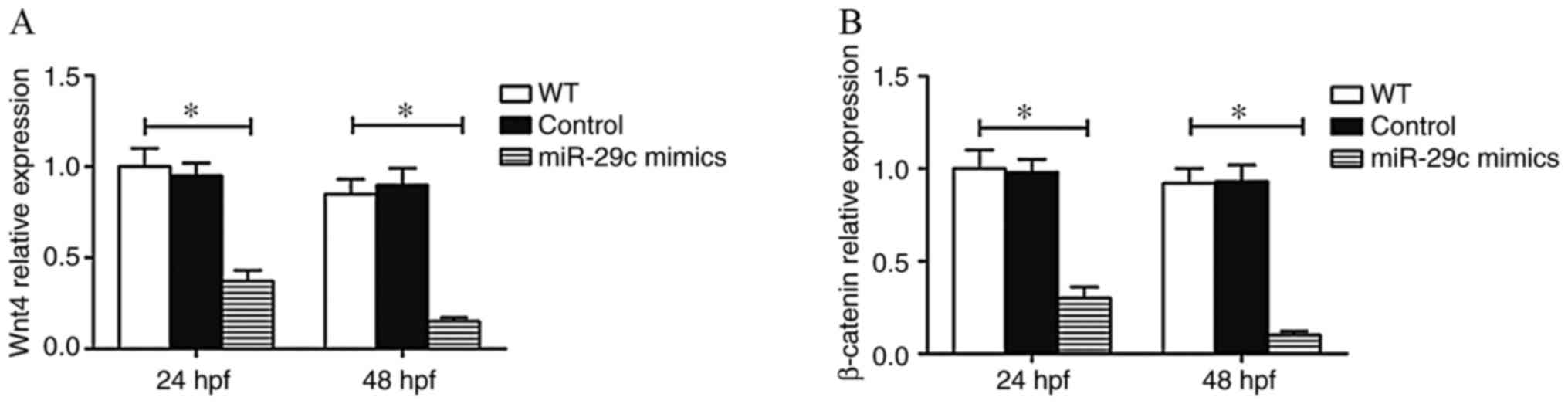
Effect of miR-29c overexpression on
the Wnt4 signaling pathway
The Wnt signaling pathway plays an important role in
embryonic development (29). Wnt
signaling has been demonstrated to play a pivotal role in cardiac
asymmetry and Kupffers vesicle development in zebrafish (30). Additionally, our previous studies
(17,18) found that Wnt4 was the target gene
of miR-29c. The present study found that the mRNA expression of
Wnt4 and β-catenin decreased in embryos injected with miR-29c
mimics (Fig. 9). This suggested
that miR-29c overexpression led to abnormal cardiac development in
zebrafish by modulating the Wnt4 signaling pathway.
Discussion
As the most common birth defect worldwide, CHD is
caused by genetic factors in ~98% cases (31,32).
Therefore, exploring the genetic factors leading to CHD is helpful
to elucidate CHD etiology. As hypothesized, the present study
confirmed the role of miR-29c.
Zebrafish and vertebrates share physiological and
morphological commonness in terms of cardiovascular development
(33,34). Moreover, unlike that in
vertebrates, severe heart malformations do not immediately lead to
zebrafish death. Even in the total absence of blood circulation,
zebrafish receive enough oxygen by passive diffusion to survive and
continue to develop in a relatively normal fashion for several days
(28,35). Secondly, zebrafish embryos are
transparent, making it convenient to perform microinjections and
observe morphological cardiac changes (28). Additionally, early cardiac
development in zebrafish embryo is similar to that in humans, such
as the migration of precursor cells to the central line, the
formation of cardiac tubes and the formation and circulation of
early ventricles (35,36). Zebrafish embryos can be used to
model the characteristics and potential molecular mechanisms of
cardiac development defects. The present study established a
zebrafish model to observe the effects of miR-29c overexpression on
embryonic heart development. The data showed that miR-29c
overexpression increased embryo mortality and decreased embryo
hatchability. In addition, miR-29c overexpression led to left-right
asymmetry defects in zebrafish embryos and affected heart
development in a dose-dependent manner, manifested by heart rate
slowdown, pericardial edema and heart looping disorder during the
early stage of development.
The potential molecular mechanisms were further
explored. Numerous signaling pathways, such as Wnt, bone
morphogenetic protein and retinoic acid, co-regulate cardiac
development (37,38). Classical and non-classical Wnt
signaling participate in cardiac differentiation in a coordinated
manner; mutations in Wnt genes can lead to CHD in several animal
models (39). As a member of the
Wnt family, Wnt4 plays an important role in regulating cell
proliferation, differentiation and apoptosis (29). It has been reported that the Wnt4
signaling (Wnt4/β-catenin) pathway is essential for cardiac
left-right patterning (30). Using
bioinformatics analysis and luciferase reporter assays, our
previous studies verified that Wnt4 was the target gene of miR-29c
(17,18). To investigate whether miR-29c
regulates cardiac function and phenotype via the Wnt4/β-catenin
signaling pathway, the present study detected mRNA expression
levels of Wnt4 and β-catenin following miR-29c mimic injection. The
expression of Wnt4/β-catenin was downregulated in embryos injected
with miR-29c mimics, suggesting that miR-29c may rely on the
Wnt4/β-catenin signaling pathway to regulate the development of the
zebrafish embryonic heart.
In conclusion, miR-29c regulated zebrafish embryonic
heart development via the Wnt4/β-catenin signaling pathway.
Inhibiting miR-29c expression may have important practical
significance for the prevention, diagnosis and treatment of
CHD.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81300127
and 81600223), the Six Talent Peaks Project in Jiangsu Province
(grant no. 2017-WSN-287), the Jiangsu Provincial Medical Youth
Talent (grant nos. QNRC2016511 and QNRC2016512), the Taizhou
Municipal Science and Technology Bureau (grant no. TS201729) and
the Taizhou People's Hospital Scientific Research Project (grant
no. ZL201622).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
YS and GS designed the study and drafted the
manuscript. YS, HL and RC performed the experiments. LZ and GS
performed the statistical analysis. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of Jiangsu Taizhou People's Hospital (Taizhou
Clinical Medical School of Nanjing Medical University).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Liu N and Olson EN: MicroRNA regulatory
networks in cardiovascular development. Dev Cell. 18:510–525. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wilczynski B and Furlong EE: Challenges
for modeling global gene regulatory networks during development:
Insights from Drosophila. Dev Biol. 340:161–169. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lints TJ, Parsons LM, Hartley L, Lyons I
and Harvey RP: Nkx-2.5: A novel murine homeobox gene expressed in
early heart progenitor cells and their myogenic descendants.
Development. 119:419–431. 1993.PubMed/NCBI
|
|
4
|
Bruneau BG, Nemer G, Schmitt JP, Charron
F, Robitaille L, Caron S, Conner DA, Gessler M, Nemer M, Seidman
CE, et al: A murine model of Holt-Oram syndrome defines roles of
the T-box transcription factor Tbx5 in cardiogenesis and disease.
Cell. 106:709–721. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Watt AJ, Battle MA, Li J and Duncan SA:
GATA4 is essential for formation of the proepicardium and regulates
cardiogenesis. Proc Natl Acad Sci USA. 101:12573–12578. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xiang R, Lei H, Chen M, Li Q, Sun H, Ai J,
Chen T, Wang H, Fang Y and Zhou Q: The miR-17-92 cluster regulates
FOG-2 expression and inhibits proliferation of mouse embryonic
cardiomyocytes. Braz J Med Biol Res. 45:131–138. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sluijter JP, van Mil A, van Vliet P, Metz
CH, Liu J, Doevendans PA and Goumans MJ: MicroRNA-1 and −499
regulate differentiation and proliferation in human-derived
cardiomyocyte progenitor cells. Arterioscler Thromb Vasc Biol.
30:859–868. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li Y and Kowdley KV: MicroRNAs in common
human diseases. Genomics Proteomics Bioinformatics. 10:246–253.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen J and Wang DZ: microRNAs in
cardiovascular development. J Mol Cell Cardiol. 52:949–957. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao Y, Ransom JF, Li A, Vedantham V, von
Drehle M, Muth AN, Tsuchihashi T, McManus MT, Schwartz RJ and
Srivastava D: Dysregulation of cardiogenesis, cardiac conduction,
and cell cycle in mice lacking miRNA-1-2. Cell. 129:303–317. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao Y, Samal E and Srivastava D: Serum
response factor regulates a muscle-specific microRNA that targets
Hand2 during cardiogenesis. Nature. 436:214–220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu N, Bezprozvannaya S, Williams AH, Qi
X, Richardson JA, Bassel-Duby R and Olson EN: microRNA-133a
regulates cardiomyocyte proliferation and suppresses smooth muscle
gene expression in the heart. Genes Dev. 22:3242–3254. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meder B, Katus HA and Rottbauer W: Right
into the heart of microRNA-133a. Genes Dev. 22:3227–3231. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu S, Cao L, Zhu J, Kong L, Jin J, Qian
L, Zhu C, Hu X, Li M, Guo X, Han S, et al: Identification of
maternal serum microRNAs as novel non-invasive biomarkers for
prenatal detection of fetal congenital heart defects. Clin Chim
Acta. 424:66–72. 2013.doi: 10.1016/j.cca.2013.05.010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu M, Chen Y, Song G, Chen B, Wang L, Li
X, Kong X, Shen Y and Qian L: MicroRNA-29c overexpression inhibits
proliferation and promotes apoptosis and differentiation in P19
embryonal carcinoma cells. Gene. 576:304–311. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen B, Song G, Liu M, Qian L, Wang L, Gu
H and Shen Y: Inhibition of miR-29c promotes proliferation, and
inhibits apoptosis and differentiation in P19 embryonic carcinoma
cells. Mol Med Rep. 13:2527–2535. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
van Almen GC, Verhesen W, van Leeuwen RE,
van de Vrie M, Eurlings C, Schellings MW, Swinnen M, Cleutjens JP,
van Zandvoort MA, Heymans S, et al: MicroRNA-18 and microRNA-19
regulate CTGF and TSP-1 expression in age-related heart failure.
Aging Cell. 10:769–779. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bakkers J: Zebrafish as a model to study
cardiac development and human cardiac disease. Cardiovasc Res.
91:279–288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Westerfield M: The Zebrafish Book. A Guide
for The Laboratory Use of Zebrafish Danio (Brachydanio) rerio. 4th
Edition. University of Oregon Press; Eugene, OR, USA: 1993
|
|
22
|
Kimmel CB, Ballard WW, Kimmel SR, Ullmann
B and Schilling TF: Stages of embryonic development of the
zebrafish. Dev Dyn. 203:253–310. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Thisse C and Thisse B: High-resolution in
situ hybridization to whole-mount zebrafish embryos. Nat Protoc.
3:59–69. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen JN and Fishman MC: Zebrafish tinman
homolog demarcates the heart field and initiates myocardial
differentiation. Development. 122:3809–3816. 1996.PubMed/NCBI
|
|
25
|
Li M, Hu X, Zhu J, Zhu C, Zhu S, Liu X, Xu
J, Han S and Yu Z: Overexpression of miR-19b impairs cardiac
development in zebrafish by targeting ctnnb1. Cell Physiol Biochem.
33:1988–2002. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang X, Zhou L, Jin J, Yang Y, Song G,
Shen Y, Liu H, Liu M, Shi C and Qian L: Knockdown of FABP3 impairs
cardiac development in Zebrafish through the retinoic acid
signaling pathway. Int J Mol Sci. 14:13826–13841. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fishman MC and Chien KR: Fashioning the
vertebrate heart: Earliest embryonic decisions. Development.
124:2099–2117. 1997.PubMed/NCBI
|
|
29
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: Components, mechanisms, and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cohen ED, Tian Y and Morrisey EE: Wnt
signaling: An essential regulator of cardiovascular
differentiation, morphogenesis and progenitor self-renewal.
Development. 135:789–798. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Blue GM, Kirk EP, Sholler GF, Harvey RP
and Winlaw DS: Congenital heart disease: Current knowledge about
causes and inheritance. Med J Aust. 197:155–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vecoli C, Pulignani S, Foffa I and
Andreassi MG: Congenital heart disease: The crossroads of genetics,
epigenetics and environment. Curr Genomics. 15:390–399. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dolk H, Loane M and Garne E; European
Surveillance of Congenital Anomalies (EUROCAT) Working Group, :
Congenital heart defects in Europe: Prevalence and perinatal
mortality, 2000 to 2005. Circulation. 123:841–849. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lin X and Xu X: Distinct functions of
Wnt/beta-catenin signaling in KV development and cardiac asymmetry.
Development. 136:207–217. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Teraoka H, Dong W and Hiraga T: Zebrafish
as a novel experimental model for developmental toxicology.
Congenit Anom (Kyoto). 43:123–132. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hu N, Sedmera D, Yost HJ and Clark EB:
Structure and function of the developing zebrafish heart. Anat Rec.
260:148–157. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Quaife NM, Watson O and Chico TJ:
Zebrafish: An emerging model of vascular development and
remodelling. Curr Opin Pharmacol. 12:608–614. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rochais F, Mesbah K and Kelly RG:
Signaling pathways controlling second heart field development. Circ
Res. 104:933–942. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Henderson DJ, Phillips HM and Chaudhry B:
Vang-like 2 and noncanonical Wnt signaling in outflow tract
development. Trends Cardiovasc Med. 16:38–45. 2006. View Article : Google Scholar : PubMed/NCBI
|