Introduction
Osteosarcoma is the most common primary malignant
bone tumor of mesenchymal origin that occurs primarily in children
and adolescents (1). It is
characterized by the production of calcified osteoid extracellular
matrix and a high frequency of inducing lung metastases, which
directly results in a high mortality rate (2). The 5-year survival rate of patients
with local osteosarcoma ranges between 65 and 75%, whereas that of
patients with metastatic osteosarcoma is <30% (3). Surgical resection, chemotherapy and
radical resection are established strategies for treating patients
with osteosarcoma (4,5). Immunotherapy or a combination of
radiotherapy and immunotherapy using nanomaterial delivery systems
have also been a research hotspot in the field (6–8).
However, the 5-year survival rate of patients with osteosarcoma
demonstrates little improvement after treatment with chemotherapy
drugs and traditional amputation (4). Therefore, it is important to identify
new effective agents for treating patients with osteosarcoma.
Certain malignant tumors present an anti-apoptotic
phenotype, which allows cancer cells to survive in local tissues
(9). Most of the current
anticancer medicines target apoptotic signaling pathways (10). Activation of caspases can be
initiated via both the extrinsic and the mitochondria-mediated
apoptotic signaling pathways (10–12).
The extrinsic pathway is triggered by activation of caspase-8 at
the plasma membrane upon ligation of the death receptor, and
subsequent cleavage of downstream effector caspases, such as
caspase-3 (10). The
mitochondria-mediated signaling pathway relies on interactions
among Bcl-2 family proteins at the mitochondrial outer membrane and
the release of cytochrome c (11). Among Bcl-2 family proteins, the
activators directly bind both anti-apoptotic proteins and
pro-apoptotic effector proteins, while sensitizers such as Bad bind
only anti-apoptotic proteins (11,12).
By competing for the BH3 binding site, sensitizers displace the
binding of activators to anti-apoptotic proteins, including Bcl-2
and Bcl-xL (11). By interacting
with the activators, pro-apoptotic effector proteins such as Bax
and Bak create openings in the outer mitochondrial membrane and
release cytochrome c, which promotes caspase-9 activation,
followed by a cascade of apoptotic proteases, including caspase-3,
resulting in cell death (12). The
increased permeability of the mitochondrial outer membrane also
results in mitochondrial depolarization with reduced mitochondrial
membrane potential (MMP) and the release of apoptotic factors,
triggering the downstream cell death processes (13–15).
Venetoclax, a direct small molecular inhibitor of Bcl-2, was
initially approved by the Food and Drug Administration (FDA) in
April 2016 and exhibited impressive outcomes particularly in
chronic lymphocytic leukemia, small lymphocytic lymphoma and acute
myeloid leukemia (16).
STAT3-mediated signaling pathways have an oncogenic
potential in cancer cell proliferation and metastasis (17). The expression levels of STAT3 have
been reported to be significantly higher in osteosarcoma tissue
compared with those in normal bone or cartilage tissue, and high
levels of STAT3 are associated with poor tumor differentiation and
metastasis (18). Five-year
overall survival and relapse-free survival rates in patients with
high STAT3 expression levels are lower compared with in those with
low STAT3 expression levels (18).
Inhibition of STAT3 has also been reported to suppress osteosarcoma
cell growth and induce apoptosis (19). Therefore, STAT3 may have an
important therapeutic and prognostic value in osteosarcoma. In
addition, STAT3 is directly involved in transcriptional regulation
of osteopontin (OPN) (20), and
increased OPN expression levels serve a critical role in
osteosarcoma, which is a potential biomarker and promising drug
target for osteosarcoma (21).
As a natural ingredient, which is primarily found in
Morus alba L., oxyresveratrol (ORES) has extensive
biological effects. Over the previous two decades, ORES has been
reported as a powerful tyrosinase activity inhibitor (22,23),
and also as having antioxidative (24,25),
anti-inflammatory (26,27), anticancer (28–30)
and anti-lipogenesis properties (31). ORES has also been observed to exert
strong neuroprotective effects, as it reduces neuronal oxidative
damage (32,33). Notably, ORES and its derivatives
have been reported to serve an efficient role against various types
of cancer, such as head and neck carcinoma (28), neuroblastoma (29), prostate (30), kidney (34) and lung cancer (35). However, it remains unknown whether
ORES has an effect on the inhibition of osteosarcoma cells and the
mechanism by which ORES inhibits tumor cell viability.
In the present study, the inhibitory effect of ORES
on Saos-2 osteosarcoma cells was determined, which indicates the
ORES is a promising agent for treating osteosarcoma.
Materials and methods
Compound and reagents
ORES (2,3′,4,5′-Tetrahydroxy-trans-stilbene,
C14H12O4; molecular weight:
244.24; purity ≥97.0%; cat. no. 29700-22-9) was purchased from
Sigma-Aldrich (Merck KGaA). DMSO was used as control. DMEM,
penicillin and streptomycin solution (100 IU/ml; 100 µg/ml), PBS,
0.25% trypsin-EDTA and enhanced chemiluminescent (ECL) substrate
were all provided by Thermo Fisher Scientific, Inc. Fetal bovine
serum (FBS) was obtained from Hangzhou Sijiqing Biological
Engineering Materials Co., Ltd. Cell Counting Kit (CCK)-8 assay
kit, MMP assay kit with JC-1, bicinchoninic acid (BCA) protein
assay kit and RIPA lysis buffer (cat. no. P0013B) were acquired
from Beyotime Institute of Biotechnology. Annexin V-FITC/propidium
iodide (PI) apoptosis detection kit was purchased from
MultiSciences (Lianke) Biotech Co., Ltd. Tris, non-fat milk and
Tween-20 were purchased from Sangon Biotech Co., Ltd. IL-6 was
purchased from PeproTech, Inc. Primary antibodies against cleaved
caspase-9 (cat. no. 20750), cleaved caspase-3 (cat. no. 9664),
GAPDH (cat. no. 5174), Bcl-2 (cat. no. 4223), Bcl-xL (cat. no.
2764), Bad (cat. no. 9239), Bax (cat. no. 5023),
phophorylated-STAT3 (P-STAT3; cat. no. 9145) and total-STAT3
(T-STAT3; cat. no. 12640) were obtained from Cell Signaling
Technologies, Inc. An antibody against OPN (cat. no. 7C5H12) was
obtained from Thermo Fisher Scientific, Inc. Horseradish peroxidase
(HRP)-conjugated anti-rabbit antibody (cat. no. 111-035-003) and
HRP-conjugated anti-mouse antibody (cat. no. 115-035-003) were
supplied by Jackson ImmunoResearch Laboratories, Inc. Methanol and
ethanol were obtained from Sinopharm Chemical Reagent Co., Ltd.
Cell culture and cell viability
assay
Saos-2 cells were obtained from the American Type
Culture Collection. Cells were grown in DMEM containing 10% FBS and
1% penicillin and streptomycin solution with 5% CO2 at
37°C. Cell passage was performed with 0.25% trypsin-EDTA.
Cell viability was detected using the CCK-8 assay
kit according to the manufacturer's instructions. Briefly, cells
were seeded in 12-well plates at a density of 4×105
cells/well. After attachment, the cells were incubated with ORES at
0, 5, 15 and 45 µM at 37°C for 48 h. The CCK-8 solution (10 µl) was
added to each well and after 1.5 h incubation, the viability of
Saos-2 cells was detected using a microplate reader at 450 nm. In
certain experiments, the Saos-2 cells were incubated with 30 µM
ORES or 30 µM ORES and 20 ng/ml IL-6 at 37°C for 24 h.
Apoptosis assay
Early and late apoptosis was detected using the
Annexin V-FITC/PI apoptosis detection kit according to the
manufacturer's instructions. Cells were seeded in 12-well plates at
a density of 4×105 cells/well. After attachment, the
cells were incubated with ORES at 0, 5, 15 and 45 µM at 37°C for 24
h. The cells were then washed twice with ice-cold PBS. After
digestion with 0.25% trypsin-EDTA, cells were collected and washed
with PBS. Cells were resuspended in 400 µl Annexin-binding buffer,
and then incubated with 5 µl Annexin V-FITC and 10 µl PI at room
temperature for 5 min. Apoptosis of Saos-2 cells was detected with
a LSRFortessa TM X-20 flow cytometer (BD Biosciences). Given that
ORES significantly inhibited Saos-2 cell viability at the
concentration between 15 and 45 µM, the Saos-2 cells was incubated
with 30 µM ORES or 30 µM ORES and 20 ng/ml IL-6 at 37°C for 24 h.
FlowJo 7.6 software (Tree Star, Inc.) was used for data
processing.
MMP measurement
Changes in MMP were detected using the MMP assay kit
with JC-1 staining, according to the manufacturer's instructions.
Cells were seeded in 12-well plates at a density of
4×105 cells/well. After attachment, cells were incubated
with ORES at 0, 5, 15 and 45 µM at 37°C for 24 h. The cells were
then harvested and washed with PBS. After incubation with JC-1
staining solution for 20 min at 37°C in the dark, cells were washed
twice with JC-1 buffer. Finally, cells were resuspended in PBS and
detected using a LSRFortessa TM X-20 flow cytometer (BD
Biosciences). FlowJo 7.6 software (FlowJo LLC) was used for data
processing. The polymer/monomer (red/green) fluorescence intensity
ratio was used to quantify the MMP.
Western blotting
Saos-2 cells were seeded in 6-well plates at
1×106 cells/well overnight at 37°C, and were then
incubated with ORES at 0, 5, 15 and 45 µM at 37°C for 24 h. In some
experiments, the Saos-2 cells were incubated with 30 µM ORES or 30
µM ORES and 20 ng/ml IL-6 at 37°C for 24 h. After washing with cold
phosphate-buffered saline (PBS) three times, cells were lysed in
RIPA lysis buffer and the protein concentration was detected using
a BCA protein assay kit, according to the manufacturer's
instructions. Each sample was adjusted to the same concentration
with lysis buffer and 20 µg protein was loaded per lane. Proteins
were then separated by SDS-PAGE on 10% gels and were transferred to
0.45 µm nitrocellulose filter membranes. The membranes were washed
and blocked in blocking buffer (5% non-fat milk) for 1 h at room
temperature. Subsequently, the membranes were incubated with
primary antibodies against cleaved caspase-9, cleaved caspase-3,
GAPDH, Bcl-2, Bcl-xL, Bad, Bax, P-STAT3, T-STAT3 and OPN (diluted
at 1:1,000) with gentle agitation overnight at 4°C. After three
washes, the membrane was incubated with HRP-conjugated anti-rabbit
secondary antibody (diluted at 1:3,000) with gentle agitation for 2
h at room temperature. Finally, the ECL reagent was used to detect
the target proteins. Semi-quantification of the bands was performed
using ImageJ software (1.48v; National Institutes of Health).
Statistical analysis
The experiments were repeated at least three times
independently, and data were presented as the mean ± SEM. One-way
ANOVA followed by Tukey's post hoc test was applied to compare the
differences between groups, using GraphPad Prism 5 (GraphPad
Software, Inc.). *P<0.05 was considered to indicate a
statistically significant difference.
Results
ORES inhibits the viability of Saos-2
cells
Although ORES has been shown to inhibit prostate and
colon cancer cell viability (30),
it remains unknown whether ORES can inhibit osteosarcoma cell
viability. As a natural phenolic compound, ORES is a resveratrol
analogue that contains an additional hydroxyl group in the aromatic
ring (22). The chemical structure
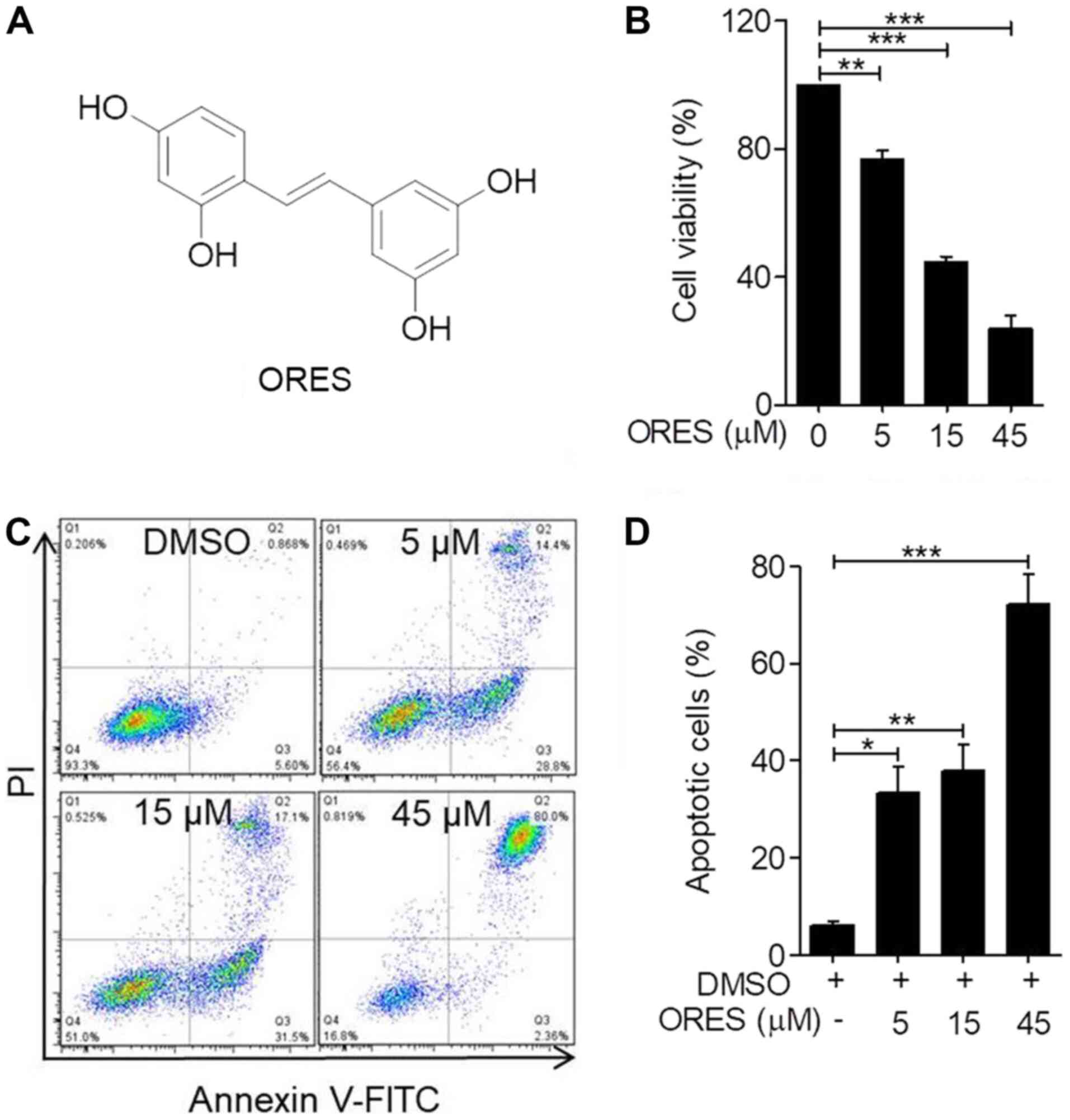
of ORES is shown in Fig. 1A. To
determine the effect of ORES on the proliferation of osteosarcoma
cells, the cell viability following ORES treatment in the Saos-2
cell line was detected. Cells were incubated with ORES at 0, 5, 15
and 45 µM for 48 h, and cell viability was detected using the CCK-8
assay. The results demonstrated that ORES significantly reduced
cell viability in a concentration-dependent manner (Fig. 1B). These results indicated that
ORES inhibits osteosarcoma cell viability.
ORES induces apoptosis of Saos-2
cells
Although ORES has been reported to induce apoptosis
of neuroblastoma cells (29), the
effect on osteosarcoma cells remains unclear. Following incubation
with ORES at 0, 5, 15 and 45 µM for 24 h, apoptotic cells were
detected using Annexin-FITC/PI staining. The results demonstrated
that treatment with ORES significantly increased the percentage of
apoptotic cells in a concentration-dependent manner compared with
that of the untreated group (Fig. 1C
and D).
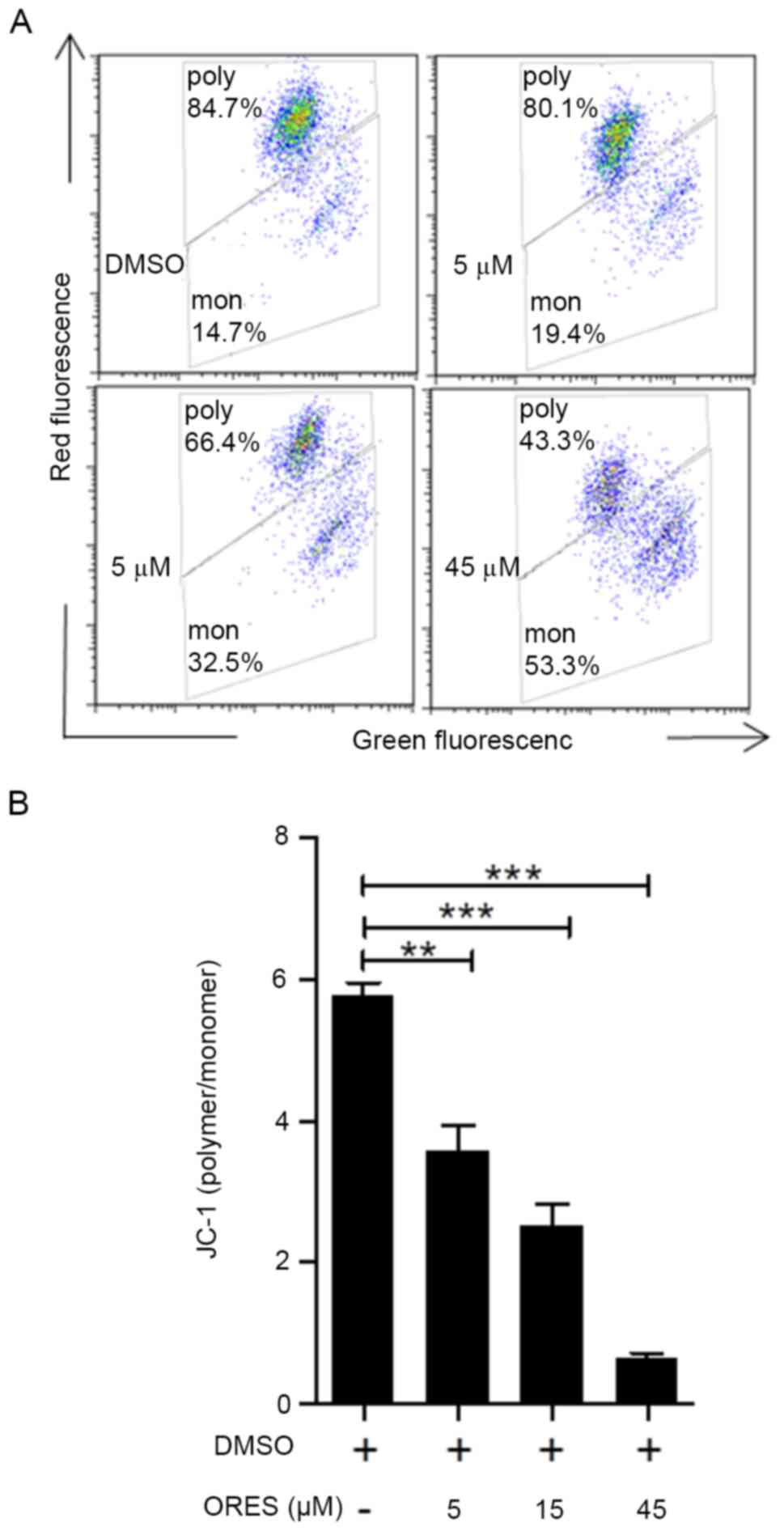
A decrease in MMP is a marker of early apoptosis. In
the mitochondria of normal cells, JC-1 is present as a polymer with
bright red fluorescence. Once the mitochondria-mediated apoptotic
pathway is initiated, JC-1 cannot aggregate and the JC-1 monomer
generates green fluorescence, according to the manufacturer's
instructions. The relative ratio of polymer/monomer (red/green)
fluorescence was used to measure the mitochondrial depolarization.
In response to ORES treatment at 0, 5, 15 and 45 µM for 24 h, cells
were clearly divided into two groups, with polymer at the top and
monomer at the bottom (Fig. 2A).
The ratio of JC-1 polymer/monomer significantly decreased in a
concentration-dependent manner in response to ORES compared with
that of the untreated group (Fig.
2B).
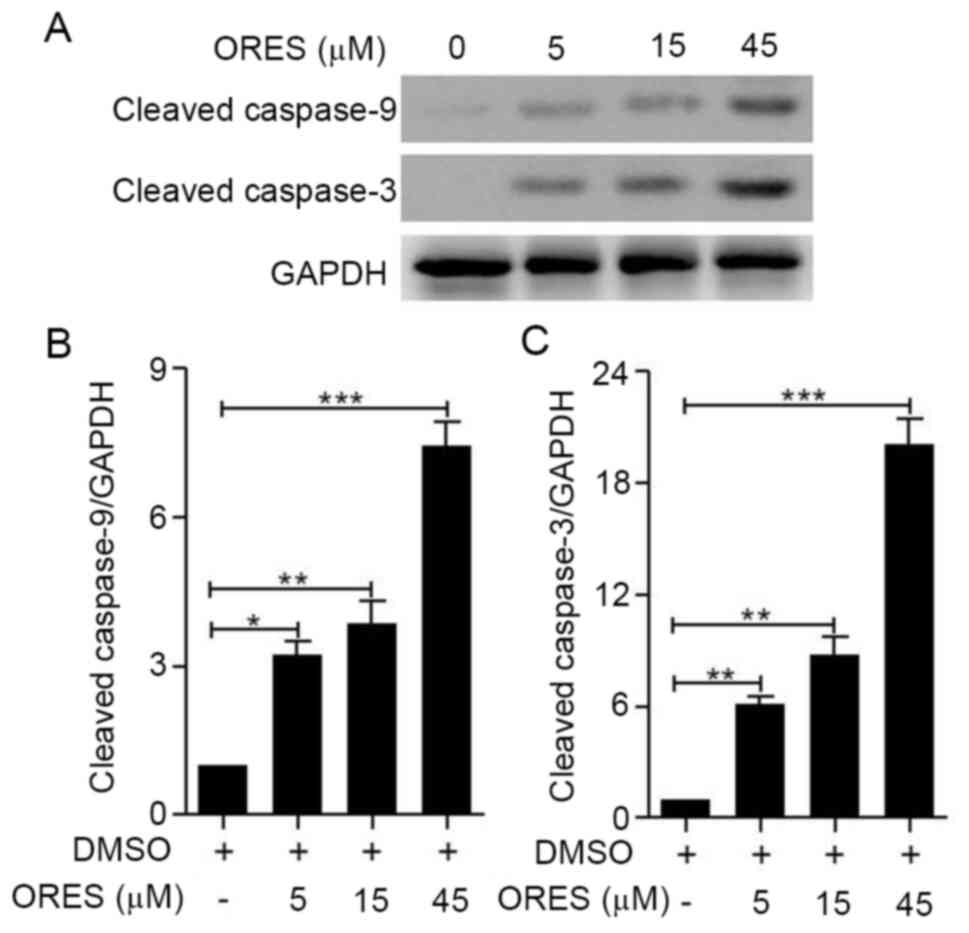
In the mitochondria-mediated pathway, cytochrome
c is released from mitochondria and induces the cleavage of
caspase-3 and caspase-9, which ultimately triggers apoptosis via
proteolysis of several cytoskeletal proteins, as well as the
cleavage of DNA via caspase-activated DNase (12). The results of the present study
demonstrated that compared with those of the untreated group, the
expression levels of cleaved caspase-9 and cleaved caspase-3 were
significantly increased by ORES treatment in a
concentration-dependent manner (Fig.
3A-C). These results suggested that ORES induces osteosarcoma
apoptosis.
ORES modulates apoptotic signaling
pathways in Saos-2 cells
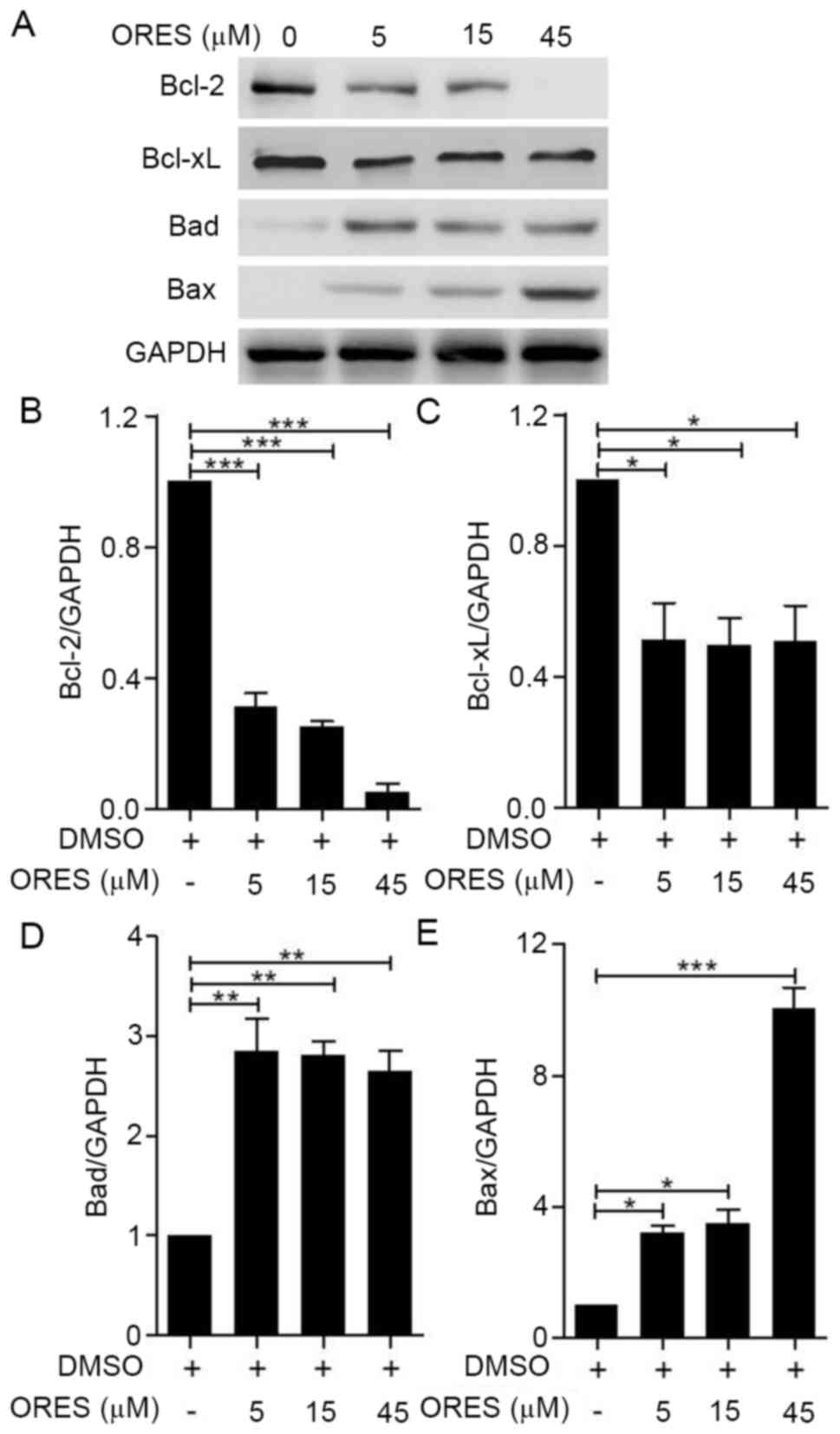
Given that ORES was demonstrated to promote
apoptosis, the mechanism by which ORES regulates the apoptotic
signaling pathways was further investigated. Western blotting was
used to detect the expression levels of Bcl-2 family proteins,
including Bcl-2, Bcl-xL, Bad and Bax (Fig. 4A). Mitochondria-mediated apoptosis
is regulated by Bcl-2 family proteins (10). Pro-apoptotic sensitizer protein Bad
can bind anti-apoptotic proteins Bcl-2 and Bcl-xL, which cause
pro-apoptotic activator proteins to dissociate from these
anti-apoptotic proteins and then bind to pro-apoptotic effector
proteins, such as Bax, to initiate apoptosis (11). Compared with those of the untreated
group, following treatment with ORES the expression levels of
anti-apoptotic proteins Bcl-2 (Fig.
4B) and Bcl-xL (Fig. 4C) were
significantly decreased, whereas the expression levels of
pro-apoptotic proteins Bad (Fig.
4D) and Bax (Fig. 4E) were
significantly increased. These results suggested that ORES may
regulate the mitochondria-mediated apoptotic signaling pathway in
Saos-2 cells.
ORES induces apoptosis and inhibits
cell viability via STAT3 phosphorylation in Saos-2 cells
Given that STAT3 serves an important role in the
proliferation and apoptosis of osteosarcoma cells (17), STAT3 activation in Saos-2 cells
upon treatment with ORES was further examined. Compared with those
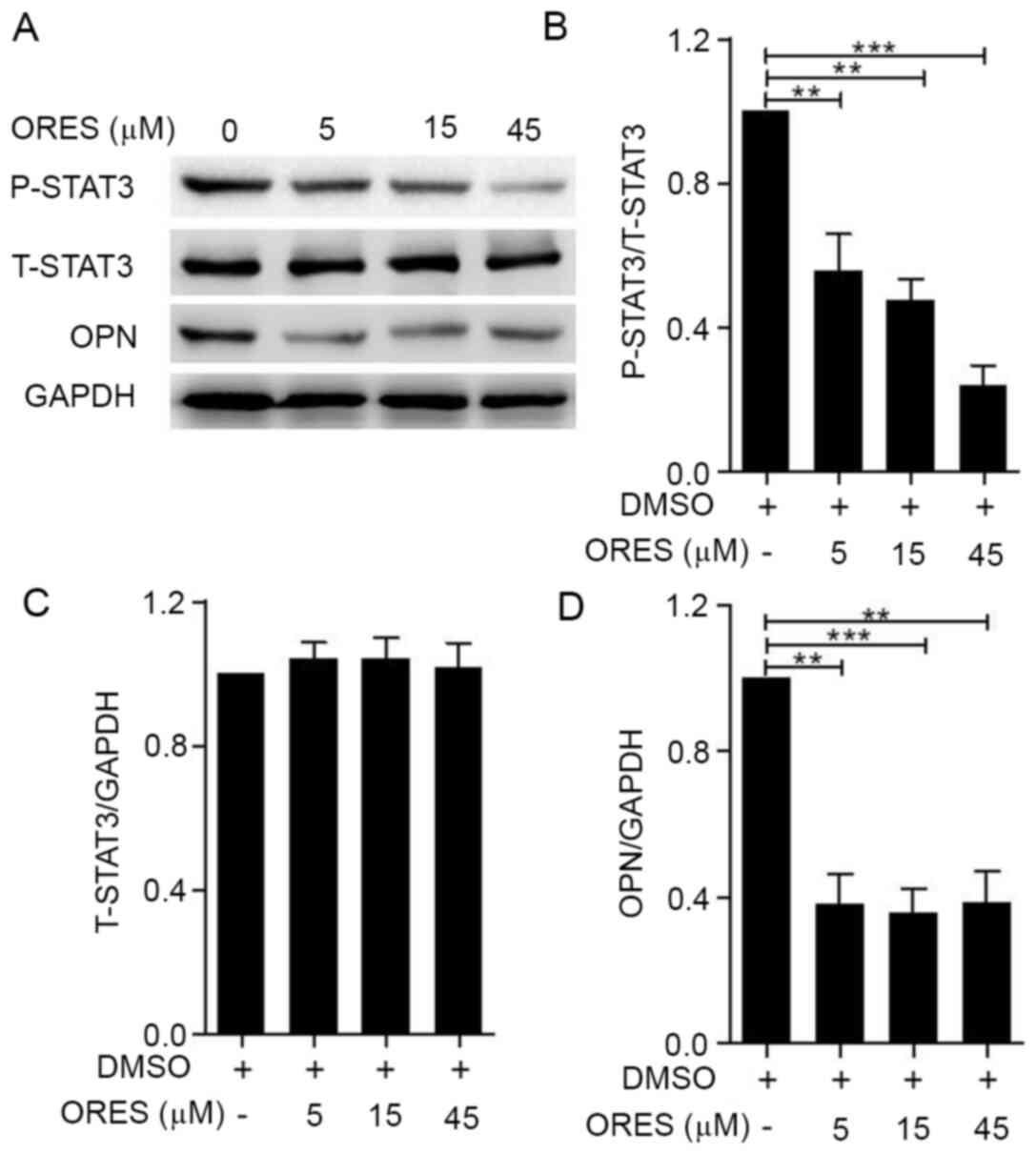
of the untreated group, after treatment with ORES at 5, 15 and 45
µM for 24 h, the expression levels of P-STAT3 were significantly
reduced in a concentration-dependent manner (Fig. 5A and B), whereas no significant
differences were observed in the expression levels of T-STAT3
(Fig. 5A and C). OPN is regulated
by STAT3 signaling, which is involved in osteosarcoma pathogenesis
(21). Compared with those of the
untreated group, treatment with ORES significantly decreased the
expression levels of OPN (Fig. 5A and
D). To determine whether ORES inhibits Saos-2 cell viability
via a reduction in STAT3 activation, IL-6 was used to activate
STAT3 signaling (36) in Saos-2
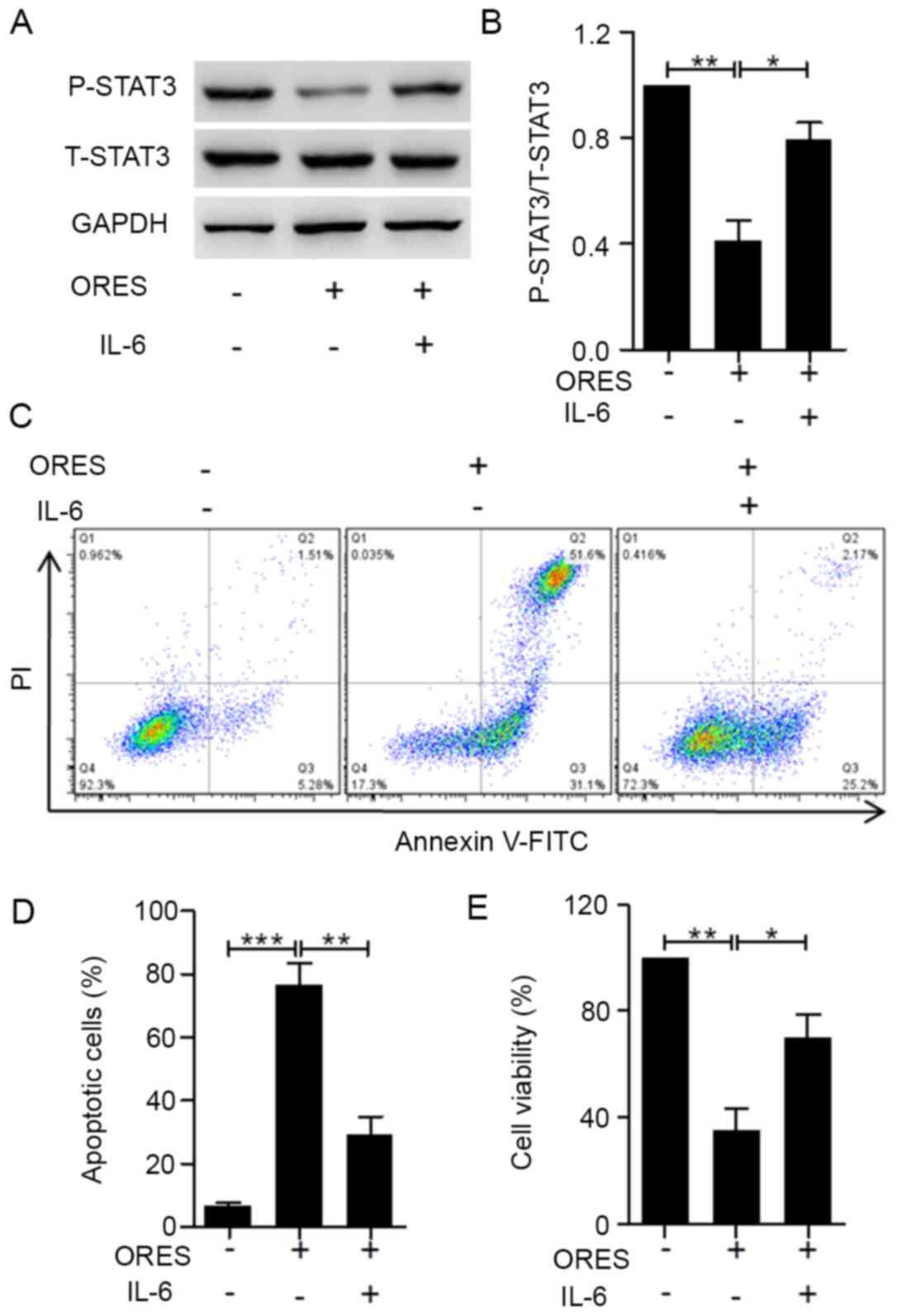
cells and reverse the effects of ORES. Upon treatment with 30 µM
ORES for 24 h, the phosphorylation of STAT3 was induced by
co-stimulation with 20 ng/ml IL-6 (Fig. 6A and B). After incubation with 20
ng/ml IL-6, the effects of ORES on the percentage of apoptotic
cells (Fig. 6C and D) and cell
viability (Fig. 6E) were reversed.
These results indicated that ORES induces apoptosis and inhibits
cell viability via partially reducing STAT3 signaling in Saos-2
cells.
Discussion
The results of the present study demonstrated that
ORES effectively inhibited cell viability and induced apoptosis of
Saos-2 cells. Upon treatment with ORES, the expression levels of
cleaved caspase-9 and cleaved caspase-3 were significantly
increased. Additionally, the expression levels of anti-apoptotic
proteins Bcl-2 and Bcl-xL were decreased, whereas the expression
levels of the pro-apoptotic proteins Bad and Bax were upregulated.
These results indicated that ORES has specific antitumor activity
against Saos-2 cells.
ORES is a natural compound derived from Morus
alba L., and its roles have been revealed in various disease
models. In macrophages, ORES has been proven to suppress
LPS-stimulated inflammatory responses by blocking the MAPK
signaling pathway (26). In
addition to its anti-inflammatory effects, ORES significantly
inhibited the proliferation of hepatocellular carcinoma (HCC) cells
(37), and head and neck squamous
cell carcinoma cells (28). In
both cases, the expression levels of vascular endothelial growth
factor (VEGF) were inhibited by ORES treatment, which indicates
that ORES may reduce micro-blood vessel density and micro-lymphatic
vessel density in tumors. As VEGF expression is regulated directly
by STAT3 in diverse human cancer cell lines (38), in addition to inhibition of cell
viability, ORES may target STAT3-mediated VEGF expression in
osteosarcoma. The inhibitory concentrations of ORES in various
tumor cells were different. The IC50 value of ORES in
T24 bladder cancer cells was 47.46 µM (39). In HCC cells, the IC50
values were nearly 80 µM (37). As
for neuroblastoma cells, the IC50 values reached 140 µM,
with little inhibitory effect against non-cancer cells, such as
Rat-2, HEK293, NIH3T3 and human peripheral blood mononuclear cells
(29). Therefore, Saos-2 cells
were relatively sensitive to ORES with an IC50 of <15
µM. In view of the broad biological activities of ORES, especially
its significant antitumor effects, further research should focus on
both cancer cells and tumor-surrounding cells, including
endothelial cells and immune cells.
It has been reported that multiple signaling
pathways influence the process of osteosarcoma cell growth and
anti-apoptosis, which can be interfered with using small molecule
compounds (40–42). For instance, caffeine has been
shown to induce apoptosis of osteosarcoma cells by inhibiting the
AKT/mTOR/ribosomal protein S6 kinase, NF-κB and MAPK
pathways (40). Glycogen synthase
kinase 3 inhibitors have also been demonstrated to inhibit
proliferation in osteosarcoma cell lines U2OS and MG-63 in
vitro (41). Messerschmitt
et al (42) reported that
specific tyrosine kinase inhibitors, such as inhibitors of EGF-R,
insulin-like growth factor 1 and Met, decreased motility and colony
formation in human osteosarcoma cell lines. Signaling pathways,
such as the hedgehog (43), Wnt
(44) and PI3K/AKT signaling
pathways (45), are all involved
in osteosarcoma proliferation. Notably, recently discovered types
of cell death, such as receptor-interacting protein (RIP)1- and
RIP3-dependent necroptosis, as well as
Ca2+/calmodulin-dependent protein kinase type II a
activity-related autophagic degradation, have been considered as
efficient anti-osteosarcoma strategies in recent research (46,47).
Resveratrol, which is structurally similar to ORES, has been shown
to induce the apoptosis of osteosarcoma cells via modulating the
microRNA139-5p/NOTCH1 signaling pathway (48). Additionally, resveratrol can
inhibit osteosarcoma MG-63 and MNNG/HOS cell proliferation and
tumorigenesis via blocking janus kinase (JAK)2/STAT3 signaling
(49). Consistent with this, in
the present study, as a potent tyrosinase inhibitor, ORES inhibited
the growth of osteosarcoma cells and promoted apoptosis by
affecting the STAT3 signaling pathway, which indicated that STAT3
may be another drug target for ORES.
STAT3 has already been confirmed to serve a crucial
role in selectively inducing and maintaining a carcinogenic
inflammatory microenvironment in response to cytokine stimulation
(17). The IL-6-JAK-STAT3
signaling axis contributes to the pathogenesis of myeloma cells by
preventing apoptosis (50). By
blocking the IL-6-OPN-STAT3 signaling pathway, the viability and
tumorigenesis of osteosarcoma cells are inhibited (21). This is consistent with the findings
of the present study, which demonstrated that compared with in the
control group, the phosphorylation of STAT3 was attenuated by ORES
treatment in Saos-2 cells. In addition, when P-STAT3 was partially
activated by IL-6, the inhibition of cell viability and apoptosis
promotion induced by ORES were also rescued. As a major cytokine in
the tumor microenvironment, IL-6 protects cancer cells against
apoptosis via multiple signaling pathways (51). The rescue of STAT3 phosphorylation
may explain in part the reversal effect of ORES. Given that
completely blocking STAT3 was demonstrated to be a risk factor
causing other diseases, such as hyper-immunoglobulin E syndrome
(52), inhibition of STAT3 should
be under strict control. Local suppression of STAT3 for a limited
period of time may convert inflammation in the tumor
microenvironment from promoting tumor growth to inhibition without
serious side effects (17).
FDA-approved tyrosine kinase inhibitors, such as sorafenib
(53) and sunitinib (54), are already in clinical use and can
indirectly inhibit STAT3 signaling, resulting in tumor cell cycle
arrest and the induction of apoptosis. The development of new STAT3
inhibitors may inhibit the viability and survival of multiple tumor
cells.
In conclusion, the present study demonstrated the
interaction between ORES and osteosarcoma. ORES induced apoptosis
and inhibited cell viability by reducing STAT3 signaling in Saos-2
cells. More comprehensive studies of ORES may establish it as a
promising agent for the future treatment of osteosarcoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81971753), the
Program for Medical Key Department of Shanghai (grant no.
ZK2019C01), the Program for the Outstanding Clinical Discipline
Project of Shanghai Pudong (grant no. PWYgy2018-09), the Key
Disciplines Group Construction Project of Pudong Health Bureau of
Shanghai (grant no. PWZxq2017-11), the Program for Outstanding
Leader of Shanghai (grant no. 046), and the Talents Training
Program of Pudong Hospital affiliated to Fudan University (grant
no. PJ201903).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
BY designed the project and drafted the manuscript.
TL and ZJ performed the experiments and drafted the manuscript. DL,
RA and XZ participated in data analysis and revised the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mirabello L, Troisi RJ and Savage SA:
International osteosarcoma incidence patterns in children and
adolescents, middle ages and elderly persons. Int J Cancer.
125:229–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Heymann MF, Lezot F and Heyman D: The
contribution of immune infiltrates and the local microenvironment
in the pathogenesis of osteosarcoma. Cell Immunol. 343:1037112019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gorlick R, Janeway K, Lessnick S, Randall
RL and Marina N; COG Bone Tumor Committee, : Children's oncology
group's 2013 blueprint for research: Bone tumors. Pediatr Blood
Cancer. 60:1009–1015. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the surveillance, epidemiology, and end results program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu KH, Lin RC, Yang JS, Yang WE, Reiter RJ
and Yang SF: Molecular and cellular mechanisms of melatonin in
osteosarcoma. Cells. 8:16182019. View Article : Google Scholar
|
|
6
|
Zhang Y, Cai L, Li D, Lao YH, Liu D, Li M,
Ding J and Chen X: Tumor microenvironment-responsive
hyaluronate-calcium carbonate hybrid nanoparticle enables effective
chemotherapy for primary and advanced osteosarcomas. Nano Res.
11:4806–4822. 2018. View Article : Google Scholar
|
|
7
|
Feng X, Xu W, Li Z, Song W, Ding J and
Chen X: Immunomodulatory nanosystems. Adv Sci (Weinh).
6:19001012019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang J, Li Z, Wang Z, Yu Y, Li D, Li B and
Ding J: Nanomaterials for combinational radio-immuno oncotherapy.
Adv Funct Mater. Oct 7–2020.(Epub ahead of print).
|
|
9
|
Lowe SW and Lin AW: Apoptosis in cancer.
Carcinogenesis. 21:485–495. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fulda S and Debatin KM: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bhola PD and Letai A: Mitochondria-judges
and executioners of cell death sentences. Mol Cell. 61:695–704.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Adams JM and Cory S: The Bcl-2 apoptotic
switch in cancer development and therapy. Oncogene. 26:1324–1337.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen G, Wang F, Trachootham D and Huang P:
Preferential killing of cancer cells with mitochondrial dysfunction
by natural compounds. Mitochondrion. 10:614–625. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu Y, Yu K, Wang G, Zhang D, Shi C, Ding
Y, Hong D, Zhang D, He H, Sun L, et al: Lanatoside C inhibits cell
proliferation and induces apoptosis through attenuating
Wnt/β-catenin/c-Myc signaling pathway in human gastric cancer cell.
Biochem Pharmacol. 150:280–292. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Z, Yu K, Hu Y, Su F, Gao Z, Hu T,
Yang Y, Cao X and Qian F: Schisantherin A induces cell apoptosis
through ROS/JNK signaling pathway in human gastric cancer cells.
Biochem Pharmacol. 173:1136732020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ashkenazi A, Fairbrother WJ, Leverson JD
and Souers AJ: From basic apoptosis discoveries to advanced
selective BCL-2 family inhibitors. Nat Rev Drug Discov. 16:273–284.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu H, Pardoll D and Jove R: STATs in
cancer inflammation and immunity: A leading role for STAT3. Nat Rev
Cancer. 9:798–809. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang YC, Zheng LH, Ma BA, Zhou Y, Zhang
MH, Zhang DZ and Fan QY: Clinical value of signal transducers and
activators of transcription 3 (STAT3) gene expression in human
osteosarcoma. Acta Histochem. 113:402–408. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang X, Goldstein D, Crowe PJ and Yang JL:
Impact of STAT3 inhibition on survival of osteosarcoma cell lines.
Anticancer Res. 34:6537–6545. 2014.PubMed/NCBI
|
|
20
|
Goel S, Sahu S, Minz RW, Singh S, Suri D,
Oh YM, Rawat A, Sehgal S and Saikia B: STAT3-mediated
transcriptional regulation of osteopontin in STAT3 loss-of-function
related hyper IgE syndrome. Front Immunol. 9:10802018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang C, Ma K and Li WY: Cinobufagin
suppresses the characteristics of osteosarcoma cancer cells by
inhibiting the IL-6-OPN-STAT3 pathway. Drug Des Devel Ther.
13:4075–4090. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim YM, Yun J, Lee CK, Lee H, Min KR and
Kim Y: Oxyresveratrol and hydroxystilbene compounds. Inhibitory
effect on tyrosinase and mechanism of action. J Biol Chem.
277:16340–16344. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ortiz-Ruiz CV, Ballesta de Los Santos M,
Berna J, Fenoll J, Garcia-Ruiz PA, Tudela J and Garcia-Canovas F:
Kinetic characterization of oxyresveratrol as a tyrosinase
substrate. IUBMB Life. 67:828–836. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Heo JI, Kim JH, Lee JM, Kho YJ, Lim SS,
Park JB, Kim J, Kim SC and Lee JY: FOXO3a Activation by
oxyresveratrol of Morus bombycis koidzumi extract mediates
antioxidant activity. Anim Cells Syst. 20:39–47. 2016. View Article : Google Scholar
|
|
25
|
Jia YN, Lu HP, Peng YL, Zhang BS, Gong XB,
Su J, Zhou Y, Pan MH and Xu L: Oxyresveratrol prevents
lipopolysaccharide/D-galactosamine-induced acute liver injury in
mice. Int Immunopharmacol. 56:105–112. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee HS, Kim DH, Hong JE, Lee JY and Kim
EJ: Oxyresveratrol suppresses lipopolysaccharide-induced
inflammatory responses in murine macrophages. Hum Exp Toxicol.
34:808–818. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wei J, Chen JR, Pais EMA, Wang TY, Miao L,
Li L, Li LY, Qiu F, Hu LM, Gao XM and Fan GW: Oxyresveratrol is a
phytoestrogen exerting anti-inflammatory effects through NF-κB and
estrogen receptor signaling. Inflammation. 40:1285–1296. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sintuyanon N, Phoolcharoen W, Pavasant P
and Sooampon S: Resveratrol demonstrated higher antiproliferative
and antiangiogenic efficacy compared with oxyresveratrol on head
and neck squamous cell carcinoma cell lines. Natural Prod Commun.
12:1781–1784. 2017.
|
|
29
|
Rahman MA, Bishayee K, Sadra A and Huh SO:
Oxyresveratrol activates parallel apoptotic and autophagic cell
death pathways in neuroblastoma cells. Biochim Biophys Acta Gen
Subj. 1861:23–36. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Matencio A, Dhakar NK, Bessone F, Musso G,
Cavalli R, Dianzani C, García-Carmona F, López-Nicolás JM and
Trotta F: Study of oxyresveratrol complexes with insoluble
cyclodextrin based nanosponges: Developing a novel way to obtain
their complexation constants and application in an anticancer
study. Carbohydr Polym. 231:1157632020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee JH, Baek SY, Jang EJ, Ku SK, Kim KM,
Ki SH, Kim CE, Park KI, Kim SC and Kim YW: Oxyresveratrol
ameliorates nonalcoholic fatty liver disease by regulating hepatic
lipogenesis and fatty acid oxidation through liver kinase B1 and
AMP-activated protein kinase. Chem Biol Interact. 289:68–74. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tadtong S, Chatsumpun N, Sritularak B,
Jongbunprasert V, Ploypradith P and Likhitwitayawuid K: Effects of
oxyresveratrol and its derivatives on cultured P19-derived neurons.
Trop J Pharm Res. 15:2619–2628. 2016. View Article : Google Scholar
|
|
33
|
Lee HJ, Feng JH, Sim SM, Lim SS, Lee JY
and Suh HW: Effects of resveratrol and oxyresveratrol on
hippocampal cell death induced by kainic acid. Anim Cells Syst
(Seoul). 23:246–252. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Duan C, Han J, Zhang C, Wu K and Lin Y:
Inhibition of kidney cancer cell growth by Mulberroside-A is
mediated via mitochondrial mediated apoptosis, inhibition of cell
migration and invasion and targeting EGFR signalling pathway. J
BUON. 24:296–300. 2019.PubMed/NCBI
|
|
35
|
Li ZR, Ma T, Guo YJ, Hu B, Niu SH, Suo FZ,
Du LN, You YH, Kang WT, Liu S, et al: Sanggenon O induced apoptosis
of A549 cells is counterbalanced by protective autophagy. Bioorg
Chem. 87:688–698. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Che Q, Xiao X, Liu M, Lu Y, Doug X and Liu
S: IL-6 promotes endometrial cancer cells invasion and migration
through signal transducers and activators of transcription 3
signaling pathway. Pathol Res Pract. 215:1523922019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu Y, Ren W, Bai Y, Wan L, Sun X, Liu Y,
Xiong W, Zhang YY and Zhou L: Oxyresveratrol prevents murine H22
hepatocellular carcinoma growth and lymph node metastasis via
inhibiting tumor angiogenesis and lymphangiogenesis. J Nat Med.
72:481–492. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Niu G, Wright KL, Huang M, Song L, Haura
E, Turkson J, Zhang S, Wang T, Sinibaldi D, Coppola D, et al:
Constitutive Stat3 activity up-regulates VEGF expression and tumor
angiogenesis. Oncogene. 21:2000–2008. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang Y, Zhang G, Li C, Wang S, Zhu M, Wang
J, Yue H, Ma X, Zhen Y and Shu X: Metabolic profile and
structure-activity relationship of resveratrol and its analogs in
human bladder cancer cells. Cancer Manag Res. 11:4631–4642. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Miwa S, Sugimoto N, Yamamoto N, Shirai T,
Nishida H, Hayashi K, Kimura H, Takeuchi A, Igarashi K, Yachie A
and Tsuchiya H: Caffeine induces apoptosis of osteosarcoma cells by
inhibiting AKT/mTOR/S6K, NF-κB and MAPK pathways. Anticancer Res.
32:3643–3649. 2012.PubMed/NCBI
|
|
41
|
Lu K, Wang X, Chen Y, Liang D, Luo H, Long
L, Hu Z and Bao J: Identification of two potential glycogen
synthase kinase 3β inhibitors for the treatment of osteosarcoma.
Acta Biochim Biophys Sin (Shanghai). 50:456–464. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Messerschmitt PJ, Rettew AN, Brookover RE,
Garcia RM, Getty PJ and Greenfield EM: Specific tyrosine kinase
inhibitors regulate human osteosarcoma cells in vitro. Clin Orthop
Relat Res. 466:2168–2175. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kumar RM and Fuchs B: Hedgehog signaling
inhibitors as anti-cancer agents in osteosarcoma. Cancers (Basel).
7:784–794. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
McQueen P, Ghaffar S, Guo Y, Rubin EM, Zi
XL and Hoangn BH: The Wnt signaling pathway: Implications for
therapy in osteosarcoma. Expert Rev Anticancer Ther. 11:1223–1232.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Qian H, Huang T, Chen Y, Li X, Gong W,
Jiang G, Zhang W, Cheng S, Li X and Li P: X-linked inhibitor of
apoptosis protein inhibitor Embelin induces apoptosis via PI3K/Akt
pathway and inhibits invasion in osteosarcoma cells. J Cancer Res
Ther. 14 (Supplement):S648–S655. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li S, Zhang T, Xu W, Ding J, Yin F, Xu J,
Sun W, Wang H, Sun M, Cai Z and Hua Y: Sarcoma-targeting
peptide-decorated polypeptide nanogel intracellularly delivers
shikonin for upregulated osteosarcoma necroptosis and diminished
pulmonary metastasis. Theranostics. 8:1361–1375. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xu J, Wang H, Hu Y, Zhang YS, Wen L, Yin
F, Wang Z, Zhang Y, Li S, Miao Y, et al: Inhibition of CaMKIIα
activity enhances antitumor effect of fullerene C60 nanocrystals by
suppression of autophagic degradation. Adv Sci (Weinh).
6:18012332019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xiao X, Zhang Y, Pan W and Chen F:
MiR-139-mediated NOTCH1 regulation is crucial for the inhibition of
osteosarcoma progression caused by resveratrol. Life Sci.
242:1172152020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Peng L and Jiang D: Resveratrol eliminates
cancer stem cells of osteosarcoma by STAT3 pathway inhibition. PLoS
One. 13:e02059182018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Catlett-Falcone R, Landowski TH, Oshiro
MM, Turkson J, Levitzki A, Savino R, Ciliberto G, Moscinski L,
Fernández-Luna JL, Nuñez G, et al: Constitutive activation of Stat3
signaling confers resistance to apoptosis in human U266 myeloma
cells. Immunity. 10:105–115. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kumari N, Dwarakanath BS, Das A and Bhatt
AN: Role of interleukin-6 in cancer progression and therapeutic
resistance. Tumour Biol. 37:11553–11572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Minegishi Y, Saito M, Tsuchiya S, Tsuge I,
Takada H, Hara T, Kawamura N, Ariga T, Pasic S, Stojkovic O, et al:
Dominant-negative mutations in the DNA-binding domain of STAT3
cause hyper-IgE syndrome. Nature. 448:1058–1062. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yang F, Van Meter TE, Buettner R, Hedvat
M, Liang W, Kowolik CM, Mepani N, Mirosevich J, Nam S, Chen MY, et
al: Sorafenib inhibits signal transducer and activator of
transcription 3 signaling associated with growth arrest and
apoptosis of medulloblastomas. Mol Cancer Ther. 7:3519–3526. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xin H, Zhang C, Herrmann A, Du Y, Figlin R
and Yu H: Sunitinib inhibition of Stat3 induces renal cell
carcinoma tumor cell apoptosis and reduces immunosuppressive cells.
Cancer Res. 69:2506–2513. 2009. View Article : Google Scholar : PubMed/NCBI
|