Introduction
Nephroblastoma is the most common malignancy of the
urinary tract in children, accounting for ~8% of all pediatric
solid tumors (1). Nephroblastoma
most commonly occurs at the ages of 1–3 years and affects ~1 in
10,000 individuals (2).
Nephroblastoma, which most commonly manifests as an abdominal mass,
is associated with high morbidity and mortality rates and adversely
affects the quality of life of the patients (3). Combination treatment with surgery,
radiotherapy and chemotherapy has greatly improved the survival
rate of patients with nephroblastoma (4). Currently, the 5-year overall
survival rate of patients with nephroblastoma is ~85%. However, due
to the insidious onset and rapid progression of the disease, 15% of
all children have a poor prognosis (5). Therefore, identifying novel
diagnostic markers and treatment approaches for children with
nephroblastoma is crucial.
Transcription factors (TF) are proteins that can
effectively bind upstream of particular genes, thus regulating
their expression and driving a series of cellular processes
(6). It has been reported that
TFs serve a central role in the development of life, while their
inactivation, which determines cell fate, is considered as an
important step in tumorigenesis (7). Homeobox (HOX) genes are involved in
regulating cell differentiation and morphological development and
are therefore at the highest level of the genetic hierarchy
(8). Homeobox protein Hox-B2
(HOXB2) is a member of the HOX family. A previous study
demonstrated that HOXB2 served an important role in the
proliferation, invasion and migration of ovarian cancer cells
(9). In addition, microRNA-139-5p
attenuated the proliferation of lung cancer cells via targeting
HOXB2 (10). Furthermore, another
study revealed that HOXB2 could promote the apoptosis of gastric
cancer cells and enhance their sensitivity to cisplatin (10). Jing et al (11) demonstrated that HOXB2 and forkhead
box C1 (FOXC1) were upregulated in nephroblastoma tissues, while
their expression was closely associated with tumor stage and lymph
node metastasis. These findings suggested that HOXB2 and FOXC1
could synergistically promote the development of malignant tumors
via enhancing the proliferative and migratory potential of
nephroblastoma cells (11).
However, the specific mechanism underlying the effects of HOXB2 on
the proliferation and migration of nephroblastoma cells remains
unclear.
In the present study, the binding potential of HOXB2
on the nucleolar spindle-associated protein 1 (NUSAP1) promoter was
predicted using the JASPAR database. NUSAP1, a microtubule-binding
protein, is highly conserved in humans and serves an indispensable
role in mitosis via regulating the proper assembly of the spindle
and the formation of chromosomes (12,13). It has been reported that, in
normal tissues, the expression of NUSAP1 is tightly regulated;
however, when NUSAP1 is abnormally upregulated, cell mitosis cannot
be terminated, resulting in infinite cell proliferation (14). This finding could explain why the
expression of NUSAP1 is abnormally elevated in cancer. Another
study analyzed the gene expression profile in nephroblastoma using
the Target Data Matrix database and showed that NUSAP1 was
significantly upregulated (15).
However, to the best of our knowledge, the role of NUSAP1 in
nephroblastoma has not been previously reported.
Therefore, the current study aimed to investigate
regulatory mechanisms in terms of TF and to predict significant
regulatory genes involved in specific signaling pathways, in order
to uncover the mechanism underlying the effect of HOXB2 and its
downstream factor NUSAP1 in nephroblastoma, thus providing novel
targets for the diagnosis and treatment of nephroblastoma.
Materials and methods
Cell culture
The nephroblastoma cell lines, GHINK-1, WiT49, 17–94
and HFWT, and 293T cells were obtained from the American Type
Culture Collection. The cells were cultured in DMEM (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.) at 37°C in a 5% CO2
incubator.
Database
The association between NUSAP1 and HOXB2 was
predicted by bioinformatics analysis using the JASPAR database
(jaspar.genereg.net).
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and was then reverse transcribed into cDNA using MMLV reverse
transcriptase (Promega Corporation) according to the manufacturer's
specifications. qPCR analysis was carried out using SYBR Green mix
(Thermo Fisher Scientific, Inc.). The PCR thermocycling conditions
applied were as follows: Initial holding period at 95°C for 30 sec,
followed by 40 cycles of 94°C for 5 sec and 60°C for 30 sec. GAPDH
served as an internal reference gene. The primer sequences used
were as follows: NUSAP1 forward, 5′-CGTCCCCTCAACTATGAACCAC-3′ and
reverse, 5′-GCGTTTCTTCCGTTGCTCTT-3′; HOXB2 forward,
5′-CGCCAGGATTCACCTTTCCTT-3′ and reverse,
5′-CCCTGTAGGCTAGGGGAGAG-3′; and GAPDH forward,
5′-AGAAGGCTGGGGCTCATTTG-3′ and reverse, 5′-AGGGGCCATCCACAGTCTTC-3′.
The quantification was performed using the 2−ΔΔCq method
(16).
Western blot analysis
Cells were lysed with RIPA buffer (Beyotime
Institute of Biotechnology) with phosphatase inhibitor AEBSF
(Beyotime Institute of Biotechnology) on ice for 30 mins. Following
centrifugation at 350 × g for 20 min at 4°C, a BCA protein assay
kit (Sigma-Aldrich; Merck KGaA) was utilized to quantify the
protein concentration. Proteins (40 µg/lane) were separated by 10%
SDS-PAGE and were then transferred onto PVDF membranes (Bio-Rad
Laboratories, Inc.). Following blocking with 5% skimmed milk for
1.5 h at room temperature, the membranes were incubated with
primary antibodies against NUSAP1 (1:1,000; cat. no. ab137230),
HOXB2 (1:1,000; cat. no. ab220390), phosphorylated (p)-PI3K
(1:1,000; cat. no. ab191606), p-Akt (1:1,000; cat. no. ab38449),
E-cadherin (1:1,000; cat. no. ab40772), N-cadherin (1:1,000; cat.
no. ab76011), Vimentin (1:1,000; cat. no. ab92547), PI3K (1:1,000;
cat. no. ab32089), Akt (1:1,000; cat. no. ab18785) and GAPDH
(1:1,000; cat. no. ab8245; all from Abcam) at 4°C overnight. The
next day, the membranes were washed and incubated with the
corresponding secondary antibodies (1:5,000; cat. no. ab150077;
Abcam) at room temperature for 1 h. For p-PI3K and p-Akt, after the
detection of PI3K and Akt, the same membrane was washed with
stripping buffer (cat. no. P0025N; Beyotime Institute of
Biotechnology), and the aforementioned protein detection steps were
repeated to detect the expression of p-PI3K and p-Akt. Protein
signals were detected using ECL detection kit (Amersham; Cytiva)
and semi-quantitative analysis was conducted using ImageJ software
(version 1.8.0; National Institutes of Health).
Cell transfection
HFWT cells were cultured in a 6-well plate at a
density of 1×106 cells/well at 37°C in a 5%
CO2 incubator for 24 h. The lentiviral vectors packaging
short hairpin RNA (shRNA) NUSAP1 (sh-NUSAP1), negative control
shRNA (sh-NC), HOXB2 overexpression plasmid (Ov-HOXB2) and Ov-NC
were obtained from Shanghai GeneChem Co., Ltd., and were then
transduced into cells at 20 nM using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The transfection efficiency was
verified by RT-qPCR and western blot analyses after 48 h.
MTT assay
Following culture for 48 h at 37°C in a 5%
CO2 incubator, HFWT cells in each well at a density of
8×103 cells/well were supplemented with 20 µl MTT
solution and incubated for an additional 3 h. Subsequently, the
supernatant was removed and 200 µl DMSO was added into each well to
dissolve formazan crystals. The absorbance at 490 nm was detected
using a microplate reader (Multiskan Sky; Thermo Fisher Scientific,
Inc.).
EdU staining
Cell proliferation was assessed using the Cell-Light
EdU DNA Cell Proliferation kit (Guangzhou RiboBio Co., Ltd.)
according to the manufacturer's protocol. Briefly, following
transfection, the HFWT cells were stained with fluorescent-labeled
EdU for 2 h at room temperature. Subsequently, cells were fixed in
4% ethanol for 30 min at room temperature and were then stained
with Apollo reaction buffer liquid at 37°C for 30 min in the dark.
Image acquisition was performed using a laser confocal microscope
(magnification, ×200).
Colony formation assay
Transfected HFWT cells were cultured normally in a
6-well plate at a density of 1×103 cells/well. Following
incubation for 2 weeks, cells were fixed with methanol for 15 min
at room temperature and stained with 0.1% crystal violet for 20
mins at room temperature. Finally, the visible colonies with a
diameter >0.5 mm were counted using ImageJ software (version
1.8.0; National Institutes of Health) and images were captured
under a microscope.
Wound healing assay
For wound healing assays, HFWT cells were grown to
100% confluence in 6-well plates after transfection. Subsequently,
two linear wounds were created in each well by scratching the
monolayer with a sterile pipette tip. The wells were then washed
with PBS and cells were cultured in serum-free DMEM for 24 h. To
assess cell migration, images of the same areas in the wells were
captured at 0 and 24 h using a light microscope following wounding.
Wound healing was evaluated by measuring the total surface area
covered by the cells.
Transwell assay
To assess cell invasion, 24-well Transwell plates
(Corning, Inc.) with 8-µm pore inserts were coated with Matrigel
(BD Biosciences) at 37°C for 30 min. The transfected HFWT cells
(5×105 cells/well) in serum-free DMEM were seeded onto
the Matrigel-coated upper chamber, while the lower chamber was
supplemented with DMEM containing 10% FBS. Following incubation for
24 h at 37°C, the cells on the upper chamber were gently removed
with cotton swabs. The invading cells in the lower chamber were
fixed in alcohol for 15 min and stained with 0.1% crystal violet
solution for 10 min at room temperature. Finally, the cells were
observed under an optical microscope (Olympus Corporation;
magnification, ×100).
Dual-luciferase reporter assay
To generate the luciferase construct, 0.5 µg vectors
containing the 3′-untranslated region (UTR) of wild-type (WT)
NUSAP1 or mutant (MUT) 3′-UTR NUSAP1using a QuickMutation kit
(Beyotime Institute of Biotechnology), with control vector or HOXB2
overexpression vector and pMIR-Renilla vector (Shanghai
GeneChem Co., Ltd) were synthesized and cloned into GV272 vectors
by GeneChem (China). HFWT cells were co-transfected using
Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) with WT or MUT 3′-UTR of NUSAP1 (NUSAP1-WT or NUSAP1-MUT,
respectively) and Ov-HOXB2 or Ov-NC for 48 h. A site directed
mutagenesis kit was used to mutant 3′-untranslated region of
NUSAP1. Following transfection, the relative luciferase activity
was determined after 48h using the Dual-Luciferase Reporter Assay
kit (Promega Corporation), according to the manufacturer's
instructions. Renilla luciferase activity was used to
normalize the firefly luciferase activity.
Chromatin immunoprecipitation (ChIP)
assay
The ChIP assay was carried out using the Chromatin
Immunoprecipitation Assay Kit (MilliporeSigma), according to the
manufacturer's instructions. Briefly, the cells were lysed with 100
µl SDS lysis buffer (Beyotime Institute of Biotechnology) and 500
µl of ChIP sonication nuclear lysis buffer was added to cell
particles and centrifuged at 1,000 × g at 4°C for 5 mins. Then, 10
µg fragmented chromatin was incubated with 2 µg anti-IgG (cat. no.
A-11031) and 5 µg anti-HOXB2 (cat. no. PA5-101640) antibodies
(1:500; both Thermo Fisher Scientific, Inc.). Subsequently, 20 µl
protein A+G agarose beads (Beyotime Institute of Biotechnology) was
added and incubated for another 4 h at 4°C. After the supernatant
was removed, the collected agarose beads were washed five times
with 100 µl PBS (0.01 M; pH 7.4) and denatured by boiling for 5
min. Total RNA was then subjected to reverse transcription to cDNA
using SMART MMLC Reverse Transcriptase (Takara Biotechnology Co.,
Ltd.). The recruited DNA was then subjected to PCR using primers
detecting the distal enhancer regions of NUSAP1 using an ABI 7500
system (Thermo Fisher Scientific, Inc.). Subsequently, the PCR
products were electrophoresed in 1% agarose gel, stained in
ethidium bromide for ~15 min, rinsed with water and photographed by
gel imaging system. Gel images and positive bands with average
optical density values were analyzed by Gel optical density
analysis software Gel pro4.0. The resulting data were normalized to
housekeeping gene β-actin.
Statistical analysis
All data are expressed as the mean ± SD of three
independent experimental repeats. All statistical analyses were
performed using GraphPad Prism 6.0 (GraphPad Software, Inc.). The
differences between multiple groups were compared by one-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
NUSAP1 knockdown attenuates the
proliferation of nephroblastoma cells
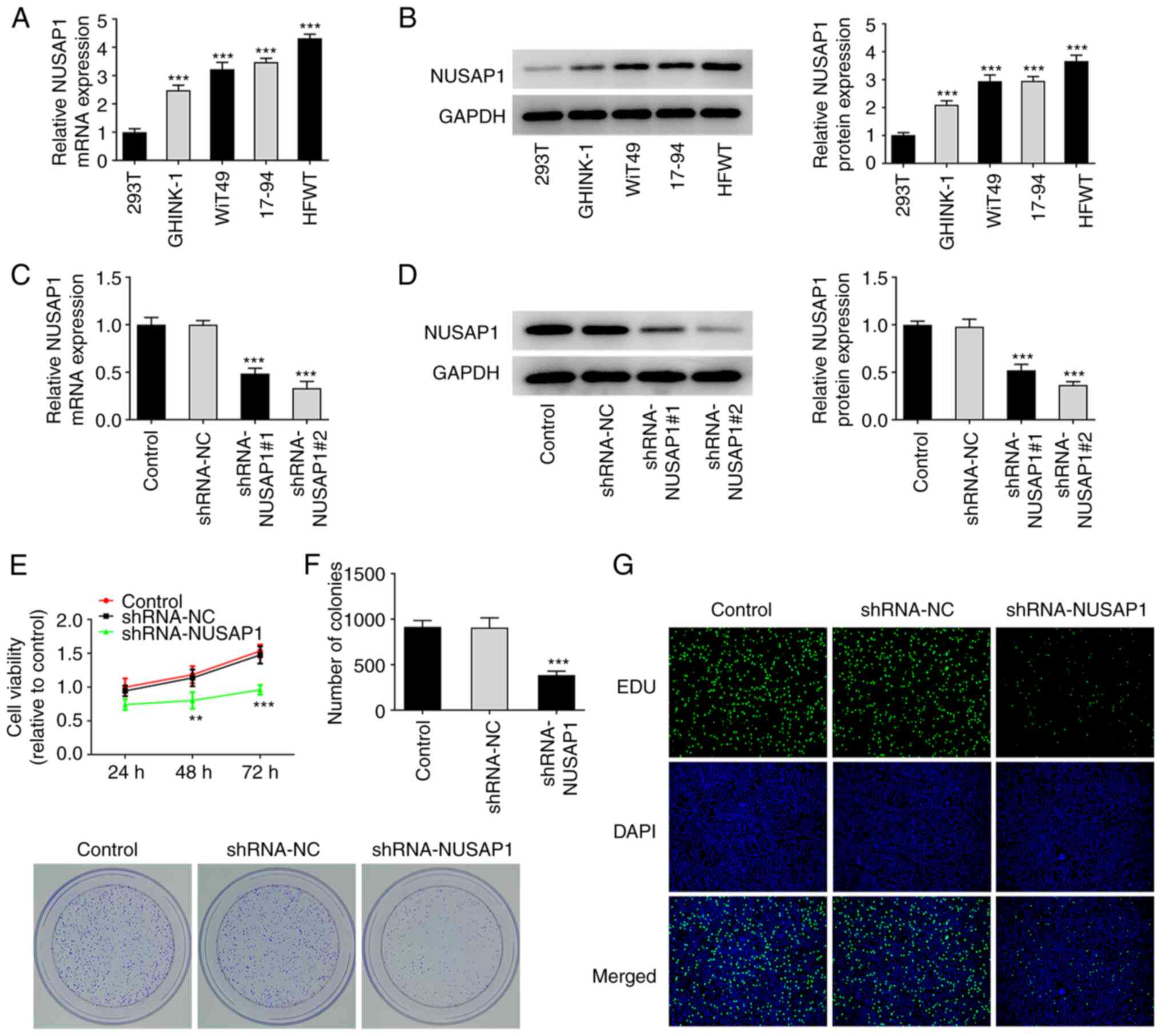
The mRNA and protein expression levels of NUSAP1 in
nephroblastoma cells were detected via RT-qPCR and western blot
analyses, respectively. The results showed that the expression of
NUSAP1 was significantly increased in nephroblastoma cell lines
compared with 293T cells (Fig. 1A and
B). The expression of NUSAP1 was more notably increased in HFWT
cells. Therefore, HFWT cells were used for the subsequent
experiments. Subsequently, the expression of NUSAP1 was knocked
down in HFWT cells following cell transfection with sh-NUSAP1 and
the transfection efficiency was evaluated using RT-qPCR and western
blot analysis. As shown in Fig. 1C
and D, the expression of NUSAP4 was significantly decreased in
cells transfected with shRNA-NUSAP1#2 and this shRNA clone was
therefore used for the subsequent experiments. To evaluate cell
viability, cells were first divided into the control, sh-NC and
sh-NUSAP1 groups followed by MTT assay. The results showed that
cell viability was significantly reduced in the sh-NUSAP1 group
compared with the sh-NC group (Fig.
1E). Cell proliferation was assessed using colony formation
assay. As shown in Fig. 1F,
compared with the sh-NC group, the proliferative ability of cells
was significantly decreased in the sh-NAUSAP1 group. The results of
EdU staining were consistent with those obtained with the colony
formation assay (Fig. 1G). The
aforementioned findings suggested that NUSAP1 silencing could
attenuate the proliferation of nephroblastoma cells.
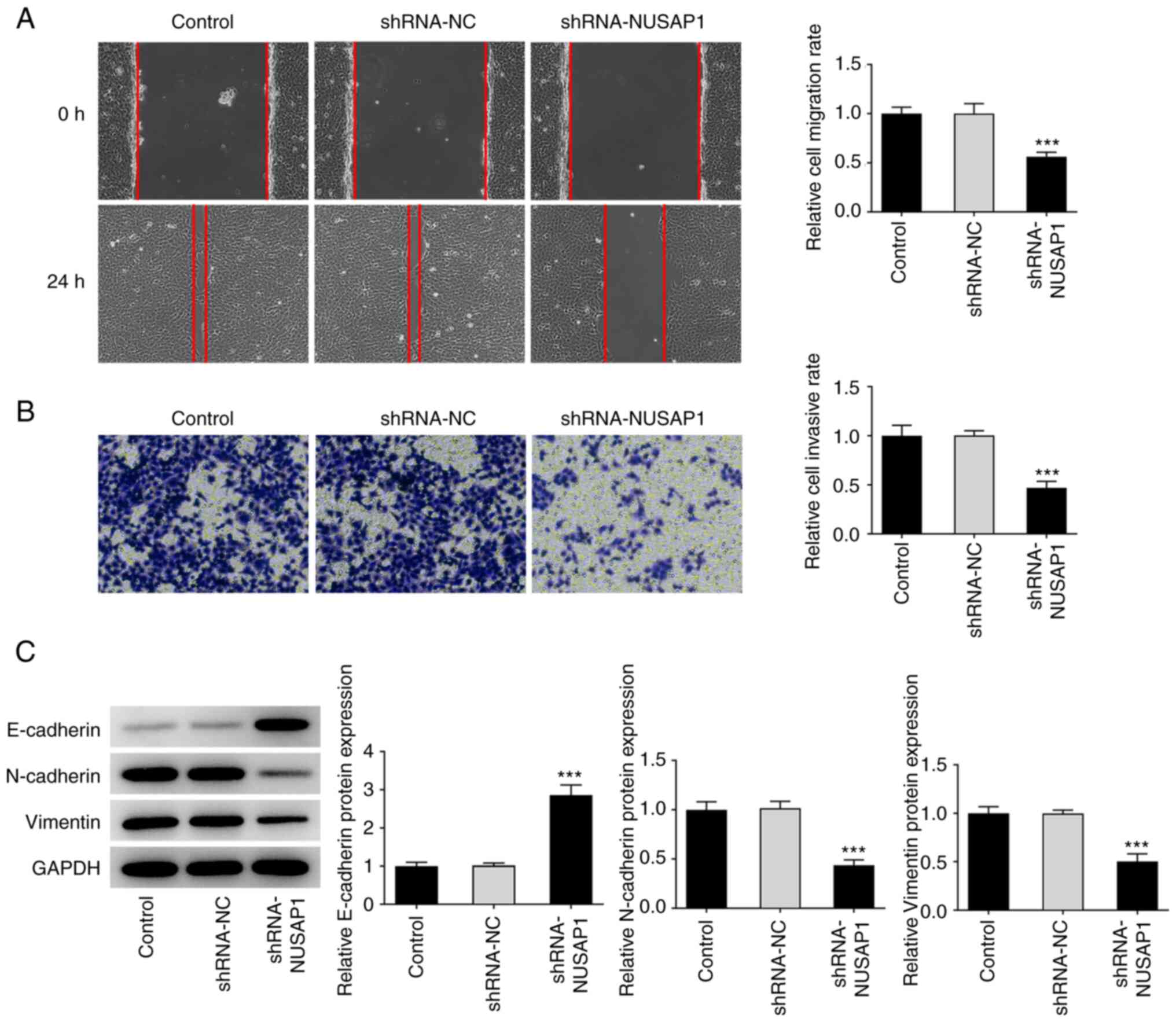
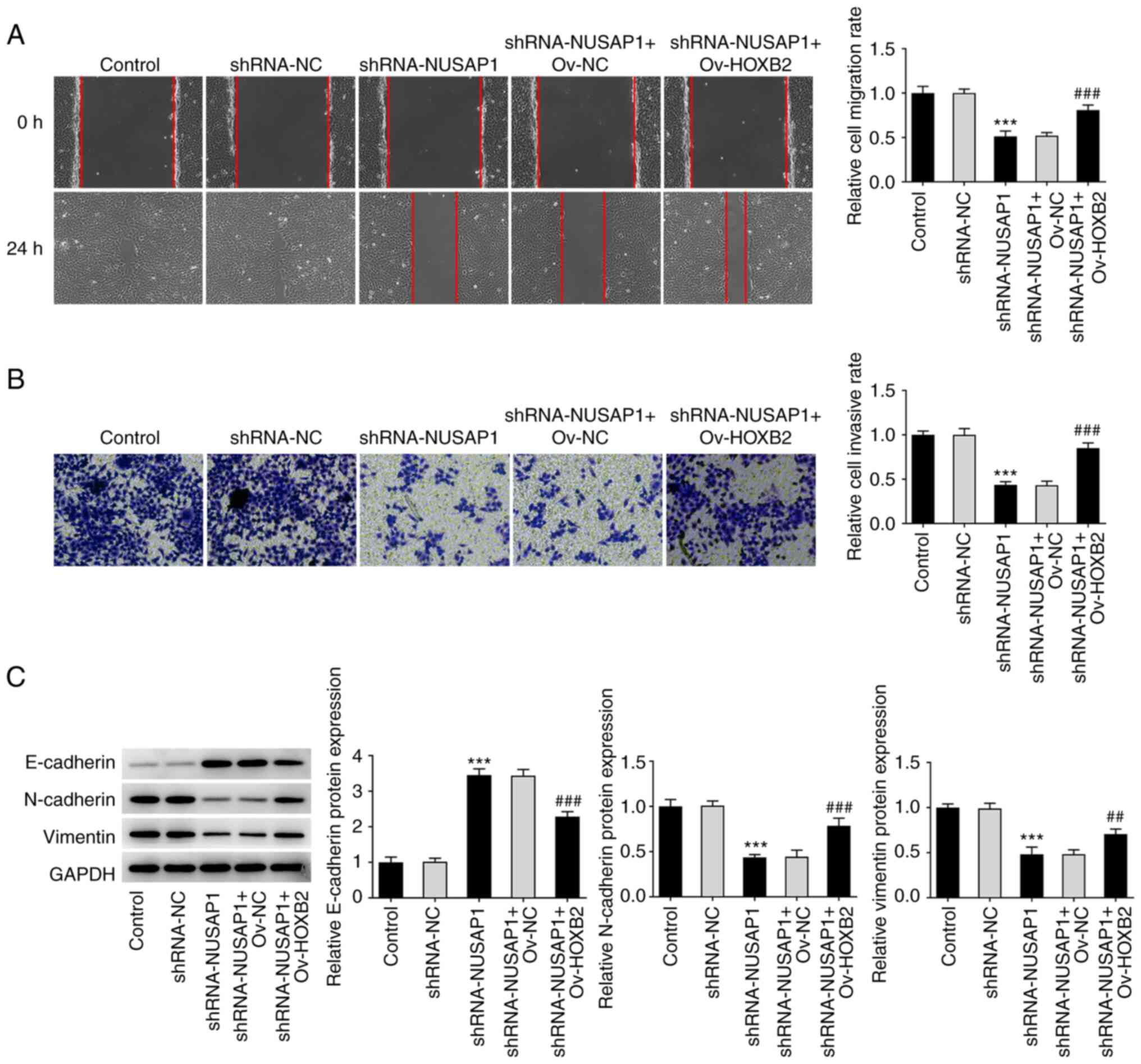
NUSAP1 silencing inhibits the
metastatic potential of nephroblastoma cells
Subsequently, wound healing and Transwell assays
were carried out to evaluate the migratory and invasive abilities
of nephroblastoma cells, respectively. The results demonstrated
that the migratory and invasive abilities of nephroblastoma cells
were notably decreased in the sh-NAUSAP1 group compared with the
sh-NC group (Fig. 2A and B).
Subsequently, western blot analysis was performed to detect the
expression levels of epithelial-to-mesenchymal transition
(EMT)-related proteins. The analysis showed that, compared with
cells transfected with sh-NC, the expression of E-cadherin was
significantly increased, while that of N-cadherin and vimentin was
notably reduced in cells in the sh-NUSAP1 group (Fig. 2C). Therefore, it was hypothesized
that NUSAP1 silencing could inhibit the metastasis of
nephroblastoma cells.
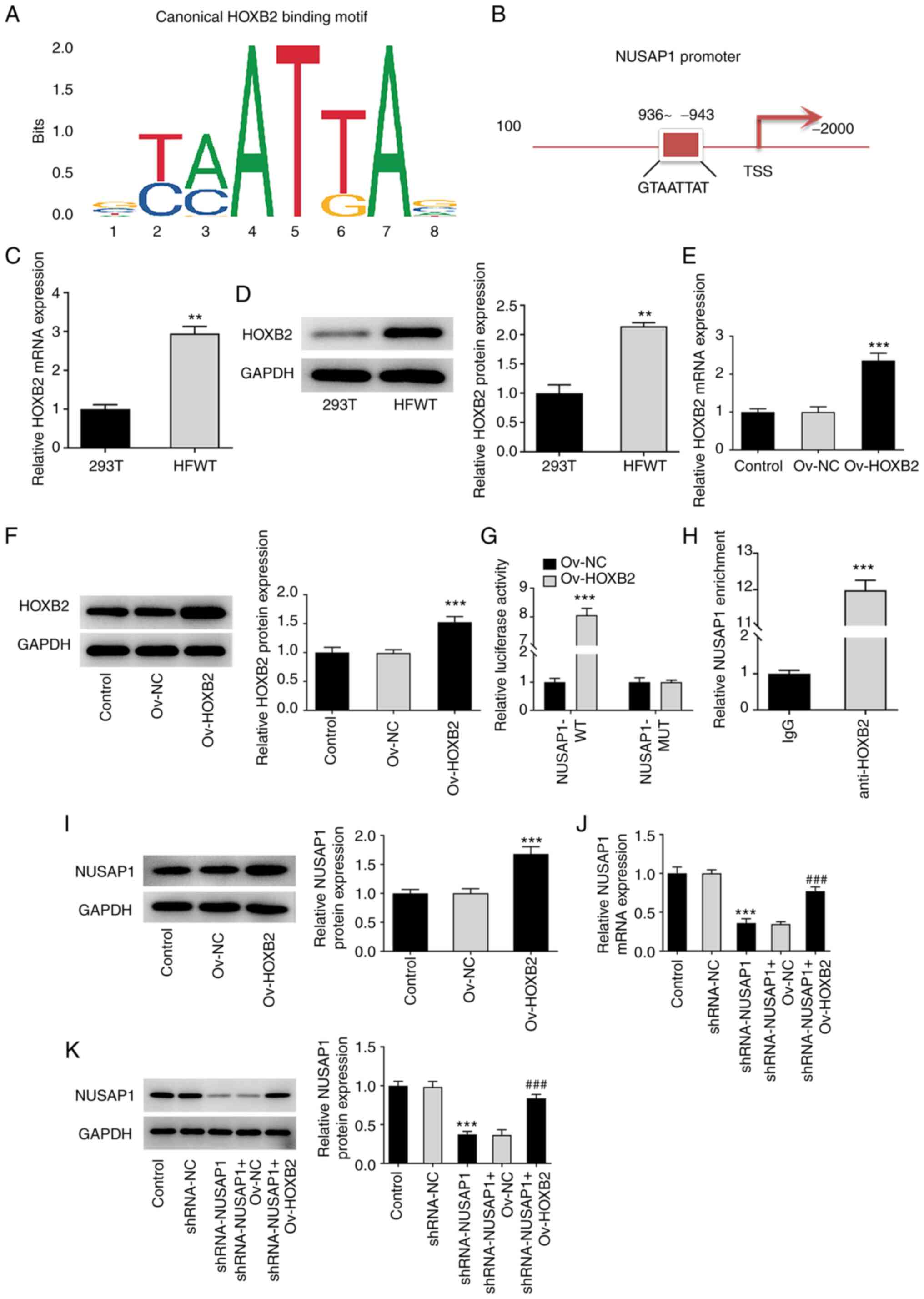
TF HOXB2 activates NUSAP1
Bioinformatics analysis using the JASPAR database
predicted a HOXB2 binding site on the NUSAP1 gene promoter
(Fig. 3A and B). Subsequently,
the expression of HOXB2 in nephroblastoma cells was assessed using
RT-qPCR and western blot analyses. It was observed that HOXB2 was
significantly upregulated in HFWT cells compared with 293T cells
(Fig. 3C and D). Then,
transfection experiments were performed to overexpress HOXB2 and
the transfection efficiency was also determined. HOXB2 was
significantly upregulated in the Ov-HOXB2 group compared with the
Ov-NC group (Fig. 3E and F). The
interaction between HOXB2 and NUSAP1 was verified by
dual-luciferase reporter assay (Fig.
3G), and was further confirmed using a ChIP assay (Fig. 3H). In addition, the expression of
NUSAP1 in the Ov-HOXB2 group was significantly increased after
overexpression of HOXB2 compared with the Ov-NC group (Fig. 3I). Subsequently, cells were
divided into the control, sh-NC, sh-NUSAP1, sh-NUSAP1 + Ov-NC and
sh-NUSAP1 + Ov-HOXB2 groups and the expression of NUSAP1 was then
determined. The results showed that HOXB2 overexpression could
reverse the sh-NUSAP1-mediated reduced NUSAP1 expression (Fig. 3J and K). These findings indicated
that HOXB2 could promote the expression of NUSAP1.
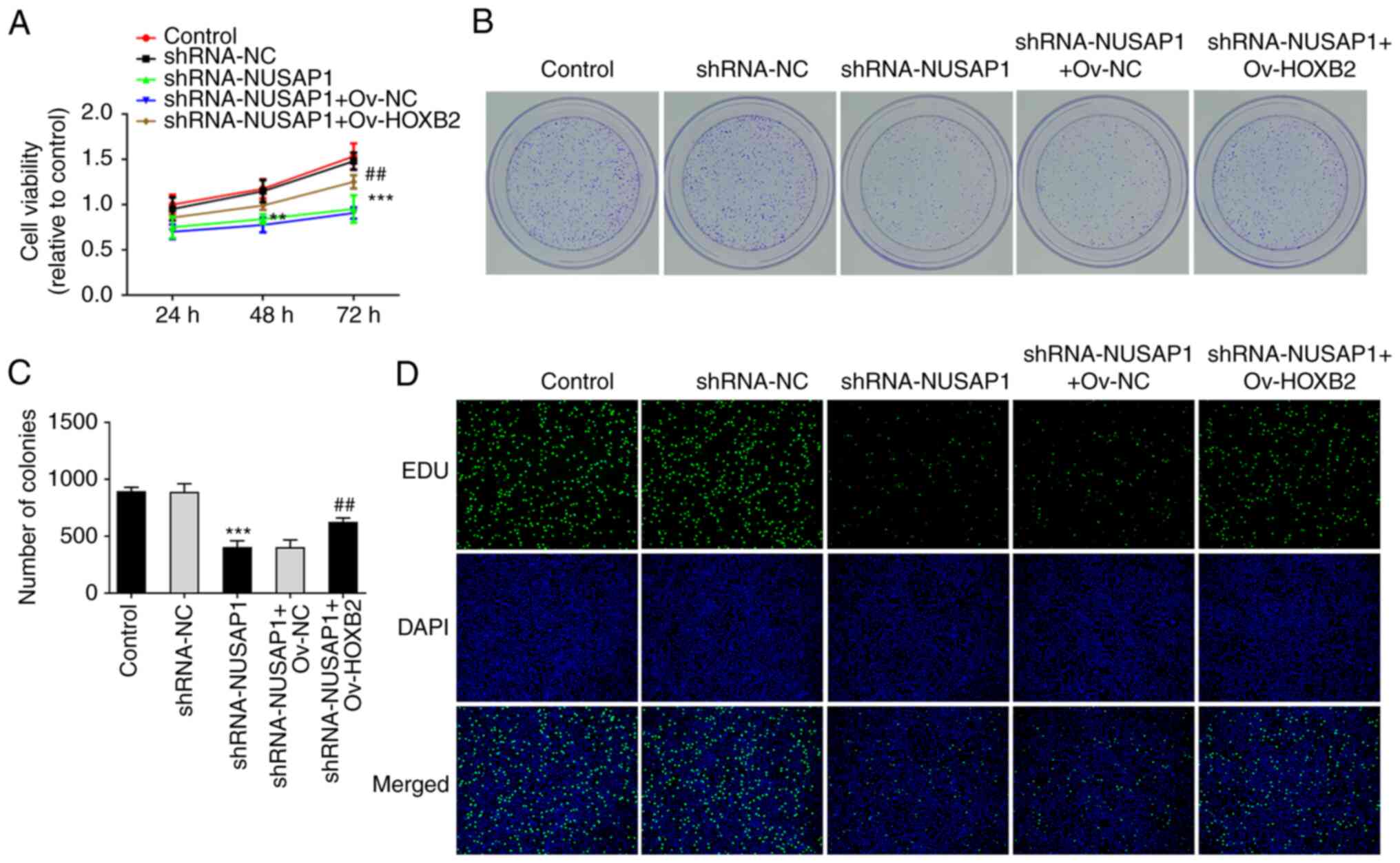
HOXB2 overexpression reverses the
inhibitory effect of NUSAP1 silencing on the proliferation and
metastatic potential of nephroblastoma cells
MTT, EdU and colony formation assays were performed
to evaluate cell proliferation. Therefore, compared with the
sh-NUSAP1 + Ov-NC group, the proliferative ability of
nephroblastoma cells in the sh-NUSAP1 + Ov-HOXB2 group was
significantly enhanced (Fig.
4A-D). Additionally, wound healing and Transwell assays showed
that compared with the sh-NUSAP1 + Ov-NC group, the invasive and
migratory abilities of nephroblastoma cells in the sh-NUSAP1 +
Ov-HOXB2 group were also significantly increased (Fig. 5A and B). To assess the expression
levels of EMT-related proteins, western blot analysis was carried
out. The analysis showed that compared with the sh-NUSAP1 + Ov-NC
group, the expression of E-cadherin was decreased, while that of
N-cadherin and Vimentin was increased in the sh-NUSAP1 + Ov-HOXB2
group (Fig. 5C). These findings
suggested that HOXB2 overexpression could reverse the inhibitory
effect of NUSAP1 knockdown on the proliferation and metastasis of
nephroblastoma cells.
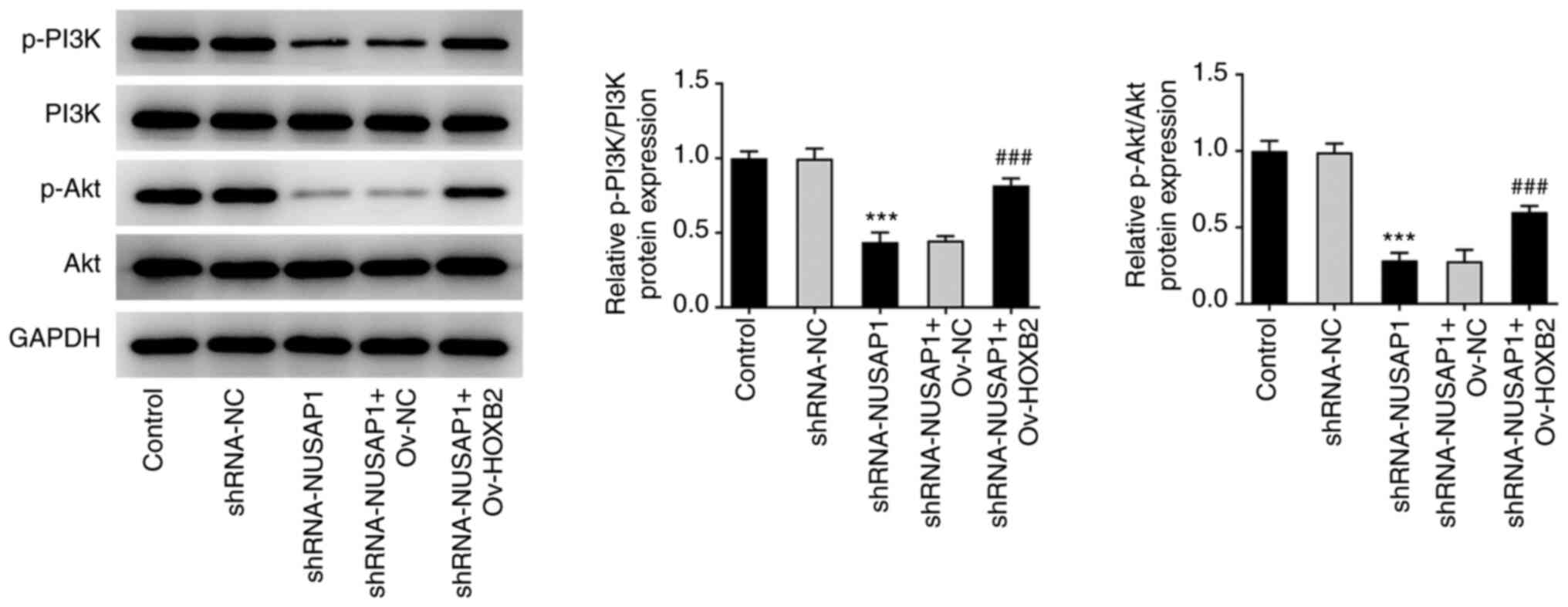
HOXB2 overexpression reverses the
inhibitory effect of NUSAP1 silencing on the proliferation and
metastatic potential of nephroblastoma cells via the PI3K/Akt
signaling pathway
Western blot analysis revealed that the expression
levels of the PI3K/Akt pathway-related proteins were abnormally
altered. Therefore, the expression levels of p-PI3K and p-Akt were
significantly decreased in the sh-NUSAP1 group compared with the
shRNA-NC group. By contrast, p-PI3K and p-Akt were significantly
upregulated in the sh-NUSAP1 + Ov-HOXB2 group compared with the
sh-NUSAP1 + Ov-NC group (Fig. 6).
The aforementioned findings suggested that HOXB2 overexpression
could reverse the inhibitory effect of NUSAP1 silencing on the
proliferation and metastasis of nephroblastoma cells, possibly via
the PI3K/Akt signaling pathway.
Discussion
Nephroblastoma is a common malignant solid tumor of
the abdomen in children, accounting for >90% of all malignant
renal pediatric tumors; improvement of treatment approaches has led
to an increase in the overall survival rate to 85% (17). However, due to tumor metastasis
and recurrence, and the development of resistance to
chemoradiotherapy, some cases remain incurable (18,19). Therefore, it is crucial to
identify novel key biomarkers for nephroblastoma, evaluate the risk
of metastasis in a timely manner, accurately predict patient
survival and improve the overall survival rate through targeted
precision therapies.
NUSAP1 is an essential microtubule-binding protein
in proliferating cells. Abnormal mitotic process during the cell
cycle is one of the most significant causes for the development of
malignant tumors (20). NUSAP1
plays a significant role in mitotic spindle assembly, chromosome
segregation and in the regulation of cell division (21). It has been reported that NUSAP1 is
highly expressed in cancer. A previous study demonstrated that
NUSAP1 was associated with poor prognosis in breast cancer, while
it was considered as a potential biomarker for ductal carcinoma
in situ (22). Based on
database analysis, NUSAP1 expression was significantly upregulated
in bladder cancer stem cells, and NUSAP1 could be used as a
therapeutic target to inhibit the characteristics of bladder cancer
stem cells (23). The abnormal
expression of NUSAP1 in ovarian cancer has good predictive value
for the occurrence and prognosis of ovarian cancer, and can be used
as a candidate target for the diagnosis and treatment of OC
(24). In addition, targeted
NUSAP1 knockdown could attenuate the growth and metastasis of
bladder cancer cells (25).
However, the specific role and mechanism of NUSAP1 in
nephroblastoma have not been previously reported. The results of
the present study demonstrated that the expression of NUSAP1 was
significantly increased in nephroblastoma cell lines. This finding
was consistent with the results reported by Zheng et al
(15). To further evaluate the
effect of NUSAP1 on the proliferative, invasive and migratory
abilities of nephroblastoma cells, the shRNA technology was used to
knock down the expression of NUSAP4. The results showed that NUSAP1
silencing significantly reduced cell viability and proliferation,
as well as the invasive and migratory abilities of nephroblastoma
cells. These results indicated that, inhibiting the expression of
NUSAP1 could significantly attenuate the malignant progression of
nephroblastoma and NUSAP1 can be used as a prognostic marker for
nephroblastoma.
The binding capacity of TF HOXB2 on the NUSAP1
promoter was predicted using the JASPAR website. However, the
effects of HOXB2 and NUSAP1 on nephroblastoma have not been
previously investigated. In the present study, the regulatory
effect of HOXB2 on NUSAP1 was verified via dual-luciferase reporter
and ChIP assays. It has been reported that HOXB2 serves a
regulatory role in tumor development, progression and cell
proliferation via acting as a TF to regulate the expression of
downstream factors (12). A
previous in vivo study on the functional screening of tumor
progression regulators revealed that HOXB2 could regulate the
development of breast cancer (26). Another study demonstrated that
HOXB2 promoted the invasion of HOP-62 non-small cell lung cancer
(NSCLC) cells via transcriptionally regulating the expression of
metastasis-related genes (27).
In addition, HOXB2 promoted glial cell proliferation and invasion
in vitro in 1,001 patients with glioma, and it was
positively correlated with tumor grade and established as an
independent prognostic biomarker (28). However, the effects of HOXB2 and
NUSAP1 on the proliferation, invasion and migration of
nephroblastoma cells remain unclear. The current study demonstrated
that HOXB2 overexpression reversed the inhibitory effect of NUSAP1
knockdown on the proliferation and metastasis of nephroblastoma
cells.
In the present study, the activation of the PI3K/Akt
signaling pathway was inhibited following NUSAP1 silencing in
nephroblastoma cells. However, this effect was reversed after HOXB2
overexpression. Therefore, it was hypothesized that HOXB2
overexpression could abrogate the inhibitory effect of NUSAP1
knockdown on the proliferation and metastasis of nephroblastoma
cells via the PI3K/Akt signaling pathway. Consistent with the
aforementioned findings, Li et al (29) showed that the expression levels of
p-PI3K and p-Akt were decreased in glioblastoma cells following
HOXB2 knockout. These results suggested that HOXB2 could promote
the activation of PI3K/Akt signaling. In addition, NUSAP1 knockdown
could also inhibit the activation of B cell translocation gene
2/PI3K/Akt signaling, thus attenuating cell proliferation and
metastasis in NSCLC (2,30). This finding was consistent with
the results of the current study.
However, there were certain limitations of the
present study. First, the mechanism underlying the involvement of
the PI3K/Akt signaling pathway was not further investigated.
Second, only in vitro experiments in nephroblastoma cell
lines were performed. Therefore, in vivo experiments are
needed to further verify the results of the present study. These
experiments will be performed in our future studies. In addition,
we will further detect the influence of HOXB2/NUSAP1 on cell
resistance caused by cisplatin or radiotherapy drugs in the next
experiment.
In conclusion, the findings of the present study
demonstrated that the TF HOXB2 could upregulate NUSAP1 to promote
the proliferation, invasion and migration of nephroblastoma cells
via the PI3K/Akt signaling pathway, thereby possibly providing
novel targets for the diagnosis and treatment of
nephroblastoma.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets generated and/or analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
BL and SF wrote the manuscript and analyzed the
data. TL, JW, ZQ, YZ and BH performed the experiments and
supervised the study. ZQ, YZ and BH searched the literature and
revised the manuscript for important intellectual content. BL and
SF confirm the authenticity of all the raw data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Charlton J, Irtan S, Bergeron C and
Pritchard-Jones K: Bilateral Wilms tumour: A review of clinical and
molecular features. Expert Rev Mol Med. 19:e82017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dome JS, Perlman EJ and Graf N: Risk
stratification for wilms tumor: current approach and future
directions. Am Soc Clin Oncol Educ Book. 215–223. 2014. View Article : Google Scholar
|
|
3
|
Dumba M, Jawad N and McHugh K:
Neuroblastoma and nephroblastoma: A radiological review. Cancer
Imaging. 15:52015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Szychot E, Apps J and Pritchard-Jones K:
Wilms' tumor: Biology, diagnosis and treatment. Transl Pediatr.
3:12–24. 2014.PubMed/NCBI
|
|
5
|
Maschietto M, Piccoli FS, Costa CM,
Camargo LP, Neves JI, Grundy PE, Brentani H, Soares FA, de Camargo
B and Carraro DM: Gene expression analysis of blastemal component
reveals genes associated with relapse mechanism in Wilms tumour.
Eur J Cancer. 47:2715–2722. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Francois M, Donovan P and Fontaine F:
Modulating transcription factor activity: Interfering with
protein-protein interaction networks. Semin Cell Dev Biol.
99:12–19. 2020. View Article : Google Scholar
|
|
7
|
Teschendorff AE, Zheng SC, Feber A, Yang
Z, Beck S and Widschwendter M: The multi-omic landscape of
transcription factor inactivation in cancer. Genome Med. 8:892016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Holland PW: Evolution of homeobox genes.
Wiley Interdiscip Rev Dev Biol. 2:31–45. 2013. View Article : Google Scholar
|
|
9
|
Yu HY and Pan SS: MiR-202-5p suppressed
cell proliferation, migration and invasion in ovarian cancer via
regulating HOXB2. Eur Rev Med Pharmacol Sci. 24:2256–2263.
2020.
|
|
10
|
Du H, Bao Y, Liu C, Zhong A, Niu Y and
Tang X: MiR1395p enhances cisplatin sensitivity in nonsmall cell
lung cancer cells by inhibiting cell proliferation and promoting
apoptosis via the targeting of Homeobox protein HoxB2. Mol Med Rep.
23:1042021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jing P, Zou J, Zhang L, Wang C, Yang Y,
Deng L and Zhao D: HOXB2 and FOXC1 synergistically drive the
progression of Wilms tumor. Exp Mol Pathol. 115:1044692020.
View Article : Google Scholar
|
|
12
|
Raemaekers T, Ribbeck K, Beaudouin J,
Annaert W, Van Camp M, Stockmans I, Smets N, Bouillon R, Ellenberg
J and Carmeliet G: NuSAP, a novel microtubule-associated protein
involved in mitotic spindle organization. J Cell Biol.
162:1017–1029. 2003. View Article : Google Scholar
|
|
13
|
Zhou Q, Lee KJ, Kurasawa Y, Hu H, An T and
Li Z: Faithful chromosome segregation in Trypanosoma brucei
requires a cohort of divergent spindle-associated proteins with
distinct functions. Nucleic Acids Res. 46:8216–8231. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Z, Guan C, Lu C, Liu Y, Ni R, Xiao M
and Bian Z: High NUSAP1 expression predicts poor prognosis in colon
cancer. Pathol Res Pract. 214:968–973. 2018. View Article : Google Scholar
|
|
15
|
Zheng H, Li BH, Liu C, Jia L and Liu FT:
Comprehensive analysis of lncRNA-Mediated ceRNA crosstalk and
identification of prognostic biomarkers in Wilms' tumor. Biomed Res
Int. 2020:49516922020.
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cunningham ME, Klug TD, Nuchtern JG,
Chintagumpala MM, Venkatramani R, Lubega J and Naik-Mathuria BJ:
Global disparities in Wilms tumor. J Surg Res. 247:34–51. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Aghaalikhani N, Rashtchizadeh N, Shadpour
P, Allameh A and Mahmoodi M: Cancer stem cells as a therapeutic
target in bladder cancer. J Cell Physiol. 234:3197–3206. 2019.
View Article : Google Scholar
|
|
19
|
Barbato L, Bocchetti M, Di Biase A and
Regad T: Cancer stem cells and targeting strategies. Cells.
8:9262019. View Article : Google Scholar
|
|
20
|
Liu X, Chen Y, Li Y, Petersen RB and Huang
K: Targeting mitosis exit: A brake for cancer cell proliferation.
Biochim Biophys Acta Rev Cancer. 1871:179–191. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mills CA, Suzuki A, Arceci A, Mo JY,
Duncan A, Salmon ED and Emanuele MJ: Nucleolar and
spindle-associated protein 1 (NUSAP1) interacts with a SUMO E3
ligase complex during chromosome segregation. J Biol Chem.
292:17178–17189. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun L, Shi C, Liu S, Zhang E, Yan L, Ji C
and Zhao Y: Overexpression of NuSAP1 is predictive of an
unfavourable prognosis and promotes proliferation and invasion of
triple-negative breast cancer cells via the Wnt/β-catenin/EMT
signalling axis. Gene. 747:1446572020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pan S, Zhan Y, Chen X, Wu B and Liu B:
Identification of biomarkers for controlling cancer stem cell
characteristics in bladder cancer by network analysis of
transcriptome data stemness indices. Front Oncol. 9:6132019.
View Article : Google Scholar
|
|
24
|
Shen J, Yu S, Sun X, Yin M, Fei J and Zhou
J: Identification of key biomarkers associated with development and
prognosis in patients with ovarian carcinoma: Evidence from
bioinformatic analysis. J Ovarian Res. 12:1102019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen Y, Zhang W, Kadier A, Zhang H and Yao
X: MicroRNA-769-5p suppresses cell growth and migration via
targeting NUSAP1 in bladder cancer. J Clin Lab Anal.
34:e231932020.
|
|
26
|
Boimel PJ, Cruz C and Segall JE: A
functional in vivo screen for regulators of tumor progression
identifies HOXB2 as a regulator of tumor growth in breast cancer.
Genomics. 98:164–172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Inamura K, Togashi Y, Ninomiya H, Shimoji
T, Noda T and Ishikawa Y: HOXB2, an adverse prognostic indicator
for stage I lung adenocarcinomas, promotes invasion by
transcriptional regulation of metastasis-related genes in HOP-62
non-small cell lung cancer cells. Anticancer Res. 28((4B)):
2121–2127. 2008.PubMed/NCBI
|
|
28
|
Pan X, Liu W, Chai Y, Wang J and Zhang Y:
Genetic and clinical characterization of HOXB2 in glioma. Onco
Targets Ther. 13:10465–10473. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li M, Wang JF, Liu B and Wang XM: Homeobox
B2 is a potential prognostic biomarker of glioblastoma. Rev Assoc
Med Bras (1992). 66:794–799. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xu Z, Wang Y, Xiong J, Cui F, Wang L and
Peng H: NUSAP1 knockdown inhibits cell growth and metastasis of
non-small-cell lung cancer through regulating BTG2/PI3K/Akt
signaling. J Cell Physiol. 235:3886–3893. 2020. View Article : Google Scholar
|