Introduction
Colorectal cancer (CRC) is a highly prevalent
malignant cancer. It is the third most commonly diagnosed type of
cancer and causes the second highest rate of mortality from cancer
worldwide (1,2). It was recently indicated that 10% of
new cancer cases globally were due to CRC and that CRC led to 9.4%
of the mortalities due to cancer (1,3). In
China, there has been an increasing trend in the incidence of CRC
for from 2000 to 2020 (4). Early
diagnosis of CRC is important for the survival of patients. It was
revealed that the 5-year survival rate for patients with CRC was up
to 90% when diagnosed before metastasis took place, while the
5-year survival rate for patients with CRC with distant metastases
was only ~12% (5). Although
invasive colonoscopy is a popular method used for the detection of
CRC, it can result in false negative colonoscopy findings in a
number of patients (6). Therefore,
more accurate and less invasive screening methods are needed for
CRC detection. The risk of CRC carcinogenesis is multifactorial and
may be associated with genetic and epigenetic genomic alterations,
age, chronic inflammation, as well as environmental and lifestyle
factors (7). During cancer
initiation and progression, the cancer cells undergo a metabolic
reprogramming, such as an altered energy metabolism and an
increased biosynthesis of macromolecules (8). The cellular metabolism regulates the
epigenetic characteristics of the tumor cells and these cells
interact with their surrounding microenvironment. These metabolic
interactions are necessary in the regulation of tumor progression
(9).
Metabolomics is a branch of systematic biology that
examines the changes of metabolites in numerous types of cancer and
other diseases (10). Metabolomics
offers understanding of carcinogenesis and potentially less
invasive methods of cancer diagnosis (11). Different specimens, including
serum, plasma, urine and stool collected from patients with CRC,
are commonly used in CRC metabolomic studies (11–13).
Alterations of multiple metabolites, including metabolites of
carbohydrates, amino acids, lipids, nucleotides and hormones, were
revealed in these CRC specimens. However, these results mainly
reflected indirect metabolic alterations in CRC cells (11,13,14).
By contrast, analysis of the CRC tissue could provide direct
metabolic profiling dates, and thus reflect higher specificity to
the metabolic consequences of the disease. This would eliminate the
non-tumor and/or body responses detected in the serum or other
specimens (15–17). It has been reported that the
alteration of metabolites in CRC tissue was implicated in energy
metabolism, amino acid metabolism, glutathione metabolism and fatty
acid metabolism and these observed metabolic changes were relevant
to the anatomical location and prognosis of CRC (15,16,18).
In the present study, ultra-high-performance liquid
chromatography (UHPLC)-mass spectrometry (MS)-based comprehensive
metabolomics analysis of metabolic profiles was performed on
resected cancer tissues from patients with CRC. Marked changes were
revealed in a number of metabolites that are involved in different
pathways in CRC. The present study showed that a number of
metabolites, which have been associated with tumorigenesis and may
provide insight for the clinical management of CRC.
Materials and methods
Patients and tissue samples
The paired CRC tissues and adjacent normal tissues
(5 cm away from the tumor margin) were collected from patients with
CRC undergoing surgical treatment in Beijing Rehabilitation
Hospital of Capital Medical University (Beijing, China) from June
2020 to January 2022. Metabolomic analyses were performed on these
35 paired samples. The characteristics of the patients with CRC
have been presented in Table I.
The patients had not received any radio- or chemotherapy before
surgery. The present study was approved by the ethics committee of
Beijing Rehabilitation Hospital of Capital Medical University
(Beijing, China) and all participants signed written informed
consent.
 | Table I.Demographics of patients with
colorectal cancer (n=35). |
Table I.
Demographics of patients with
colorectal cancer (n=35).
|
Characteristics | Value |
|---|
| Sex, n (%) |
|
|
Male | 23 (65.7) |
|
Female | 12 (34.2) |
| Age, years (mean ±
SD) | 70.0±14.0 |
| Anatomic location
in the colon, n (%) |
|
| Right
side | 10 (28.6) |
| Left
side | 13 (37.1) |
|
Rectum | 12 (34.2) |
| Disease stage, n
(%) |
|
| I | 6 (17.1) |
| II | 14 (37.1) |
|
III | 9 (22.9) |
| IV | 6 (17.1) |
| Tumor
differentiation, n (%) |
|
|
Poor | 5 (14.2) |
|
Moderate | 25 (71.4) |
|
Good | 5 (14.2) |
Tissue procurement
The tissue sample preparation and metabolite
extraction was conducted according to the methods described by
Romisch-Margl et al (19).
After surgical resection, tissue samples were immediately stored at
−80°C until used for metabolomics analyses. Tissue samples (20 µg)
were thawed on ice for 30 min. After homogenization, the metabolite
extraction procedure was carried out as follows: A total of 400 µl
70% methanol-water and internal standard extractant (Table II) were added to each sample, the
mixture was vortexed for 2 min, frozen in liquid nitrogen for 5
min, placed on dry ice for 5 min, and then thawed on ice for 5 min.
After the sample was vortexed for 2 min, it was inverted three
times, then centrifuged at 16,113.6 × g at 4°C for 10 min.
Subsequently, 300 µl of the supernatant was transferred to a new
tube and stored at −20°C for 30 min to precipitate proteins to
avoid any interference of the proteins in the analytic procedure.
After final centrifugation at 16,113.6 × g at 4°C for 3 min, 200 µl
supernatant was collected for analysis by Allwegene Technology Co.,
Ltd. The molecules of internal standards are presented in Table II.
| Table II.Molecules used as internal
standards. |
Table II.
Molecules used as internal
standards.
| Name | CAS no. | Concentration |
|---|
|
L-2-chlorophenylalanine | 103616-89-3 | 1 µg/ml |
|
(2H3)-L-Carnitine HCl | 350818-62-1 | 1 µg/ml |
|
4-Fluoro-L-α-phenylglycine | 19883-57-9 | 1 µg/ml |
| L-Phenylalanine
(2–13C, 99%) | 63-91-2 | 1 µg/ml |
|
(2H5)-Hippuric acid | 53518-98-2 | 1 µg/ml |
|
(2H5)-Kynurenic acid | 350820-13-2 | 1 µg/ml |
|
(2H5)-Phenoxy acetic acid | 154492-74-7 | 1 µg/ml |
UHPLC-MS/MS analysis
UHPLC chromatographic separation was performed using
an EXIONLC System (SCIEX) installed with a Waters Acquity UHPLC HSS
T3 1.8 µm 2.1×100 mm column (Waters Corporation). The details of
the internal standard are provided in Table II. The mobile phase A consisted of
water (and 0.04% acetic acid), and the mobile phase B consisted of
acetonitrile (and 0.04% acetic acid). The following gradient
program was used: 98% A, 0 min; 5% A, 11 min; 5% A, 12 min; 5% A,
12.1 min; 95% A, 14 min; and 95% A, 15 min. The flow rate was set
to 0.5 ml/min. The injection volume was 2 µl, and the samples were
maintained at 4°C in the autosampler.
Linear ion trap (LIT) and triple quadrupole (QQQ)
scans were acquired on a triple quadrupole-linear ion trap mass
spectrometer (QTRAP), QTRAP® LC-MS/MS System (SCIEX),
equipped with an electrospray ionization (ESI) Turbo Ion-Spray
interface, operating in positive and negative ion mode and
controlled by Analyst software (version 1.6.3; SCIEX). The ESI
source operation parameters were as follows: Source temperature
500°C; ion spray voltage 5,500 V (positive), −4,500 V (negative);
ion source gas I, ion source gas II and curtain gas were set to 55,
60 and 25 psi, respectively; the collision gas parameter was set at
high. Instrument tuning and mass calibration were performed with 10
and 100 µmol/l polypropylene glycol solutions in QQQ and LIT modes,
respectively. A specific set of multiple reaction monitoring (MRM)
transitions were monitored for each period according to the
metabolites eluted within this period. Briefly, in MRM mode, the
instrument first selected a parent ion of the target molecule based
on specific mass/change ratio (m/z) values, and any initial
interference were removed. The parent ion then undergoes
collision-induced dissociation. The resultant fragment ions are
then filtered through the triple four-pole filter to select the
characteristic fragment ions, eliminating non-target ion
interference, making the quantification more accurate and improving
repeatability.
Data analysis
Based on the metabolome standard database
(AllwegeneDB; Allwegene Technology Co., Ltd., qualitative analysis
was carried out according to the retention time, parent ion pair
information and secondary spectrum data. After obtaining the data
of all the tissue samples in the present study, the mass spectrum
files of the samples were opened with multiquant software (version
3.0.3; Shanghai AB SCIEX Analytical Instrument Trading Co.) to
carry out the integration and correction of the chromatographic
peaks. The peak area of each chromatographic peak represents the
relative content of the corresponding substance. All of the
chromatographic peak area integration data were exported and
saved.
Principal component analysis (PCA) was performed
using the statistics function prcomp within R (version 3.5.1;
www.r-project.org). The data was unit variance
scaled before unsupervised PCA. The hierarchical cluster analysis
(HCA) results of samples and metabolites were presented as heatmaps
with dendrograms, while Pearson correlation coefficients (PCC)
between samples were calculated by the cor function in R and
presented only as heatmaps. Both HCA and PCC were carried out using
the R package ComplexHeatmap (version 1.20.0) (20). For HCA, the normalized signal
intensities of the metabolites (unit variance scaling) were
visualized as a color spectrum. Significantly regulated metabolites
between groups were determined by variable important in projection
(VIP) ≥1 and absolute Log2 fold change (FC) ≥1. VIP values were
extracted from the orthogonal partial least-squares discriminant
analysis (OPLS-DA) results, which also contain score plots and
permutation plots that were generated using the R package
MetaboAnalystR (version 3.5.1; http://www.metaboanalyst.ca/) (21). The data were log transformed (log2)
and mean centered before OPLS-DA. In order to avoid overfitting, a
permutation test (200 permutations) was performed. MetaboAnalystR
was also used to analyze the significantly different levels of the
metabolites between the cancer and adjacent tissues, in which a
paired Student's t-test was applied.
Identified metabolites were annotated using the
Kyoto Encyclopedia of Genes and Genomes (KEGG) Compound database
(http://www.kegg.jp/kegg/compound/),
and annotated metabolites were then mapped to the KEGG Pathway
database (http://www.kegg.jp/kegg/pathway.html). Significantly
enriched pathways were identified according to hypergeometric test
P-value. All differential metabolites in the present study were
examined in PubMed (https://pubmed.ncbi.nlm.nih.gov) and The Human
Metabolome Database (https://hmdb.ca/) in order to
identify them in previously published studies.
Statistical analysis
With SPSS (version 20; IBM Corp.), statistical
analysis was performed for the comparisons of the differential
metabolites between cancer tissue and sex, grade of
differentiation, tumor location and tumor stage in each group.
Non-parametric tests were used to analyze the data of each group
after the normality of data was assessed using the Shapiro-Wilk
test. The Mann-Whitney U test was used to assessed the different
metabolite levels between male and female patients. A
Kruskal-Wallis test was used to compare the data among three or
four subgroups of patients depending on the grade of
differentiation, tumor location and tumor stage, and the Dunn's
post hoc test was used for comparison among the subgroups when
statistical difference was reached after the Kruskal-Wallis test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Metabolic differences between CRC and
normal colon tissues
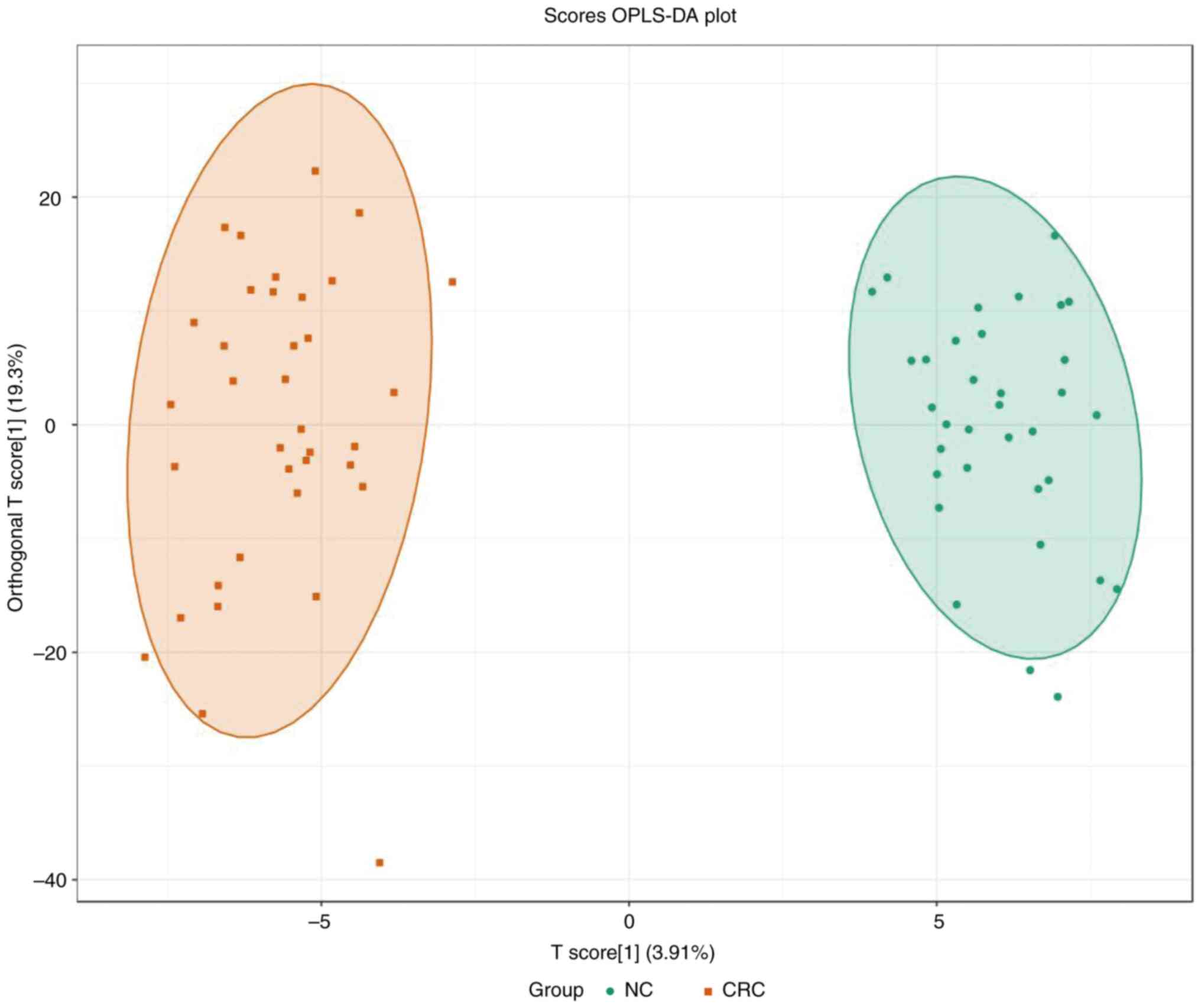
PCA was performed to identify the differences
between the paired CRC tissue and normal adjacent colon tissue,
which was collected from patients with CRC (Fig. 1). A separation tendency between the
normal and the CRC tissues indicated differences in the levels of
the metabolites between the two groups. A total of 927 metabolites
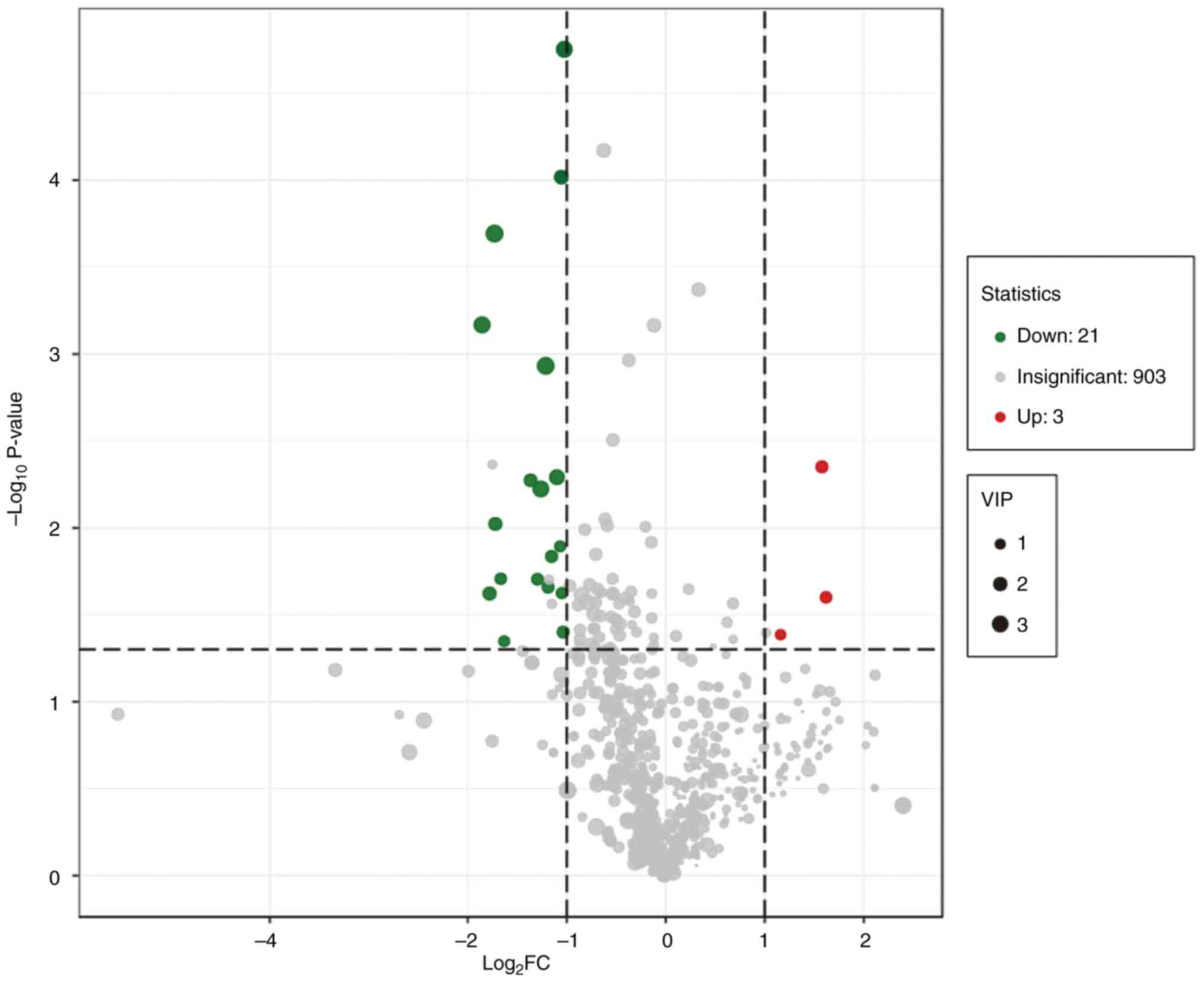
were detected. Using a univariate analysis, 24 differential
metabolites were identified, which had a FC of either ≥2 or <0.5
between normal and CRC tissue (P<0.05). Among them, the levels
of three metabolites were upregulated and 21 were downregulated in
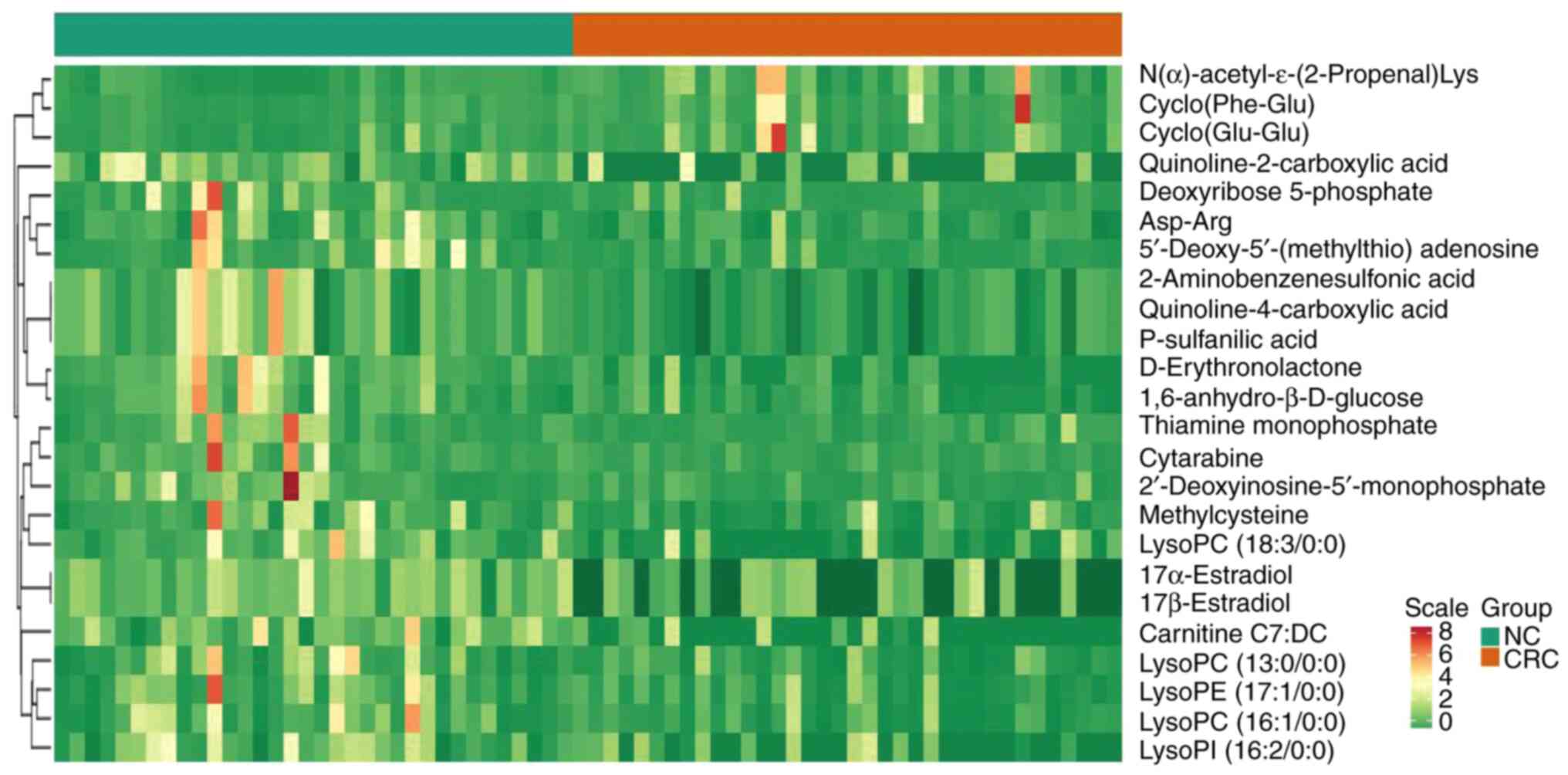
CRC tissue (Figs. 2 and 3 and Table
III). In Fig. 3, the heatmap
indicated the changes of all 24 differential metabolites in the 35
patients included in the present study.
| Table III.Differential metabolites in
colorectal cancer tissue vs. adjacent normal tissues. |
Table III.
Differential metabolites in
colorectal cancer tissue vs. adjacent normal tissues.
| Compounds | Class | VIP | P-value | Fold change |
|---|
| Cyclo(Glu-Glu) | Cyclodipeptide | 1.14 | 0.041 | 2.24 |
| Cyclo(Phe-Glu) | Cyclodipeptide | 1.50 | 0.025 | 3.07 |
|
N-α-acetyl-ε-(2-propenal)-Lys | Amino acid | 1.63 | 0.004 | 2.98 |
|
Quinoline-2-carboxylic acid | Amino acid | 3.51 | 0.001 | −2.33 |
|
Quinoline-4-carboxylic acid | Amino acid | 1.99 | 0.001 | −2.08 |
|
D-Erythronolactone | Carbohydrate | 3.20 | 0.001 | −3.57 |
|
1,6-anhydro-β-D-glucose | Carbohydrate | 1.83 | 0.005 | −2.56 |
| Asp-Arg | Dipeptide | 1.31 | 0.013 | −2.08 |
| Methylcysteine | Amino acid | 1.60 | 0.022 | −2.27 |
| P-sulfanilic
acid | Amino acid | 1.99 | <0.001 | −2.08 |
| Cytarabine | Nucleotide | 2.00 | 0.024 | −3.45 |
|
5′-Deoxy-5′-(methylthio) adenosine | Nucleotide | 1.92 | 0.009 | −3.33 |
| Deoxyribose
5-phosphate | Nucleotide | 1.45 | 0.020 | −3.23 |
|
2′-Deoxyinosine-5′-monophosphate | Nucleotide | 1.28 | 0.045 | −3.13 |
| Carnitine
C7:DC | Lipid | 3.52 | <0.001 | −3.33 |
| LysoPC
(18:3/0:0) | Lipid | 3.06 | 0.006 | −2.38 |
| LysoPC
(16:1/0:0) | Lipid | 1.66 | 0.015 | −2.22 |
| LysoPC
(13:0/0:0) | Lipid | 1.40 | 0.024 | −2.08 |
| LysoPI
(16:2/0:0) | Lipid | 2.60 | 0.005 | −2.13 |
| LysoPE
(17:1/0:0) | Lipid | 1.60 | 0.040 | −2.04 |
| Thiamine
monophosphate | Heterocyclic
compound | 1.52 | 0.020 | −2.44 |
| 17α-Estradiol | Hormone | 2.90 | <0.001 | −2.04 |
| 17β-Estradiol | Hormone | 2.90 | <0.001 | −2.04 |
|
2-Aminobenzenesulfonic acid | Benzene
derivatives | 1.99 | <0.001 | −2.08 |
Without stratification, the levels of two
carbohydrate metabolites, D-erythronolactone and
1,6-anhydro-β-D-glucose, were decreased in CRC tissue compared with
the levels in normal tissue. The levels of the amino acid
metabolite, N-α-acetyl-ε-(2-propenal)-Lys, and two cyclodipeptides
(CDPs), cyclo(Glu-Glu) and cyclo(Phe-Leu), were increased in CRC
tissue, while other amino acid metabolites, quinaldic acid (also
referred to quinoline-2-carboxylic acid) and quinoline-4-carboxylic
acid were decreased in CRC tissue compared with the levels in
normal tissue. The results demonstrated that the levels of
nucleotide metabolites were consistently deceased in CRC tissue
compared with normal tissue. These included cytarabine,
5′-deoxy-5′-(methylthio) adenosine, deoxyribose 5-phosphate and
2′-deoxyinosine-5′-monophosphate. Similarly, lipid metabolites,
such as carnitine, phosphatidylcholine (PC) derivative lysoPC,
including two sub-types lysophosphatidylethanolamine (lysoPE) and
lysophosphatidylinositol, were decreased in the CRC tissue compared
with normal tissue. Furthermore, the levels of 17α- and
17β-estradiol were lower in CRC tissue compared with normal tissue.
Lastly, the level of thiamine monophosphate, a phosphorylated form
of thiamine (vitamin B1) that is required to maintain sodium and
potassium gradients for conducting nerve impulses in the central
nervous system (22), was
decreased in CRC tissue compared with normal tissue (Table III).
The 24 differential metabolites identified in the
present study were examined in public databases, which revealed
that 11 of the metabolites had been reported previously (Table IV). Among the 11 metabolites, six
were revealed in human CRC tissues, and seven in other human
samples, such as plasma, urine or stool samples. A total of four
metabolites were reported in mouse samples, including tumor tissue,
plasma or stool samples.
| Table IV.Comparison of the differential
metabolites identified in the present study with those reported in
other published reports. |
Table IV.
Comparison of the differential
metabolites identified in the present study with those reported in
other published reports.
|
| Patient studies
(Refs.) | Animal studies
(Refs.) |
|---|
|
|
|
|
|---|
| Compounds | CRC tissues | Other samples |
Samplesa |
|---|
| Asp-Arg | (56) | - | - |
| Carnitine
C7:DC | (57–60) | (61)b,(62,63)c | - |
| LysoPC
(18:3/0:0) | (56,64,65) | (66)b | - |
| LysoPC
(16:1/0:0) | (58,65,67,68) | (69,70)b | - |
| LysoPE
(17:1/0:0) | (65) | - | - |
| Methylcysteine | - | (71)c | - |
|
5′-Deoxy-5′-(methylthio) adenosine | - | (26)d | - |
| P-sulfanilic
acid | (72) | - | (73) |
|
1,6-anhydro-β-D-glucose | - | (26)d | (74,75) |
|
N-α-acetyl-ε-(2-propenal)-Lys | - | (76)b,(61)d | (57,77)b,(75) |
| Deoxyribose
5-phosphate | - | - | (78)d |
Comparison of the differential
metabolites based on sex, anatomic location, stage and grade of
differentiation
No statistical differences were revealed when
comparing the changes in the levels of the metabolites based on sex
or on the grade of tumor differentiation (data not shown). The
levels of the metabolites were subsequently compared based on the
anatomic location of the tumors and it was revealed that the right
side of the colon had higher levels of lysoPE (17:1/0:0) (10.45
fold) compared with the left side of the colon (0.56 fold) and
rectum (1.07 fold) (P<0.05), there was no difference in the
level of lysoPE (17:1/0:0) between the left side of the colon and
the rectum (data not shown). The present study subsequently
analyzed the levels of the metabolites based on disease stage, and
six metabolites were revealed to have different levels at different
stages (Table V). In stage I, the
levels of three metabolites, 2-aminobenzenesulfonic acid,
P-sulfanilic acid and quinoline-4-carboxylic acid, were lower
compared with the other stages, while N-α-acetyl-ε-(2-propenal)-Lys
was higher in stage III compared with the other stages. The levels
of methylcysteine and 5′-deoxy-5′-(methylthio) adenosine varied at
the different stages of tumorigenesis.
| Table V.Differential metabolite changes in
colorectal cancer at different stages. |
Table V.
Differential metabolite changes in
colorectal cancer at different stages.
|
| Fold change in
different stages |
|---|
|
|
|
|---|
| Metabolite
name | I | II | III | IV |
|---|
|
2-Aminobenzenesulfonic acid | 0.26a | 0.85 | 0.83 | 0.67 |
| P-sulfanilic
acid | 0.26a | 0.85 | 0.83 | 0.67 |
|
Quinoline-4-carboxylic acid | 0.28a | 0.47 | 0.74 | 0.64 |
|
N-α-acetyl-ε-(2-propenal)-Lys | 11.02a | 1.82 | 23.28b | 3.00 |
| Methylcysteine | 0.10b | 143.45c | 0.46 | 1.17 |
|
5′-Deoxy-5′-(methylthio) adenosine | 0.15d | 0.99 | 2.38 | 1.32 |
Identification of distinct metabolites
in enriched pathways
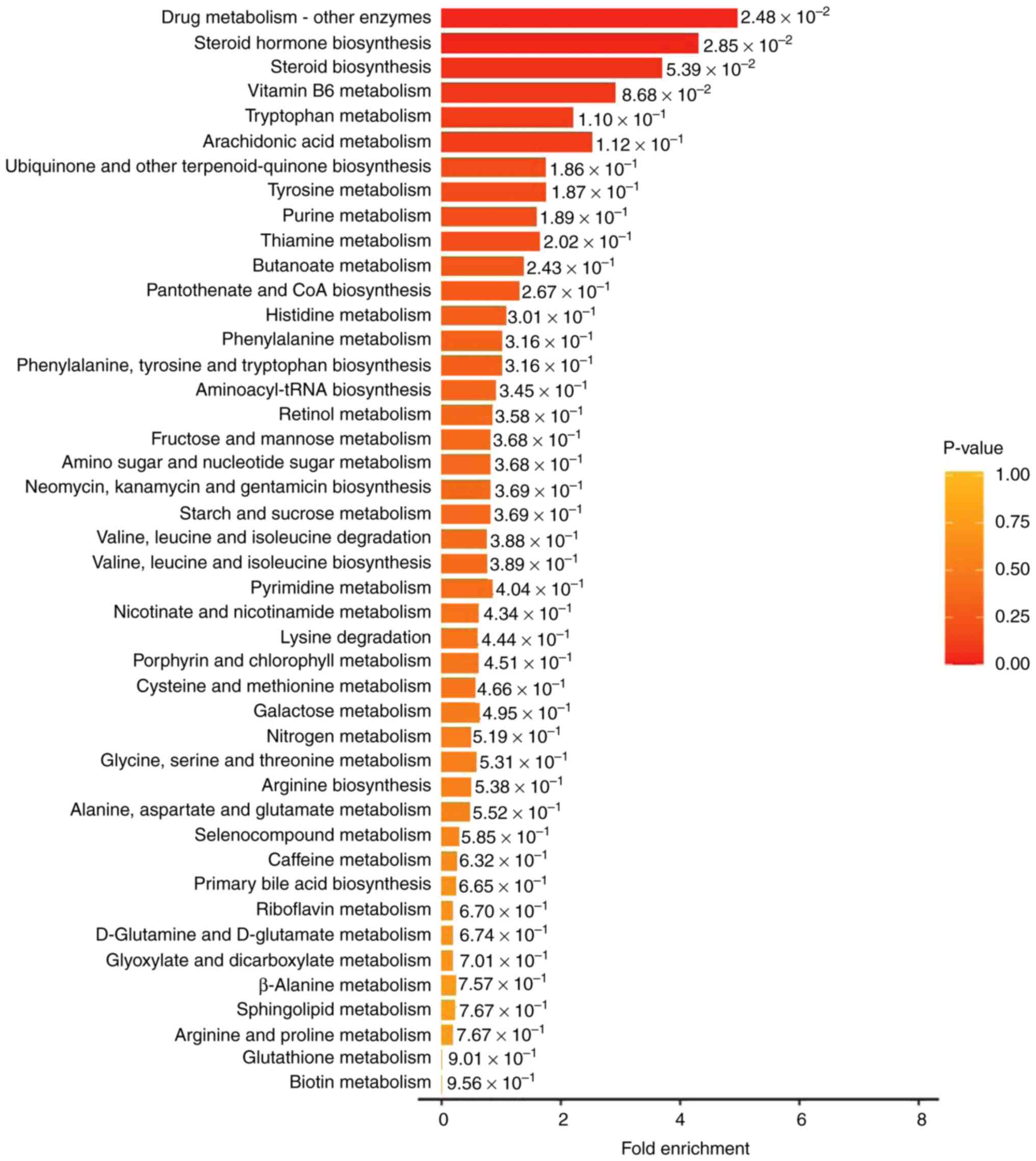
To identify distinct metabolite-associated pathways,
the 24 differential metabolites were imported into KEGG for pathway
enrichment analysis. KEGG is a public database providing integrated
metabolic pathways (23). Using
KEGG, the enriched pathways of the metabolites in the present study
were identified as the metabolism of amino acids, carbohydrates,
lipids, nucleotides, hormones and vitamins, and thermogenesis. The
classification annotation and differential abundance score of the
differential metabolites analyzed using KEGG are presented in
Fig. 4. There was a synergistic or
mutually exclusive relationship between the different
metabolites.
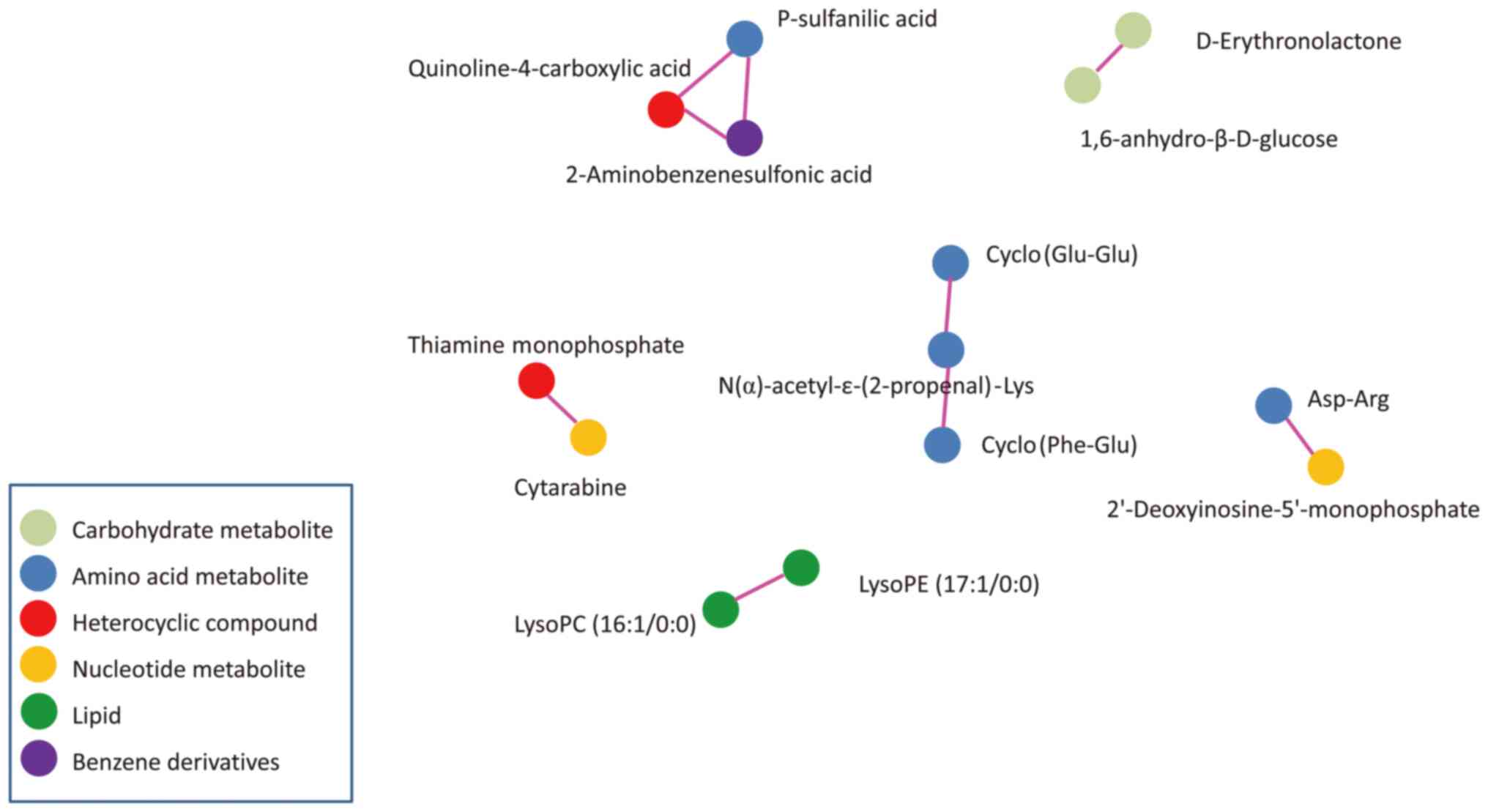
Pearson correlation analysis was used to measure the
metabolic proxies between significantly different metabolites
(Fig. 5). This analysis is helpful
to further understand the relationship between metabolites in the
process of biological metabolism. These differential metabolites
were carbohydrate metabolites, amino acid metabolites, heterocyclic
compounds, nucleotide metabolites, lipids and benzene
derivatives.
Discussion
In the present study, a total of 24 differential
metabolites were identified between CRC tissue and adjacent normal
tissues. These metabolites were associated with multi-pathways,
including energy, carbohydrate, amino acid, lipid, nucleotide,
vitamin metabolism and hormone. In CRC tissue two carbohydrate
metabolites were downregulated compared with normal tissue,
indicating an alteration of carbohydrate metabolism. Previous
evidence suggests that the energy of CRC cells may rely on the
reversed Warburg effect (24,25).
In other reports, 1,6-anhydro-β-D-glucose levels were revealed to
be normal in the feces of patients with CRC (26) and in the urine of patients with
lung cancer (27). This
discordance might be related to the different specimens used.
The present data demonstrated that there were
increases and decreases in different amino acid metabolites in CRC
tissue compared with normal tissue. Tumor cells increase both the
synthesis and degradation of proteins. These changes may indicate
the active metabolism of protein for cell proliferation, as well as
the degradation of protein in tumor cells (28). Notably, two CDPs, cyclo(Glu-Glu)
and cyclo(Phe-Glu), were increased in CRC tissues. To the best of
our knowledge, this is the first time that CDPs were found in CRC
tissue, and compared between tumor tissue and adjacent normal
tissues in patients with CRC. CDPs are the smallest of the cyclic
peptides and occur naturally, forming a large class of secondary
metabolites produced by microorganisms, plants and mammals. CDPs
possess specific chemical and diverse biological functions such as
antitumor, antimicrobial and radical-scavenging activities
(29). Although the source and
effects of CDPs in tissues are unknown, they are likely either
endogenous or microbial metabolites, since bacteria that exist in
CRC tissue serve a role in the tumorigenesis of CRC (30,31).
Asp-Arg is a structural unit of cyanophycin, a biopolymer of long
chains of Asp-Arg. Cyanophycin is synthesized by numerous types of
bacteria, of which cyanobacteria is the most well-known (32). It is possible that the changes in
the levels of Asp-Arg reflect the microbiota changes in the colon.
Future studies will compare the microbiota between the tumor
samples and normal samples to verify the source of CDPs and
Asp-Arg.
The alterations of lipid metabolism in cancer cells
are well known (33). Cancer
progression is associated with de novo lipogenesis and
supply from the microenvironment. The high energy requirement of
CRC during processes of malignant transformation and cancer cell
proliferation lead to the dysfunction of lipid metabolism (33). In agreement with previous studies,
lysoPCs, the product of the degradation of PC, was markedly
decreased in CRC tissue compared with normal tissue, and it has
been proposed that the increased breakdown of the hemolytic
metabolic product of phosphatidylcholine increased cancer
malignancy (11,34). The reason for the reduction of
lipid metabolites could be due to an increased degradation and
reduced supply from the microenvironment (9). Cleavage of lysoPC may increase free
fatty acid, and subsequently this free fatty acid can be used by
cancer cells for energy via fatty acid oxidation (33). However, carnitine, a major
transporter for fatty acid acyl moieties entering the mitochondrial
matrix for oxidation (35), was
decreased in CRC tissue compared with normal tissue in the present
study, which was not in agreement with previous reports (11,36).
Carnitine is also considered to inhibit cell proliferation and
increase apoptosis (37). The
decrease of carnitine indicated the reduction of lipid metabolism
in the mitochondria. Although lipid metabolites have been proposed
as diagnostic and prognostic predictors for patients with CRC
(38,39), the alteration of lipids in CRC
tissue is inconsistent, and as such, the influence of lipids on
tumorigenesis is inconclusive (40).
Nucleotide metabolism serves a role in
carcinogenesis and cancer progression. It not only meets the
requirements of cell proliferation but also affects a variety of
activities beyond proliferation, such as, tumor immunity, antitumor
drug resistance and interactions of the tumor cell with the
microenvironment (41). Cancer
cells tend to use the de novo nucleotide synthesis pathway
(42). In the present study, the
levels of nucleotide metabolites, including cytarabine,
5′-deoxy-5′-(methylthio) adenosine, deoxyribose 5-phosphate and
2′-deoxyinosine-5′-monophosphate, were decreased in CRC tissues
compared with normal tissue. It was hypothesized that the decrease
of these metabolites may indicate the insufficient replenishment of
nucleotide metabolites for active nucleotide synthesis in CRC
cells. Unlike lipids and proteins, cancer cells may not be
obtaining sufficient amounts of nucleotides and their components
from the tumor microenvironment (9). The decreased levels of
5′-deoxy-5′-(methylthio) adenosine in CRC tissues, indicated
alterations in the cysteine and methionine metabolism pathways.
Decreases in the 17α- and 17β-estradiol levels in
the tumor tissues regardless of sex is consistent with the notion
that estrogen has an anti-tumor effect in CRC, presumably by
activating the estrogen receptor (ER)-β (43,44).
ER-β-modulated transcription events resulted in the stimulation of
proapoptotic signaling, regulation of mismatch repair proteins as
well as the modulation of the antitumor immune response (45). Quinaldic acid, a tryptophan
metabolite and a derivative of kynurenic acid, induced apoptosis in
colon cancer cells via the activation of the p53 tumor suppressor
but had no toxicity to non-cancerous colon cell lines (46,47).
Therefore, decreased levels of quinaldic acid contributed to colon
cancer cell growth. Notably, the levels of 2-aminobenenesulfonic
acid, quinoline-4-carboxylic acid and p-sulfanilic acid were
revealed to be correlated to one another, even though they are in
three different KEGG pathways. In fact, sulfanilic acid (also
referred to as 4-aminobenzenesulfonic acid) is an intermediate
product of azo dyes, plant protectives and detergents (48). In the present study, it was
revealed that the levels of these metabolites were correlated to
the stage of CRC (Table V). It is
unclear the resource of sulfanilic acid. Since sulfanilic acid is
degraded by Pseudomonas paucimobilis, an aerobic bacterium,
perhaps bacterial colonization may be increased in the tumor.
In the present study, the level of lysoPE (17:1/0:0)
was increased on the right-side of the colon compared with the
left-side of the colon and rectum. Additionally, there were
metabolite profile differences between the right- and left-side
colon tissues in healthy individuals (49). Cai et al (16) revealed that there were differences
in metabolites in early-stage CRC tissues between the right- and
left-side of the colon. The anatomic location of the CRC was
relevant to the development and prognosis of the patients with CRC
(50). In the present study, the
changes of six differential metabolites in different stages of CRC
were significant, including 2-aminobenzenesulfonic acid,
p-sulfanilic acid, quinoline-4-carboxylic acid,
N-α-acetyl-ε-(2-propenal)-Lys, methylcysteine and
5′-deoxy-5′-(methylthio) adenosine. These metabolites are involved
in amino acid and nucleotide metabolism. Although previous studies
compared the changes of differential metabolites in different
stages of CRC tissues, there were various results in different
studies (11,16,51,52).
The data of the present study demonstrated that the level of
differential metabolites were regardless of sex and
differentiations of CRC. Since the sample size in the present study
was small, further studies are required to confirm the results of
the present study.
The detection of metabolites within cancer tissue
directly reflects the true metabolic state of the cancer cell
unlike the use of other specimens, such as serum, urine and stool.
However, unlike proteomics, genomics or transcriptomics,
metabolomics only represents a transient phenotypic state that may
vary rapidly (11). The results of
the present study agreed with the findings of a number of previous
studies, while they differed from others (11,36,53).
The differential metabolic profile identified in the present study
was unique and different from other studies (Table IV). There were multiple factors
that could have caused the variations in the metabolites between
the different studies, such as different preparation methods,
samples collected from patients of different ages, weight, diet and
even circadian rhythm (54), or
different analytic platforms and processing software (11,36,55).
Further conclusive studies are required before any clear
explanations can be offered for the different changes in the
metabolites.
The present study had a number of limitations.
Firstly, the present study had a small sample size. The
verification of the results in another set of patients compared
with the one used for metabolomics analysis could not be performed
because of the lack of available samples. Further investigation
using a larger sample size would increase the accuracy and
reproducibility of the study. Secondly, the present study is
cross-sectional. Further longitudinal studies or experimental
studies could provide further evidence to confirm the relationship
between metabolite levels and CRC, and may elucidate the mechanisms
of the alterations in the tumorigenesis of CRC. Thirdly, the
difference between cancer tissues and adenomatous polyps, a
precursor lesion for the majority of CRC, could have been compared
in order to investigate the metabolic changes in the transition to
malignancy. Fourthly, if the metabolite changes could be confirmed
using blood samples from patients, it could aid the establishment
of biomarkers for CRC. The patients of the present study are being
followed-up for further investigation of the relationship between
the alteration of metabolism and prognosis. Additionally, future
study will compare the alteration of metabolites in tissue and
blood.
In conclusion, the present study revealed that the
alteration of metabolism in CRC cells could provide novel insights
to decode an unknown aspect of colon carcinogenesis. The
differential levels of the metabolites may serve as biomarkers in
CRC. With collective efforts, further studies on CRC metabolites
may lead to an improved diagnosis or treatment of CRC.
Acknowledgements
The authors would like to thank Dr. Chuixiong Chen
from Key Tech Statistics and Science Company (Beijing, China) for
his work on the statistical analysis.
Funding
This study was supported by National Natural Science Foundation
of China (grant no. 81370487), Scientific Research Fund of Beijing
Rehabilitation Hospital, Capital Medical University (grant nos.
2017-004, 2021-018, 2019-043, 2019R-001 and 2020-056 and 2021-015)
and Scientific Research Fund of Beijing Anorectal Society (grant
no. 2020ABCP002). These funds were used for the design of the
study, sample collection, data analysis, interpretation of the data
and in writing the manuscript.
Availability of data and materials
The datasets generated during the current study are
available in the MetaboLights repository, https://www.ebi.ac.uk/metabolights/ (study no.
MTBLS8090).
Authors' contributions
QG, CK, JZ, SJ and FC conceived and designed the
study. CK, JZ, MZ, XL and MX recruited patients and collected
samples. JZ, MZ, MX, YW, XL and DD collected data. QG, FC, MZ CK,
SJ and XL were involved in data analysis and the literature search.
JZ, MZ, YW, DD and MX were involved in project administration. QG,
SJ, CK and MZ wrote the manuscript. QG and CK confirm the
authenticity of all the raw data. QG, CK, MX and MZ acquired
funding. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
All procedures used in this research were approved
by the Ethics Committee of Beijing Rehabilitation Hospital
(approval no. 2020bkkyLW003). Written informed consent was obtained
from all study participants. All methods performed in this study
were in accordance with the Declaration of Helsinki.
Patient consent for publication
All study participants gave consent to publish the
data of the study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Baidoun F, Elshiwy K, Elkeraie Y, Merjaneh
Z, Khoudari G, Sarmini MT, Gad M, Al-Husseini M and Saad A:
Colorectal cancer epidemiology: Recent trends and impact on
outcomes. Curr Drug Targets. 22:998–1009. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD, Fuchs HE and Jemal
A: Cancer statistics, 2021. CA Cancer J Clin. 71:7–33. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xia C, Dong X, Li H, Cao M, Sun D, He S,
Yang F, Yan X, Zhang S, Li N and Chen W: Cancer statistics in China
and United States, 2022: Profiles, trends, and determinants. Chin
Med J (Engl). 135:584–590. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu X, Lin H and Li S: Prognoses of
different pathological subtypes of colorectal cancer at different
stages: A population-based retrospective cohort study. BMC
Gastroenterol. 19:1642019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kanth P and Inadomi JM: Screening and
prevention of colorectal cancer. BMJ. 374:n18552021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Martinez-Reyes I and Chandel NS: Cancer
metabolism: Looking forward. Nat Rev Cancer. 21:669–680. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vander Heiden MG and DeBerardinis RJ:
Understanding the intersections between metabolism and cancer
biology. Cell. 168:657–669. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Klassen A, Faccio AT, Canuto GA, da Cruz
PL, Ribeiro HC, Tavares MF and Sussulini A: Metabolomics:
Definitions and significance in systems biology. Adv Exp Med Biol.
965:3–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gold A, Choueiry F, Jin N, Mo X and Zhu J:
The Application of metabolomics in recent colorectal cancer
studies: A state-of-the-art review. Cancers (Basel). 14:7252022.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Erben V, Bhardwaj M, Schrotz-King P and
Brenner H: Metabolomics biomarkers for detection of colorectal
neoplasms: A systematic review. Cancers (Basel). 10:2462018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hashim NAA, Ab-Rahim S, Suddin LS, Saman
MSA and Mazlan M: Global serum metabolomics profiling of colorectal
cancer. Mol Clin Oncol. 11:3–14. 2019.PubMed/NCBI
|
|
14
|
Di Giovanni N, Meuwis MA, Louis E and
Focant JF: Specificity of metabolic colorectal cancer biomarkers in
serum through effect size. Metabolomics. 16:882020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shen Y, Sun M, Zhu J, Wei M, Li H, Zhao P,
Wang J, Li R, Tian L, Tao Y, et al: Tissue metabolic profiling
reveals major metabolic alteration in colorectal cancer. Mol Omics.
17:464–471. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cai Y, Rattray NJW, Zhang Q, Mironova V,
Santos-Neto A, Muca E, Vollmar AKR, Hsu KS, Rattray Z, Cross JR, et
al: Tumor tissue-specific biomarkers of colorectal cancer by
anatomic location and stage. Metabolites. 10:2572020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao P, Zhou C, Zhao L, Zhang G and Zhang
Y: Tissue amino acid profile could be used to differentiate
advanced adenoma from colorectal cancer. J Pharm Biomed Anal.
118:349–355. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hirayama A, Kami K, Sugimoto M, Sugawara
M, Toki N, Onozuka H, Kinoshita T, Saito N, Ochiai A, Tomita M, et
al: Quantitative metabolome profiling of colon and stomach cancer
microenvironment by capillary electrophoresis time-of-flight mass
spectrometry. Cancer Res. 69:4918–4925. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Romisch-Margl W, Prehn C, Bogumil R,
Röhring C, Suhre K and Adamski J: Procedure for tissue sample
preparation and metabolite extraction for high-throughput targeted
metabolomics. Metabolomics. 8:133–142. 2012. View Article : Google Scholar
|
|
20
|
Xu Y, Wang X, Han D, Wang J, Luo Z, Jin T,
Shi C, Zhou X, Lin L and Shan J: Revealing the mechanism of Jiegeng
decoction attenuates bleomycin-induced pulmonary fibrosis via
PI3K/Akt signaling pathway based on lipidomics and transcriptomics.
Phytomedicine. 102:1542072022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiao Y, Liu H, Li H, Liu Q, Lu Q, Varshney
RK, Chen X and Honget Y: Widely targeted metabolomics characterizes
the dynamic changes of chemical profile in postharvest peanut
sprouts grown under the dark and light conditions. Food Sci
Technol. 152:1122832021.
|
|
22
|
McCann A, Midttun Ø, Whitfield KC, Kroeun
H, Borath M, Sophonneary P, Ueland PM and Green TJ: Comparable
performance characteristics of plasma thiamine and erythrocyte
thiamine diphosphate in response to thiamine fortification in rural
cambodian women. Nutrients. 9:6762017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: New perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45:D353–D361. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chekulayev V, Mado K, Shevchuk I, Koit A,
Kaldma A, Klepinin A, Timohhina N, Tepp K, Kandashvili M, Ounpuu L,
et al: Metabolic remodeling in human colorectal cancer and
surrounding tissues: Alterations in regulation of mitochondrial
respiration and metabolic fluxes. Biochem Biophys Rep. 4:111–125.
2015.PubMed/NCBI
|
|
25
|
Satoh K, Yachida S, Sugimoto M, Oshima M,
Nakagawa T, Akamoto S, Tabata S, Saitoh K, Kato K, Sato S, et al:
Global metabolic reprogramming of colorectal cancer occurs at
adenoma stage and is induced by MYC. Proc Natl Acad Sci USA.
114:E7697–E7706. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Goedert JJ, Sampson JN, Moore SC, Xiao Q,
Xiong X, Hayes RB, Ahn J, Shi J and Sinha R: Fecal metabolomics:
Assay performance and association with colorectal cancer.
Carcinogenesis. 35:2089–2096. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stretch C, Eastman T, Mandal R, Eisner R,
Wishart DS, Mourtzakis M, Prado CM, Damaraju S, Ball RO, Greiner R
and Baracos VE: Prediction of skeletal muscle and fat mass in
patients with advanced cancer using a metabolomic approach. J Nutr.
142:14–21. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vettore L, Westbrook RL and Tennant DA:
New aspects of amino acid metabolism in cancer. Br J Cancer.
122:150–156. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mishra AK, Choi J, Choi SJ and Baek KH:
Cyclodipeptides: An overview of their biosynthesis and biological
activity. Molecules. 22:17962017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mima K, Nishihara R, Qian ZR, Cao Y,
Sukawa Y, Nowak JA, Yang J, Dou R, Masugi Y, Song M, et al:
Fusobacterium nucleatum in colorectal carcinoma tissue and patient
prognosis. Gut. 65:1973–1980. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cheng Y, Ling Z and Li L: The intestinal
microbiota and colorectal cancer. Front Immunol. 11:6150562020.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sharon I, Grogg M, Hilvert D and Schmeing
TM: Structure and Function of the β-Asp-Arg Polymerase Cyanophycin
Synthetase 2. ACS Chem Biol. 17:670–679. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Santos CR and Schulze A: Lipid metabolism
in cancer. FEBS J. 279:2610–2623. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao Z, Xiao Y, Elson P, Tan H, Plummer
SJ, Berk M, Aung PP, Lavery IC, Achkar JP, Li L, et al: Plasma
lysophosphatidylcholine levels: Potential biomarkers for colorectal
cancer. J Clin Oncol. 25:2696–2701. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Console L, Scalise M, Mazza T, Pochini L,
Galluccio M, Giangregorio N, Tonazzi A and Indiveri C: Carnitine
traffic in cells. Link with cancer. Front Cell Dev Biol.
8:5838502020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang F, Zhang Y, Zhao W, Deng K, Wang Z,
Yang C, Ma L, Openkova MS, Hou Y and Li K: Metabolomics for
biomarker discovery in the diagnosis, prognosis, survival and
recurrence of colorectal cancer: A systematic review. Oncotarget.
8:35460–35472. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dionne S, Elimrani I, Roy MJ, Qureshi IA,
Sarma DR, Levy E and Seidman EG: Studies on the chemopreventive
effect of carnitine on tumorigenesis in vivo, using two
experimental murine models of colon cancer. Nutr Cancer.
64:1279–1287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun Y, Liu B, Chen Y, Xing Y and Zhang Y:
Multi-Omics prognostic signatures based on lipid metabolism for
colorectal cancer. Front Cell Dev Biol. 9:8119572021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kurabe N, Hayasaka T, Ogawa M, Masaki N,
Ide Y, Waki M, Nakamura T, Kurachi K, Kahyo T, Shinmura K, et al:
Accumulated phosphatidylcholine (16:0/16:1) in human colorectal
cancer; possible involvement of LPCAT4. Cancer Sci. 104:1295–1302.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pakiet A, Kobiela J, Stepnowski P,
Sledzinski T and Mika A: Changes in lipids composition and
metabolism in colorectal cancer: A review. Lipids Health Dis.
18:292019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ma J, Zhong M, Xiong Y, Gao Z, Wu Z, Liu Y
and Hong X: Emerging roles of nucleotide metabolism in cancer
development: Progress and prospect. Aging. 13:13349–13358. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lane AN and Fan TW: Regulation of
mammalian nucleotide metabolism and biosynthesis. Nucleic Acids
Res. 43:2466–2485. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Haziman AA, Ravinderan S, Thangavelu T and
Thomas W: A novel role for estrogen-induced signaling in the
colorectal cancer gender bias. Ir J Med Sci. 188:389–395. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ditonno I, Losurdo G, Rendina M, Pricci M,
Girardi B, Ierardi E and Di Leo A: Estrogen receptors in colorectal
cancer: Facts, novelties and perspectives. Curr Oncol.
28:4256–4263. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Caiazza F, Ryan EJ, Doherty G, Winter DC
and Sheahan K: Estrogen receptors and their implications in
colorectal carcinogenesis. Front Oncol. 5:192015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Langner E, Jeleniewicz W, Turski WA and
Plech T: Quinaldic acid induces changes in the expression of p53
tumor suppressor both on protein and gene level in colon cancer
LS180 cells. Pharmacol Rep. 71:189–193. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Langner E, Walczak K, Jeleniewicz W,
Turski WA and Rajtar G: Quinaldic acid inhibits proliferation of
colon cancer ht-29 cells in vitro: Effects on signaling pathways.
Eur J Pharmacol. 757:21–27. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Perei K, Rakhely G, Kiss I, Polyák B and
Kovács KL: Biodegradation of sulfanilic acid by Pseudomonas
paucimobilis. Appl Microbiol biotechnol. 55:101–107. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Baxter BA, Parker KD, Nosler MJ, Rao S,
Craig R, Seiler C and Ryan EP: Metabolite profile comparisons
between ascending and descending colon tissue in healthy adults.
World J Gastroenterol. 26:335–352. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wong MCS, Huang J, Lok V, Wang J, Fung F,
Ding H and Zheng ZJ: Differences in incidence and mortality trends
of colorectal cancer worldwide based on sex, age, and anatomic
location. Clin Gastroenterol Hepatol. 19:955–966.e61. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Mirnezami R, Jimenez B, Li JV, Kinross JM,
Veselkov K, Goldin RD, Holmes E, Nicholson JK and Darzi A: Rapid
diagnosis and staging of colorectal cancer via high-resolution
magic angle spinning nuclear magnetic resonance (HR-MAS NMR)
spectroscopy of intact tissue biopsies. Ann Surg. 259:1138–1149.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jimenez B, Mirnezami R, Kinross J, Cloarec
O, Keun HC, Holmes E, Goldin RD, Ziprin P, Darzi A and Nicholson
JK: 1H HR-MAS NMR spectroscopy of tumor-induced local metabolic
‘field-effects’ enables colorectal cancer staging and
prognostication. J Proteome Res. 12:959–968. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tian JS, Xue WN, Yin HH, Zhang N, Zhou J,
Long Z, Wu C, Liang Z, Xie K, Li S, et al: Differential metabolic
alterations and biomarkers between gastric cancer and colorectal
cancer: A systematic review and meta-analysis. Onco Targets Ther.
13:6093–6108. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kosmides AK, Kamisoglu K, Calvano SE,
Corbett SA and Androulakis IP: Metabolomic fingerprinting:
Challenges and opportunities. Crit Rev Biomed Eng. 41:205–221.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yusof HM, Ab-Rahim S, Suddin LS, Saman MSA
and Mazlan M: Metabolomics profiling on different stages of
colorectal cancer: A systematic review. Malays J Med Sci. 25:16–34.
2018.PubMed/NCBI
|
|
56
|
Long Z, Zhou J, Xie K, Wu Z, Yin H, Daria
V, Tian J, Zhang N, Li L, Zhao Y, et al: Metabolomic markers of
colorectal tumor with different clinicopathological features. Front
Oncol. 10:9812020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Manna SK, Tanaka N, Krausz KW, Haznadar M,
Xue X, Matsubara T, Bowman ED, Fearon ER, Harris CC, Shah YM and
Gonzalez FJ: Biomarkers of coordinate metabolic reprogramming in
colorectal tumors in mice and humans. Gastroenterology.
146:1313–1324. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Shen X, Cai Y, Lu L, Huang H, Yan H, Paty
PB, Muca E, Ahuja N, Zhang Y, Johnson CH and Khan SA: Asparagine
metabolism in tumors is linked to poor survival in females with
colorectal cancer: A cohort study. Metabolites. 12:1642022.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ong ES, Zou L, Li S, Cheah PY, Eu KW and
Ong CN: Metabolic profiling in colorectal cancer reveals signature
metabolic shifts during tumorigenesis. Mol Cell Proteomics. Feb
10–2010.doi: 10.1074/mcp.M900551-MCP200 (Epub ahead of print).
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Li Z, Deng X, Luo J, Lei Y, Jin X, Zhu J
and Lv G: Metabolomic comparison of patients with colorectal cancer
at different anticancer treatment stages. Front Oncol.
11:5743182021. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Brown DG, Rao S, Weir TL, O'Malia J, Bazan
M, Brown RJ and Ryan EP: Metabolomics and metabolic pathway
networks from human colorectal cancers, adjacent mucosa, and stool.
Cancer Metab. 4:112016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Liesenfeld DB, Habermann N, Toth R, Owen
RW, Frei E, Staffa J, Schrotz-King P, Klika KD and Ulrich CM:
Changes in urinary metabolic profiles of colorectal cancer patients
enrolled in a prospective cohort study (ColoCare). Metabolomics.
11:998–1012. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Udo R, Katsumata K, Kuwabara H, Enomoto M,
Ishizaki T, Sunamura M, Nagakawa Y, Soya R, Sugimoto M and Tsuchida
A: Urinary charged metabolite profiling of colorectal cancer using
capillary electrophoresis-mass spectrometry. Sci Rep. 10:210572020.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Williams MD, Zhang X, Park JJ, Siems WF,
Gang DR, Resar LM, Reeves R and Hill HH Jr: Characterizing
metabolic changes in human colorectal cancer. Anal Bioanal Chem.
407:4581–4595. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wang Y, Hinz S, Uckermann O, Hönscheid P,
von Schönfels W, Burmeister G, Hendricks A, Ackerman JM, Baretton
GB, Hampe J, et al: Shotgun lipidomics-based characterization of
the landscape of lipid metabolism in colorectal cancer. Biochim
Biophys Acta Mol Cell Biol Lipids. 1865:1585792020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Shen S, Yang L, Li L, Bai Y, Cai C and Liu
H: A plasma lipidomics strategy reveals perturbed lipid metabolic
pathways and potential lipid biomarkers of human colorectal cancer.
J Chromatogr B Analyt Technol Biomed Life Sci. 1068–1069. 41–48.
2017.PubMed/NCBI
|
|
67
|
Cai Y, Rattray NJW, Zhang Q, Mironova V,
Santos-Neto A, Hsu KS, Rattray Z, Cross JR, Zhang Y, Paty PB, et
al: Sex differences in colon cancer metabolism reveal a novel
subphenotype. Sci Rep. 10:49052020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ecker J, Benedetti E, Kindt ASD, Höring M,
Perl M, Machmüller AC, Sichler A, Plagge J, Wang Y, Zeissig S, et
al: The colorectal cancer lipidome: Identification of a robust
Tumor-Specific lipid species signature. Gastroenterology.
161:910–923.e19. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kuhn T, Floegel A, Sookthai D, Johnson T,
Rolle-Kampczyk U, Otto W, von Bergen M, Boeing H and Kaaks R:
Higher plasma levels of lysophosphatidylcholine 18:0 are related to
a lower risk of common cancers in a prospective metabolomics study.
BMC Med. 14:132016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Holowatyj AN, Gigic B, Herpel E, Scalbert
A, Schneider M and Ulrich CM; MetaboCCC Consortium; ColoCare Study,
: Distinct molecular phenotype of sporadic colorectal cancers among
young patients based on multiomics analysis. Gastroenterology.
158:1155–1158.e2. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zarei I, Baxter BA, Oppel RC, Borresen EC,
Brown RJ and Ryan EP: Plasma and urine metabolite profiles impacted
by increased dietary navy bean intake in colorectal cancer
survivors: A randomized-controlled trial. Cancer Prev Res (Phila).
14:497–508. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lin L, Zeng X, Liang S, Wang Y, Dai X, Sun
Y and Wu Z: Biomarkers of coordinate metabolic reprogramming and
the construction of a co-expression network in colorectal cancer.
Ann Transl Med. 10:11152022. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Li M, Wu S, Zhuang C, Shi C, Gu L, Wang P,
Guo F, Wang Y and Liu Z: Metabolomic analysis of circulating tumor
cells derived liver metastasis of colorectal cancer. Heliyon.
9:e125152023. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Montrose DC, Zhou XK, Kopelovich L,
Yantiss RK, Karoly ED, Subbaramaiah K and Dannenberg AJ: Metabolic
profiling, a noninvasive approach for the detection of experimental
colorectal neoplasia. Cancer Prev Res (Phila). 5:1358–1367. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Yoshie T, Nishiumi S, Izumi Y, Sakai A,
Inoue J, Azuma T and Yoshida M: Regulation of the metabolite
profile by an APC gene mutation in colorectal cancer. Cancer Sci.
103:1010–1021. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Long Y, Sanchez-Espiridion B, Lin M, White
L, Mishra L, Raju GS, Kopetz S, Eng C, Hildebrandt MAT, Chang DW,
et al: Global and targeted serum metabolic profiling of colorectal
cancer progression. Cancer. 123:4066–4074. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Luo Z, Wang H, Lin S, Liao L, Cai L, Zhang
X, Tan Y and Shen M: Study on the levels of N-nitrosamine compounds
and untargeted metabolomics in patients with colorectal cancer.
Anal Bioanal Chem. 414:3483–3496. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Yang F, DeLuca JAA, Menon R,
Garcia-Vilarato E, Callaway E, Landrock KK, Lee K, Safe SH, Chapkin
RS, Allred CD and Jayaraman A: Effect of diet and intestinal AhR
expression on fecal microbiome and metabolomic profiles. Microb
Cell Fact. 19:2192020. View Article : Google Scholar : PubMed/NCBI
|