Introduction
Epidemiological studies show that head and neck
squamous cell carcinoma (HNSCC) is the sixth most commonly
occurring human neoplasm and includes cancers of the oral cavity,
larynx, nasal passages, pharynx and salivary glands. HNSCC accounts
for approximately 10% of the total cancer burden in men in Europe
(1). In 2008, there were 47,560
estimated new cases of head and neck cancers in the US, with 11,260
fatalities (2). Although certain
independent risk factors for the incidence of HNSCC such as alcohol
and tobacco exposure or a family history of malignancy have been
revealed (3,4), complete molecular pathogenesis in
these human malignancies has yet to be adequately elucidated.
Secreted protein, acidic and rich in cysteine
(SPARC)has generated considerable interest as a tumor-associated
protein for its diverse actions and complex functions. An enhanced
expression of SPARC is associated with a highly aggressive tumor
phenotype in melanomas and gliomas, as supported by previous
functional verification (5–8). However, other studies reported that
SPARC acts as a tumor suppressor gene (TSG) in pancreatic
adenocarcinoma, acute myeloid leukemia and ovarian and colorectal
carcinomas (9–12). Furthermore, recent studies have
shown that SPARC mediates the interaction between cells and the
extracellular environment as a matricellular protein and is
associated with chemotherapy sensitivity (13,14).
However, the role played by SPARC in the tumorigenesis of various
common HNSCCs remains to be clarified. The present study aimed to
investigate the mechanism underlying the abnormal expression of
SPARC in laryngeal and hypopharyngeal carcinomas, the two most
common HNSCCs. The clinical significance of aberrant promoter
methylation was also investigated.
Materials and methods
HNSCC cell lines, primary tumors and
matched specimens of normal epithelia
Human laryngeal carcinoma cell line Hep-2 was
maintained in our laboratory. The cancer cell lines Tu212 and Tu686
were established from primary hypopharyngeal carcinoma, and primary
tongue carcinomas, respectively, from the University of Texas, M.D.
Anderson Cancer Center (Houston, TX, USA), as previously described
(15), together with HNSCC cell
lines M2E and M4E were kindly provided by Dr He Zhu (XiangYa
Central Experiment Laboratory, Changsha, China). A total of 51
primary tumor biopsies of laryngeal and hypopharyngeal carcinoma,
with 9 samples of normal epithelia from areas adjacent to the
tumors, were included in the present study. The tumor biopsies and
matched normal epithelia were obtained at the Department of
Otolaryngology-Head and Neck Surgery, First Affiliated Hospital of
Guangxi Medical University (Nanning, China), between July 2008 and
December 2009. Tissues from the patients were immediately frozen in
liquid nitrogen and stored at −180˚C until required. Diagnoses were
established according to the World Health Organization
classification. None of the patients had been administered with
preoperative radiation or chemotherapy. Formal written consent was
obtained from all patients and approval was obtained from the local
ethics committee.
Semi-quantitative reverse transcription
polymerase chain reaction (RT-PCR)
Total RNA of the cell lines, primary tumor biopsies
and adjacent normal epithelia were isolated with TRIzol reagent
(Invitrogen, Carlsbad, CA, USA). First strand cDNA was synthesized
with M-MLV reverse transcriptase (Promega, Madison, WI, USA)
according to the manufacturer's instructions. Total RNA (2 μg) was
used for each reaction. The primer sequences used were previously
reported (9); forward:
5′-AAGCTCACTGGCATGGCCTT-3′ and reverse:
5′-CTCTCTTCCTCTTGTGCTCTTG-3′ at 61˚C for 30 sec and 72˚C for 1 min
at 24 cycles (GAPDH). PCR was carried out in a total volume of 25
μl. The PCR mixture contained 10 pmol of each primer, 100 pmol of
deoxynucleoside triphosphate, 1X PCR buffer, 1 unit of ExTaq HS
polymerase (Takara), and 2 μl of cDNA. The amplified PCR products
were then identified on 2% agarose gels. Images of ethidium
bromide-stained agarose gels were acquired using a CCD camera
(Bio-Rad, Hercules, CA, USA) and semi-quantitative analysis was
performed using Quantity-One software, version 4.4.0 (Bio-Rad).
Sodium bisulphite modification of genomic
DNA
High-molecular weight genomic DNA was extracted from
cell lines and biopsies using a conventional phenol/chloroform
method. The sodium bisulphite modification procedure was a slight
modification of the protocol previously reported (16). Briefly, 500 ng of genomic DNA was
denatured in 0.3 M NaOH for 15 min at 37˚C, and then mixed with 2
volumes of 2% low melting point agarose gels. Agarose/DNA mixtures
were then pipetted into chilled mineral oil to form agarose beads.
Aliquots of 200 μl of 5 M bisulphite solution (2.5 M sodium
metabisulphite and 100 mM hydroquinone; Sigma, St. Louis, MO, USA)
were added to each tube containing a single bead. The bisulphite
reaction was then carried out by incubating the reaction mixture
for 4 h at 50˚C in the dark. Treatments were stopped by
equilibration against 1 ml of TE buffer followed by desulphonation
in 500 μl of 0.2 M NaOH. Finally, the beads were washed with 1 ml
of H2O and directly used for PCRs.
Methylation-specific PCR (MSP)
The methylation status of the SPARC promoter region
was determined by MSP. Primers distinguishing unmethylated (U) and
methylated (M) alleles were designed to amplify the sequence within
the CpG island around exon 1 of SPARC, as previously described
(9). Each MSP reaction contained 20
ng of sodium bisulphite-modified DNA, 10 pmol of each primer, 100
pmol of deoxynucleoside triphosphate, 1X PCR buffer, and 1 unit of
ExTaq HS polymerase in a final reaction volume of 25 μl. Cycling
conditions were as previously described (9). PCR products were separated on 2%
agarose gels, stained with ethidium bromide and visualized under UV
illumination. For cases with borderline results, PCR analyses were
repeated.
5-Aza-2′-deoxycytidine treatment
(5-Aza-dC)
The human laryngeal carcinoma cell line Hep-2 and
hypopharyngeal carcinoma cell line Tu212 were selected to perform
5-aza-dC treatment. Cells were seeded into 6-well plates with a
density of 2×105 cells/well. Medium containing 0, 5 or
10 μM DNA methyltransferase inhibitor 5-aza-dC (Sigma) was added 24
h later and replaced every 24 h. After 72 h of demethylation
treatment, the restoration of SPARC expression was examined by
RT-PCR, as mentioned above.
Bisulphite genomic sequencing (BGS)
Sodium bisulphite-modified DNA was subjected to PCR
with primers designed to amplify nucleotides from −29 to +191 bp
relative to the transcription start site of the SPARC gene, as
previously described (9). PCR was
carried out in a total volume of 25 μl, containing 20 ng of sodium
bisulphite-modified DNA as a template, 10 pmol of each primer, 100
pmol of deoxynucleoside triphosphate, 1X PCR buffer, and 1 unit of
ExTaq HS polymerase (Takara). PCR products were then gel-purified
and cloned using the pMD18-T vector (Takara) and JM109 competent
E. coli cells. Colonies were grown on agar plates and five
colonies of each sample were randomly selected. Plasmids were then
isolated and purified. Sequencing was carried out using a BigDye
terminator cycle sequencing kit 3.0 (Applied Biosystems, Carlsbad,
CA, USA) on an ABI 3100 sequencer according to the manufacturer's
instructions.
Statistical analysis
An independent sample t-test was used to compare
transcription expression levels between methylated tumor tissues
and normal epithelia, or between methylated and unmethylated tumor
tissues. The correlation between numbers of methylated samples and
clinicopathological characteristics was analyzed by the
χ2 or Fisher's exact test. P<0.05 was considered to
be statistically significant.
Results
Expression and methylation status of
SPARC in cancer cell lines
Initially, transcription levels of SPARC were
examined by RT-PCR in various HNSCC cell lines, including laryngeal
and hypopharyngeal carcinoma cell lines. SPARC expression was
undetectable in 100% (5/5) of HNSCC cell lines. However, SPARC was
expressed at an easily detected level in normal epithelial
specimens (Fig. 1A). The
methylation status of SPARC was investigated by an MSP assay, and
aberrant promoter methylation was detected in all of the cell
lines, but not in any of the normal epithelial specimens (Fig. 1B).
Transcription levels and aberrant DNA
methylation of SPARC in primary tumors of laryngeal and
hypopharyngeal carcinoma
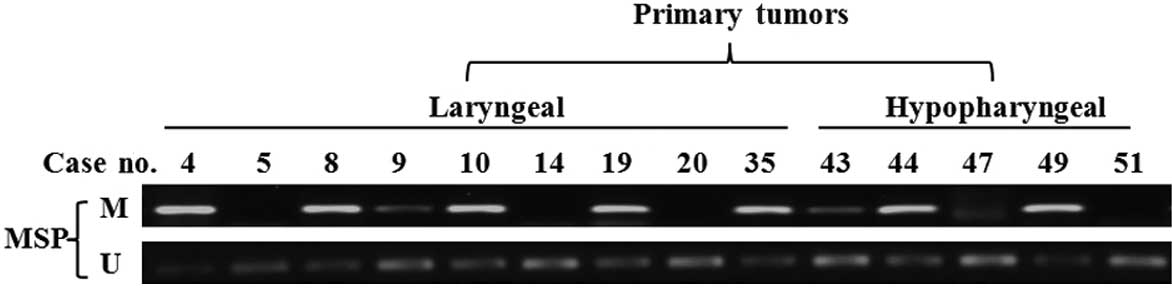
The methylation status of the SPARC promoter was
detected in 51 primary tumor biopsies. The MSP assay showed that
SPARC promoter was hypermethylated in 56.1% (23/41) of laryngeal
carcinoma and 70.0% (7/10) of hypopharyngeal carcinoma specimens
(Fig. 2A), but only in 11.1% (1/9)
of samples of normal epithelia (Fig.
2B). In order to evaluate whether methylation is involved in
the down-regulation of SPARC in laryngeal and hypopharyngeal
carcinoma, transcription levels in methylated tumor, unmethylated
tumor and normal epithelial tissues were examined. The
transcription expression levels of SPARC in methylated-tumor
tissues were found to be significantly lower than those of
unmethylated tumor tissues (p<0.05, Fig. 2C), but no significant difference was
observed between primary tumors and normal epithelia. This negative
result may be attributed either to a smaller number of normal
epithelial samples as compared to that required for statistical
analysis or to an enhanced stromal expression of SPARC in primary
tumors, as previously described (9).
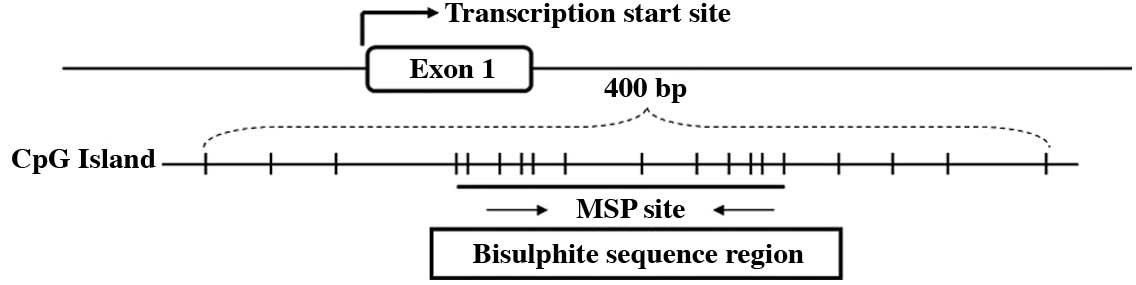
Bisulphite genomic sequence analysis of
CpG sites in the SPARC promoter region
Bisulphite genomic sequencing was used to determine
the detailed methylation status of the SPARC promoter region −29 to
+191 bp relative to the transcription start site of the SPARC gene
in 2 carcinoma cell lines, 3 laryngeal carcinoma cases and 1
hypopharyngeal carcinoma case with their matched normal epithelia.
The majority of the CpG sites were heavily methylated in cancer
cell lines and primary tumors, but almost all of the CpG sites
showed non-methylation in matched normal epithelia. Fig. 3A shows the typical CpG island
spanning the SPARC exon 1 and Fig.
3B shows representative results of the BGS analysis.
Restoration of SPARC expression following
pharmacological demethylation treatment
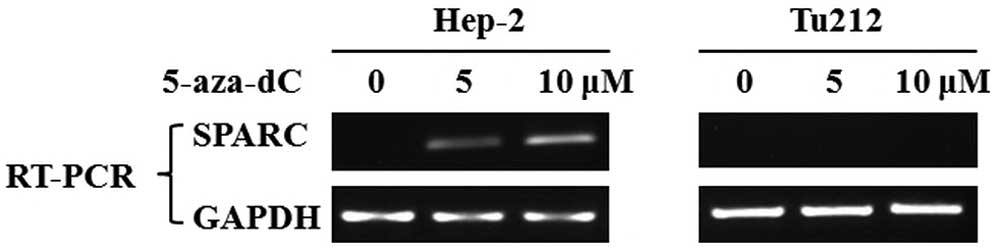
To determine whether SPARC is inactivated by
aberrant methylation, laryngeal and hypopharyngeal carcinoma cell
lines Hep-2 and Tu212 were treated with the methyltransferase
inhibitor 5-aza-dC and then examined using RT-PCR analysis. SPARC
expression was restored by pharmacological demethylation treatment
in the Hep-2 cell line by 5 or 10 μM 5-aza-dC, but not in the Tu212
cell line (Fig. 4).
Correlation between methylation status
and clinicopathological characteristics in laryngeal and
hypopharyngeal carcinoma
An analysis of the association of SPARC promoter
hypermethylation and clinicopathological characteristics of
laryngeal and hypopharyngeal carcinoma patients revealed no
significant correlation between aberrant methylation and age,
location or T stage. However, SPARC promoter hypermethylation was
found to be associated with lymph node metastasis (p<0.01)
(Table I).
 | Table ICorrelation between
clinicopathological characteristics and SPARC promoter
methylation. |
Table I
Correlation between
clinicopathological characteristics and SPARC promoter
methylation.
| No. | Promoter
methylation | p-valuea |
|---|
| Age |
| <60 | 29 | 51.7% (15/29) | 0.237 |
| ≥60 | 22 | 68.2% (15/22) | |
| Location |
| Larynx | 41 | 56.1% (23/41) | 0.423 |
| Hypopharynx | 10 | 70% (7/10) | |
| T stage |
| T1-2 | 16 | 50.0% (8/16) | 0.387 |
| T3-4 | 35 | 62.9% (22/35) | |
| N stage |
| N0 | 21 | 28.6% (6/21) | 0.000 |
| N1-2 | 30 | 80.0% (24/30) | |
Discussion
In addition to genetic changes, the transcriptional
inactivation of TSGs in human tumors by promoter hypermethylation
has been extensively studied. Methylation-associated silencing of
TSGs in HNSCC has been reported (17–19).
However, further identification of cancer-related genes is required
in order to gain a better understanding of the precise molecular
mechanism underlying the development of HNSCC.
As a prototypic matricellular protein, SPARC plays a
significant role in carcinogenesis, tumor progression, invasion and
metastasis by affecting the cell shape, differentiation,
attachment, migration, proliferation and growth factor activity.
The silencing of SPARC in various HNSCC cell lines, including those
of laryngeal and hypopharyngeal carcinoma, was first observed in
the present study (Fig. 1A). The
MSP assay also indicated that the CpG island of the SPARC promoter
was frequently methylated in these cell lines (Fig. 1B). The methylation status of the
SPARC promoter in primary tumor biopsies of laryngeal and
hypopharyngeal carcinoma was investigated. High frequencies of
SPARC promoter hypermethylation (58.8%, 30/51) were detected in
primary tumors, but were almost entirely absent from adjacent
normal epithelia. Notably, aberrant methylation was found even in
early stage (T1 or T2) cases (50.0%, 8/16). This finding may
reflect the fact that hypermethylation of SPARC is a frequent and
tumor-specific early event in laryngeal and hypopharyngeal
carcinoma. Patients lacking SPARC expression were associated with a
poorer prognosis in a survival analysis of colorectal carcinoma
(12). In our study, statistical
analysis showed that SPARC promoter hypermethylation was associated
with lymph node metastasis (p<0.01). This result suggests that
SPARC methylation serves as a biomolecular marker for the diagnosis
and prognosis of laryngeal and hypopharyngeal carcinoma.
As with the MSP assay, a subsequent BGS analysis
showed that the SPARC promoter was heavily methylated in
SPARC-silenced cancer cell lines (Fig.
3B). A heavy degree of methylation was observed in primary
tumor biopsies (Fig. 3B), despite
the inevitable normal tissue contamination in the biopsy samples
without micro-dissection. Furthermore, we showed that SPARC
expression was rescued by 5-aza-dC treatment (Fig. 4). Although demethylation treatment
failed to restore SPARC expression in the Tu212 cell line, our
results indicated that the epigenetic inactivation of SPARC by
aberrant promoter methylation is a significant, if not the only,
mechanism contributing to the loss of SPARC expression in laryngeal
and hypopharyngeal carcinoma.
The question of whether SPARC is a TSG or an
oncogene involved in human cancer remains unresolved, and the
precise role played by SPARC in human cancer has yet to be
clarified. An enhanced SPARC expression has been observed in oral
squamous cell carcinoma and other HNSCC (20,21).
On the other hand, the inactivation of SPARC by aberrant DNA
methylation has also been reported in pancreatic, ovarian, colon
and lung cancers (9,11,12,22).
With regards to SPARC function, exogenous SPARC inhibits the
proliferation, movement and migration of human cancer cell lines
(11,23), which is consistent with the results
of an in vivo study (14). A
recent study also provided evidence that exogenous SPARC enhances
the apoptosis of colon cancer cells via the caspase 8 pathway
(24).
In conclusion, SPARC is inactivated in laryngeal and
hypopharyngeal carcinoma by promoter hypermethylation, which
promotes the expression of these HNSCCs. Studies investigating the
in-depth mechanism of SPARC are required to determine whether its
promoter hypermethylation is likely to serve as a biomolecular
indicator for prognosis, as well as whether the selective rescue of
SPARC is a promising strategy for increasing the efficacy of cancer
therapy in laryngeal and hypopharyngeal carcinoma patients.
Acknowledgements
This study was supported by the Natural Science
Foundation of China (No. 30960416 to Z.Z.) and the Science and
Technology Department of Guangxi Province, China (No. 0728052 to
Z.Z). We are grateful to Dr Y. Chen (Department of Epidemiology and
Health Statistics, Public Health of Guangxi Medical University) for
her enthusiastic assistance in the statistical analysis. Dr H.
Zhu's (XiangYa Central Experiment Laboratory, Changsha, China)
provision of HNSCC cell lines is also acknowledged.
References
|
1
|
Ferlay J, Autier P, Boniol M, Heanue M,
Colombet M and Boyle P: Estimates of the cancer incidence and
mortality in Europe in 2006. Ann Oncol. 18:581–592. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics. CA Cancer J Clin.
58:71–96. 2008.
|
|
3
|
DeRienzo DP, Greenberg SD and Fraire AE:
Carcinoma of the larynx. Changing incidence in women. Arch
Otolaryngol Head Neck Surg. 117:681–684. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen YJ, Chang JT, Liao CT, Wang HM, Yen
TC, Chiu CC, Lu YC, Li HF and Cheng AJ: Head and neck cancer in the
betel quid chewing area: recent advances in molecular
carcinogenesis. Cancer Sci. 99:1507–1514. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rempel SA, Golembieski WA, Ge S, Lemke N,
Elisevich K, Mikkelsen T and Gutierrez JA: SPARC: a signal of
astrocytic neoplastic transformation and reactive response in human
primary and xenograft gliomas. J Neuropathol Exp Neurol.
57:1112–1121. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Seno T, Harada H, Kohno S, Teraoka M,
Inoue A and Ohnishi T: Downregulation of SPARC expression inhibits
cell migration and invasion in malignant gliomas. Int J Oncol.
34:707–715. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ledda F, Bravo AI, Adris S, Bover L,
Mordoh J and Podhajcer OL: The expression of the secreted protein
acidic and rich in cysteine (SPARC) is associated with the
neoplastic progression of human melanoma. J Invest Dermatol.
108:210–214. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Horie K, Tsuchihara M and Nakatsura T:
Silencing of secreted protein acidic and rich in cysteine inhibits
the growth of human melanoma cells with G arrest induction. Cancer
Sci. 101:913–919. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sato N, Fukushima N, Maehara N,
Matsubayashi H, Koopmann J, Su GH, Hruban RH and Goggins M:
SPARC/osteonectin is a frequent target for aberrant methylation in
pancreatic adenocarcinoma and a mediator of tumor-stromal
interactions. Oncogene. 22:5021–5030. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Di Martino JF, Lacayo NJ, Varadi M, Li L,
Saraiya C, Ravindranath Y, Yu R, Sikic BI, Raimondi SC and Dahl GV:
Low or absent SPARC expression in acute myeloid leukemia with MLL
rearrangements is associated with sensitivity to growth inhibition
by exogenous SPARC protein. Leukemia. 20:426–432. 2006.
|
|
11
|
Socha MJ, Said N, Dai Y, Kwong J,
Ramalingam P, Trieu V, Desai N, Mok SC and Motamed K: Aberrant
promoter methylation of SPARC in ovarian cancer. Neoplasia.
11:126–13512. 2009.PubMed/NCBI
|
|
12
|
Yang E, Kang HJ, Koh KH, Rhee H, Kim NK
and Kim H: Frequent inactivation of SPARC by promoter
hypermethylation in colon cancers. Int J Cancer. 121:567–575. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cheetham S, Tang MJ, Mesak F, Kennecke H,
Owen D and Tai IT: SPARC promoter hypermethylation in colorectal
cancers can be reversed by 5-Aza-2'deoxycytidine to increase SPARC
expression and improve therapy response. Br J Cancer. 98:1810–1819.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bull-Phelps SL, Carbon J, Miller A,
Castro-Rivera E, Arnold S, Brekken RA and Lea JS: Secreted protein
acidic and rich in cysteine as a regulator of murine ovarian cancer
growth and chemosensitivity. Am J Obstet Gynecol. 200:180–187.
2009.PubMed/NCBI
|
|
15
|
Zhang X, Chen ZG, Choe MS, Lin Y, Sun SY,
Wieand HS, Shin HJ, Chen A, Khuri FR and Shin DM: Tumor growth
inhibition by simultaneously blocking epidermal growth factor
receptor and cyclooxygenase-2 in a xenograft model. Clin Cancer
Res. 11:6261–6269. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Olek A, Oswald J and Walter J: A modified
and improved method for bisulphite based cytosine methylation
analysis. Nucleic Acids Res. 24:5064–5066. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Maruya S, Issa JP, Weber RS, Rosenthal DI,
Haviland JC, Lotan R and El-Naggar AK: Differential methylation
status of tumor-associated genes in head and neck squamous
carcinoma: incidence and potential implications. Clin Cancer Res.
10:3825–3830. 2004. View Article : Google Scholar
|
|
18
|
Bennett KL, Karpenko M, Lin MT, Claus R,
Arab K, Dyckhoff G, Plinkert P, Herpel E, Smiraglia D and Plass C:
Frequently methylated tumor suppressor genes in head and neck
squamous cell carcinoma. Cancer Res. 68:4494–4499. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bennett KL, Lee W, Lamarre E, Zhang X,
Seth R, Scharpf J, Hunt J and Eng C: HPV status-independent
association of alcohol and tobacco exposure or prior radiation
therapy with promoter methylation of FUSSEL18, EBF3, IRX1, and
SEPT9, but not SLC5A8, in head and neck squamous cell carcinomas.
Genes Chromosomes Cancer. 49:319–326. 2010.
|
|
20
|
Choi P, Jordan CD, Mendez E, Houck J, Yueh
B, Farwell DG, Futran N and Chen C: Examination of oral cancer
biomarkers by tissue microarray analysis. Arch Otolaryngol Head
Neck Surg. 134:539–546. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chin D, Boyle GM, Williams RM, Ferguson K,
Pandeya N, Pedley J, Campbell CM, Theile DR, Parsons PG and Coman
WB: Novel markers for poor prognosis in head and neck cancer. Int J
Cancer. 113:789–797. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Suzuki M, Hao C, Takahashi T, Shigematsu
H, Shivapurkar N, Sathyanarayana UG, Iizasa T, Fujisawa T,
Hiroshima K and Gazdar AF: Aberrant methylation of SPARC in human
lung cancers. Br J Cancer. 92:942–948. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen G, Tian X, Liu Z, Zhou S, Schmidt B,
Henne-Bruns D, Bachem M and Kornmann M: Inhibition of endogenous
SPARC enhances pancreatic cancer cell growth: modulation by
FGFR1-III isoform expression. Br J Cancer. 102:188–195. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang MJ and Tai IT: A novel interaction
between procaspase 8 and SPARC enhances apoptosis and potentiates
chemotherapy sensitivity in colorectal cancers. J Biol Chem.
282:34457–34467. 2007. View Article : Google Scholar
|