Introduction
Breast cancer is the leading cause of death from
cancer in America, and its incidence is currently increasing in
China. By 2021, it is anticipated to have increased substantially
from the current rate, estimated at 10–60 cases per 100,000 women,
to more than 100 new cases per 100,000 women aged 55–69 years
(1,2). Breast cancer has been suggested to be
caused by interactions between genetic and environmental factors.
Of these factors, genetic factors, such as family history and
gene-related variants, may play an important role in the
development and progression of breast cancer.
Signaling from transforming growth factor-β (TGF-β)
was conducted through serine-threonine kinases of transmembrane
receptors, including TGFBR1 and TGFBR2. TGF-β1 is the most abundant
form of TGF-β and regulates cellular processes by binding to
TGFBR2, which then activates TGFBR1 through phosphorylation
(3). Inactivation of the TGF-β
signaling pathway may lead to acquisition of resistance to the
anti-mitogenic effects of TGF-β and contribute to tumor development
and progression (4,5). Therefore, defective expression or
inactivation of TGF-β1 and its receptors, such as TGFBR2, may play
a significant role in carcinogenesis. Compelling evidence shows
that TGF-β signaling may play a crucial role in mammary development
(6,7) and a complex role in breast cancer
tumorigenesis via both tumor-suppressor and oncogenic activities
(8).
Although a number of polymorphisms in TGFB1
and its receptor TGFBR2 were originally reported in the
western population (9,10), no frequency of the polymorphisms
G-800A, codon25 and codon263 of TGFB1 was detected in the
Chinese population (11). Two
polymorphisms (C-509T and T+29C) in TGFB1 have been
associated with increased levels of TGF-β1 in serum from breast
cancer patients (12). The
TGFBR2 −875A allele was reported to enhance transcriptional
activity in normal epithelial cells (10). Based on previous findings, we
hypothesize that polymorphisms affecting the activity of genes
involved in the TGF-β signaling pathway play a role in the
development of breast cancer. However, little is known about the
relationship between TGFBR2 G-875A and breast cancer risk in
the Chinese population. Furthermore, the correlation of the
expression of estrogen receptor (ER), progesterone receptor (PR)
and human epidermal growth factor receptor-2 (HER2) in breast
cancer tissue with TGFB1 C-509T, T+29C and TGFBR2
G-875A polymorphisms remains to be determined. In this study, three
functional polymorphisms, TGFB1 C-509T, T+29C and
TGFBR2 G-875A, were selected to evaluate whether they modify
predisposition to breast cancer.
Materials and methods
Study participants
A total of 170 breast cancer patients and 178
cancer-free female controls were enrolled in this study. Fresh
surgically resected tissues (n=82) and archived paraffin-embedded
specimens (n=88) from patients diagnosed with breast cancer between
2006 and 2010 were selected from the Second Affiliated Hospital of
Soochow University, China. In addition, blood samples from 178
cancer-free donors, frequency-matched to the patients by age
(Table I), were collected for use
as controls at the same hospital. With respect to
immunohistochemistry, expression data on ER were available for 138,
PR for 138 and HER2 for 103 patients among all of the breast cancer
cases. Immunohistochemistry staining permitted the detection and
localization of ER, PR and HER2 within sections of
paraffin-embedded tissues. Positive expression is considered as
>20% of tumor cell nuclei staining, borderline is 5–19% and
negative is <5%. Both borderline and overtly positive results
were considered as positive (13,14).
HER2 expression was considered as positive (score 3+) and negative
(score 0–1+). Samples with a score of 2+ were excluded from the
analysis (15). This study was
approved by the Academic Advisory Board of Soochow University.
 | Table ICharacteristics of breast cancer
cases and controls. |
Table I
Characteristics of breast cancer
cases and controls.
| Cases | Controls | P-valuea |
|---|
| Age (mean ±
SD) | 53.75±11.22 | 52.67±10.28 | 0.35 |
| Tumor stage |
| 0 | 3 | | |
| I | 26 | | |
| II | 34 | | |
| III | 60 | | |
| IV | 0 | | |
| Missing | 47 | | |
| Cancer type |
| Intraductal
carcinoma | 5 | | |
| Infiltrative
ductal carcinoma | 158 | | |
| Infiltrative
lobular carcinoma | 1 | | |
| Mucinous
adenocarcinoma | 4 | | |
| Adenocarcinoma
infiltrating | 2 | | |
| Histologic
grade |
|
Well-differentiated | 5 | | |
| Moderately
differentiated | 115 | | |
| Poorly
differentiated | 12 | | |
| Missing | 26 | | |
| Tumor size |
| ≤2 cm | 38 | | |
| 2.1–5 cm | 81 | | |
| >5 cm | 4 | | |
| Missing | 47 | | |
| Lymph node
status |
| Positive | 61 | | |
| Negative | 63 | | |
| Not examined | 46 | | |
| Tumor
subtypesb |
|
ER+ | 88 | | |
|
ER− | 50 | | |
|
PR+ | 88 | | |
|
PR− | 50 | | |
| ER+ and
PR+ | 74 | | |
| ER+ or
PR+ | 28 | | |
| ER− and
PR− | 36 | | |
|
HER2+ | 9 | | |
|
HER2− | 94 | | |
DNA isolation and genotyping
Genomic DNA from fresh tissues and peripheral blood
samples was extracted using the proteinase K digestion standard
method. Isolation of DNA from paraffin-embedded tissues was
performed using a microwave-based DNA extraction method, as
previously described (16,17). Single nucleotide polymorphism (SNP)
analysis was performed using a polymerase chain
reaction-restriction fragment length polymorphism (PCR-RFLP) assay.
In total, two SNPs in TGFB1 and one SNP in TGFBR2
were genotyped. C-509T and G-875A were located in the promoter
region of TGFB1 and TGFBR2, respectively.
Additionally, T+29C was located in exon 1 of TGFB1. The
primer sequences, annealing temperatures, sizes of PCR products and
restriction enzymes are shown in Table
II. The PCR reaction was carried out in a total volume of 25
μl, containing 50–100 ng of genomic DNA, 1 unit of Ex Taq DNA
polymerase (Takara, Japan), 0.2 μmol/l of each primer, 1X Ex Taq
Buffer (Mg2+ Plus) and 0.25 mmol/l of each
deoxynucleotide triphosphate. Briefly, PCR amplification was
performed according to the following conditions: initial
denaturation at 95°C for 5 min followed by 35 cycles of 94°C for 45
sec, 63 or 59°C for 45 sec and 72°C for 45 sec. PCR was completed
by a final extension cycle at 72°C for 10 min.
| Table IIPrimers and restriction enzymes used
in PCR-RFLP assays for genotyping. |
Table II
Primers and restriction enzymes used
in PCR-RFLP assays for genotyping.
| Gene | Polymorphism | Primer sequence
5′→3′ | T (°C) | Size (bp) | Restriction
enzyme |
|---|
| TGFB1 | C-509T | F: TTG AGT GAC AGG
AGG CTG CTT A
R: GCT GGG AAA CAA GGT AGG AGA A | 63 | 178 | Eco81I |
| T+29C | F: CCA CCA CAC CAG
CCC TGT T
R: TCC GCT TCA CCA GCT CCA T | 63 | 186 | MspA1I |
| TGFBR2 | G-875A | F: GGA ATG TCT TGG
GCA AAT CT
R: ACC TGA ATG CTT GTG CTT TTA TT | 59 | 152 | TaaI |
Statistical analysis
The independent samples t-test was used to compare
the difference in age between breast cancer patients and the
controls. Differences in the distributions of genotypes and alleles
of TGFB1 and TGFBR2 variants between patients and
controls were evaluated using the χ2 test. The odds
ratios (ORs), their 95% confidence intervals (CIs) and the P-value
were assessed by logistic regression analyses, which were adjusted
for age. Hardy-Weinberg equilibrium (HWE) was tested by a
goodness-of-fit χ2 test. Statistical analysis was
performed using SPSS 16.0.
Results
Association of polymorphisms in TGFB1 and
TGFBR2 with risk of breast cancer
As summarized in Table
III, the genotype and allele frequencies were obtained for
TGFB1 C-509T, T+29C and TGFBR2 G-875A in breast
cancer cases and controls. The genotype distributions for the three
polymorphisms did not deviate from HWE in the patients (P=0.17,
0.22 and 1.00, respectively) or controls (P=0.55, 0.14 and 0.54,
respectively). No statistical difference was found between the
patients and controls for TGFB1 C-509T, T+29C genotype and
allele frequencies. Additionally, no overall association was found
between TGFBR2 G-875A genotype and breast cancer risk.
However, the frequency of the TGFBR2 -875A allele was
marginally higher in cancer-free individuals than that in breast
cancer patients (24.2 vs. 17.9%, P=0.05), indicating that
TGFBR2 −875A may predispose to breast cancer (adjusted
OR=0.69, 95% CI 0.48–0.99).
| Table IIIGenotype and allele distributions for
the polymorphisms of TGFB1 and TGFBR2 in breast
cancer patients and controls. |
Table III
Genotype and allele distributions for
the polymorphisms of TGFB1 and TGFBR2 in breast
cancer patients and controls.
| Gene | Genotype | Case | Control | OR (95% CI) | P-value |
|---|
| |
|
| | |
|---|
| | n | % | n | % | | |
|---|
| TGFB1 | C-509T | | | | | | |
| CC | 28 | 16.5 | 41 | 23.0 | 1 | |
| CT | 93 | 54.7 | 84 | 47.2 | 1.35
(0.73–2.50) | 0.35 |
| TT | 49 | 28.8 | 53 | 29.8 | 1.59
(0.91–2.81) | 0.11 |
| CT/TT | 142 | 83.5 | 137 | 77.0 | 1.50
(0.88–2.56) | 0.14 |
| C | 149 | 43.8 | 166 | 46.6 | 1 | |
| T | 191 | 56.2 | 190 | 53.4 | 1.12
(0.83–1.51) | 0.46 |
| T+29C | | | | | | |
| TT | 38 | 22.4 | 49 | 27.5 | 1 | |
| TC | 76 | 44.7 | 79 | 44.4 | 1.24
(0.73–2.10) | 0.43 |
| CC | 56 | 32.9 | 50 | 28.1 | 1.45
(0.82–2.57) | 0.20 |
| TC/CC | 132 | 77.6 | 129 | 72.5 | 1.32
(0.81–2.15) | 0.27 |
| T | 152 | 44.7 | 177 | 49.7 | 1 | |
| C | 188 | 55.3 | 179 | 50.3 | 1.22
(0.91–1.65) | 0.19 |
| TGFBR2 | G-875A | | | | | | |
| GG | 114 | 67.1 | 104 | 58.4 | 1 | |
| GA | 51 | 30.0 | 62 | 34.8 | 0.74
(0.47–1.17) | 0.20 |
| AA | 5 | 2.9 | 12 | 6.7 | 0.38
(0.13–1.12) | 0.08 |
| GA/AA | 56 | 32.9 | 74 | 41.5 | 0.68
(0.44–1.06) | 0.09 |
| G | 279 | 82.1 | 270 | 75.8 | 1 | |
| A | 61 | 17.9 | 86 | 24.2 | 0.69
(0.48–0.99) | 0.05 |
Relationship of TGFBR2 −875A allele to
breast cancer with the expression of ER, PR and HER2
Based on the effects of hormones on breast
carcinogenesis, the specimens were divided into various subgroups,
including ER−, ER+, PR−,
PR+, ER+PR+,
ER+PR−/ER−PR+ and
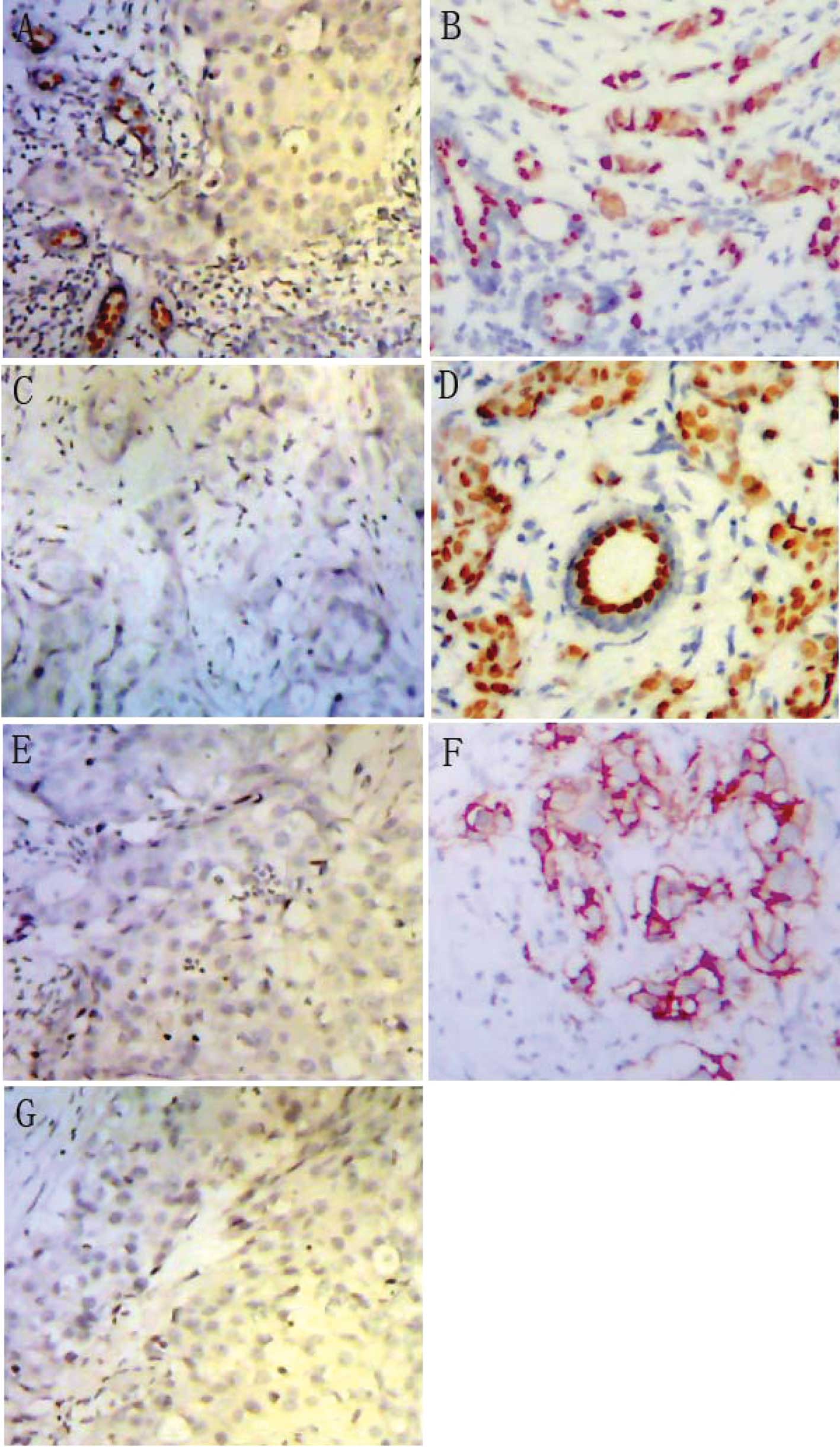
ER−PR− (Fig.
1A–D). The samples were additionally classified into subgroups
of HER2− and HER2+ (Fig. 1E and F). Information on ER, PR and
HER2 expression was available for 138, 138 and 103 patients with
breast cancer, respectively. In detail, 88 ER+, 50
ER−, 88 PR+, 50 PR−, 74
ER+PR+, 28
ER+PR−/ER−PR+, 36
ER−PR−, 94 HER2− and 9
HER2+ breast cancer cases were included (Table I). When performing stratification by
ER, PR and HER2 expression, we found no difference in the
frequencies of TGFB1 C-509T, T+29C and TGFBR2 G-875A
between ER+ and ER−, PR+ and
PR−, and HER2+ and HER2− breast
cancer cases (data available upon request). However, when comparing
the ER+, ER−, PR+, PR−,
ER+PR+,
ER+PR−/ER−PR+,
ER−PR−, HER2−, HER2+
breast cancer cases to the controls, our results showed that
TGFBR2 G-875A was associated with a decreased risk of breast
cancer with ER+ (OR=0.57, 95% CI 0.35–0.92),
PR+ (OR=0.54, 95% CI 0.34–0.88),
ER+PR+ (OR=0.55, 95% CI 0.33–0.92) and
HER2− (OR=0.55, 95% CI 0.34–0.88) under the dominant
genetic model (Table IV).
| Table IVAssociation of TGFBR2 G-875A
genotypes and alleles with ER, PR and HER2 expression. |
Table IV
Association of TGFBR2 G-875A
genotypes and alleles with ER, PR and HER2 expression.
| Genotype | Cases (%)/Controls
(%) | OR (95% CI) | P-value |
|---|
| ER+
breast cancer cases vs. controls |
| GG | 62 (70.5)/104
(58.4) | 1 | |
| GA | 25 (28.4)/62
(34.8) | 0.66
(0.37–1.16) | 0.15 |
| AA | 1 (1.1)/12
(6.7) | 0.14
(0.02–1.12) | 0.06 |
| GA+AA | 26 (29.5)/74
(41.6) | 0.58
(0.33–1.00) | 0.05 |
| G | 149 (84.7)/270
(75.8) | 1 | |
| A | 27 (15.3)/86
(24.2) | 0.57
(0.35–0.92) | 0.02 |
| PR+
breast cancer cases vs. controls |
| GG | 64 (72.7)/104
(58.4) | 1 | |
| GA | 22 (25)/62
(34.8) | 0.57
(0.32–1.01) | 0.06 |
| AA | 2 (2.3)/12
(6.7) | 0.27
(0.06–1.26) | 0.10 |
| GA+AA | 24 (27.3)/74
(41.5) | 0.52
(0.30–0.91) | 0.02 |
| G | 150 (85.2)/270
(75.8) | 1 | |
| A | 26 (14.8)/86
(24.2) | 0.54
(0.34–0.88) | 0.01 |
| ER+ and
PR+ breast cancer cases vs. controls |
| GG | 53 (71.6)/104
(58.4) | 1 | |
| GA | 20 (27)/62
(34.8) | 0.62
(0.34–1.14) | 0.13 |
| AA | 1 (1.4)/12
(6.7) | 0.17
(0.02–1.31) | 0.09 |
| GA+AA | 21 (28.4)/74
(41.5) | 0.55
(0.31–0.99) | 0.05 |
| G | 126 (85.1)/270
(75.8) | 1 | |
| A | 22 (14.9)/86
(24.2) | 0.55
(0.33–0.92) | 0.02 |
| HER2−
breast cancer cases vs. controls |
| GG | 67 (71.3)/104
(58.4) | 1 | |
| GA | 26 (27.7)/62
(34.8) | 0.62
(0.36–1.09) | 0.09 |
| AA | 1 (1.1)/12
(6.7) | 0.13
(0.02–1.01) | 0.05 |
| GA+AA | 27 (28.8)/74
(41.5) | 0.54
(0.32–0.93) | 0.03 |
| G | 160 (85.1)/270
(75.8) | 1 | |
| A | 28 (14.9)/86
(24.2) | 0.55
(0.34–0.88) | 0.01 |
Discussion
In this study, no difference was found in the
genotype and allele frequencies of TGFB1 C-509T and T+29C
between breast cancer patients and cancer-free female controls.
This finding is consistent with a meta-analysis involving 10,633
cases and 13,648 controls for the C-509T polymorphism and 20,022
cases and 24,423 controls for T+29C (18). A previous study concerning T+29C
showed decreased risk for Japanese pre-menopausal women with
CC genotype, but not for post-menopausal women (19). Kaklamani et al found that
breast cancer patients carrying the TGFB1*CC
allele of the polymorphism T+29C were more likely to have
ER− and PR− tumors (14). Cox et al found that an 18%
decreased risk of ER+/PR+ breast cancer
occurred among women heterozygous at C-509T and a 38% decrease
occurred among women homozygous for the T allele as compared to
those with CC genotype (20). In this study, breast cancer cases
were stratified with ER, PR and HER2 expression. No significant
association was found between the two polymorphisms (C-509T and
T+29C) and breast cancer risk in the Chinese population.
Additionally, previous studies indicated that the
TGFBR2 G-875A variant was not associated with breast cancer
risk in the European population (21). However, results of the present study
have shown that the −875A allele was marginally associated with a
decreased risk of breast cancer. These findings are consistent with
previous ones indicating that the TGFBR2 G-875A polymorphism
significantly correlates with a decreased risk of gastric and
esophageal squamous cell carcinomas in the Chinese population
(22,23). In particular, we found that the
TGFBR2 G-875A polymorphism may modify the risk of breast
cancer with ER+, PR+,
ER+PR+ and HER2−.
Activation of the TGF-β-mediated signal transduction
is subject to hormonal regulation (24). TGF-β signal transduction may play a
crucial role in the activation of various complex TGF-β receptors,
including TGFBR1 and TGFBR2 (25).
The expression of TGFBR2 is hormonally regulated and
anti-estrogen may induce activation of the TGFBR2 promoter
(26). For instance, breast cancer
cell lines expressing ER+ are refractory to TGF-β
effects, whereas ER− cells are sensitive (27). Loss or undetectable expression of
TGFBR2 has been reported to contribute to TGF-β resistance in
ER+ breast cancer cells (28). Paiva et al have suggested
that the absence of TGFBR2 was associated with poorer prognosis in
HER2− breast tumors (15). TGFBR2 −875A allele was proven
to enhance the transcriptional activity of TGFBR2 in normal
epithelial cells (10). In this
study, breast cancer subgroups, comprising ER+,
PR+, ER+PR+ and HER2−,
had a higher frequency of TGFBR2 −875G than the controls,
indicating that the TGFBR2 −875A allele correlated
significantly to a decreased risk of breast cancer with
ER+, PR+, ER+PR+ and
HER2−.
In conclusion, the present study suggests that the
functional polymorphism G-875A in TGFBR2, but neither of the
common genetic variants (C-509T and T-29C) in TGFB1, modify
the predisposition to breast cancer in Chinese females. Notably,
the TGFBR2 −875A allele is significantly associated with a
decreased risk of breast cancer with ER+, PR+
and HER2− expression.
Acknowledgements
The authors sincerely acknowledge the participation
and cooperation of the patients with NSCLC and the individuals with
no history of cancer. This study was supported by grants from the
Program for New Century Excellent Talents in University
(NCET-09-0165), the Science and Technology Committee of Jiangsu
Province (BK2008162), the SRF for ROCS, State Education Ministry
(2008890), the Qing-Lan Project of Education Bureau of Jiangsu
Province, the ‘333’ Project of Jiangsu Province Government, and the
Soochow Scholar Project of Soochow University (all to H.-T.
Zhang).
References
|
1
|
Ziegler RG, Anderson WF and Mh G:
Increasing breast cancer incidence in China: the numbers add up.
JNCI. 100:1339–1341. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Linos E, Rosner BA and Linos K: Effects of
reproductive and demographic changes on breast cancer incidence in
China: a modeling analysis. JNCI. 100:1352–1359. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Massague J and Chen YG: Controlling
TGF-beta signaling. Genes Dev. 14:627–644. 2000.
|
|
4
|
Gobbi H, Arteaga CL, Jensen RA, Simpson
JF, Dupont WD, Olson SJ, Schuyler PA, Plummer WD Jr and Page DL:
Loss of expression of transforming growth factor beta type II
receptor correlates with high tumour grade in human breast in situ
and invasive carcinomas. Histopathology. 36:168–177. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gobbi H, Dupont WD, Simpson JF, Plummer WD
Jr, Schuyler PA, Olson SJ, Arteaga CL and Page DL: Transforming
growth factor-beta and breast cancer risk in women with mammary
epithelial hyperplasia. J Natl Cancer Inst. 91:2096–2101. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Daniel CW and Robinson SD: Regulation of
mammary growth and function by TGF-beta. Mol Reprod Dev.
32:145–151. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barcellos-Hoff MH and Ewan KB:
Transforming growth factor-beta and breast cancer: mammary gland
development. Breast Cancer Res. 2:92–99. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
White RL: Tumor suppressing pathways.
Cell. 92:591–592. 1998. View Article : Google Scholar
|
|
9
|
Cambien F, Ricard S, Troesch A, Mallet C,
Generenaz L, Evans A, Arveiler D, Luc G, Ruidavets JB and Poirier
O: Polymorphisms of the transforming growth factor-beta 1 gene in
relation to myocardial infarction and blood pressure. The Etude
Cas-Temoin de l’Infarctus du Myocarde (ECTIM) Study. Hypertension.
28:881–887. 1996.PubMed/NCBI
|
|
10
|
Seijo ER, Song H, Lynch MA, Jennings R,
Qong X, Lazaridis E, Muro-Cacho C, Weghorst CM and Munoz-Antonia T:
Identification of genetic alterations in the TGFbeta type II
receptor gene promoter. Mutat Res. 483:19–26. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Y, Liu B, Jin M, Ni Q, Liang X, Ma
X, Yao K, Li Q and Chen K: Genetic polymorphisms of transforming
growth factor-beta1 and its receptors and colorectal cancer
susceptibility: a population-based case-control study in China.
Cancer Lett. 275:102–108. 2009. View Article : Google Scholar
|
|
12
|
Dunning AM, Ellis PD, McBride S, et al: A
transforming growth factorbeta1 signal peptide variant increases
secretion in vitro and is associated with increased incidence of
invasive breast cancer. Cancer Res. 63:2610–2615. 2003.
|
|
13
|
Onitilo AA, Engel JM, Greenlee RT and
Mukesh BN: Breast cancer subtypes based on ER/PR and Her2
expression: comparison of clinicopathologic features and survival.
Clin Med Res. 7:4–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kaklamani VG, Baddi L, Liu J, et al:
Combined genetic assessment of transforming growth factor-beta
signaling pathway variants may predict breast cancer risk. Cancer
Res. 65:3454–3461. 2005.PubMed/NCBI
|
|
15
|
Paiva CE, Drigo SA, Rosa FE, Moraes Neto
FA, Caldeira JR, Soares FA, Domingues MA and Rogatto SR: Absence of
transforming growth factor-beta type II receptor is associated with
poorer prognosis in HER2-negative breast tumours. Ann Oncol.
21:734–740. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Banerjee SK, Makdisi WF, Weston AP,
Mitchell SM and Campbell DR: Microwave-based DNA extraction from
paraffin-embedded tissue for PCR amplification. Biotechniques.
18:768–770. 772–763. 1995.PubMed/NCBI
|
|
17
|
Sato Y, Sugie R, Tsuchiya B, Kameya T,
Natori M and Mukai K: Comparison of the DNA extraction methods for
polymerase chain reaction amplification from formalin-fixed and
paraffin-embedded tissues. Diagn Mol Pathol. 10:265–271. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qi X, Zhang F, Yang X, Fan L, Zhang Y,
Chen L, Zhou Y, Chen X, Zhong L and Jiang J: Transforming growth
factor-beta1 polymorphisms and breast cancer risk: a meta-analysis
based on 27 case-control studies. Breast Cancer Res Treat.
122:273–279. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hishida A, Iwata H, Hamajima N, Matsuo K,
Mizutani M, Iwase T, Miura S, Emi N, Hirose K and Tajima K:
Transforming growth factor B1 T29C polymorphism and breast cancer
risk in Japanese women. Breast Cancer. 10:63–69. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cox DG, Penney K, Guo Q, Hankinson SE and
Hunter DJ: TGFB1 and TGFBR1 polymorphisms and breast cancer risk in
the Nurses’ Health Study. BMC Cancer. 7:1752007.PubMed/NCBI
|
|
21
|
Jin Q, Hemminki K, Grzybowska E, Klaes R,
Soderberg M, Zientek H, Rogozinska-Szczepka J, Utracka-Hutka B,
Pamula J, Pekala W and Forsti A: Polymorphisms and haplotype
structures in genes for transforming growth factor beta1 and its
receptors in familial and unselected breast cancers. Int J Cancer.
112:94–99. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jin G, Wang L, Chen W, Hu Z, Zhou Y, Tan
Y, Wang J, Hua Z, Ding W, Shen J, Zhang Z, Wang X, Xu Y and Shen H:
Variant alleles of TGFB1 and TGFBR2 are associated with a decreased
risk of gastric cancer in a Chinese population. Int J Cancer.
120:1330–1335. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jin G, Deng Y, Miao R, Hu Z, Zhou Y, Tan
Y, Wang J, Hua Z, Ding W, Wang L, Chen W, Shen J, Wang X, Xu Y and
Shen H: TGFB1 and TGFBR2 functional polymorphisms and risk of
esophageal squamous cell carcinoma: a case-control analysis in a
Chinese population. J Cancer Res Clin Oncol. 134:345–351. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Knabbe C, Lippman ME, Wakefield LM,
Flanders KC, Kasid A, Derynck R and Dickson RB: Evidence that
transforming growth factor-beta is a hormonally regulated negative
growth factor in human breast cancer cells. Cell. 48:417–428. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wrana JL, Attisano L, Wieser R, Ventura F
and Massague J: Mechanism of activation of the TGF-beta receptor.
Nature. 370:341–347. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Buck M, von der Fecht J and Knabbe C:
Antiestrogenic regulation of transforming growth factor beta
receptors I and II in human breast cancer cells. Ann NY Acad Sci.
963:140–143. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Arteaga CL, Tandon AK, von Hoff DD and
Osborne CK: Transforming growth factor beta: potential autocrine
growth inhibitor of estrogen receptor-negative human breast cancer
cells. Cancer Res. 48:3898–3904. 1988.
|
|
28
|
Kalkhoven E, Roelen BA, de Winter JP,
Mummery CL, van den Eijnden-van Raaij AJ, van der Saag PT and van
der Burg B: Resistance to transforming growth factor beta and
activin due to reduced receptor expression in human breast tumor
cell lines. Cell Growth Differ. 6:1151–1161. 1995.PubMed/NCBI
|