Introduction
The loss of cell-cell adhesion is a significant step
in tumor cell metastasis (1).
Claudins are a family of tight junction proteins that regulate cell
adhesion and polarity. Thus, they appear to be significant in
regulating the process of metastasis. First described in 1998,
claudins contain four transmembrane domains and, to date, 24 family
members have been identified. These proteins have molecular weights
of 20–27 kDa and are widely expressed in the majority of epithelial
cells (2,3). Certain claudin family members have
been identified to be abnormally regulated in several types of
cancer in humans. In particular, claudin-7 was observed to be
downregulated in breast cancer and head and neck squamous cell
carcinoma. These findings suggest that claudin-7 may be involved in
the epithelial-mesenchymal transition (EMT). Following the
restoration of claudin-7 levels, cancer cells have been
demonstrated to exhibit decreased motility and invasion abilities
(4,5).
Endometrial cancer is the primary gynecological
malignancy in the majority of countries (6). In the USA, ∼8,010 fatalities resulting
from endometrial cancer and 47,130 new cases are predicted this
year (6–8). At the time of diagnosis, ∼25% of
patients present with regional or distant metastases (stages III or
IV). Notably, the prognosis associated with this group is usually
unfavorable (9). Therefore,
specific strategies targeting tumor invasion should be a priority.
However, detailed mechanisms governing endometrial cancer
metastasis are not well defined. Previous studies have suggested
that claudins may play a critical role in this process.
In endometrial cancer, several of the claudins,
including claudin-1, -3 and -4, have previously been demonstrated
to be involved in maintaining tight junctions and in preventing
tumor cell dispersion and invasion (10,11).
In the present study, the functions of claudin-7 were investigated
in endometrial cancer.
Materials and methods
Immunohistochemical staining assay
Human endometrial cancer tissue microarrays,
comprising 31 pairs of endometrial cancer tissues and their
corresponding normal endometrial tissues, were purchased from
Shanghai Xinchao Biotechnology (Shanghai, China). All tissues had
been acquired via surgical resection. The cancer cases were
classified and graded according to the criteria of the
International Federation of Obstetrics and Gynecology (FIGO, 2009).
This study was approved by the ethics committee of the
International Peace Maternity Child Health Hospital, Shanghai
Jiaotong University. In brief, the tissue sample slides were
rehydrated and antigen retrieval was performed in the microwave
with ethylenediaminetetraacetic acid (EDTA; pH 8.0). The slides
were then incubated with primary anti-claudin-7 antibodies
(1:1,000; Epitomics, Inc., Burlingame, CA, USA) overnight at 4°C.
Antibody staining was visualized with 3,3′-diaminobenzidine (DAB;
Invitrogen Life Technologies, Carlsbad, CA, USA). For evaluation,
the following criteria were used: 0, no expression (complete
negative staining); 1, weak expression (1–15% positive staining);
2, moderate expression (16–49% positive staining); and 3, strong
expression (50–100% positive staining). Scores of 0 and 1 were
defined as negative expression. All slides were independently
scored by two pathologists.
Cancer cell lines and cultures
The RL95-2, Ishikawa, AN3CA and KLE endometrial
cancer cell lines, were routinely cultured in Dulbecco’s modified
Eagle’s medium (DMEM)/F12 (Gibco, Auckland, New Zealand)
supplemented with 10% fetal bovine serum (Biowest, Nuaillé, France)
at 37°C with 5% CO2.
RNA extraction and quantitative real-time
reverse transcription polymerase chain reaction (RT-PCR)
Total RNA was extracted using TRIzol reagent
(Invitrogen Life Technologies) and reverse transcribed with the RT
kit from Takara Biotechnology Co., Ltd. (Dalian, China). The
primers used for claudin-7 were as follows: Forward, 5′-AGAG CACG G
G GATGATGAG-3′ a nd reverse, 5′-CACCCATGGCTATACGGGC-3′. The PCR
conditions were as follows: 95°C for 30 sec, 35 cycles at 95°C for
5 sec, then 60°C for 30 sec. β-actin was used as an endogenous
control. The relative mRNA expression was calculated using the
2−ΔΔCt comparative CT method.
Western blot analysis
The primary antibodies for claudin-7 were obtained
from Epitomics, Inc. The mouse monoclonal
anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH), horseradish
peroxidase (HRP)-conjugated anti-rabbit and anti-mouse secondary
antibodies were purchased from Boshide Biotechnology Company
(Wuhan, China). The specific bands were developed with enhanced
chemiluminescence (Beyotime, Shanghai, China).
Silencing claudin-7 in Ishikawa
cells
Claudin-7-specific siRNA was purchased from Shanghai
Genepharma Co., Ltd. (China). The cells were cultured in 6-well
plates for 24 h prior to being transfected with Lipofectamine 2000
(Invitrogen Life Technologies), according to the manufacturer’s
instructions.
Overexpressing claudin-7 in AN3CA
cells
The plasmid pcDNA3.1-claudin-7 was purchased from
Genearray Biotech Company (Shanghai, China) and verified by
sequence analysis. The AN3CA cells were transfected with
pcDNA3.1-claudin-7 or an empty vector using Lipofectamine 2000.
Subsequently, real-time PCR and western blot analysis were
performed to verify the changes in claudin-7 expression.
Proliferation assay
The proliferation of the Ishikawa and AN3CA cells
was determined by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay. Briefly, 4×103 cells were seeded in each well of
a 96-well plate and incubated overnight. At the appropriate time
(48 and 72 h), the cells were incubated with 10 μl MTT (5
mg/ml; Sigma, St. Louis, MO, USA) for 4 h. Formazan crystals were
subsequently dissolved in 100 μl dimethylsulfoxide (DMSO;
Sigma). The absorbance of the solution was measured at 570 nm. All
measurements were repeated in triplicate.
Transwell assay
The transwell chamber system (Applied Biosystems,
Foster City, CA, USA) was used to investigate cellular invasive
abilities. Briefly, 1×104 Ishikawa or AN3CA cells were
seeded into the upper chamber. Subsequently, 500 μl
conditioned medium was added to the bottom chamber. Following 24 h
of incubation, the cells that had migrated to the bottom membrane
were stained and counted under a microscope. This experiment was
performed in triplicate.
Statistical analysis
P<0.05 was considered to indicate a statistically
significant difference. SPSS software, version 16.0, (SPSS Inc.,
Chicago, IL, USA) was used for the statistical analysis. The
analysis was performed using either a χ2 test or a
t-test.
Results
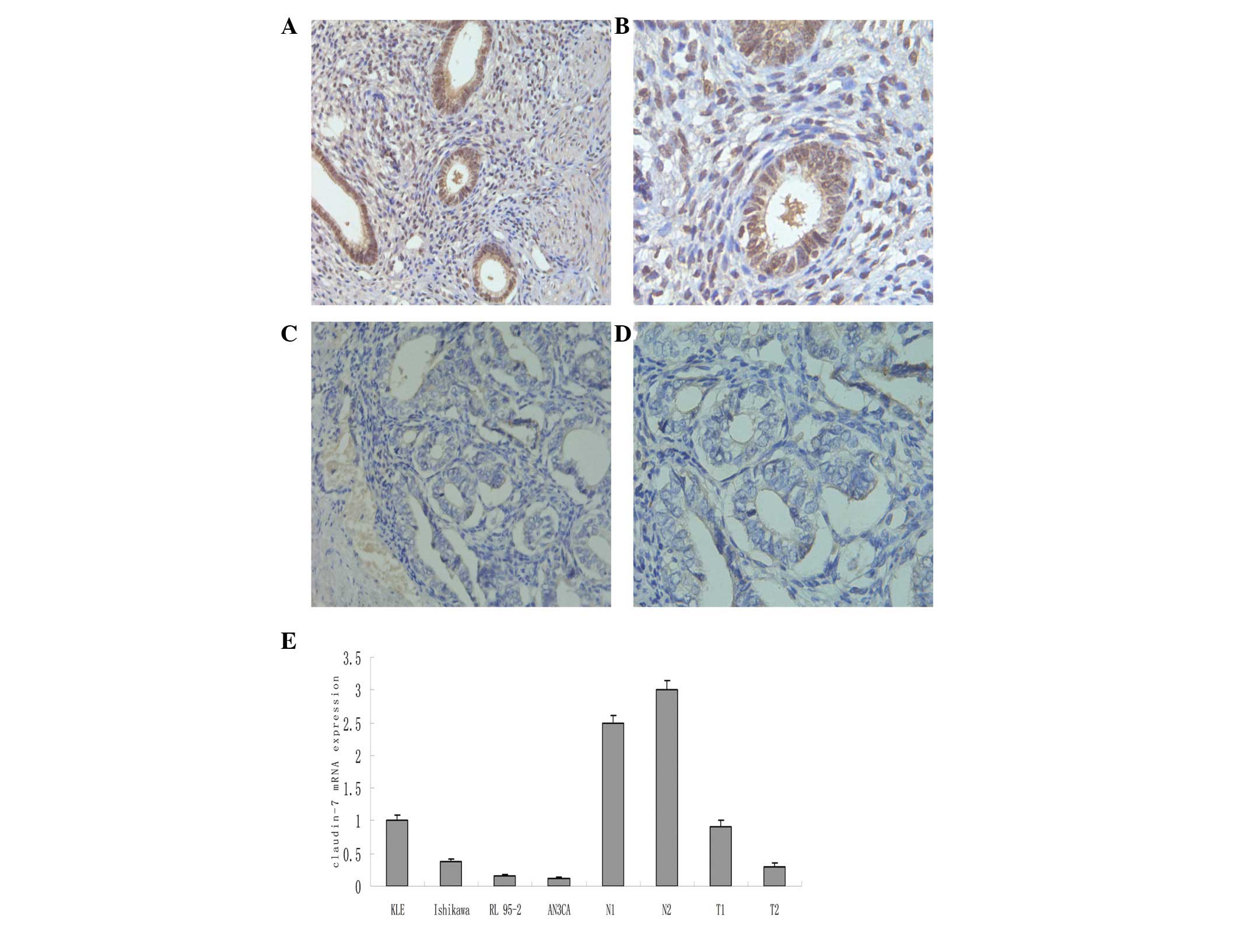
Claudin-7 expression in human endometrial
cancer
Compared with the corresponding normal endometrial
tissues, the endometrial cancer tissues exhibited significantly
downregulated claudin-7 expression (Fig. 1, P<0.01). In the cancer tissues,
approximately half of the patients (14/31, 45.2%) demonstrated a
loss or the total negative expression of claudin-7. A real-time
RT-PCR analysis of the KLE, RL 95-2 and Ishikawa cells revealed
that these cell lines positively expressed claudin-7, and that by
contrast, the AN3CA cells were claudin-7-negative (Fig. 1).
Correlations between claudin-7 expression
and clinicopathological characteristics
According to the statistical analysis, the reduced
expression of claudin-7 was significantly correlated with the tumor
stage (P=0.023) and histological grade (P=0.018), but not with the
remaining characteristics that were examined (Table I).
 | Table I.Correlations between claudin-7
expression and clinicopathological characteristics in 31
endometrial carcinoma samples. |
Table I.
Correlations between claudin-7
expression and clinicopathological characteristics in 31
endometrial carcinoma samples.
| Variables | No. of patients | Claudin-7 expression
| P-valuea |
|---|
| Negative | Positive |
|---|
| Age (years) | | | | |
| <60 | 17 | 7 | 10 | 0.715 |
| ≥60 | 14 | 7 | 7 | |
| Stage | | | | |
| I | 21 | 9 | 12 | 0.023 |
| II | 5 | 2 | 3 | |
| III | 4 | 2 | 2 | |
| IV | 1 | 1 | 0 | |
| Grade | | | | |
| 1 | 23 | 8 | 15 | 0.018 |
| 2 | 5 | 3 | 2 | |
| 3 | 3 | 3 | 0 | |
| Myometrial
invasion | | | | |
| <1/2 | 19 | 9 | 10 | 0.560 |
| ≥1/2 | 12 | 5 | 7 | |
| ER expression | | | | |
| Negative | 9 | 5 | 4 | 0.125 |
| Positive | 22 | 10 | 12 | |
| PR expression | | | | |
| Negative | 5 | 2 | 3 | 0.318 |
| Positive | 26 | 12 | 14 | |
| p53 expression | | | | |
| Negative | 23 | 10 | 13 | 0.266 |
| Positive | 8 | 3 | 5 | |
Correlations between claudin-7 expression
and other immunohistochemical parameters
No correlation was observed between claudin-7
expression and ER, PR or p53 expression (P=0.125, P=0.318 and
P=0.266, respectively; Table
I).
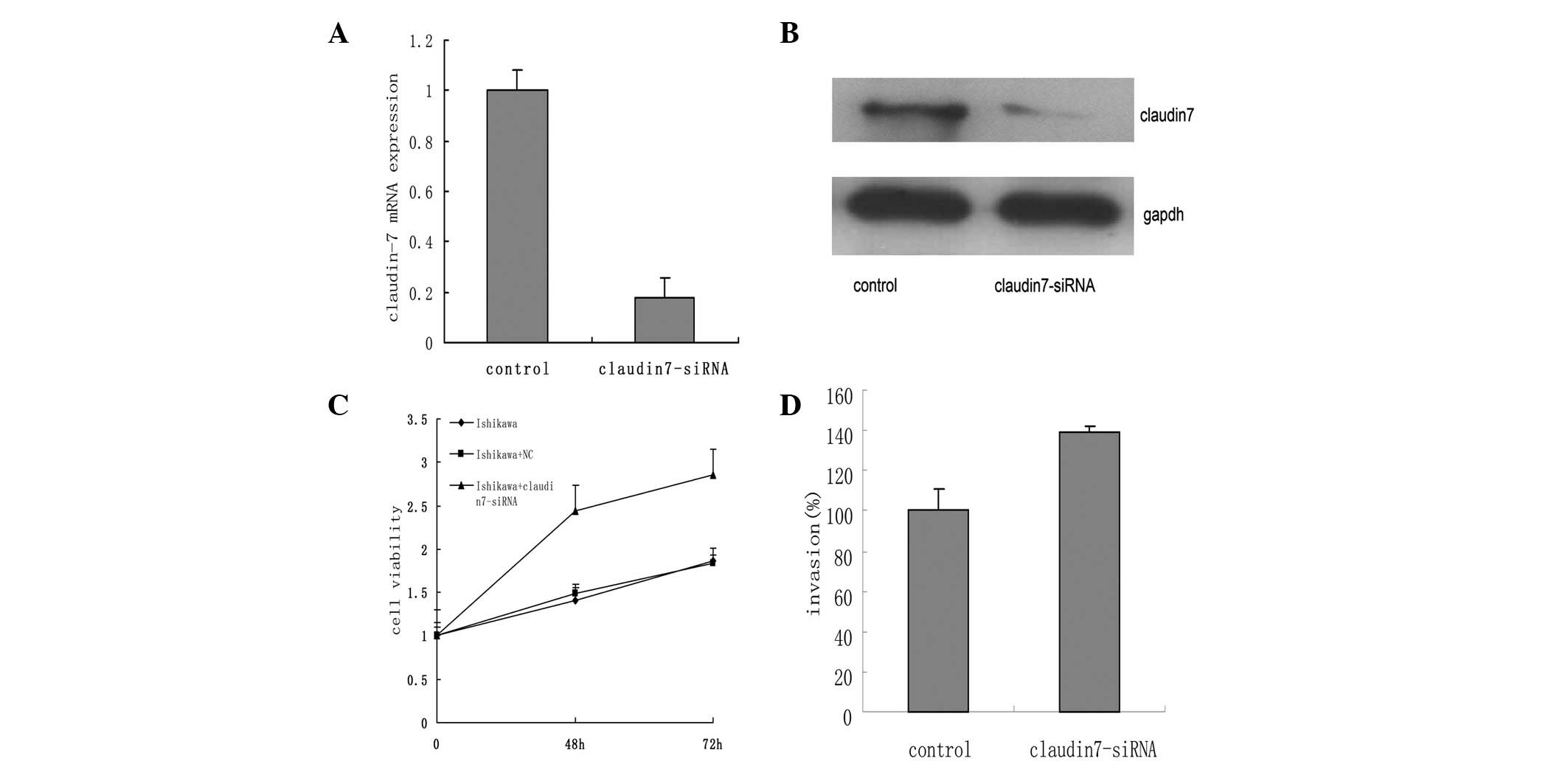
Claudin-7 silencing increases the
proliferation and invasion of Ishikawa cells
RNA interference resulted in an ∼90% knockdown of
claudin-7 mRNA levels in the Ishikawa cells (Fig. 2A and B). The MTT assay revealed that
the Ishikawa cells treated with claudin-7-siRNA possessed a higher
growth rate compared with the untreated and empty vector (negative
control) Ishikawa cells (P=0.032; Fig.
2C). In the transwell assay, the claudin-7-silenced Ishikawa
cells were more invasive than the control group (P=0.02; Fig. 2D).
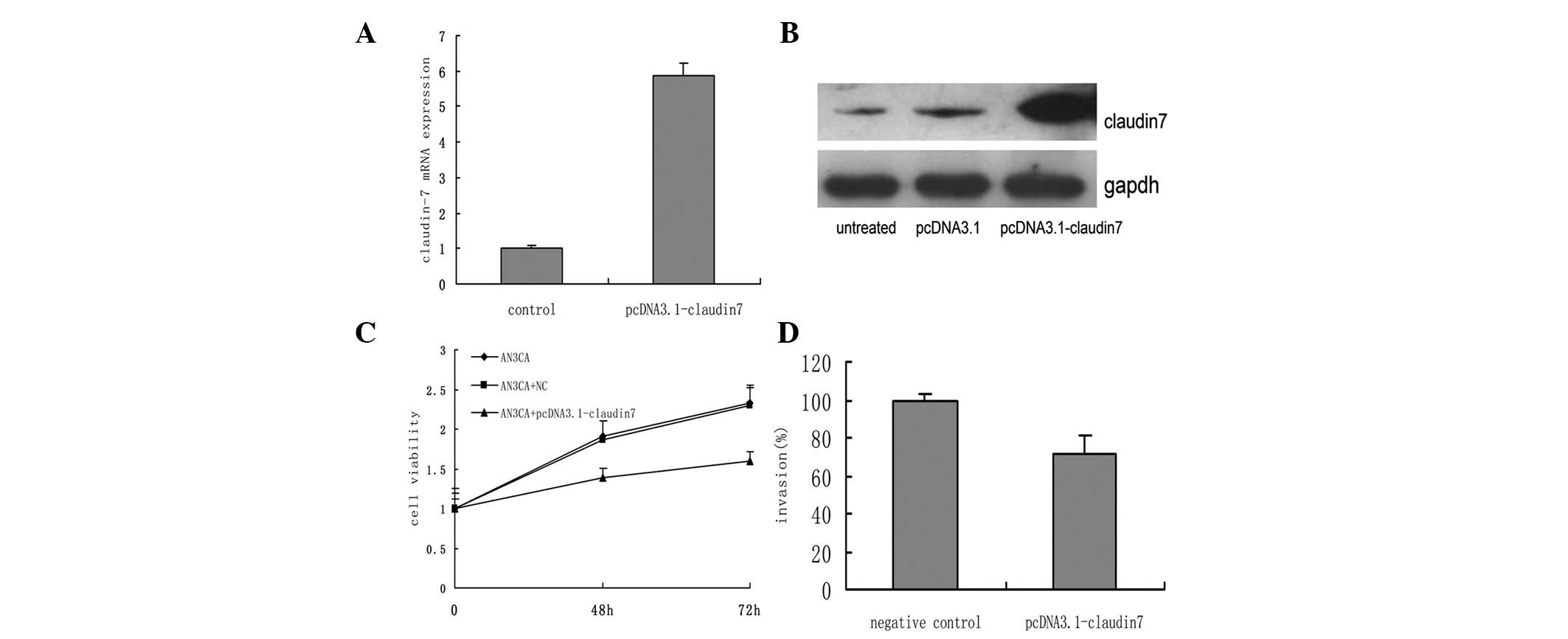
Claudin-7 overexpression decreases the
tumorigenic properties of AN3CA cells
The transfection of the AN3CA cells with
pcDNA3.1-claudin-7 resulted in the upregulation of claudin-7 at the
mRNA and protein levels (Fig. 3A and
B). Cellular proliferation and invasion were significantly
suppressed following transfection (P=0.021 and P=0.012, Fig. 3C and D, respectively), compared with
the untreated and empty vector (negative control) AN3CA cells.
Discussion
In numerous types of human cancer, the number of
cell-cell junctions decreases, permitting the escape of cancer
cells from their primary sites, along with the acquisition of
invasive and metastatic properties (12,13).
Therefore, targeting cell-cell junctions may be a valid strategy
for the treatment of cancer. A number of claudins, including
claudin-7, have been identified to be downregulated in various
types of human cancer. Claudin-7 has been shown to be downregulated
in head and neck squamous cell carcinoma, invasive ductal breast
cancer and invasive esophageal cancer (4,5,14).
However, these studies did not analyze the manner in which
claudin-7 functions to inhibit cancer cells from undergoing
invasion and metastasis. Thus, the present study investigated the
expression and potential mechanisms governing claudin-7 in
endometrial cancer.
The study observed that claudin-7 was frequently
down-regulated in human endometrial cancer tissues. To investigate
the mechanism by which claudin-7 affects endometrial cancer cells,
claudin-7-specific siRNA was first transfected into Ishikawa cells
(those demonstrating normal claudin-7 expression). The results
demonstrated that invasion was significantly upregulated in the
Ishikawa cells. Cellular proliferation was also significantly
upregulated. Full-length claudin-7 cDNA was then cloned and
overexpressed in the AN3CA cells (which were claudin-7-negative),
and MTT and invasion assays confirmed similar effects.
It has been suggested that the effects of claudin-7
were generated through the attenuated activation of MAPK/ERK
signaling (14). The interactions
between the MAPK pathway and tight junction proteins have
previously been identified. For example, the tight junction
membrane protein, occludin, participates in the activation of the
MAPK signaling pathway (15). In
the hepatic cell lines derived from occludin-deficient mice, MAPK
activation was revealed to be downregulated, triggering apoptosis
and increasing claudin-2 expression. The PI3K/Akt pathway may also
interact with tight junction proteins. It has been also
demonstrated, in these hepatic cell lines, that the activation of
Akt is decreased, while cell apoptosis is increased (16).
Overall, the present study demonstrated that
claudin-7 is frequently downregulated in endometrial cancer, and
that this is correlated with the tumor stage and histological
grade. The ectopic expression of claudin-7 significantly regulates
the proliferation and invasion of endometrial cancer cells.
Therefore, these findings may provide a potential therapeutic
target for the treatment of endometrial cancer.
Acknowledgements
This study was supported by the
National Natural Science Foundation of China (grant no. 81172477),
the Youth Foundation of Shanghai Hygiene Bureau (grant no. 044Y06)
and the Natural Science Foundation of Shanghai Science and
Technology (grant no. 11ZR1440800).
References
|
1.
|
Hirohashi and Kanai Y: Cell adhesion
system and human cancer morphogenesis. Cancer Sci. 94:575–581.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
González-Mariscal L, Betanzos A, Nava P
and Jaramillo BE: Tight junction proteins. Prog Biophys Mol Biol.
81:1–44. 2003.
|
|
3.
|
Morita K, Furuse M, Fujimoto K and Tsukita
S: Claudin multigene family encoding four-transmembrane domain
protein components of tight junction strands. Proc Natl Acad Sci
USA. 96:511–516. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Kominsky SL, Argani P, Korz D, Evron E,
Raman V, Garrett E, Rein A, Sauter G, et al: Loss of the tight
junction protein claudin-7 correlates with histological grade in
both ductal carcinoma in situ and invasive ductal carcinoma of the
breast. Oncogene. 22:2021–2033. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Al Moustafa AE, Alaoui-Jamali MA, Batist
G, Hernandez-Perez M, Serruya C, Alpert L, Black MJ, Sladek R and
Foulkes WD: Identification of genes associated with head and neck
carcinogenesis by cDNA microarray comparison between matched
primary normal epithelial and squamous carcinoma cells. Oncogene.
21:2634–2640. 2002.
|
|
6.
|
Amant F, Moerman P, Neven P, Timmerman D,
Van Limbergen E and Vergote I: Endometrial cancer. Lancet.
366:491–505. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
American Cancer Society: Cancer Facts
& Figures 2011. American Cancer Society; Atlanta: 2011
|
|
8.
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar
|
|
9.
|
Dedes KJ, Wetterskog D, Ashworth A, Kaye
SB and Reis-Filho JS: Emerging therapeutic targets in endometrial
cancer. Nat Rev Clin Oncol. 8:261–271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Pan XY, Wang B, Che YC, Weng ZP, Dai HY
and Peng W: Expression of claudin-3 and claudin-4 in normal,
hyperplastic, and malignant endometrial tissue. Int J Gynecol
Cancer. 17:233–241. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Santin AD, Zhan F, Cane’ S, Bellone S,
Palmieri M, Thomas M, et al: Gene expression fingerprint of uterine
serous papillary carcinoma: identification of novel molecular
markers for uterine serous cancer diagnosis and therapy. Br J
Cancer. 92:1561–1573. 2005. View Article : Google Scholar
|
|
12.
|
Moldvay J, Jäckel M, Páska C, Soltész I,
Schaff Z and Kiss A: Distinct claudin expression profile in
histologic subtypes of lung cancer. Lung Cancer. 57:159–167. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Swisshelm K, Macek R and Kubbies M: Role
of claudins in tumorigenesis. Adv Drug Deliv Rev. 57:919–928. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Lioni M, Brafford P, Andl C, Rustgi A,
El-Deiry W, Herlyn M and Smalley KS: Dysregulation of claudin-7
leads to loss of E-cadherin expression and the increased invasion
of esophageal squamous cell carcinoma cells. Am J Pathol.
170:709–721. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Lu Z, Ding L, Hong H, Hoggard J, Lu Q and
Chen YH: Claudin-7 inhibits human lung cancer cell migration and
invasion through ERK/MAPK signaling pathway. Exp Cell Res.
317:1935–1946. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Murata M, Kojima T, Yamamoto T, Go M,
Takano K, Osanai M, Chiba H and Sawada N: Down-regulation of
survival signaling through MAPK and Akt in occludin-deficient mouse
hepatocytes in vitro. Exp Cell Res. 310:140–151. 2005. View Article : Google Scholar : PubMed/NCBI
|